
Combination of Spirulina platensis, Ganoderma lucidum and Moringa oleifera Improves Cardiac Functions and Reduces Pro-Inflammatory Biomarkers in Preclinical Models of Short-Term Doxorubicin-Mediated Cardiotoxicity: New Frontiers in Cardioncology?
 ,
,  , , ,
, , ,  , ,
, ,  , and
, and 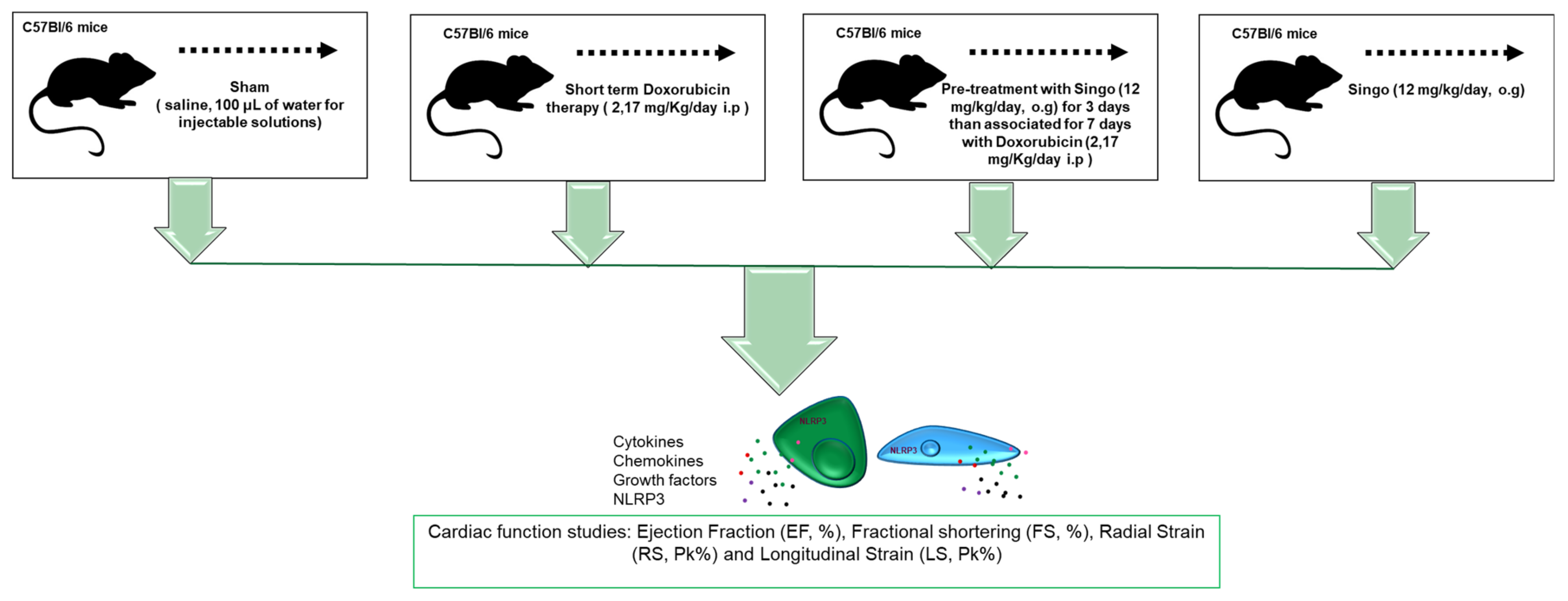{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
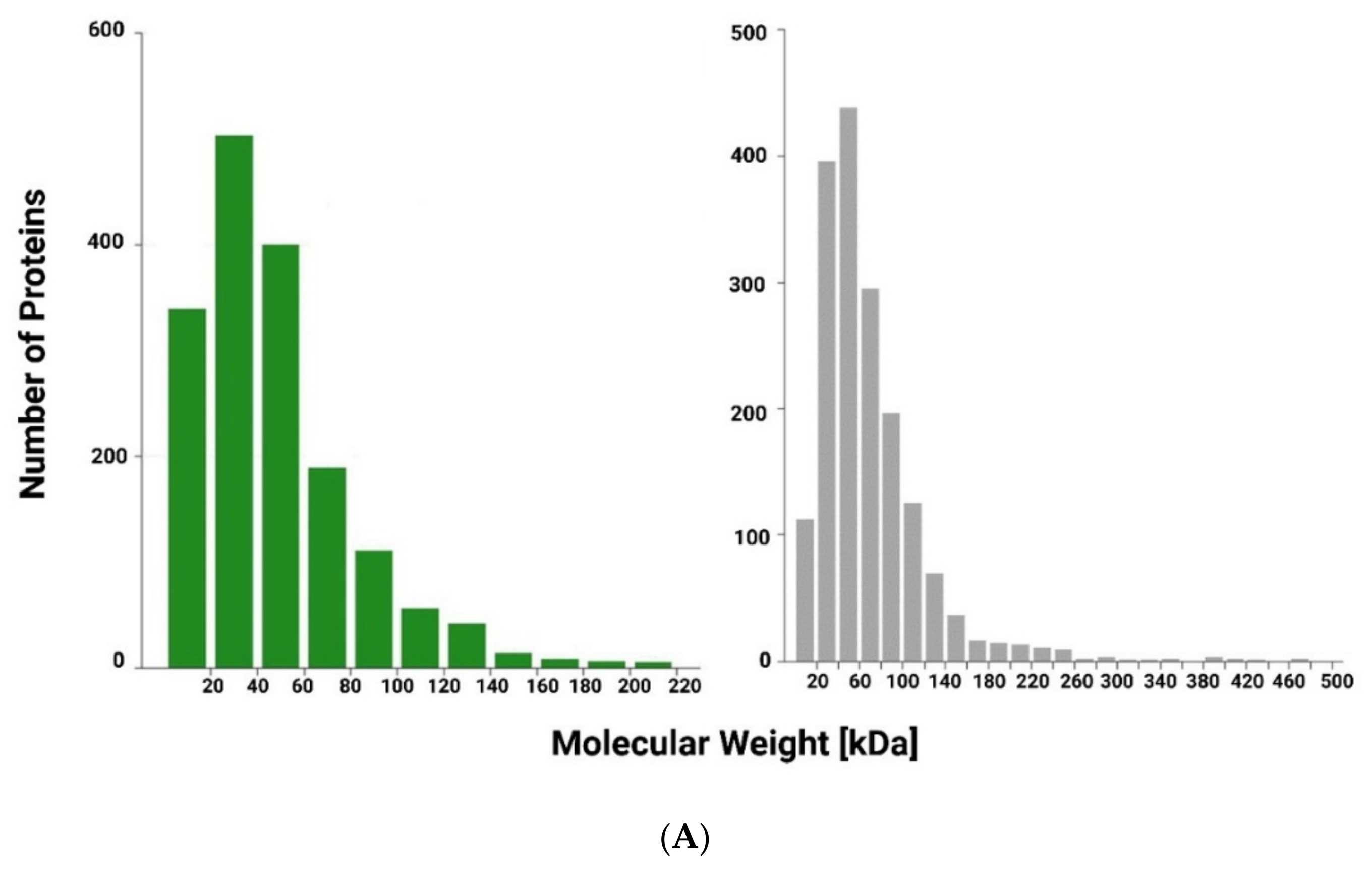
2.1. Protein Digestion of Spirulina platensis and Moringa oleifera Extracts
2.1.1. Protein Content Determination
2.1.2. Derivatization and Digestion
2.1.3. Desalting
