
Engineering Aspergillus oryzae for the Heterologous Expression of a Bacterial Modular Polyketide Synthase


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
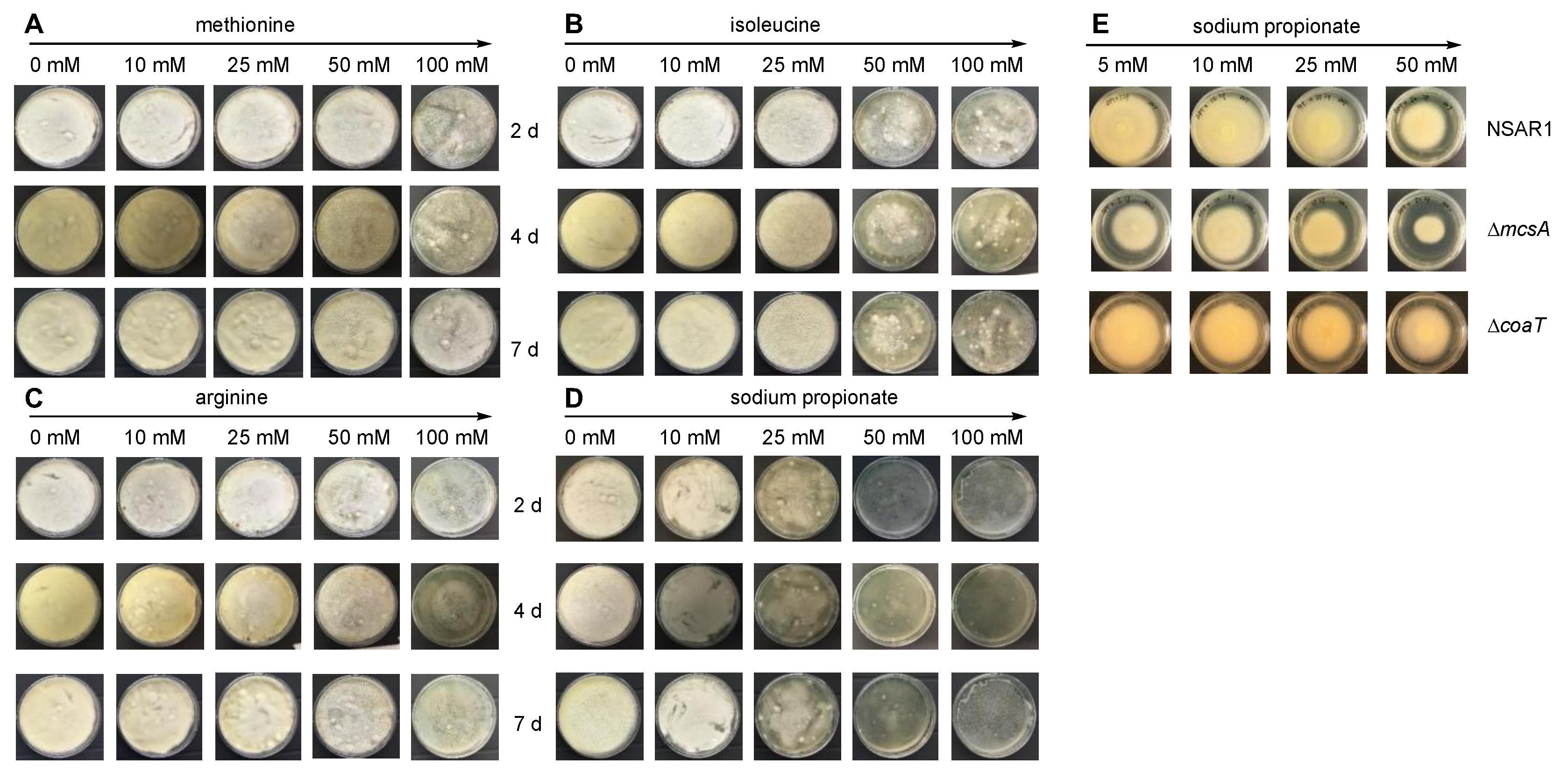
2.2. Propionyl-CoA Toxicity Assessment to A. oryzae NSAR1
2.3. PCR-Based Gene Identification
2.4. Construction of pTYGS·arg·pccABE
2.5. Construction of pTYGS·arg·debs1te·pccABE
2.6. PEG-Mediated Transformation of A. oryzae NSAR1
2.7. TKL Fermentation, Extraction and Analysis
2.8. TKL Quantification
2.9. Gene Knockout of Degradation Pathways
3. Results
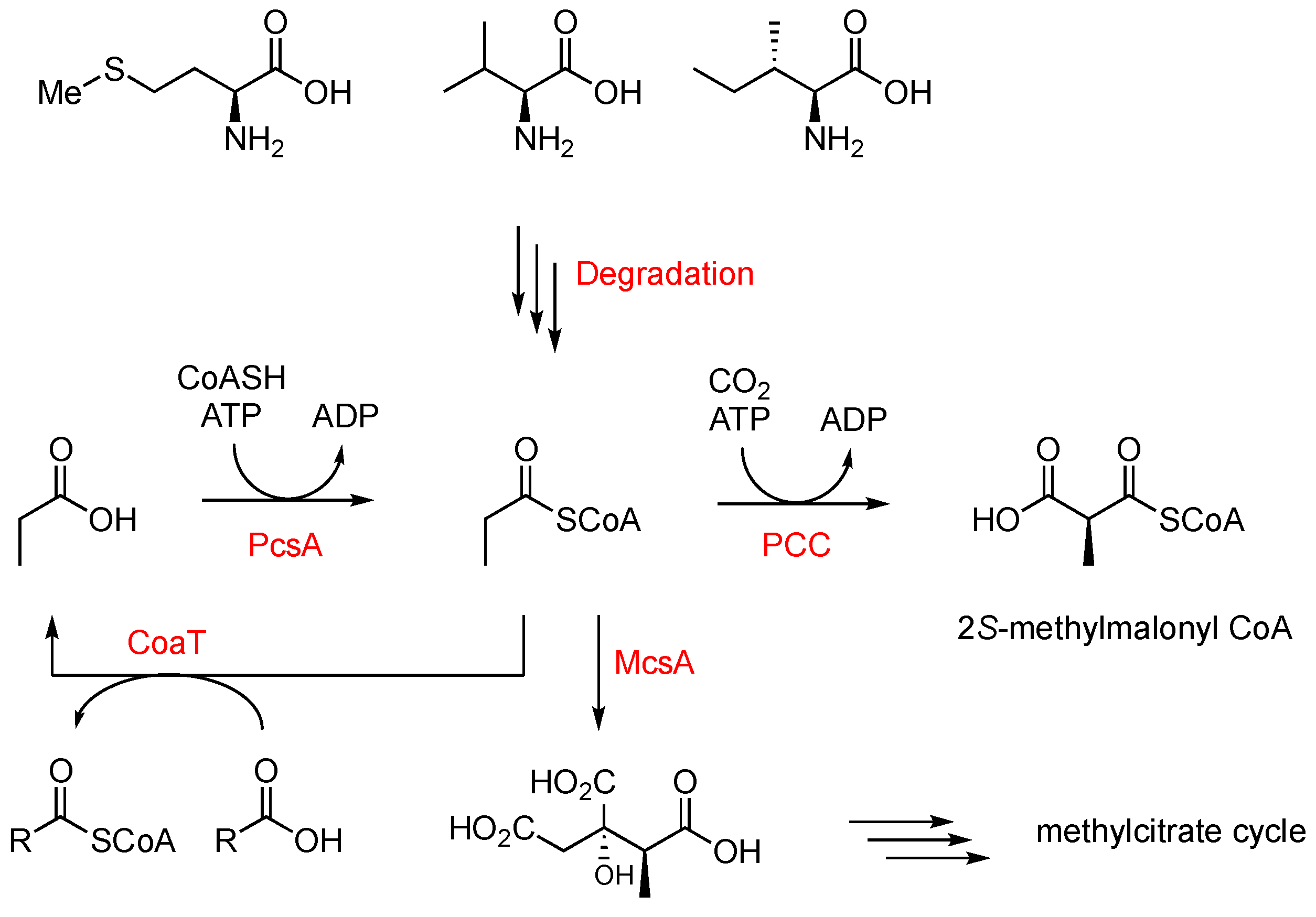
3.1. Investigating Propionyl-CoA Metabolism in A. oryzae
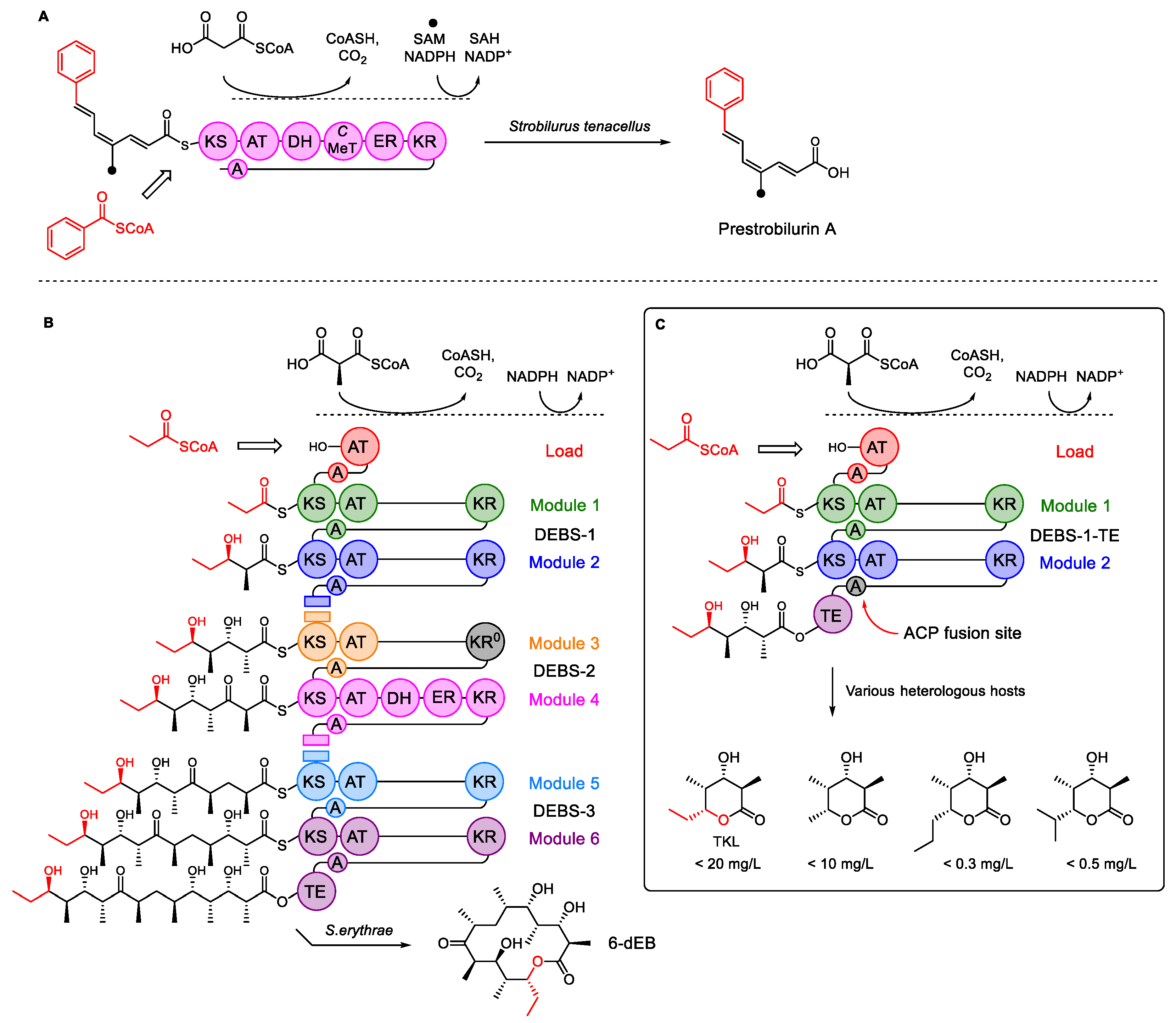
3.2. Introducing a Methylmalonyl-CoA Pathway into A. oryzae NSAR1
3.3. Heterologous Expression of DEBS1-TE in A. oryzae to form TKL
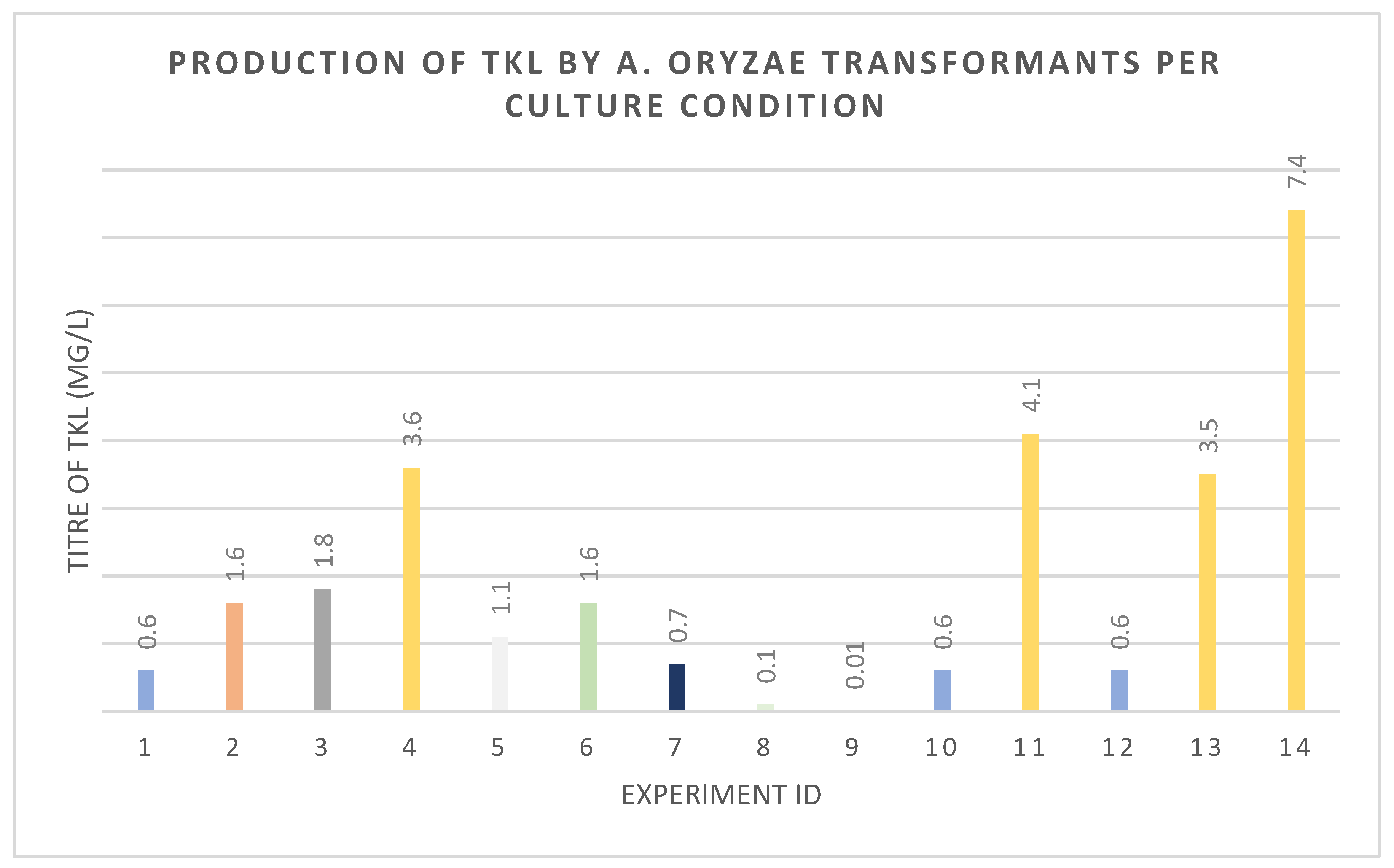
3.4. Improving Titers of TKL through Metabolic Engineering of A. oryzae Primary Metabolism and Sodium Propionate Feeding Methods
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Zhang, Q.; Deng, W.; Qian, J.; Zhang, S.; Liu, W. Toward Improvement of Erythromycin A Production in an IndustrialSaccharopolyspora erythraea Strain via Facilitation of Genetic Manipulation with an Artificial attB Site for Specific Recombination. Appl. Environ. Microbiol. 2011, 77, 7508–7516. [Google Scholar] [CrossRef] [Green Version]
- Miyazawa, T.; Fitzgerald, B.J.; Keatinge-Clay, A.T. Preparative production of an enantiomeric pair by engineered polyketide synthases. Chem. Commun. 2021, 57, 8762–8765. [Google Scholar] [CrossRef]
- Cook, T.B.; Pfleger, B.F. Leveraging synthetic biology for producing bioactive polyketides and non-ribosomal peptides in bacterial heterologous hosts. MedChemComm 2019, 10, 668–681. [Google Scholar] [CrossRef]
- Yang, D.; Park, S.Y.; Park, Y.S.; Eun, H.; Lee, S.Y. Metabolic Engineering of Escherichia coli for Natural Product Biosynthesis. Trends Biotechnol. 2020, 38, 745–765. [Google Scholar] [CrossRef]
- Bond, C.; Tang, Y.; Li, L. Saccharomyces cerevisiae as a tool for mining, studying and engineering fungal polyketide synthases. Fungal Genet. Biol. 2016, 89, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Harvey, C.J.B.; Tang, M.; Schlecht, U.; Horecka, J.; Fischer, C.R.; Lin, H.-C.; Li, J.; Naughton, B.; Cherry, J.; Miranda, M.; et al. HEx: A heterologous expression platform for the discovery of fungal natural products. Sci. Adv. 2018, 4, eaar5459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Liu, Y.; Du, G.; Ledesma-Amaro, R.; Liu, L. Microbial Chassis Development for Natural Product Biosynthesis. Trends Biotechnol. 2020, 38, 779–796. [Google Scholar] [CrossRef] [PubMed]
