1. Introduction
Methanol is a promising feedstock alternative to sugar-based raw materials for the bioproduction of fuels, specialty chemicals, polymers, and other value-added products due to its abundance and relatively low cost [
1,
2,
3,
4]. Methanol is also the major impurity in crude glycerol, reaching relatively high levels that can vary considerably from batch to batch and, although it can be removed by evaporation, this process is energy demanding [
5,
6]. Therefore, the utilization of methanol as co-substrate by methanol-tolerant methylotrophic yeasts would increase the feasibility of bioprocesses that use crude glycerol as substrate [
5]. Methanol is also present, at relatively low concentrations, in hydrolysates from pectin-rich agro-industrial residues given that the D-galacturonic acid monomers are methyl-esterified in different positions [
7,
8,
9]. Differently from methylotrophic yeast species, the preferred yeast cell factory
Saccharomyces cerevisiae, is not able to use methanol as sole carbon source but there are successful examples of
S. cerevisiae metabolic engineering for direct methanol utilization [
2,
10,
11]. Although being a promising carbon source for metabolically competent yeast strains, methanol toxicity can limit the productivity of methanol-based biomanufacturing [
3]. For this reason, the identification of genes/proteins whose expression is required for maximum tolerance to methanol in the model yeast species
S. cerevisiae is important for enlightening the mechanisms underlying methanol toxicity in methylotrophic yeasts and in other eukaryotes as well as for guiding the development of more robust yeast strains, in particular methylotrophic yeast strains [
3]. Genome-wide approaches have been enabling a holistic view and a deeper understanding of the molecular mechanisms and signaling pathways involved in the global response and adaptation of yeasts to sublethal concentrations of toxicants by allowing the identification of genes and pathways involved in the toxicological response and required for maximum tolerance [
12,
13]. The genome-wide identification of genes that are determinants of tolerance to methanol is a first step to allow the improvement of yeast robustness and the objective of the present study. Such chemogenomic analysis, using a
S. cerevisiae deletion mutant collection, has been explored before to identify genes required for maximum tolerance to a variety of relevant chemical stresses including compounds of biotechnological, agronomical and pharmaceutical interest [
12,
13].
The methanol detoxification pathway in
S. cerevisiae involves two reactions: (i) the oxidation of methanol to formaldehyde carried out by alcohol dehydrogenases, and (ii) the oxidation of formaldehyde to formic acid catalyzed by an aldehyde dehydrogenase [
14]. The genome-wide response of yeast to methanol, based on transcriptomic analyses, was reported in two studies. Results suggest that the major cellular targets for methanol and formaldehyde toxicity are membrane structure and proteins, respectively [
14], and that the response to methanol also includes the up-regulation, at different levels and depending on the yeast strain, of genes of mitochondrial and peroxisomal metabolism, alcohol and formate dehydrogenation, glutathione metabolism, at different levels, [
15]. The screening of the same yeast deletion mutant collection used in our study was carried out for the identification of formaldehyde tolerance determinants [
16,
17]. Among them, DNA repair mechanisms were found to underlie formaldehyde tolerance, consistent with the alkylating activity of this compound [
16,
17]. Concerning formic acid, produced in the last step of methanol detoxification in yeast [
14], another chemogenomic analysis performed in our lab, indicates an enrichment of tolerance genes involved in intracellular trafficking and protein synthesis, cell wall and cytoskeleton organization, carbohydrate metabolism, lipid, amino acid and vitamin metabolism, response to stress, chromatin remodeling, transcription, and internal pH homeostasis [
18]. This study also confirms the involvement of the Haa1 transcription factor and the Haa1-regulon in the tolerance to formic acid [
18], as described for acetic acid [
19].
Although the available studies on the determinants and signaling pathways involved in yeast tolerance to methanol are scarce, several reports on tolerance to ethanol toxicity are available, in particular at the genome-wide level [
20,
21,
22,
23,
24,
25]. Due to the structural similarity of these short chain alcohols, the knowledge gathered for ethanol can be useful to understand methanol toxicity and tolerance in yeast. The screening of the yeast disruptome carried out in our laboratory for ethanol, using the same experimental methodology applied in the present work for methanol, identified as enriched in the obtained dataset genes associated with intracellular organization, biogenesis, and transport regarding the vacuole, the peroxisome, the endosome, and the cytoskeleton and the transcriptional machinery [
22]. The clustering of the encoded proteins, based on their known physical and genetic interactions, highlighted the importance of the vacuolar protein sorting machinery, the vacuolar H (
+)-ATPase (V-ATPase) complex, and the peroxisome protein import machinery [
22]. Several plasma-membrane H
+-ATPase and vacuolar H
+-ATPase (V-ATPase) genes which are essential for maintaining the intracellular pH at physiological values [
26,
27,
28,
29], were also found in other genome-wide screenings as ethanol tolerance determinants [
20,
22,
23,
24,
25]. As a lipophilic agent, ethanol leads to the perturbation of plasma membrane lipid organization and consequently to the increase of its non-specific permeability, disrupting membrane biological function as a matrix for proteins and, thus, affecting their activity [
30,
31]. Alterations of plasma membrane lipid composition is among the responses of the yeast cell considered useful to counteract those perturbations, namely at the level of sterol and unsaturated fatty acids composition and content [
29,
32,
33]. Peroxisomal function is also responsible for yeast tolerance to ethanol with phospholipid biosynthesis since cells with abnormal peroxisomal function are unable to regulate the composition of membrane phospholipids, compromising membrane remodeling to overcome ethanol stress [
22,
25].
