
Stress-Activated Protein Kinase Signalling Regulates Mycoparasitic Hyphal-Hyphal Interactions in Trichoderma atroviride

 , and
, and 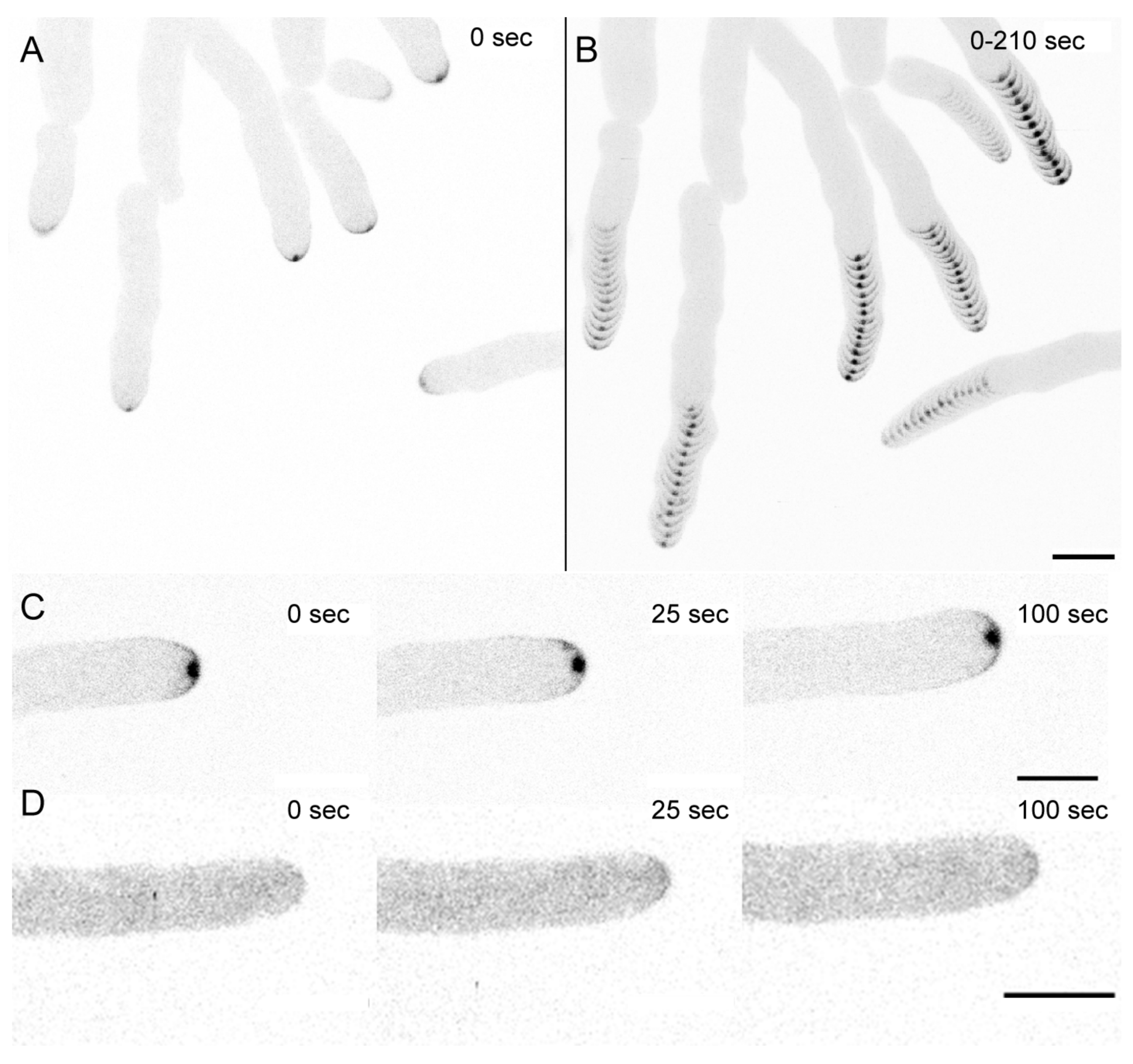{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains
2.2. Dual-Culture Confrontation Assays
2.3. Cloning of CRIB GTPase-Activity Reporters
2.4. T. atroviride Transformation
2.5. Confocal Laser Scanning Microscopy
2.6. Wide-Field Fluorescence Microscopy
2.7. Live-Cell Imaging Experimentation
2.8. Pharmacologial Inhibition of GTPase Activity
2.9. Image Post-Processing and Automated Time Course Analysis
2.10. Manual Image Data Quantification and Statistical Analyses
3. Results
3.1. Dynamic Recruitment of the CRIB Reporter Reflects Apical GTPase Activity in T. atroviride Hyphae
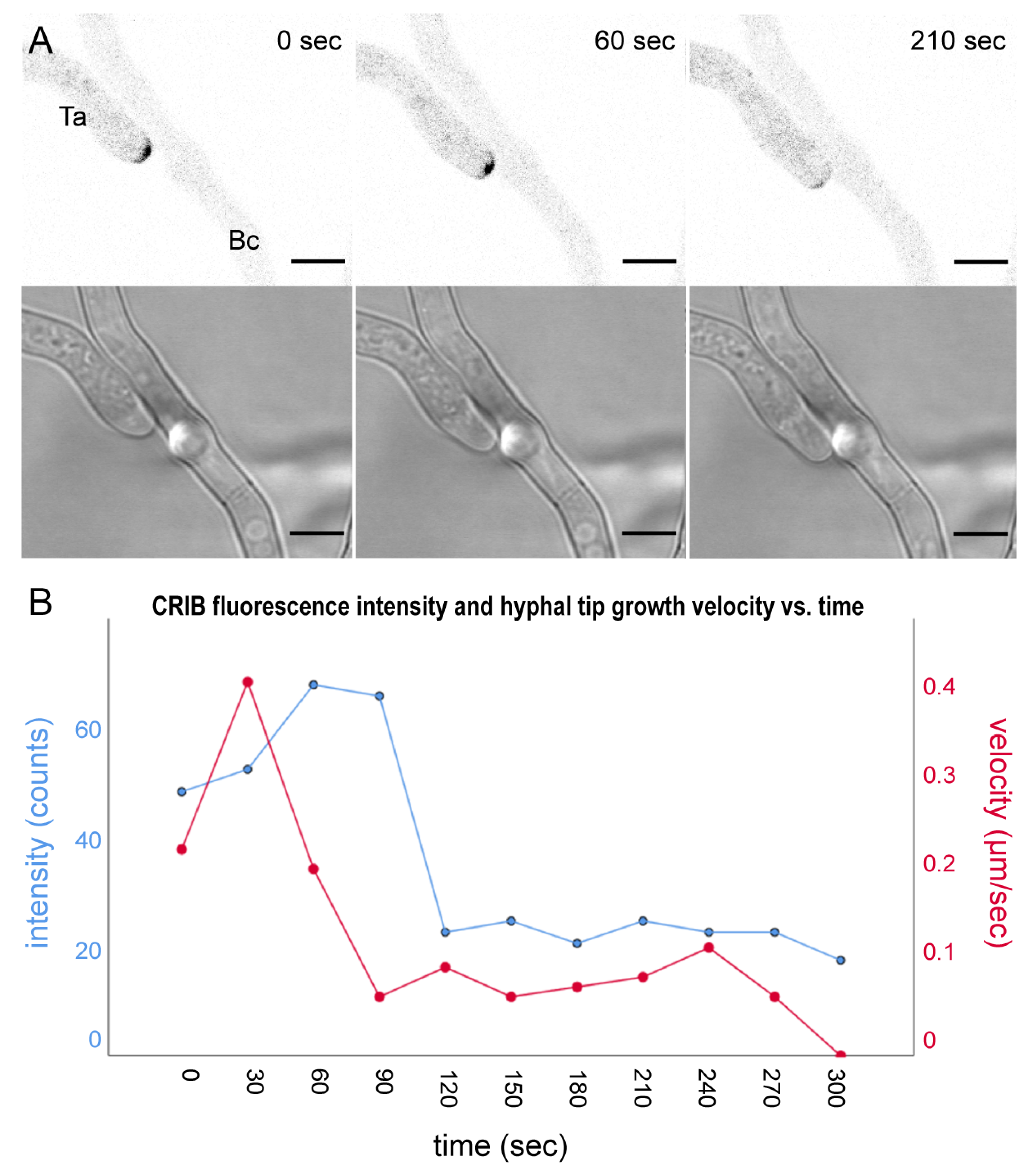
3.2. The Onset of Prey-Induced Tip Depolarisation during the Pre-Contact Phase Occurs Abruptly and Results in Growth Arrest
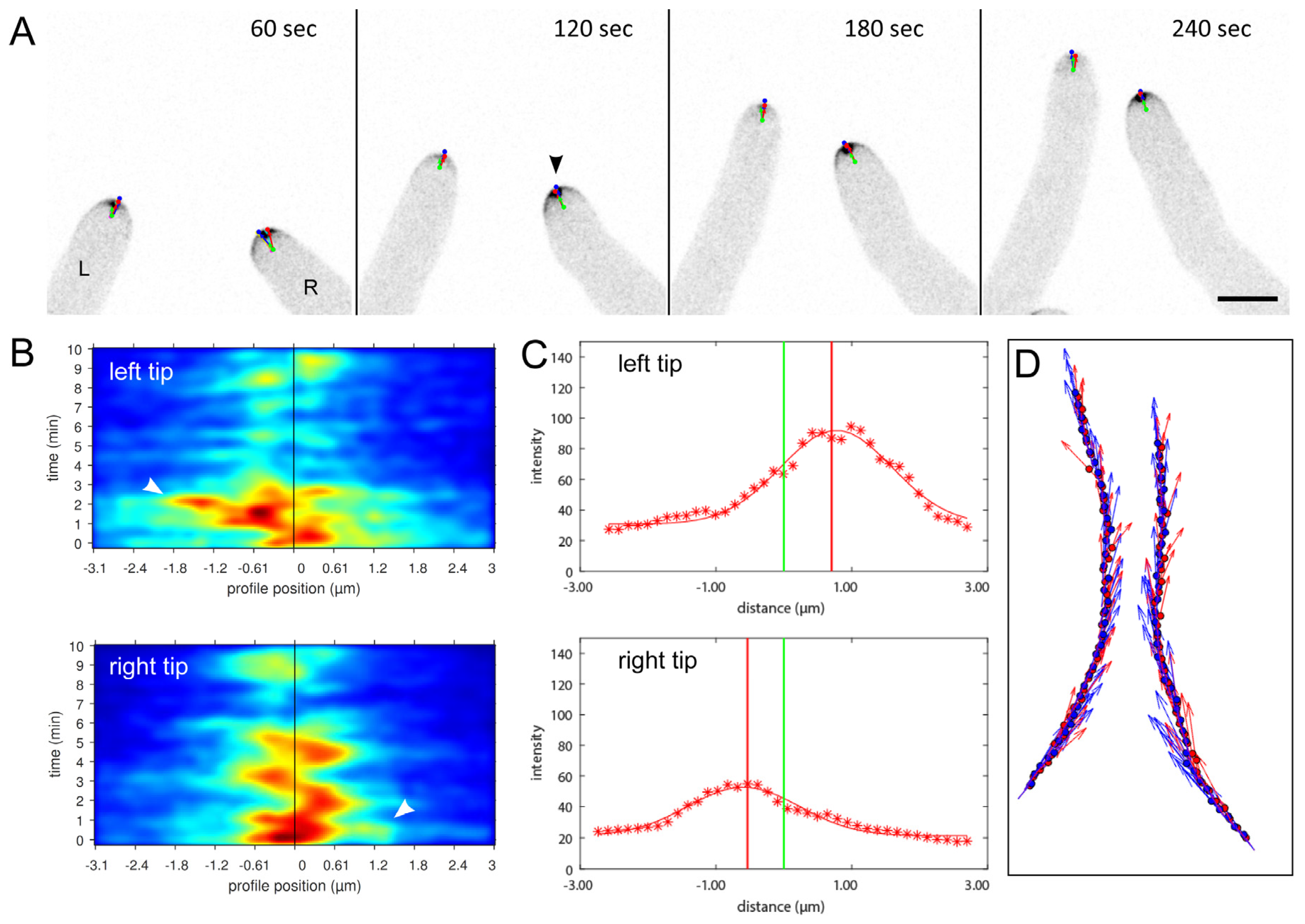
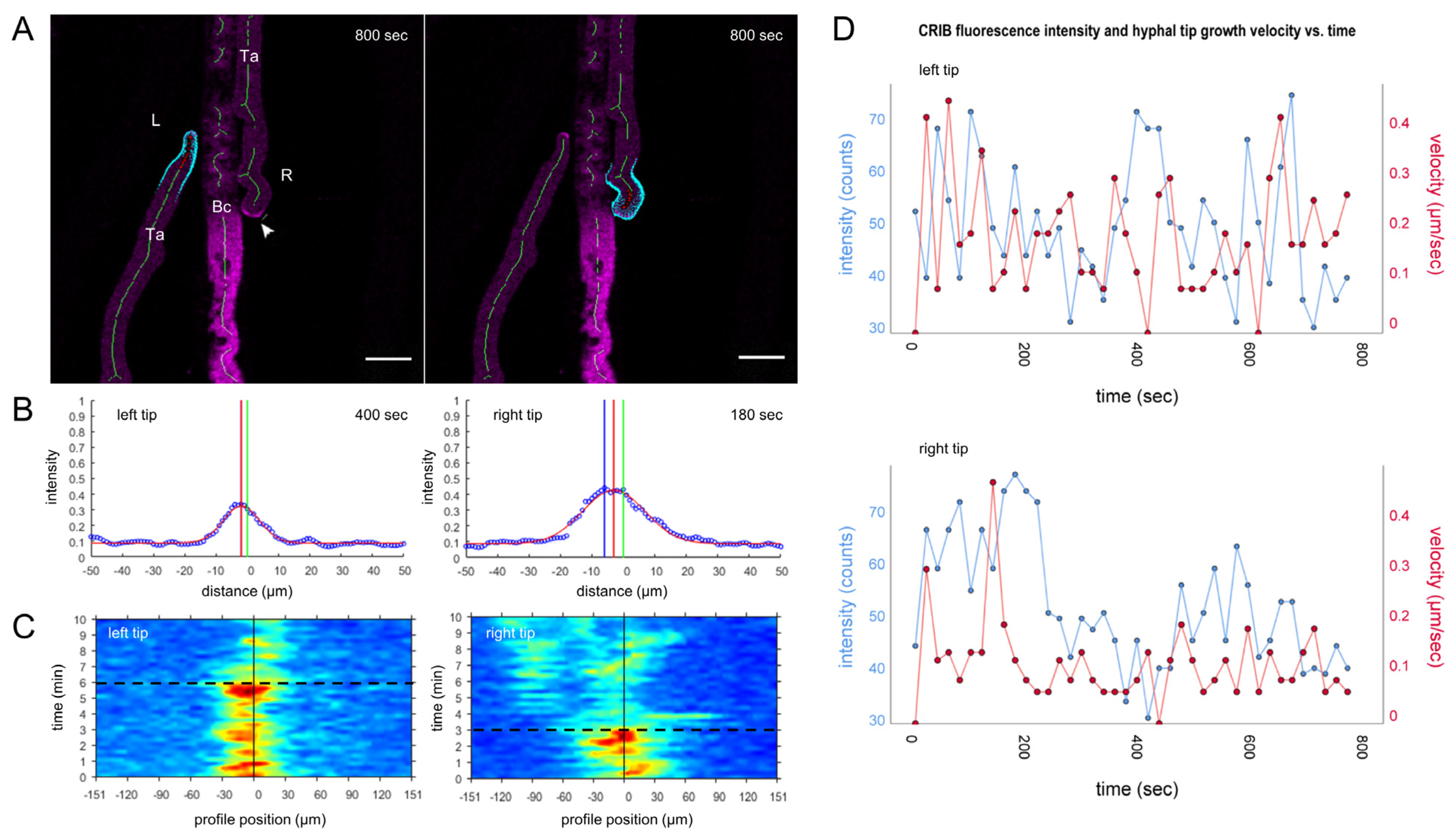
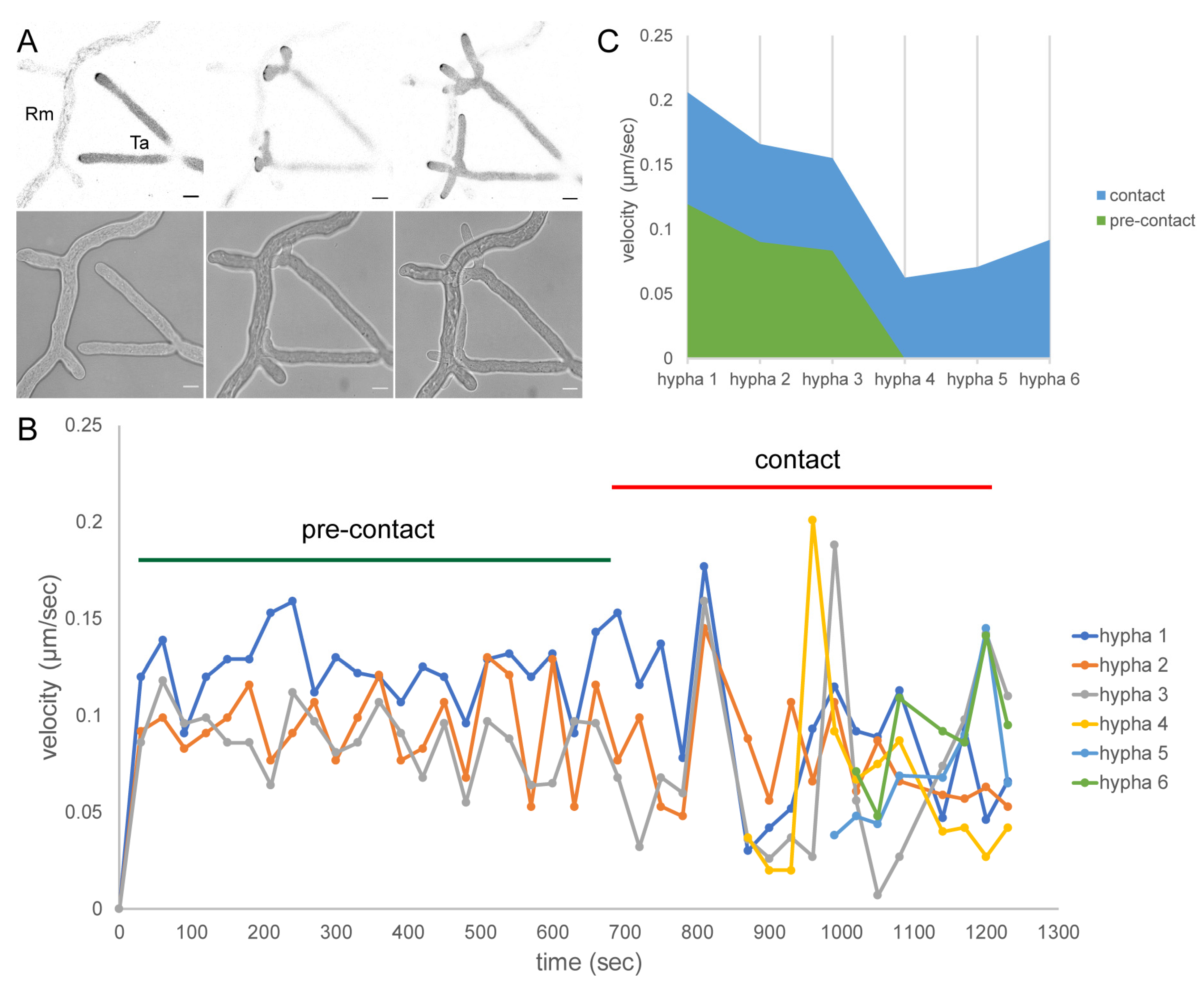
3.3. Mycoparasitic Hyphae Switch Chemotropic Behaviour While Establishing Contact to Prey Hyphae
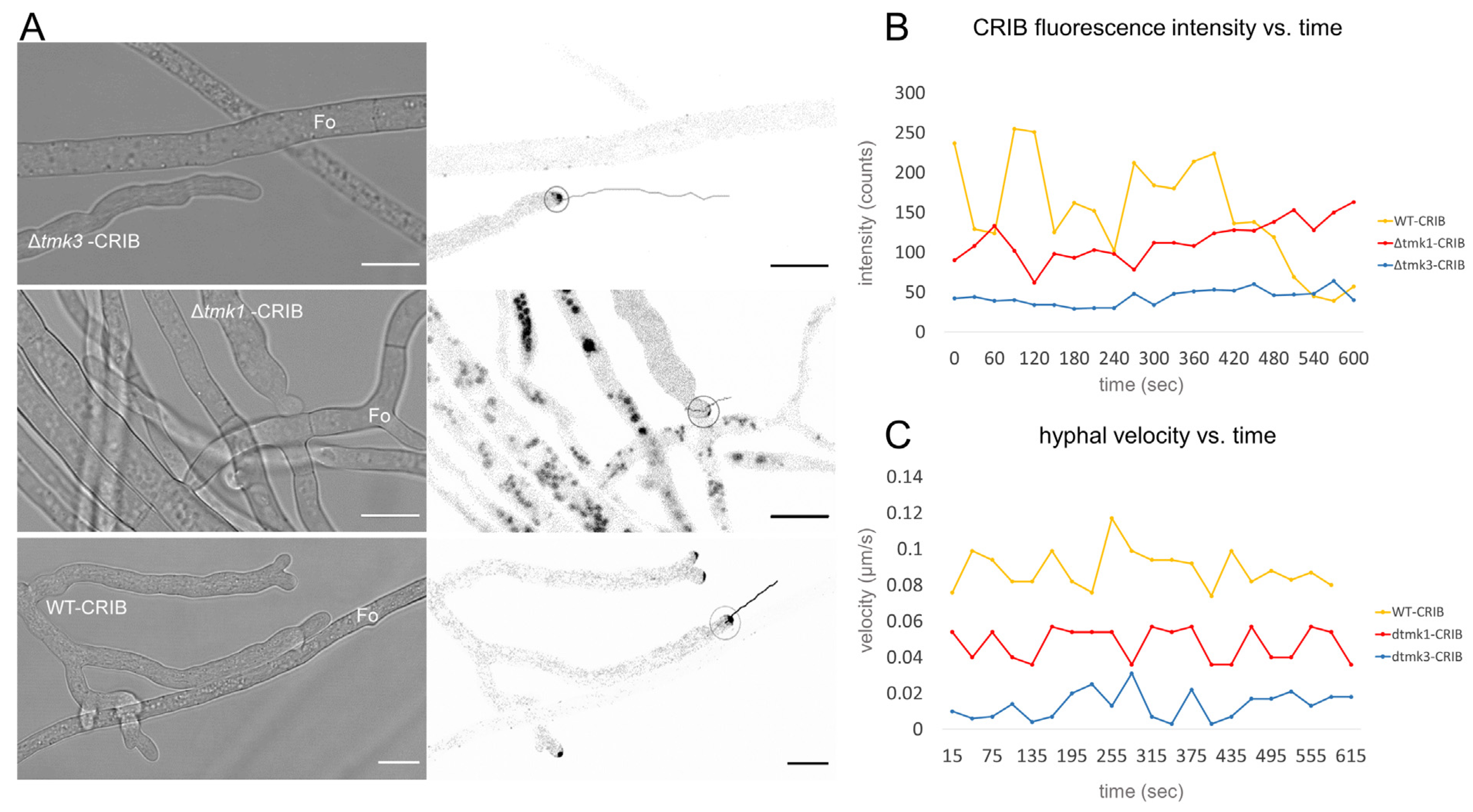
3.4. Tmk1 Is Required to Establish Contact with Prey Hyphae
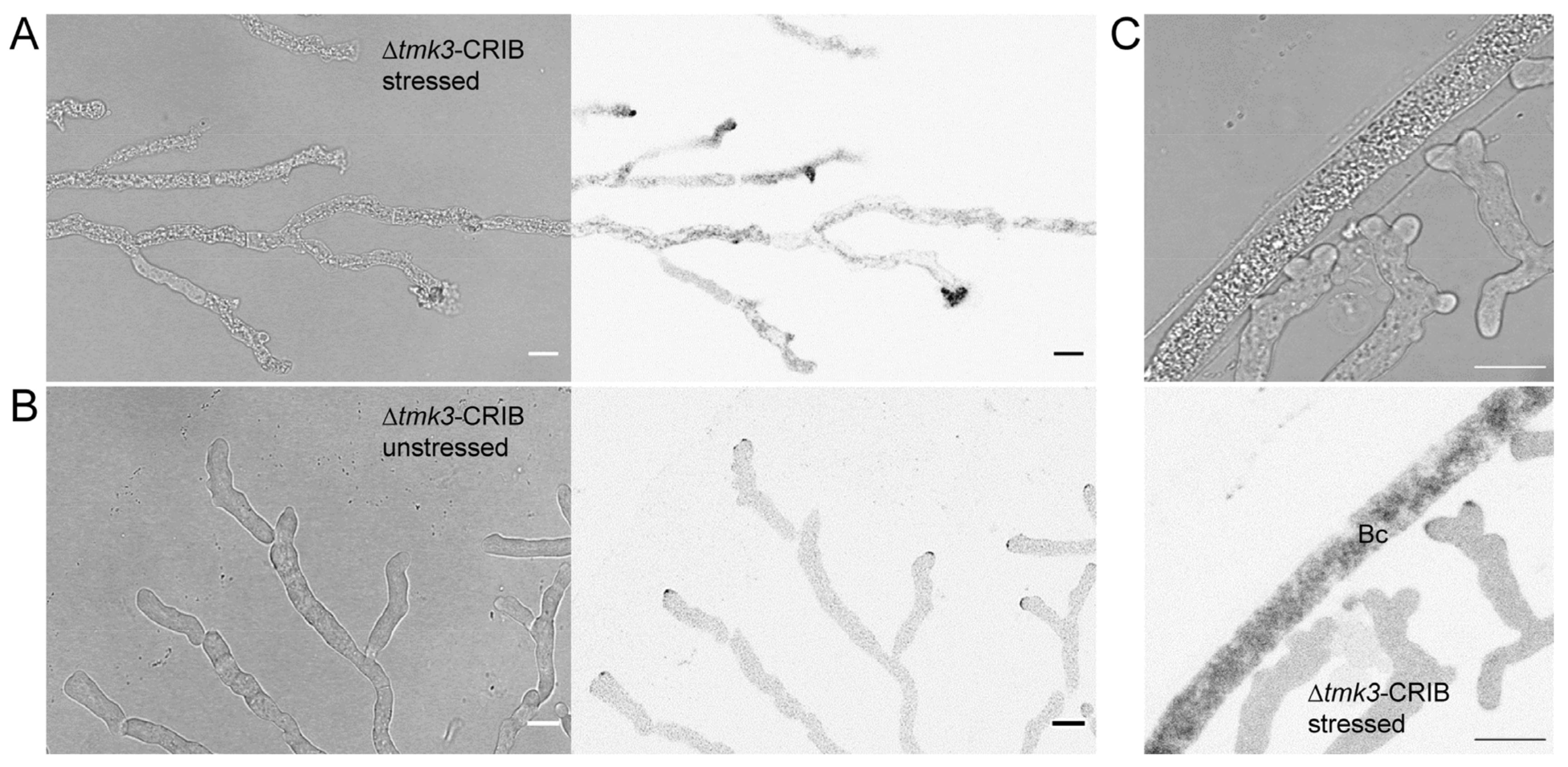
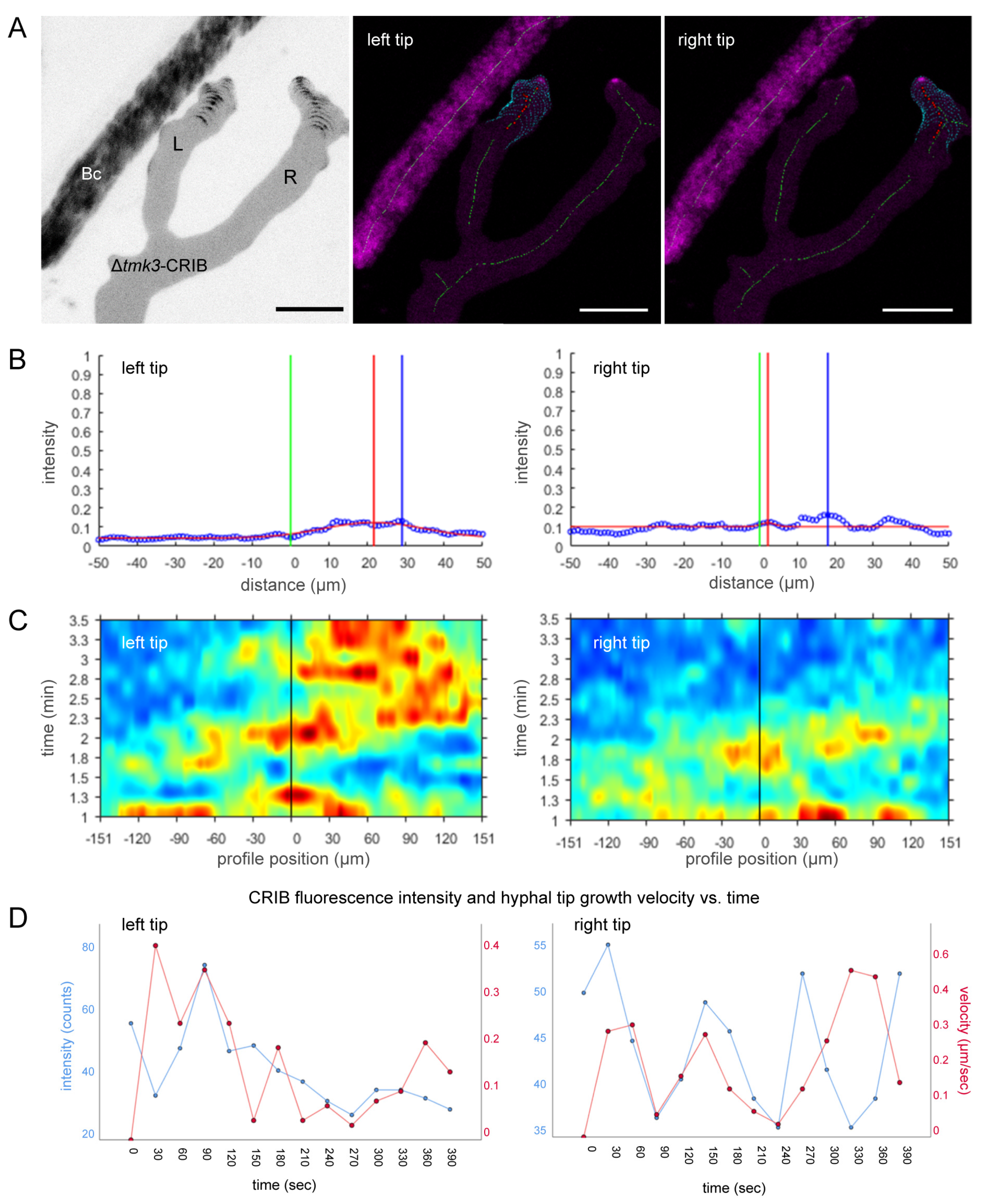
3.5. Loss of Tmk3 Reduces Hyphal Survival during Pre-Contact and Impairs the Formation of Effective GTPase Activity Clusters
4. Discussion
4.1. CRIB Reporters Facilitate Quantification of Hyphal Morphogenesis in Mycoparasitic Interactions
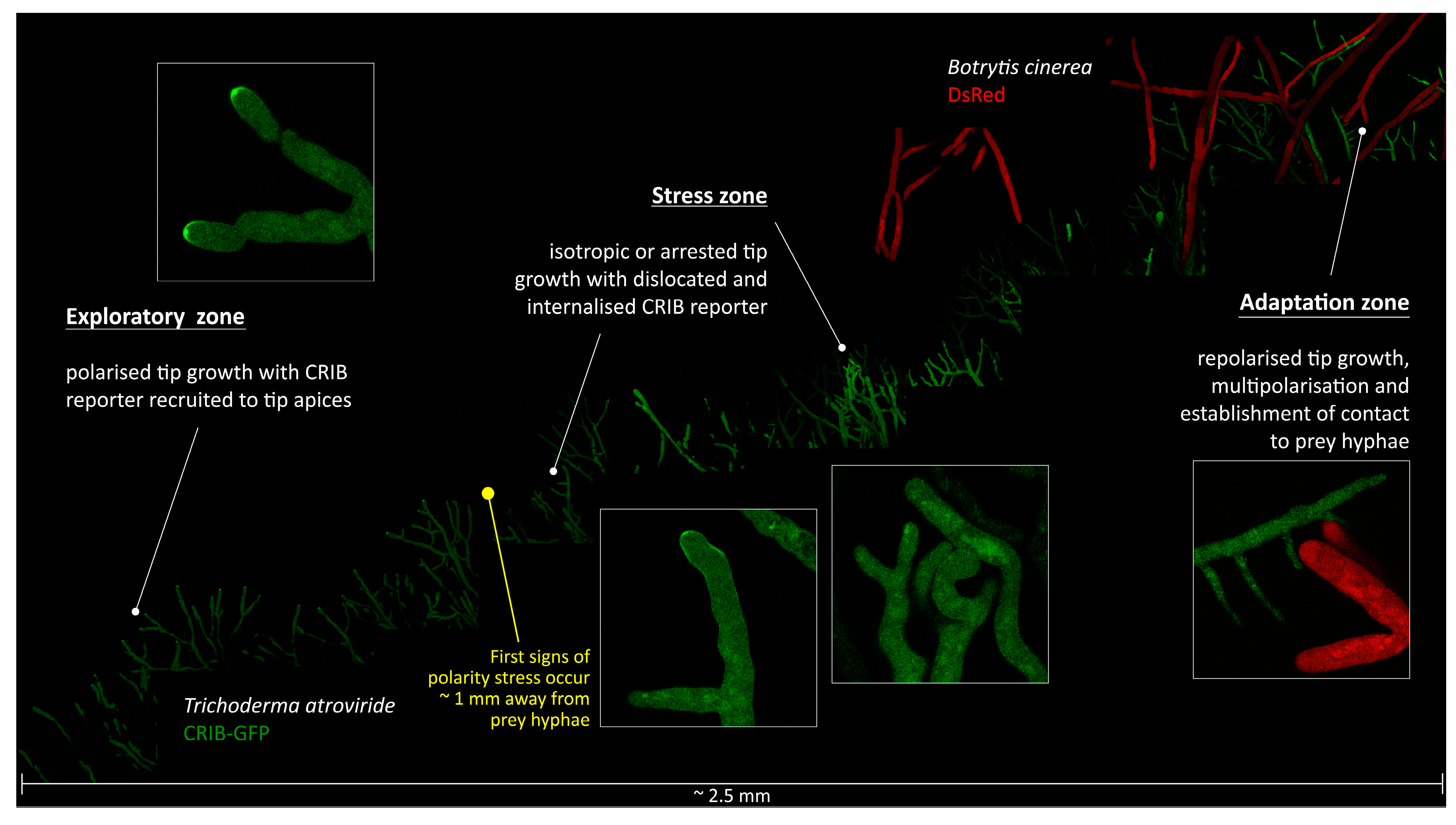
4.2. T. atroviride Has to Overcome Prey-Induced Polarity Stress to Initiate Mycoparasitic Morphogenesis
4.3. The MAP/SAP Kinases Are Required for Polarity Stress Adaptation and Contact-Induced Mycoparasitism
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Błaszczyk, L.; Siwulski, M.; Sobieralski, K.; Lisiecka, J.; Jędryczka, M. Trichoderma spp.—Application and prospects for use in organic farming and industry. J. Plant Prot. Res. 2014, 54, 309–317. [Google Scholar] [CrossRef]
- Qualhato, T.F.; Lopes, F.A.C.; Steindorff, A.S.; Brandão, R.S.; Jesuino, R.S.A.; Ulhoa, C.J. Mycoparasitism studies of Trichoderma species against three phytopathogenic fungi: Evaluation of antagonism and hydrolytic enzyme production. Biotechnol. Lett. 2013, 35, 1461–1468. [Google Scholar] [CrossRef] [PubMed]
- Dubey, S.C.; Suresh, M.; Singh, B. Evaluation of Trichoderma species against Fusarium oxysporum f. sp. ciceris for integrated management of chickpea wilt. Biol. Control 2007, 40, 118–127. [Google Scholar] [CrossRef]
- Chet, I.; Harman, G.E.; Baker, R. Trichoderma hamatum. Its hyphal interactions with Rhizoctonia solani and Pythium spp. Microb. Ecol. 1981, 7, 29–38. [Google Scholar] [CrossRef]
- Chet, I. Mycoparasitism—Recognition, physiology and ecology. New Directions in Biological Control. Alternatives for Suppressing Agricultural Pests and Diseases. In Proceedings of the UCLA Colloquim, Frisco, CO, USA, 20–27 January 1990; pp. 725–733. [Google Scholar]