- Daba, G.M.; Mostafa, F.A.; Elkhateeb, W.A. The ancient koji mold (Aspergillus oryzae) as a modern biotechnological tool. Bioresour. Bioprocess. 2021, 8, 1–17. [Google Scholar] [CrossRef]
- He, Y.; Cox, R.J. The molecular steps of citrinin biosynthesis in fungi. Chem. Sci. 2016, 7, 2119–2127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, K.; Szwalbe, A.J.; Mulholland, N.P.; Vincent, J.L.; Bailey, A.M.; Willis, C.L.; Simpson, T.J.; Cox, R.J. Heterologous Production of Fungal Maleidrides Reveals the Cryptic Cyclization Involved in their Biosynthesis. Angew. Chem. Int. Ed. 2016, 55, 6784–6788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahlert, L.; Bassiony, E.F.; Cox, R.; Skellam, E.J. Diels–Alder Reactions during the Biosynthesis of Sorbicillinoids. Angew. Chem. Int. Ed. 2020, 59, 5816–5822. [Google Scholar] [CrossRef] [Green Version]
- Kahlert, L.; Villanueva, M.; Cox, R.J.; Skellam, E.J. Biosynthesis of 6-Hydroxymellein Requires a Collaborating Polyketide Synthase-like Enzyme. Angew. Chem. Int. Ed. 2021, 60, 11423–11429. [Google Scholar] [CrossRef]
- Feng, J.; Surup, F.; Hauser, M.; Miller, A.; Wennrich, J.-P.; Stadler, M.; Cox, R.J.; Kuhnert, E. Biosynthesis of oxygenated brasilane terpene glycosides involves a promiscuous N-acetylglucosamine transferase. Chem. Commun. 2020, 56, 12419–12422. [Google Scholar] [CrossRef]
- Schor, R.; Schotte, C.; Wibberg, D.; Kalinowski, J.; Cox, R.J. Three previously unrecognised classes of biosynthetic enzymes revealed during the production of xenovulene A. Nat. Commun. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nofiani, R.; De Mattos-Shipley, K.; Lebe, K.E.; Han, L.-C.; Iqbal, Z.; Bailey, A.; Willis, C.L.; Simpson, T.J.; Cox, R.J. Strobilurin biosynthesis in Basidiomycete fungi. Nat. Commun. 2018, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kahlert, L.; Schotte, C.; Cox, R.J. Total Mycosynthesis: Rational Bioconstruction and Bioengineering of Fungal Natural Products. Synthesis 2021, 53, 2381–2394. [Google Scholar] [CrossRef]