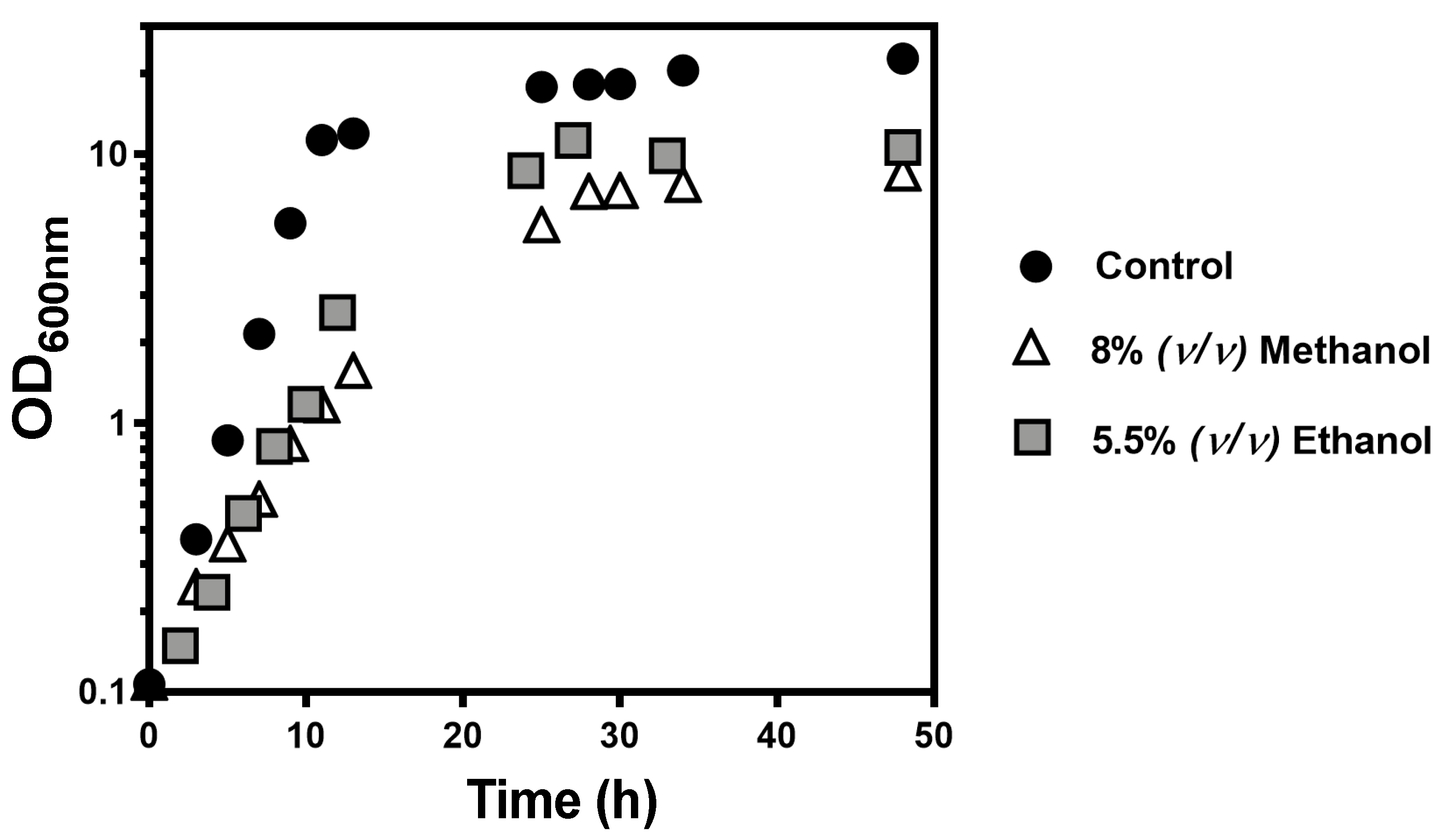
The first goal of the present study was to get insights into the global mechanisms underlying methanol toxicity through the identification of tolerance determinant genes by screening the entire Euroscarf haploid deletion mutant collection grown in YPD medium supplemented with 8% (
v/
v) methanol at 35 °C. This chemogenomic analysis was extended to an equivalent growth inhibitory concentration of ethanol of 5.5% (
v/
v) performed under identical experimental conditions to compare the mechanisms underlying methanol and ethanol toxicity and tolerance. This was considered essential because the various available genome-wide studies that allowed the identification of genetic determinants of ethanol tolerance were performed under different experimental conditions: in rich medium with ethanol concentrations of 7% (
v/
v) [
23], 10% (
v/
v) [
20,
21] and 11% (
v/
v) [
24] and in minimal medium supplemented with 8% (
v/
v) ethanol [
22]. Moreover, the criteria used to identify genes that when deleted lead to ethanol susceptibility phenotypes varied. The results obtained in the present work indicate that, despite the similarities identified for a vast number of genetic determinants of tolerance to these two alcohols, DNA repair and membrane remodeling are among the more specific responses to counteract methanol toxicity. Results from this genome-wide search for genes that confer tolerance to methanol in
S. cerevisiae can now be explored for the rational genetic manipulation of yeasts to obtain more robust strains capable to cope with stressing methanol concentrations, in particular of methylotrophic yeasts.
4. Discussion
The chemogenomics analysis performed in this work provides new information on methanol toxicity and tolerance mechanisms in yeast, being the first disruptome study conducted to unveil methanol tolerance determinants at a genome-wide scale. Although the identification of methanol tolerance determinants in yeast was the main objective of this study, we also intended to compare the major determinants of tolerance to methanol versus ethanol to get clues on more specific methanol toxicity mechanisms and to identify relevant tolerance determinants common to both alcohols to guide future efforts concerning yeast robustness engineering. The yeast disruptome was previously screened for susceptibility to ethanol stress in several studies [
20,
21,
22,
23,
24,
25]. However, differences registered between the data obtained in the various experimental analyses suggest that the genetic background, growth media, level of stress, and other environmental conditions influence the effect of gene expression in yeast tolerance [
22]. To address this issue, two datasets were obtained in this work by testing equivalent sub-lethal inhibitory concentrations of methanol and ethanol under the same experimental conditions. Among the 402 determinants of tolerance to methanol-induced stress, identified in YPD growth medium supplemented with 8% (
v/
v) at 35 °C, 235 were specific to methanol, thus not shared with the ethanol dataset obtained for 5.5% (
v/
v) ethanol. The main feature that clearly distinguishes methanol from ethanol tolerance determinants relates with DNA repair mechanisms presumably required for overcoming methanol-induced stress. Among the eight
RAD genes (a designation due to the sensitivity of the corresponding mutants to exposure to X-rays [
46]) exclusively present in the methanol dataset and involved in several mechanisms of DNA repair, such as recombinational repair and double-strand break repair [
47,
48] are genes, previously identified as formaldehyde tolerance determinants: the
RAD18,
RAD27,
RAD5,
RAD51, and
RAD57 [
16]. The mechanisms proposed to be required for formaldehyde tolerance involve homologous recombination and nucleotide excision repair; while homologous recombination is considered the preferred mechanism to repair damage due to chronic exposure to formaldehyde, nucleotide excision repair is the preferred mechanism to repair acute exposure [
17]. The identification of genes involved in homologous recombination (Rad51, Rad54, Rad57) and in nucleotide excision repair (Rad33) indicates that both mechanisms are important for methanol tolerance.