- Viterbo, A.; Horwitz, B.A. Mycoparasitism. In Cellular and Molecular Biology of Filamentous Fungi; Borkovich, K.A., Ebbole, D.J., Eds.; ASM Press: Washington, DC, USA, 2010; pp. 676–693. [Google Scholar] [CrossRef]
- Miyoshi, M. Ueber chemotropismus der Pilze. Bot. Zeitung 1984, 52, 1–28. [Google Scholar]
- Moreno-Ruiz, D.; Lichius, A.; Turrà, D.; Di Pietro, A.; Zeilinger, S. Chemotropism Assays for Plant Symbiosis and Mycoparasitism Related Compound Screening in Trichoderma atroviride. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Brunner, K.; Peterbauer, C.K.; Mach, R.L.; Lorito, M.; Zeilinger, S.; Kubicek, C.P. The Nag1 N-acetylglucosaminidase of Trichoderma atroviride is essential for chitinase induction by chitin and of major relevance to biocontrol. Curr. Genet. 2003, 43, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codón, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar]
- Viterbo, A.; Inbar, J.; Hadar, Y.; Chet, I. Plant Disease Biocontrol and Induced Resistance via Fungal Mycoparasites. In The Mycota. A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research, 2nd ed.; Kubicek, C.P., Druzhinina, I.S., Eds.; Springer: Berlin, Germany, 2007; pp. 127–146. [Google Scholar]
- Seidl, V.; Song, L.; Lindquist, E.; Gruber, S.; Koptchinskiy, A.; Zeilinger, S.; Schmoll, M.; Martínez, P.; Sun, J.; Grigoriev, I.; et al. Transcriptomic response of the mycoparasitic fungus Trichoderma atroviride to the presence of a fungal prey. BMC Genom. 2009, 10, 567. [Google Scholar] [CrossRef] [Green Version]
- Cortés, C.; Gutierrez, A.; Olmedo, V.; Inbar, J.; Chet, I.; Herrera-Estrella, A. The expression of genes involved in parasitism by Trichoderma harzianum is triggered by a diffusible factor. Mol. Gen. Genet. 1998, 260, 218–225. [Google Scholar] [CrossRef]
- Zeilinger, S.; Galhaup, C.; Payer, K.; Woo, S.L.; Mach, R.L.; Fekete, C.; Lorito, M.; Kubicek, C.P. Chitinase gene expression during mycoparasitic interaction of Trichoderma harzianum with its host. Fungal Genet. Biol. 1999, 26, 131–140. [Google Scholar] [CrossRef]
- Kullnig, C.; Mach, R.L.; Lorito, M.; Kubicek, C.P. Enzyme Diffusion from Trichoderma atroviride (=T. harzianum P1) to Rhizoctonia solani Is a Prerequisite for Triggering of Trichoderma ech42 Gene Expression before Mycoparasitic Contact. Appl. Environ. Microbiol. 2000, 66, 2232–2234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reithner, B.; Ibarra-Laclette, E.; Mach, R.L.; Herrera-Estrella, A. Identification of mycoparasitism-related genes in Trichoderma atroviride. Appl. Environ. Microbiol. 2011, 77, 4361–4370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viterbo, A.; Montero, M.; Ramot, O.; Friesem, D.; Monte, E.; Llobell, A.; Chet, I. Expression regulation of the endochitinase chit36 from Trichoderma asperellum (T. harzianum T-203). Curr. Genet. 2002, 42, 114–122. [Google Scholar] [CrossRef]