- Alberti, F.; Khairudin, K.; Venegas, E.R.; Davies, J.; Hayes, P.M.; Willis, C.L.; Bailey, A.M.; Foster, G.D. Heterologous expression reveals the biosynthesis of the antibiotic pleuromutilin and generates bioactive semi-synthetic derivatives. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Bailey, A.; Alberti, F.; Kilaru, S.; Collins, C.; De Mattos-Shipley, K.; Hartley, A.J.; Hayes, P.; Griffin, A.; Lazarus, C.M.; Cox, R.; et al. Identification and manipulation of the pleuromutilin gene cluster from Clitopilus passeckerianus for increased rapid antibiotic production. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rendsvig, J.K.H.; Futyma, M.E.; Jarczynska, Z.D.; Mortensen, U.H. Filamentous Fungi as Hosts for Heterologous Production of Proteins and Secondary Metabolites in the Post-Genomic Era. In Genetics and Biotechnology; The Mycota (A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research); Benz, J.P., Schipper, K., Eds.; Springer: Cham, Switzerland, 2020; Volume 2, pp. 227–265. [Google Scholar]
- Thynne, E.; Mead, O.; Chooi, Y.-H.; McDonald, M.C.; Solomon, P.S. Acquisition and Loss of Secondary Metabolites Shaped the Evolutionary Path of Three Emerging Phytopathogens of Wheat. Genome Biol. Evol. 2019, 11, 890–905. [Google Scholar] [CrossRef]
- Cox, R. Oxidative rearrangements during fungal biosynthesis. Nat. Prod. Rep. 2014, 31, 1405–1424. [Google Scholar] [CrossRef] [Green Version]
- Cortes, J.; Wiesmann, K.E.H.; Roberts, G.A.; Brown, M.J.B.; Staunton, J.; Leadlay, P.F. Repositioning of a Domain in a Modular Polyketide Synthase to Promote Specific Chain Cleavage. Science 1995, 268, 1487–1489. [Google Scholar] [CrossRef]
- Pfeifer, B.A.; Admiraal, S.J.; Gramajo, H.; Cane, D.E.; Khosla, C. Biosynthesis of Complex Polyketides in a Metabolically Engineered Strain of E. coli. Science 2001, 291, 1790–1792. [Google Scholar] [CrossRef]
- Kao, C.M.; Luo, G.; Katz, L.; Cane, D.E.; Khosla, C. Manipulation of macrolide ring size by directed mutagenesis of a modular polyketide synthase. J. Am. Chem. Soc. 1995, 117, 9105–9106. [Google Scholar] [CrossRef]
- Kim, B.S.; Cropp, T.A.; Florova, G.; Lindsay, Y.; Sherman, D.H.; Reynolds, K.A. An Unexpected Interaction between the Modular Polyketide Synthases, Erythromycin DEBS1 and Pikromycin PikAIV, Leads to Efficient Triketide Lactone Synthesis. Biochemistry 2002, 41, 10827–10833. [Google Scholar] [CrossRef] [PubMed]