Although there are differences between the major methanol or ethanol tolerance determinants, there are tolerance mechanisms in common. Since straight-chain alcohols toxicity can be related with the octanol-water partition coefficient log P
ow value [
49], and log P
ow values for methanol and ethanol are −0.74 and −0.30, respectively [
50], compared with methanol, ethanol is more lipophilic and, therefore, expectably, more toxic as the result of membrane targeting [
51,
52]. Consistent with the concept that membranes are molecular targets for methanol and ethanol toxic effects, genes required for membrane composition control were identified as required for alcohol tolerance, in particular genes involved in the biosynthesis of ergosterol, phospholipids, and sphingolipids [
20,
21,
22,
23,
24,
25]. In this context, the TF Ume6, an important ethanol tolerance determinant [
20,
21,
22,
24] that is known to regulate phospholipid biosynthetic gene expression [
53], was found in methanol and ethanol datasets. Together with other tolerance genes involved in the establishment of membrane composition, those genes reinforce the idea of the importance of membrane remodeling to counteract alcohol stress. Alcohol-induced permeabilization of plasma membrane leads to the increased passive influx of ions, in particular protons, across plasma membrane, contributing to cytosolic acidification and to the dissipation of the electrochemical membrane potential [
36,
37,
54]. Since the H
+-ATPase, present at the vacuolar membrane (V-ATPase), is involved in the regulation of intracellular pH homeostasis [
36] it is understandable that several subunits of this multimeric protein were identified as required for methanol and ethanol tolerance. Methanol is likely an inducer of intracellular and vacuolar acidification, as described for ethanol [
22,
55,
56] and the maintenance of intracellular pH (pH
i) homeostasis is affected by V-ATPase defects, emphasizing the importance of this cellular function in tolerance to various straight-chain alcohols [
21]. The biosynthesis of the reserve polysaccharides, glycogen and trehalose also appears to be relevant for yeast tolerance to methanol and ethanol, as suggested by the identification of two genes involved in the synthesis of trehalose (
TPS1 and
TPS2) and one gene required for glycogen degradation (
GPH1) in the datasets. Trehalose and glycogen accumulate under stress conditions and are important in carbon storage and as compatible solutes [