- Howell, C.R. Mechanisms Employed by Trichoderma Species in the Biological Control of Plant Diseases. The History and Evolution of Current Concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Lu, Z.; Tombolini, R.; Woo, S.; Zeilinger, S.; Lorito, M.; Jansson, J.K. In vivo study of Trichoderma-pathogen-plant interactions, using constitutive and inducible green fluorescent protein reporter systems. Appl. Environ. Microbiol. 2004, 70, 3073–3081. [Google Scholar] [CrossRef] [Green Version]
- Riquelme, M.; Martinez-Nunez, L. Hyphal ontogeny in Neurospora crassa. a model organism for all seasons. F1000Research 2016, 5, 2801. [Google Scholar] [CrossRef] [Green Version]
- Jaffe, A.B.; Hall, A. Rho GTPases: Biochemistry and biology. Annu. Rev. Cell Dev. Biol. 2005, 21, 247–269. [Google Scholar] [CrossRef] [Green Version]
- Virag, A.; Lee, M.P.; Si, H.; Harris, S.D. Regulation of hyphal morphogenesis by cdc42 and rac1 homologues in Aspergillus nidulans. Mol. Microbiol. 2007, 66. [Google Scholar] [CrossRef] [Green Version]
- Bishop, A.L.; Hall, A. Rho GTPases and their effector proteins. Biochem. J. 2000, 348, 241–255. [Google Scholar] [CrossRef]
- Chiou, J.G.; Balasubramanian, M.K.; Lew, D.J. Cell Polarity in Yeast. Ann. Rev. Cell Dev. Biol. 2017, 15, 365–391. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Pryciak, P.M. Identification of Novel Membrane-binding Domains in Multiple Yeast Cdc42 Effectors. Mol. Biol. Cell. 2007, 18, 4945–4956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudolph, M.G.; Bayer, P.; Abo, A.; Kuhlmann, J.; Vetter, I.R.; Wittinghofer, A. The Cdc42/Rac Interactive Binding Region Motif of the Wiskott Aldrich Syndrome Protein (WASP) Is Necessary but Not Sufficient for Tight Binding to Cdc42 and Structure Formation. J. Biol. Chem. 1998, 273, 18067–18076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen, D.; Mott, H.R.; Laue, E.D.; Lowe, P.N. Residues in Cdc42 That Specify Binding to Individual CRIB Effector Proteins. Biochemistry 2000, 39, 1243–1250. [Google Scholar] [CrossRef]
- Ozbudak, E.M.; Becskei, A.; van Oudenaarden, A. A system of counteracting feedback loops regulates Cdc42p activity during spontaneous cell polarization. Dev. Cell. 2005, 9, 565–571. [Google Scholar] [CrossRef] [Green Version]
- Lichius, A.; Goryachev, A.B.; Fricker, M.D.; Obara, B.; Castro-Longoria, E.; Read, N.D. CDC-42 and RAC-1 regulate opposite chemotropisms in Neurospora crassa. J. Cell Sci. 2014, 127, 1953–1965. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, P.K.; Latha, J.; Hadar, R.; Horwitz, B.A. TmkA, a Mitogen-Activated Protein Kinase of Trichoderma virens, Is Involved in Biocontrol Properties and Repression of Conidiation in the Dark. Eukaryot. Cell 2003, 2, 446–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reithner, B.; Schuhmacher, R.; Stoppacher, N.; Pucher, M.; Brunner, K.; Zeilinger, S. Signaling via the Trichoderma atroviride mitogen-activated protein kinase Tmk 1 differentially affects mycoparasitism and plant protection. Fungal Genet. Biol. 2007, 44, 1123–1133. [Google Scholar] [CrossRef] [Green Version]