- Pieper, R.; Luo, G.; Cane, D.E.; Khosla, C. Cell-free synthesis of polyketides by recombinant erythromycin polyketide synthases. Nat. Cell Biol. 1995, 378, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Brock, M.; Buckel, W. On the mechanism of action of the antifungal agent propionate. JBIC J. Biol. Inorg. Chem. 2004, 271, 3227–3241. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-Q.; Keller, N.P. Blockage of methylcitrate cycle inhibits polyketide production in Aspergillus nidulans. Mol. Microbiol. 2004, 52, 541–550. [Google Scholar] [CrossRef]
- Lazarus, C.M.; Williams, K.; Bailey, A.M. Reconstructing fungal natural product biosynthetic pathways. Nat. Prod. Rep. 2014, 31, 1339–1347. [Google Scholar] [CrossRef]
- Zhang, Y.-Q.; Brock, M.; Keller, N.P. Connection of Propionyl-CoA Metabolism to Polyketide Biosynthesis in Aspergillus nidulans. Genetics 2004, 168, 785–794. [Google Scholar] [CrossRef] [Green Version]
- Brock, M.; Fischer, R.; Linder, D.; Buckel, W. Methylcitrate synthase from Aspergillus nidulans: Implications for propionate as an antifungal agent. Mol. Microbiol. 2000, 35, 961–973. [Google Scholar] [CrossRef] [PubMed]
- Fleck, C.B.; Brock, M. Characterization of an acyl-CoA: Carboxylate CoA-transferase from Aspergillus nidulans involved in propionyl-CoA detoxification. Mol. Microbiol. 2008, 68, 642–656. [Google Scholar] [CrossRef]
- Diacovich, L.; Peirú, S.; Kurth, D.; Rodríguez, E.; Podestá, F.; Khosla, C.; Gramajo, H. Kinetic and Structural Analysis of a New Group of Acyl-CoA Carboxylases Found in Streptomyces coelicolor A3 (2). J. Biol. Chem. 2002, 277, 31228–31236. [Google Scholar] [CrossRef] [Green Version]
- Vandova, G.A.; O’Brien, R.V.; Lowry, B.; Robbins, T.F.; Fischer, C.R.; Davis, R.W.; Khosla, C.; Harvey, C.J.B.; Hillenmeyer, M.E. Heterologous expression of diverse propionyl-CoA carboxylases affects polyketide production in Escherichia coli. J. Antibiot. 2017, 70, 859–863. [Google Scholar] [CrossRef] [Green Version]
- Pahirulzaman, K.A.K.; Williams, K.; Lazarus, C.M. A Toolkit for Heterologous Expression of Metabolic Pathways in Aspergillus oryzae. Methods Enzymol. 2012, 517, 241–260. [Google Scholar] [CrossRef] [PubMed]