57] and trehalose exerts a protective effect on biomembranes, avoiding desiccation and protein denaturation [
58].
TPS1 and
GPH1 were also found in other chemogenomic studies as ethanol tolerance determinants [
22,
23,
24,
25].
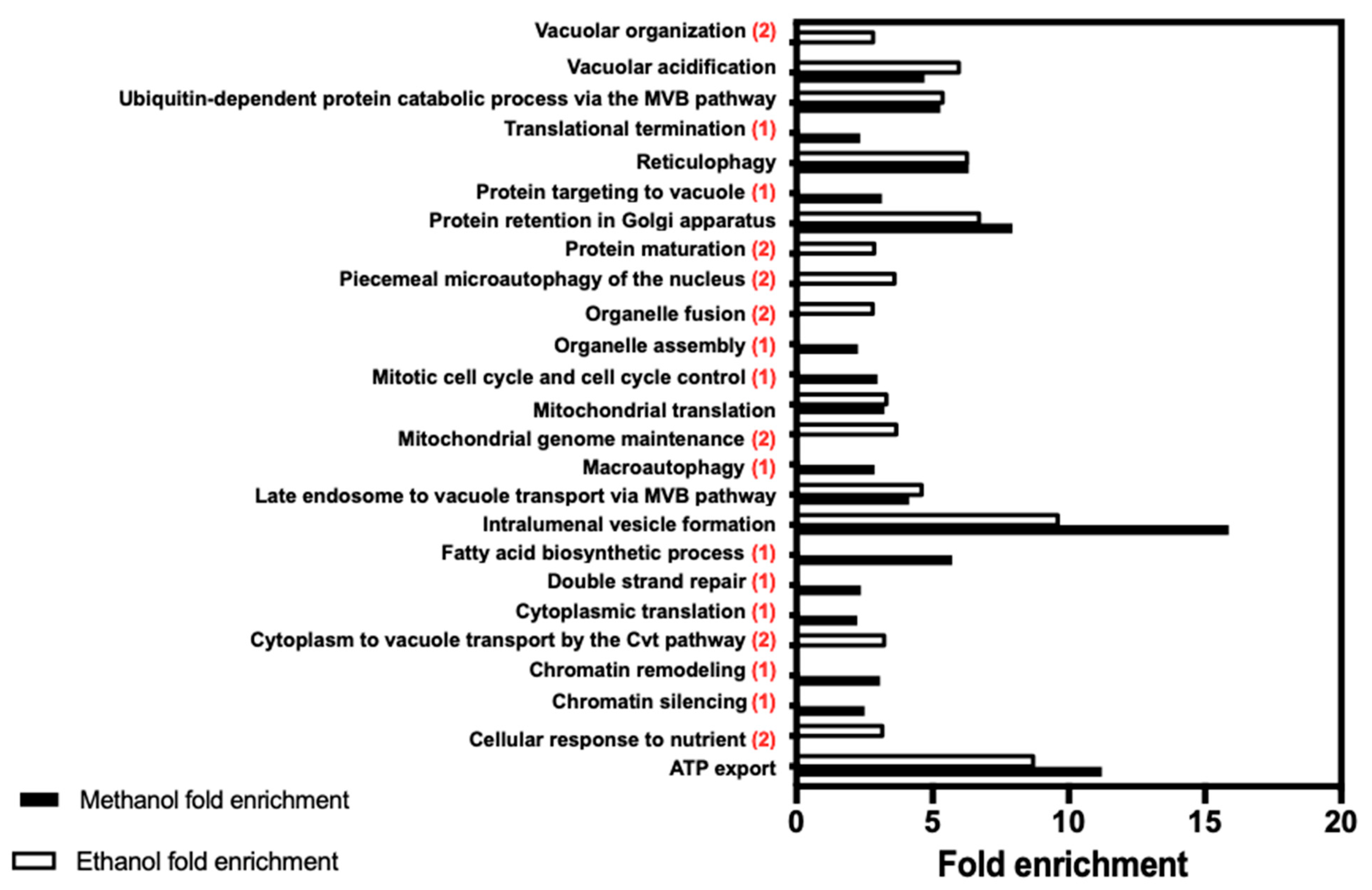
The importance of autophagic processes in methanol and ethanol tolerance is also suggested by our study, a role that can be related with cell protection against DNA and other macromolecules and organelle damaging [
34,
59]. In fact, autophagy is considered a central component of the global stress response [
59]. Reticulophagy, a type of selective autophagy [
34], is an enriched biological function in both datasets. These results are consistent with previous studies reporting that ethanol exposure can lead to endoplasmic reticulum stress, contributing to impaired protein folding and inducing the unfolded protein response [
60]. Methanol also decreases the level of hydration of proteins, leading to tertiary structure modifications, in which polar groups are exposed and can interact mutually [
39,
61]. Formaldehyde, due to reaction with the amino- and sulfhydryl- groups in small molecules, peptides, proteins and nucleic acids, contributes to the formation of inter- and intramolecular bridges [
39,
62]. These conformational changes, caused by methanol or formaldehyde, are comparable to the effect of ethanol inducing endoplasmic reticulum stress, consistent with a common cellular response to these alcohols. The de novo synthesized proteins from the endoplasmic reticulum, destined for secretion from the cell, endocytic processes or to the plasma membrane are delivered to the Golgi apparatus [
63]. The retention of proteins in the Golgi apparatus, suggested by specific methanol and ethanol tolerance determinants obtained in the two datasets of this study, in particular
DID4,
VPS1,
VPS27,
VPS36,
VPS4, and
VPS5 genes, corroborates the occurrence of changes in protein structure induced by the alcohols linking protein defects due to alcohol exposure to protein sequestration in the Golgi. Furthermore, the processes of intracellular trafficking, including vacuolar protein targeting, endosome transport, and transport mediated by the endosomal sorting complexes needed for transport (ESCRT-I, -II, and -III), as well as ubiquitin-dependent protein sorting to the vacuole, were found to confer methanol tolerance, similarly to what was described for ethanol [
20,
21,
22,
23,