- Gruber, S.; Zeilinger, S. The transcription factor Ste12 mediates the regulatory role of the Tmk1 MAP kinase in mycoparasitism and vegetative hyphal fusion in the filamentous fungus Trichoderma atroviride. PLoS ONE 2014, 9, e111636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Dong, Y.; Zhao, Q.; Wang, F.; Liu, K.; Jiang, B.; Fang, X. Identification of the role of a MAP kinase Tmk2 in Hypocrea jecorina (Trichoderma reesei). Sci. Rep. 2014, 4, 6732. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, M.; Li, L.; Dong, Y.; Jiang, Y.; Liu, K.; Zhang, R.; Jiang, B.; Niu, K.; Fang, X. Role of Trichoderma reesei mitogen-activated protein kinases (MAPKs) in cellulase formation. Biotechnol. Biofuels 2017, 10, 99. [Google Scholar] [CrossRef] [PubMed]
- de Paula, R.G.; Antoniêto, A.C.C.; Carraro, C.B.; Lopes, D.C.B.; Persinoti, G.F.; Peres, N.T.A.; Martinez-Rossi, N.M.; Silva-Rocha, R.; Silva, R.N. The Duality of the MAPK Signaling Pathway in the Control of Metabolic Processes and Cellulase Production in Trichoderma reesei. Sci. Rep. 2018, 8, 14931. [Google Scholar] [CrossRef]
- Zheng, D.; Zhang, S.; Zhou, X.; Wang, C.; Xiang, P.; Zheng, Q.; Xu, J.R. The FgHOG1 pathway regulates hyphal growth, stress responses, and plant infection in Fusarium graminearum. PLoS ONE 2012, 7, e49495. [Google Scholar] [CrossRef]
- Saito, H.; Posas, F. Response to Hyperosmotic Stress. Genetics 2012, 192, 289–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Zhao, Q.; Yang, J.; Jiang, B.; Wang, F.; Liu, K.; Fang, X. A Mitogen-Activated Protein Kinase Tmk3 Participates in High Osmolarity Resistance, Cell Wall Integrity Maintenance and Cellulase Production Regulation in Trichoderma reesei. PLoS ONE 2013, 8, e072189. [Google Scholar] [CrossRef] [Green Version]
- Medina-Castellanos, E.; Esquivel-Naranjo, E.U.; Heil, M.; Herrera-Estrella, A. Extracellular ATP activates MAPK and ROS signaling during injury response in the fungus Trichoderma atroviride. Front. Plant Sci. 2014, 5, 659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.A.; Morgan, B.A.; Quinn, J. Stress signalling to fungal stress-activated protein kinase pathways. FEMS Microbiol. Lett. 2010, 306, 1–8. [Google Scholar] [CrossRef]
- Moreno-Ruiz, D.; Fuchs, A.; Missbach, K.; Schuhmacher, R.; Zeilinger, S. Influence of Different Light Regimes on the Mycoparasitic Activity and 6-Pentyl-α-pyrone Biosynthesis in Two Strains of Trichoderma atroviride. Pathogens 2020, 9, 860. [Google Scholar] [CrossRef]
- Schumacher, J. Tools for Botrytis cinerea: New expression vectors make the gray mold fungus more accessible to cell biology approaches. Fungal Genet. Biol. 2012, 49, 483–497. [Google Scholar] [CrossRef]
- Lichius, A.; Seidl-Seiboth, V.; Seiboth, B.; Kubicek, C.P. Nucleo-cytoplasmic shuttling dynamics of the transcriptional regulators XYR1 and CRE1 under conditions of cellulase and xylanase gene expression in Trichoderma reesei. Mol. Microbiol. 2014, 94, 1162–1178. [Google Scholar] [CrossRef]