- Lambalot, R.H.; Gehring, A.M.; Flugel, R.S.; Zuber, P.; LaCelle, M.; Marahiel, M.A.; Reid, R.; Khosla, C.; Walsh, C.T. A new enzyme superfamily—the phosphopantetheinyl transferases. Chem. Biol. 1996, 3, 923–936. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Komatsu, M.; Shin-ya, K.; Omura, S.; Ikeda, H. Distribution and functional analysis of the phosphopantetheinyl transferase superfamily in Actinomycetales microorganisms. Proc. Nat. Acad. Sci. USA 2018, 115, 6828–6833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutka, S.C.; Bondi, S.M.; Carney, J.R.; Da Silva, N.A.; Kealey, J.T. Metabolic pathway engineering for complex polyketide biosynthesis inSaccharomyces cerevisiae. FEMS Yeast Res. 2006, 6, 40–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissman, K.J.; Hong, H.; Oliynyk, M.; Siskos, A.P.; Leadlay, P.F. Identification of a Phosphopantetheinyl Transferase for Erythromycin Biosynthesis in Saccharopolyspora erythraea. ChemBioChem 2004, 5, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Brock, M. Role of Cellular Control of Propionyl-CoA Levels for Microbial Pathogenesis. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 3279–3291. [Google Scholar]
- Nielsen, M.L.; Albertsen, L.; Lettier, G.; Nielsen, J.B.; Mortensen, U.H. Efficient PCR-based gene targeting with a recyclable marker for Aspergillus nidulans. Fungal Genet. Biol. 2006, 43, 54–64. [Google Scholar] [CrossRef]
- Tokuoka, M.; Tanaka, M.; Ono, K.; Takagi, S.; Shintani, T.; Gomi, K. Codon Optimization Increases Steady-State mRNA Levels in Aspergillus oryzae Heterologous Gene Expression. Appl. Environ. Microbiol. 2008, 74, 6538–6546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, R.J.; Hitchman, T.S.; Byrom, K.J.; Findlow, I.C.; A Tanner, J.; Crosby, J.; Simpson, T.J. Post-translational modification of heterologously expressed Streptomyces type II polyketide synthase acyl carrier proteins. FEBS Lett. 1997, 405, 267–272. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Garcia, R.A.; Nielsen, L.K.; Marcellin, E. Heterologous Production of 6-Deoxyerythronolide B in Escherichia coli through the Wood Werkman Cycle. Metabolites 2020, 10, 228. [Google Scholar] [CrossRef]
- Zhang, H.; Boghigian, B.A.; Pfeifer, B.A. Investigating the role of native propionyl-CoA and methylmalonyl-CoA metabolism on heterologous polyketide production in Escherichia coli. Biotech. Bioeng. 2010, 105, 567–573. [Google Scholar] [CrossRef]
- Maerker, C.; Rohde, M.; Brakhage, A.A.; Brock, M. Methylcitrate synthase from Aspergillus fumigatus. Propionyl-CoA affects polyketide synthesis, growth and morphology of conidia. FEBS J. 2005, 272, 3615–3630. [Google Scholar] [CrossRef]