24]. The endosomal sorting complexes, as well as ubiquitin-dependent sorting to the vacuole, are important for degradation of methanol/formaldehyde- and ethanol-induced misfolded proteins that were likely to suffer damage due to alcohol toxicity.
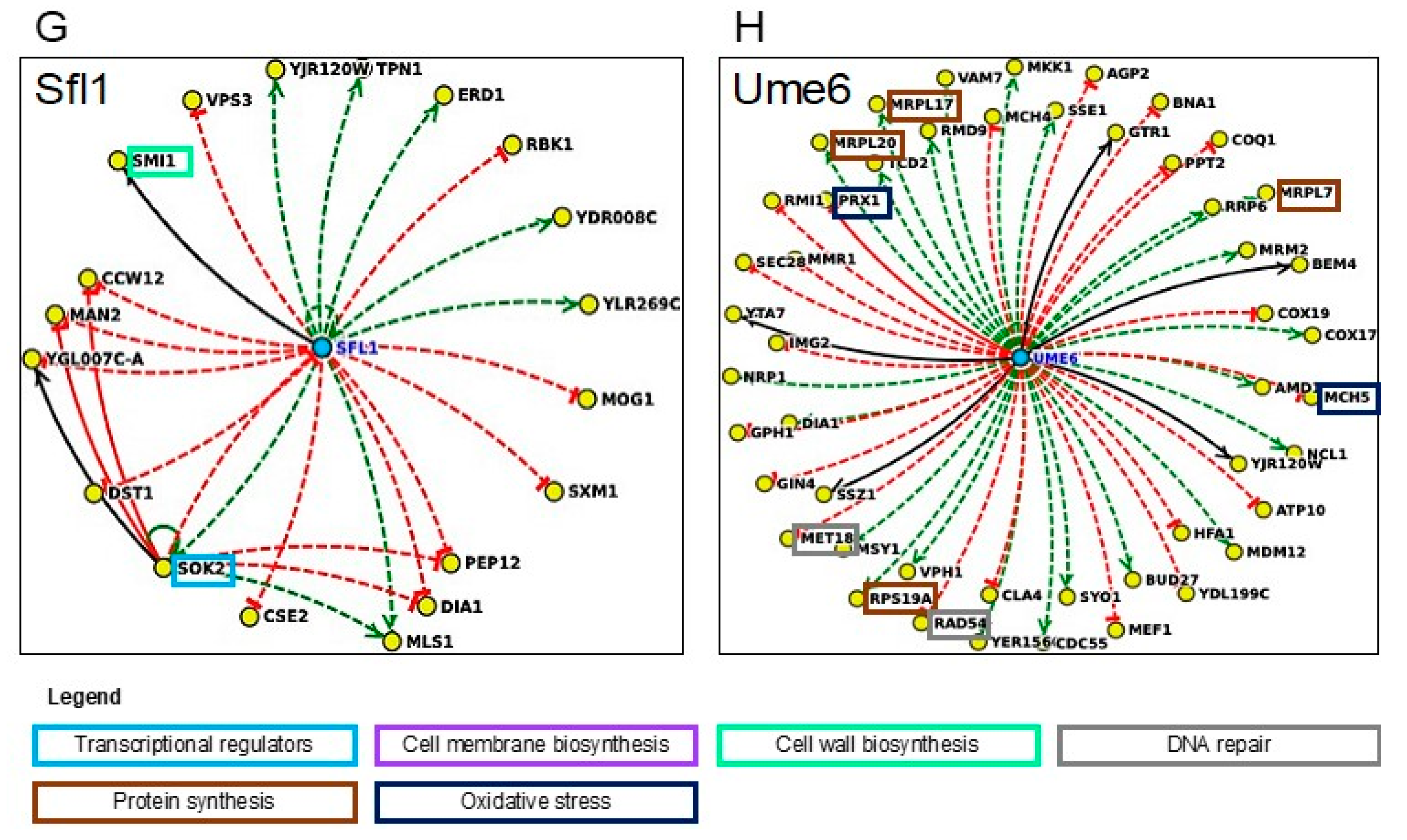
Concerning the putative regulatory networks involved in methanol tolerance, twelve TFs were identified in our chemogenomic analysis. Five of them are relevant to overcome the stress induced by either methanol or ethanol: Cbf1, Rpn4, Sfl1, Sfp1, and Ume6. Regarding Rpn4 and Sfp1, these TFs reportedly confer tolerance to a wide variety of environmental stresses [
64,
65,
66]. The regulation data available in Yeastract pointed out Rpn4 as a major regulator: 5834 genes of yeast genome are under Rpn4 control and above 90% of the alcohol tolerance genes present in both datasets are known to be under Rpn4 regulation. This TF was previously reported to be activated by ethanol shock [
67] or short exposure to this alcohol [
68]. Rpn4 stimulates the expression of proteasome subunit genes as well as of genes involved DNA repair [
66], which are enriched biological functions in the methanol dataset. The regulation of the DNA-damage response is also dependent on Sfp1 [
69], which was previously identified as an ethanol tolerance determinant [
22]. Cbf1 regulates around 80% of methanol and ethanol datasets, being the second major regulator of alcohol tolerance genes. The Yeastract tool “Rank by TF”, enabling automatic selection and ranking of transcription factors potentially involved in the regulation of the genes of methanol and ethanol datasets, attributed a significant p-value to Cbf1 and Rpn4 (<0.05 in both datasets) and for Sfp1 (<0.06 and 0.08, in methanol and ethanol datasets, respectively). These results emphasize the importance of these TFs in the regulation of methanol and ethanol tolerance genes and its potential as candidates for alcohol tolerance engineering. Although the genetic engineering of TFs by modulating their activity can lead to an imbalance in metabolic reactions, this strategy has been proposed as an important approach for improvement of yeast tolerance to several toxicants. Successful approaches using this strategy includes Haa1 engineering to increase
S cerevisiae acetic acid tolerance [
70] or the tolerance to a mixture of acetic acid and furfural by the increased co-expression of Haa1 and Tye7 [
71] or Ace2 and Sfp1 [
65]. To improve methanol and ethanol tolerance, Cbf1, Rpn4 and Sfp1 are promising molecular targets of genetic engineering. Recently, the heterologous expression of IrrE, a global regulatory protein from the prokaryotic organism