- Salzmann, L. Generation of Live-Cell Imaging Markers for Mycoparasitism in Trichoderma atroviride. Master’s Thesis, University of Innsbruck, Innsbruck, Austria, 2017. [Google Scholar]
- Freitag, M.; Hickey, P.C.; Raju, N.B.; Selker, E.U.; Read, N.D. GFP as a tool to analyse the organization, dynamics and function of nuclei and microtubules in Neurospora crassa. Fungal Genet. Biol. 2004, 41, 897–910. [Google Scholar] [CrossRef] [PubMed]
- Teerawanichpan, P.; Hoffman, T.; Ashe, P.; Datla, R.; Selvaraj, G. Investigations of combinations of mutations in the jellyfish green fluorescent protein (GFP) that afford brighter fluorescence, and use of a version (VisGreen) in plant, bacterial, and animal cells. Biochim. Biophys. Acta 2007, 1770, 1360–1368. [Google Scholar] [CrossRef]
- Kück, U.; Hoff, B. Application of the nourseothricin acetyltransferase gene (nat1) as dominant marker for transformation of filamentous fungi. Fungal Genet. Rep. 2006, 53, 9–11. [Google Scholar] [CrossRef] [Green Version]
- Atanasova, L.; Gruber, S.; Lichius, A.; Radebner, T.; Abendstein, L.; Münsterkötter, M.; Stralis-Pavese, N.; Łabaj, P.P.; Kreil, D.P.; Zeilinger, S. The Gpr1-regulated Sur7 family protein Sfp2 is required for hyphal growth and cell wall stability in the mycoparasite Trichoderma atroviride. Sci. Rep. 2018, 8, 12064. [Google Scholar] [CrossRef]
- Gruber, F.; Visser, J.; Kubicek, C.P.; de Graaff, L.H. The development of a heterologous transformation system for the cellulolytic fungus Trichoderma reesei based on a pyrG-negative mutant strain. Curr. Genet. 1990, 18. [Google Scholar] [CrossRef]
- Hickey, P.C.; Swift, S.R.; Roca, M.G.R.; Read, N.D. Live-cell imaging of filamentous fungi using vital fluorescent dyes and confocal microscopy. Methods Microbiol. 2004, 34, 63–87. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Tinevez, J.Y.; Perry, N.; Schindelin, J.; Hoopes, G.M.; Reynolds, G.D.; Laplantine, E.; Bednarek, S.Y.; Shorte, S.Y.; Eliceiri, K.W. TrackMate: An open and extensible platform for single-particle tracking. Methods 2017, 115, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.D. Cdc42/Rho GTPases in fungi: Variations on a common theme. Mol. Microbiol. 2011, 79, 1123–1127. [Google Scholar] [CrossRef] [PubMed]
- Tatebe, H.; Nakano, K.; Maximo, R.; Shiozaki, K. Pom1 DYRK regulates localization of the Rga4 GAP to ensure bipolar activation of Cdc42 in fission yeast. Curr. Biol. 2008, 18, 322–330. [Google Scholar] [CrossRef] [Green Version]
- Corvest, V.; Bogliolo, S.; Follette, P.; Arkowitz, R.A.; Bassilana, M. Spatiotemporal regulation of Rho1 and Cdc42 activity during Candida albicans filamentous growth. Mol. Microbiol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Heim, R.; Cubitt, A.B.; Tsien, R.Y. Improved green fluorescence. Nature 1995, 373, 663–664. [Google Scholar] [CrossRef] [PubMed]