- Domin, N.; Wilson, D.; Brock, M. Methylcitrate cycle activation during adaptation of Fusarium solani and Fusarium verticillioides to propionyl-CoA-generating carbon sources. Microbiology 2009, 155, 3903–3912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubey, M.K.; Broberg, A.; Jensen, D.; Karlsson, M. Role of the methylcitrate cycle in growth, antagonism and induction of systemic defence responses in the fungal biocontrol agent Trichoderma atroviride. Microbiology 2013, 159, 2492–2500. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-H.; Han, Y.-K.; Yun, S.-H.; Lee, Y.-W. Roles of the Glyoxylate and Methylcitrate Cycles in Sexual Development and Virulence in the Cereal Pathogen Gibberella zeae. Eukaryot. Cell 2009, 8, 1155–1164. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Wang, H.; Zhu, S.; Wang, J.; Liu, X.; Lin, F.; Lu, J. The Methylcitrate Cycle is Required for Development and Virulence in the Rice Blast Fungus Pyricularia oryzae. Mol. Plant-Microbe Interact. 2019, 32, 1148–1161. [Google Scholar] [CrossRef]
- Santos, L.P.A.; Assunção, L.D.P.; Lima, P.D.S.; Tristão, G.B.; Brock, M.; Borges, C.L.; Silva-Bailao, M.; Soares, C.M.D.A.; Bailao, A. Propionate metabolism in a human pathogenic fungus: Proteomic and biochemical analyses. IMA Fungus 2020, 11, 1–16. [Google Scholar] [CrossRef]
- Catenazzi, M.C.E.; Jones, H.; Wallace, I.; Clifton, J.; Chong, J.P.J.; Jackson, M.A.; Macdonald, S.; Edwards, J.; Moir, J.W.B. A large genomic island allows Neisseria meningitidis to utilize propionic acid, with implications for colonization of the human nasopharynx. Mol. Microbiol. 2014, 93, 346–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otzen, C.; Bardl, B.; Jacobsen, I.D.; Nett, M.; Brock, M. Candida albicans utilizes a modified β-oxidation pathway for the degradation of toxic propionyl-CoA. J. Biol. Chem. 2014, 289, 8151–8169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dayem, L.C.; Carney, J.R.; Santi, D.V.; Pfeifer, B.A.; Khosla, C.; Kealey, J.T. Metabolic engineering of a methylmalonyl-CoA mutase-epimerase pathway for complex polyketide biosynthesis in Escherichia coli. Biochemistry 2002, 41, 5193–5201. [Google Scholar] [CrossRef]