Deinococcus radiodurans, was engineered to improve yeast tolerance to inhibitors present in lignocellulose hydrolysates or to high temperatures [
72].
Concerning the TFs encoded by genes only found in the methanol dataset,
GLN3,
IXR1,
NGG1,
OPI1,
RPH1,
SOK2, and
STB5, they are involved in several bioprocesses, such as nitrogen catabolism [
73], oxygen sensing [
45], histone acetylation and demethylation [
74,
75], lipid biosynthesis [
76], autophagy [
77], pseudohyphal growth [
78], and oxidative stress response [
79].
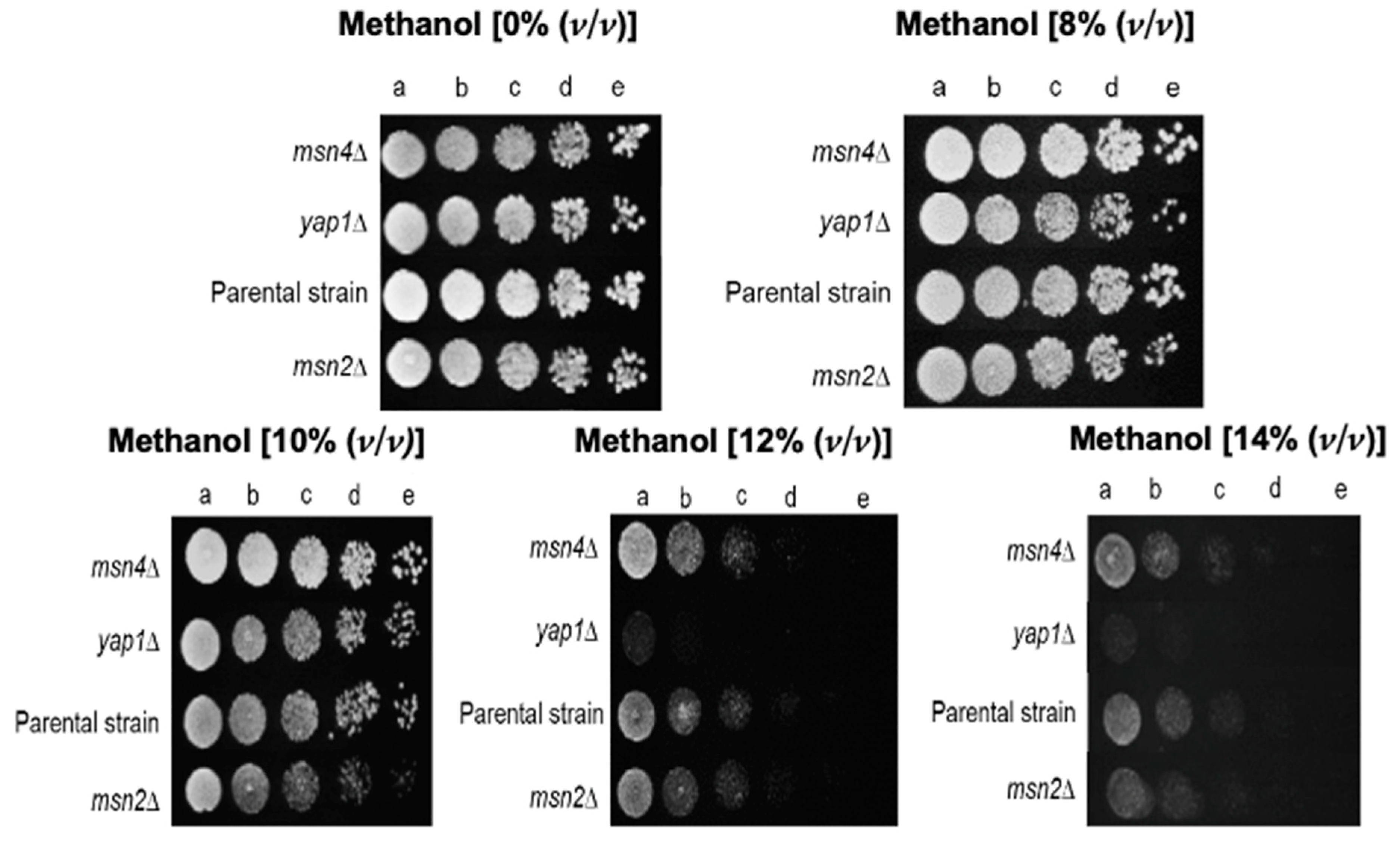
Ixr1 and Opi1 are suggested to play a significant role in methanol tolerance since the corresponding deletion mutants exhibited a marked methanol susceptibility phenotype. Ixr1 is a transcriptional repressor of hypoxic genes’ transcription during normoxia [
45] and in the corresponding deletion mutant,
ixr1Δ, the expression of genes related to ribosomal genes are downregulated [
44]. Ixr1 also controls the levels of dNTPs in the cell required for DNA synthesis and repair [
80]. Additionally,
IXR1 may be involved in the oxidative stress response, since the mutant
ixr1Δ is more susceptible to peroxides [
81] and its promoter region has a binding site for
STB5, a regulator of the oxidative stress response [
44]. Opi1 is a transcriptional repressor involved in the regulation of phospholipid synthesis in response to inositol availability [
76]. The deletion of the
OPI1 gene was found to jeopardize mitochondrial metabolism, by decreasing the levels of cardiolipin by 50%, resulting in low cytochrome content and high mitochondrial DNA instability [
82]. Our results suggest, for the first time, the importance of mitochondrial genome in methanol tolerance, similarly to what has been proposed for ethanol [
22,
83]. Given that
IXR1 is involved in the regulation of around 30% of the genes in the methanol dataset, including other transcription factors encoding genes (
OPI1,
RPH1,
SFP1,
SOK2,
STB5, and
UME6) [
42,
43,
44,
45] and the deletion mutant exhibits a strong methanol susceptibility phenotype, this regulator, together with Opi1 can be, in principle, considered promising candidate molecular targets to be tested for TF engineering in
S. cerevisiae.
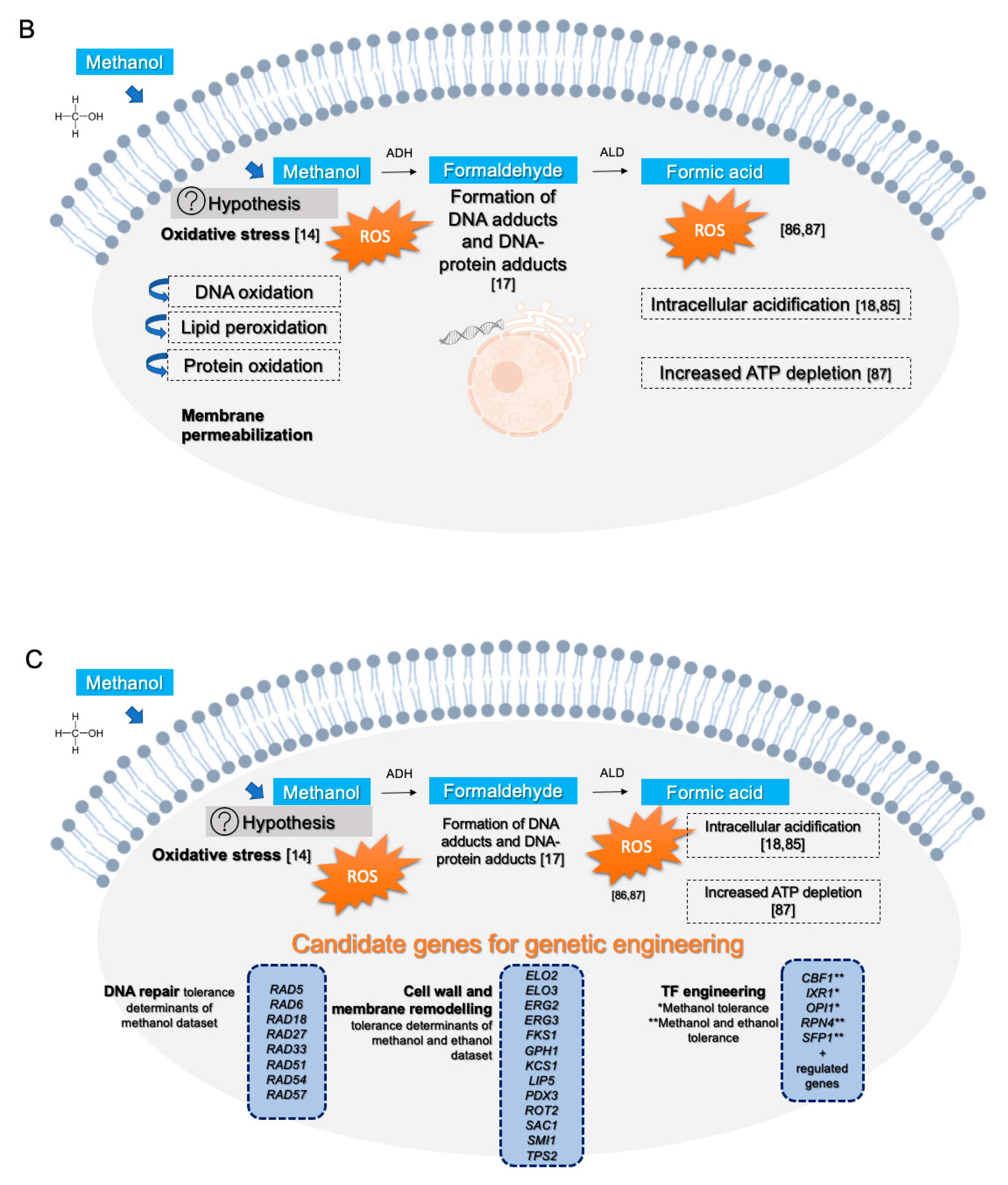
Methanol oxidation is accompanied by the production of free radicals in complex eukaryotes [