- Zeilinger, S.; Omann, M. Trichoderma biocontrol: Signal transduction pathways involved in host sensing and mycoparasitism. Gene Regul. Syst. Biol. 2007, 1, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Druzhinina, I.S.; Seidl-Seiboth, V.; Herrera-Estrella, A.; Horwitz, B.A.; Kenerley, C.M.; Monte, E.; Mukherjee, P.K.; Zeilinger, S.; Grigoriev, I.V.; Kubicek, C.P. Trichoderma: The genomics of opportunistic success. Nat. Rev. Microbiol. 2011, 9, 749–759. [Google Scholar] [CrossRef]
- Mukherjee, M.; Mukherjee, P.K.; Horwitz, B.A.; Zachow, C.; Berg, G.; Zeilinger, S. Trichoderma-plant-pathogen interactions: Advances in genetics of biological control. Indian J. Microbiol. 2012, 52, 522–529. [Google Scholar] [CrossRef] [Green Version]
- López-Berges, M.S.; Hera, C.; Sulyok, M.; Schäfer, K.; Capilla, J.; Guarro, J.; Di Pietro, A. The velvet complex governs mycotoxin production and virulence of Fusarium oxysporum on plant and mammalian hosts. Mol. Microbiol. 2013, 87, 49–65. [Google Scholar] [CrossRef]
- Bonfante, P.; Genre, A. Mechanisms underlying beneficial plant-fungus interactions in mycorrhizal symbiosis. Nat. Commun. 2010, 1, 48. [Google Scholar] [CrossRef] [Green Version]
- Lichius, A.; Lord, K.M. Chemoattractive Mechanisms in Filamentous Fungi. Open Mycol. J. 2014, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Turrà, D.; Di Pietro, A. Chemotropic sensing in fungus-plant interactions. Curr. Opin. Plant Biol. 2015, 26, 135–140. [Google Scholar] [CrossRef]
- Inbar, J.; Chet, I. Biomimics of fungal cell-cell recognition by use of lectin-coated nylon fibers. J. Bacteriol. 1992, 174, 1055–1059. [Google Scholar] [CrossRef] [Green Version]
- Inbar, J.; Chet, I. A newly isolated lectin from the plant pathogenic fungus Sclerotium rolfsii: Purification, characterization and role in mycoparasitism. Microbiology 1994, 140, 651–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inbar, J.; Chet, I. The Role of Lectins in Recognition and Adhesion of the Mycoparasitic Fungus Trichoderma spp. In Toward Anti-Adhesion Therapy for Microbial Diseases to Its Host; Itzhak, K., Itzhak, O., Eds.; Springer: Boston, MA, USA, 1996; Volume 408, pp. 229–231. [Google Scholar]
- Benhamou, N.; Chet, I. Hyphal interaction between Trichoderma harzianum and Rhizoctonia solani: Ultrastructure and gold cytochemistry of the mycoparasitic process. Phytopathology 1993, 83, 1062–1071. [Google Scholar] [CrossRef]
- Elad, Y.; Barak, R.; Chet, I. Possible role of lectins in mycoparasitism. J. Bacteriol. 1983, 154, 1431–1435. [Google Scholar] [CrossRef] [Green Version]
- Murmanis, L.L.; Highley, T.L.; Ricard, J. Hyphal interaction of Trichoderma harzianum and Trichoderma polysporum with wood decay fungi. Mater. Org. 1988, 23, 271–279. [Google Scholar]
- Watters, M.K.; Humphries, C.; de Vries, I.; Griffiths, A.J.F. A homeostatic set point for branching in Neurospora crassa. Mycol. Res. 2000, 104, 557–563. [Google Scholar] [CrossRef]