- De Mattos-Shipley, K.M.J.; Greco, C.; Heard, D.M.; Hough, G.; Mulholland, N.P.; Vincent, J.L.; Micklefield, J.; Simpson, T.J.; Willis, C.L.; Cox, R.J.; et al. The cycloaspeptides: Uncovering a new model for methylated nonribosomal peptide biosynthesis. Chem. Sci. 2018, 9, 4109–4117. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.-L.; Friedrich, S.; Yin, S.; Piech, O.; Williams, K.; Simpson, T.J.; Cox, R.J. Molecular basis of methylation and chain-length programming in a fungal iterative highly reducing polyketide synthase. Chem. Sci. 2019, 10, 8478–8489. [Google Scholar] [CrossRef] [PubMed]
- Márquez-Fernández, O.; Trigos, A.; Ramos-Balderas, J.L.; Viniegra-González, G.; Deising, H.B.; Aguirre, J. Phosphopantetheinyl Transferase CfwA/NpgA Is Required for Aspergillus nidulans Secondary Metabolism and Asexual Development. Eukaryot. Cell 2007, 6, 710–720. [Google Scholar] [CrossRef] [Green Version]
- Zainudin, N.A.I.M.; Condon, B.; De Bruyne, L.; Van Poucke, C.; Bi, Q.; Li, W.; Höfte, M.; Turgeon, B.G. Virulence, Host-Selective Toxin Production, and Development of Three Cochliobolus Phytopathogens Lacking the Sfp-Type 4′-Phosphopantetheinyl Transferase Ppt1. Mol. Plant-Microbe Interact. 2015, 28, 1130–1141. [Google Scholar] [CrossRef] [Green Version]
- Albermann, S.; Elter, T.; Teubner, A.; Krischke, W.; Hirth, T.; Tudzynski, B. Characterization of novel mutants with an altered gibberellin spectrum in comparison to different wild-type strains of Fusarium fujikuroi. Appl. Microbiol. Biotechnol. 2013, 97, 7779–7790. [Google Scholar] [CrossRef]
- Leng, Y.; Zhong, S. Sfp-type 4′-phosphopantetheinyl transferase is required for lysine synthesis, tolerance to oxidative stress and virulence in the plant pathogenic fungus Cochliobolus sativus. Mol. Plant Pathol. 2012, 13, 375–387. [Google Scholar] [CrossRef]
- Velazquez-Robledo, R.; Contreras-Cornejo, H.A.; Macias-Rodriguez, L.; Hernandez-Morales, A.; Aguirre, J.; Casa-Flores, S.; Lopez-Bucio, J.; Herrera-Estrella, A. Role of the 4-Phosphopantetheinyl Transferase of Trichoderma virens in Secondary Metabolism and Induction of Plant Defense Responses. MPMI 2011, 12, 1459–1471. [Google Scholar] [CrossRef] [Green Version]