39] and, in yeast, oxidative stress was pointed out as a major consequence of exposure to methanol [
14]. Formic acid is also an oxidative stress inducer and, in mammalians, formate binds to cytochrome c, inhibiting the last step of the electron transport chain in mitochondria [
84] and to the disruption of the proton gradient and the consequent decrease of ATP synthesis [
85]. In yeast, formic acid can lead to the rapid burst of intracellular reactive oxygen species [
86] and, consequently, to oxidative stress [
87]. In addition to
IXR1 and
STB5, other oxidative stress responsive genes were found to be determinants of methanol tolerance, including genes encoding antioxidant enzymes such as superoxide dismutase (Sod1), methionine-S-sulfoxide reductase (Mrx2), peroxiredoxin (Prx1), as well as Gsh1, a gamma glutamylcysteine synthetase involved in the first step of glutathione biosynthesis. Additionally, two transporters, Fen2 and Mch5 involved in overcoming oxidative stress [
88,
89] are relevant due to the inability of the corresponding deletion mutants to grow in methanol. Fen2 is a H
+-pantothenate (vitamin B5) symporter [
90] and mutations in
FEN2 lead to reduced biosynthesis of ergosterol and fatty acids [
90], which emphasizes the importance of membrane composition and properties in methanol tolerance. In addition, the
Corynebacterium glutamicum Cgl0833, a Na+/panthothenate symporter, is required for methanol tolerance [
91]. Mch5 is a transporter from the major facilitator superfamily that proceeds to the uptake of riboflavin [
92], a vitamin that is required for the activity of glutathione reductase in the FAD coenzyme form [
88]. Although many important TFs for methanol tolerance were identified in this genome-wide study, the major regulator of the response to oxidative stress, Yap1 were not identified but, for higher levels of methanol stress,
YAP1 expression was confirmed as a critical methanol tolerance determinant.
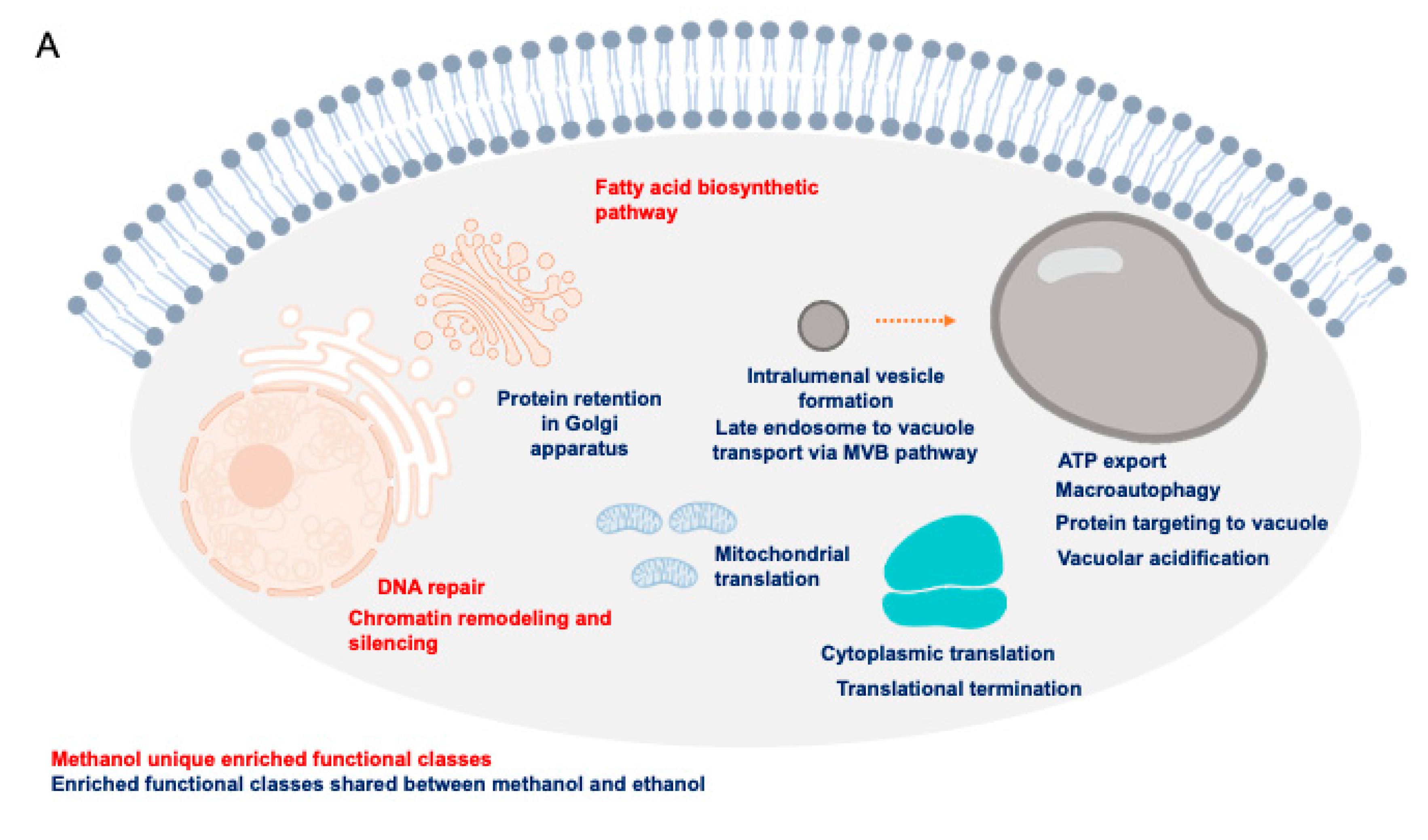
Despite the potential of methanol as a (co)substrate for the biotechnology industry, the global mechanisms by which this alcohol exerts its toxicity are still unclear. Based on the results from this study and on the above referred discussion, a schematic model on the hypothesized methanol toxicity and tolerance mechanisms are presented, specifying the more specific mechanisms and those shared with ethanol (
Figure 7A). The reported toxicity mechanisms of the detoxification intermediates formaldehyde and formic acid are also included (
Figure 7B).