- Riquelme, M.; Bartnicki-Garcia, S. Key differences between lateral and apical branching in hyphae of Neurospora crassa. Fungal Genet. Biol. 2004, 41, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, K.S.; Gow, N.A.; Davidson, F.A.; Gadd, G.M. Regulation of vectorial supply of vesicles to the hyphal tip determines thigmotropism in Neurospora crassa. Fungal Biol. 2014, 118, 287–294. [Google Scholar] [CrossRef]
- Thomson, D.D.; Wehmeier, S.; Byfield, F.R.J.; Janmey, P.A.; Caballero-Lima, D.; Crossley, A.; Brand, A.C. Contact-induced apical asymmetry drives the thigmotropic responses of Candida albicans hyphae. Cell. Microbiol. 2015, 17, 342–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benhamou, N.; Chet, I. Cellular and Molecular Mechanisms Involved in the Interaction between Trichoderma harzianum and Pythium ultimum. Appl. Environ. Microbiol. 1997, 63, 2095–2099. [Google Scholar] [CrossRef] [Green Version]
- El-Debaiky, S.A. Antagonistic studies and hyphal interactions of the new antagonist Aspergillus piperis against some phytopathogenic fungi in vitro in comparison with Trichoderma harzianum. Microb. Pathog. 2017, 113, 135–143. [Google Scholar] [CrossRef]
- Turrà, D.; El Ghalid, M.; Rossi, F.; Di Pietro, A. Fungal pathogen uses sex pheromone receptor for chemotropic sensing of host plant signals. Nature 2015, 527, 521–524. [Google Scholar] [CrossRef]
- Delgado-Jarana, J.; Sousa, S.; González, F.; Rey, M.; Llobell, A. ThHog1 controls the hyperosmotic stress response in Trichoderma harzianum. Microbiology 2006, 152, 1687–1700. [Google Scholar] [CrossRef] [Green Version]
- Atriztán-Hernández, K.; Moreno-Pedraza, A.; Winkler, R.; Markow, T.; Herrera-Estrella, A. Trichoderma atroviride from Predator to Prey: Role of the Mitogen-Activated Protein Kinase Tmk3 in Fungal Chemical Defense against Fungivory by Drosophila melanogaster Larvae. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef] [Green Version]
- Reiser, V.; Salah, S.M.; Anmerer, G. Polarized localization of yeast Pbs2 depends on osmostress, the membrane protein Sho1 and Cdc42. Nat. Cell Biol. 2000, 2, 620–627. [Google Scholar] [CrossRef] [PubMed]
- Raitt, D.C.; Posas, F.; Saito, H. Yeast Cdc42 GTPase and Ste20 PAK-like kinase regulate Sho1-dependent activation of the Hog1 MAPK pathway. EMBO J. 2000, 19, 4623–4631. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Zhou, X.; Zhao, X.; Xu, J.R. The PMK1 MAP Kinase Pathway and Infection-Related Morphogenesis. In Advances in Genetics, Genomics and Control of Rice Blast Disease; Wang, G.L., Barbara, V., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 13–21. [Google Scholar]
- Turrà, D.; Nordzieke, D.; Vitale, S.; El Ghalid, M.; Di Pietro, A. Hyphal chemotropism in fungal pathogenicity. Semin. Cell Dev. Biol. 2016, 57, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Elion, E.A.; Satterberg, B.; Kranz, J.E. FUS3 phosphorylates multiple components of the mating signal transduction cascade: Evidence for STE12 and FAR1. Mol. Biol. Cell 1993, 4, 495–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tedford, K.; Kim, S.; Sa, D.; Stevens, K.; Tyers, M. Regulation of the mating pheromone and invasive growth responses in yeast by two MAP kinase substrates. Curr. Biol. 1997. [Google Scholar] [CrossRef] [Green Version]