- Horbach, R.; Graf, A.; Weihmann, F.; Antelo, L.; Mathea, S.; Liermann, J.C.; Opatz, T.; Thines, E.; Aguirre, J.; Deising, H.B. Sfp-Type 4′-Phosphopantetheinyl Transferase Is Indispensable for Fungal Pathogenicity. Plant Cell 2009, 21, 3379–3396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenke-Kodama, H.; Sandmann, A.; Müller, R.; Dittmann, E. Evolutionary Implications of Bacterial Polyketide Synthases. Mol. Biol. Evol. 2005, 22, 2027–2039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertrand, R.L.; Sorensen, J.L. Lost in Translation: Challenges with Heterologous Expression of Lichen Polyketide Synthases. Chem. Sel. 2019, 4, 6473–6483. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time after Inoculation (Days)/Sodium Propionate Inoculation Concentration (mM) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Condition | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 |
| A | - | - | - | 50 | Extract | ||||||
| B | 12.5 | 12.5 | 12.5 | 12.5 | Extract | ||||||
| C | 50 mM infusion with pump | Extract | |||||||||
| D | 10 | 10 | 10 | 10 | 10 | Extract | |||||
| E | 50 mM infusion with pump | Extract | |||||||||
| F | 6.25 | 6.25 | 6.25 | 6.25 | 6.25 | 6.25 | 6.25 | 6.25 | Extract | ||
| G | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | Extract |
| H | 2 | 2 | 2 | 2 | 2 | Extract | |||||
| I | 10 mM infusion with pump | Extract | |||||||||
| Experiment ID | A. oryzae Host | Expression Vectors | Condition |
|---|---|---|---|
| 1 | NSAR1 | pTYGS-arg-pccABE-debs1te-egfp + pTYGS-sC-sepptII | A |
| 2 | B | ||
| 3 | C | ||
| 4 | D | ||
| 5 | E | ||
| 6 | F | ||
| 7 | G | ||
| 8 | H | ||
| 9 | I | ||
| 10 | NSAR1 | pTYGS-arg-pccABE-debs1te + pTYGS-sC-sepptII | A |
| 11 | D | ||
| 12 | NSAR1 | pTYGS-arg-pccABE-debs1te-egfp + pTYGS-sC-sepptII-pcsA | A |
| 13 | D | ||
| 14 | NSAR1 ΔmcsA | pTYGS-arg-pccABE-debs1te-egfp + pTYGS-sC-sepptII | D |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, J.; Hauser, M.; Cox, R.J.; Skellam, E. Engineering Aspergillus oryzae for the Heterologous Expression of a Bacterial Modular Polyketide Synthase. J. Fungi 2021, 7, 1085. https://doi.org/10.3390/jof7121085
Feng J, Hauser M, Cox RJ, Skellam E. Engineering Aspergillus oryzae for the Heterologous Expression of a Bacterial Modular Polyketide Synthase. Journal of Fungi. 2021; 7(12):1085. https://doi.org/10.3390/jof7121085
Chicago/Turabian StyleFeng, Jin, Maurice Hauser, Russell J. Cox, and Elizabeth Skellam. 2021. "Engineering Aspergillus oryzae for the Heterologous Expression of a Bacterial Modular Polyketide Synthase" Journal of Fungi 7, no. 12: 1085. https://doi.org/10.3390/jof7121085