Results from this genome-wide search for methanol tolerance genes can also be explored for the rational genomic manipulation of the yeast cell to obtain more robust strains capable of coping with high methanol or high methanol and ethanol concentrations by exploring information on a cumulative inhibitor tolerance phenotype, as recently reported for
S cerevisiae engineering towards improved tolerance to for the multiple inhibitors present in lignocellulosics-based fermentations [
93]. Since TFs engineering is an emerging, although somewhat controversial, strategy to increase yeast tolerance to different biotechnological relevant stresses [
65,
70,
71], it would be interesting to explore the modulation of the expression or the alteration of the amino acid sequence of the TFs here identified as promising (
Figure 7C). Among them are Cbf1, Rpn4 and Sfp1, for alcohol tolerance, and Ixr1 and Opi1, specifically for methanol tolerance. The knowledge here obtained, and the list of genes provided in this study can be considered an important starting point for the improvement of yeast tolerance to methanol or to methanol and ethanol in biotechnologically relevant yeast species for which the necessary genome sequence and editing tools are currently available or could be developed. The exploitation of the bioinformatics tool NCYeastract database (Non- Conventional Yeastract;
http://yeastract-plus.org/ncyeastract/) [
38] will facilitate the identification of orthologous genes in the yeast species currently included in the database as well as the regulatory associations already described for other yeast species, especially for
S. cerevisiae in the sister database Yeastract (
http://yeastract-plus.org/yeastract/scerevisiae/index.php), using the new tools for cross-species transcription regulation comparison [
38]. In particular, this is currently a useful resource to guide the genetic engineering of biotechnologically relevant yeasts, such as the methylotrophic yeast species
Komagataella phaffii and the oleaginous yeast species
Yarrowia lipolytica [
38].




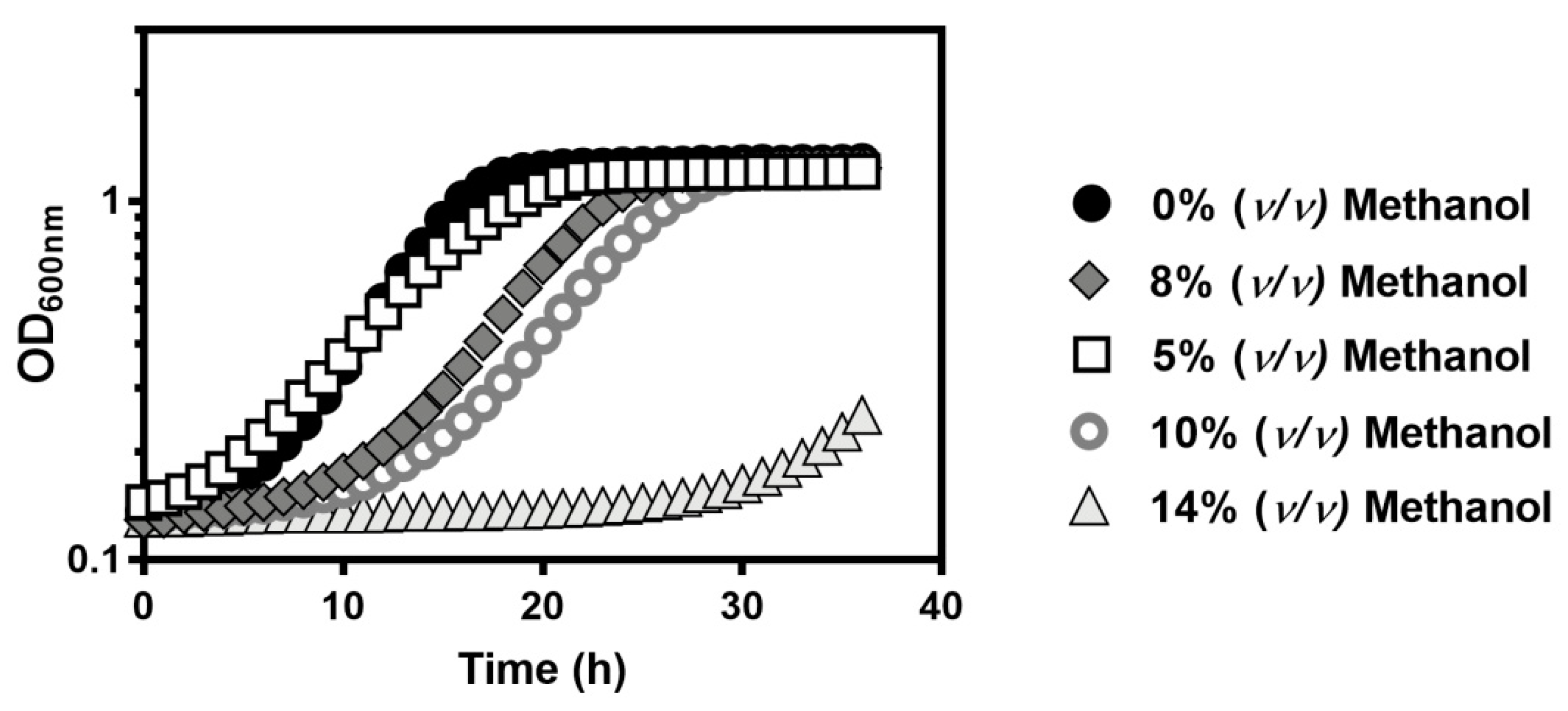
 ) with 5% (
) with 5% (  ), 8% (
), 8% (  ), 10% (
), 10% (  ), or 14% (v/v) (
), or 14% (v/v) (  ) of methanol. Growth was followed in a microplate reader by measuring the optical density at 595 nm of a 96-well plate incubated at 35 °C with orbital agitation, for 36 h.
) of methanol. Growth was followed in a microplate reader by measuring the optical density at 595 nm of a 96-well plate incubated at 35 °C with orbital agitation, for 36 h.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}