- Matheos, D.; Metodiev, M.; Muller, E.; Stone, D.; Rose, M.D. Pheromone-induced polarization is dependent on the Fus3p MAPK acting through the formin Bni1p. J. Cell Biol. 2004, 165, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Ly, N.; Cyert, M.S. Calcineurin, the Ca2+-dependent phosphatase, regulates Rga2, a Cdc42 GTPase-activating protein, to modulate pheromone signaling. Mol. Biol. Cell 2017, 28, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Momany, M.; Westfall, P.J.; Abramowsky, G. Aspergillus nidulans swo Mutants Show Defects in Polarity Establishment, Polarity Maintenance and Hyphal Morphogenesis. Genetics 1999, 151, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Seiler, S.; Plamann, M. The genetic basis of cellular morphogenesis in the filamentous fungus Neurospora crassa. Mol. Biol. Cell 2003, 14, 4352–4364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, Y.; Knechtle, P.; Wendland, J.; Helfer, H.; Philippsen, P. A Ras-like GTPase is involved in hyphal growth guidance in the filamentous fungus Ashbya gossypii. Mol. Biol. Cell 2004, 15, 4622–4632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichius, A.; Yáñez-Gutiérrez, M.E.; Read, N.D.; Castro-Longoria, E. Comparative live-cell imaging analyses of SPA-2, BUD-6 and BNI-1 in Neurospora crassa reveal novel features of the filamentous fungal polarisome. PLoS ONE 2012, 7, e30372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutavchiev, D.R.; Leda, M.; Sawin, K.E. Remodeling of the Fission Yeast Cdc42 Cell-Polarity Module via the Sty1 p38 Stress-Activated Protein Kinase Pathway. Curr. Biol. 2016, 26, 2921–2928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, P.; Soto, T.; Gómez-Gil, E.; Cansado, J. Functional interaction between Cdc42 and the stress MAPK signaling pathway during the regulation of fission yeast polarized growth. Int. Microbiol. 2020, 23, 31–41. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Ruiz, D.; Salzmann, L.; Fricker, M.D.; Zeilinger, S.; Lichius, A. Stress-Activated Protein Kinase Signalling Regulates Mycoparasitic Hyphal-Hyphal Interactions in Trichoderma atroviride. J. Fungi 2021, 7, 365. https://doi.org/10.3390/jof7050365
Moreno-Ruiz D, Salzmann L, Fricker MD, Zeilinger S, Lichius A. Stress-Activated Protein Kinase Signalling Regulates Mycoparasitic Hyphal-Hyphal Interactions in Trichoderma atroviride. Journal of Fungi. 2021; 7(5):365. https://doi.org/10.3390/jof7050365
Chicago/Turabian StyleMoreno-Ruiz, Dubraska, Linda Salzmann, Mark D. Fricker, Susanne Zeilinger, and Alexander Lichius. 2021. "Stress-Activated Protein Kinase Signalling Regulates Mycoparasitic Hyphal-Hyphal Interactions in Trichoderma atroviride" Journal of Fungi 7, no. 5: 365. https://doi.org/10.3390/jof7050365