2.1.4. Peptide Separation
2.2. Mass Spectrometry and Protein Analysis
2.3. Determination of Total Polysaccharides Content in Aqueous Extract of G. Lucidum
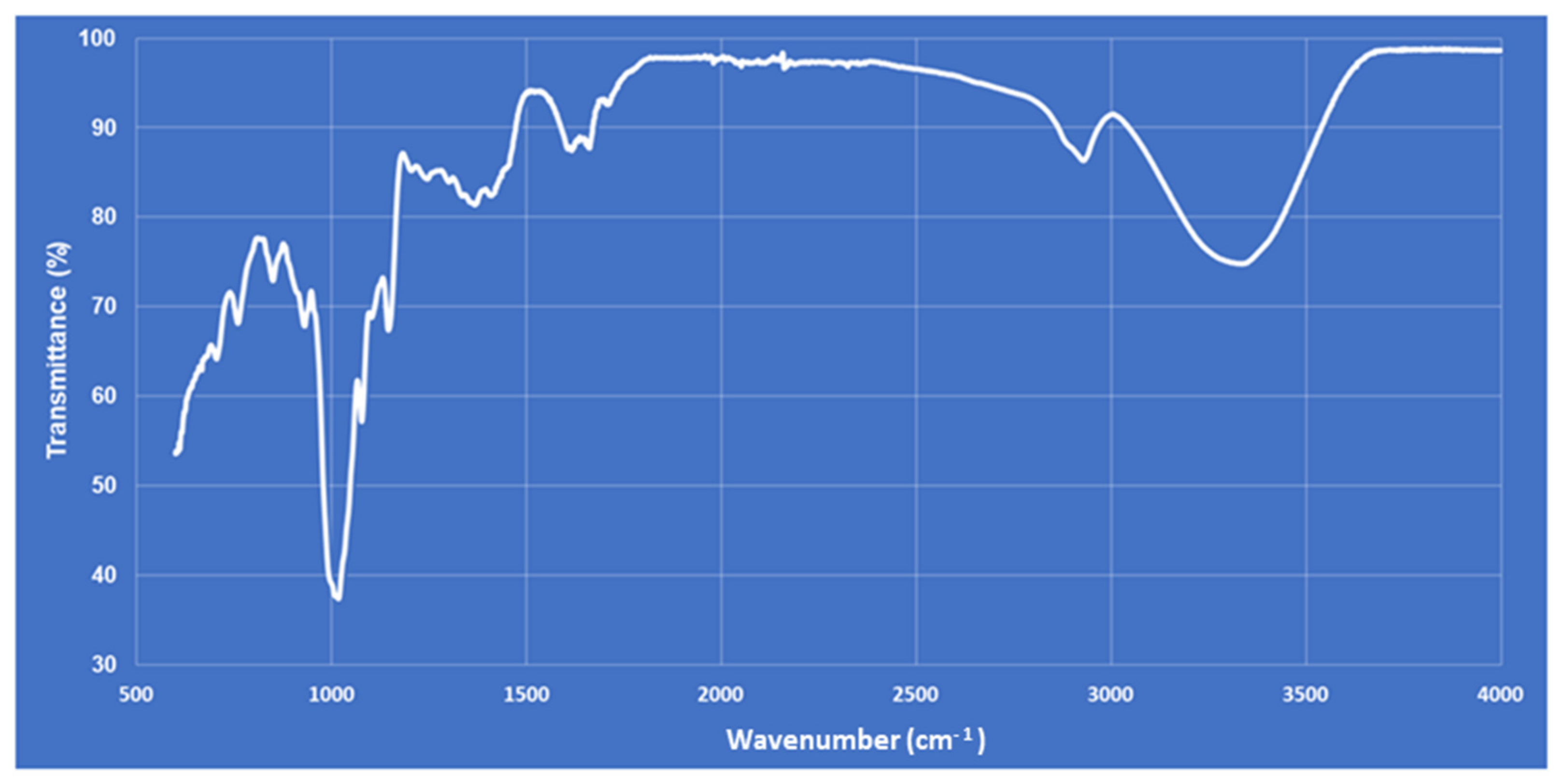
2.4. Polysaccharide Extraction and Fourier Transform Infrared (FT-IR) Characterization
2.5. Animal Studies
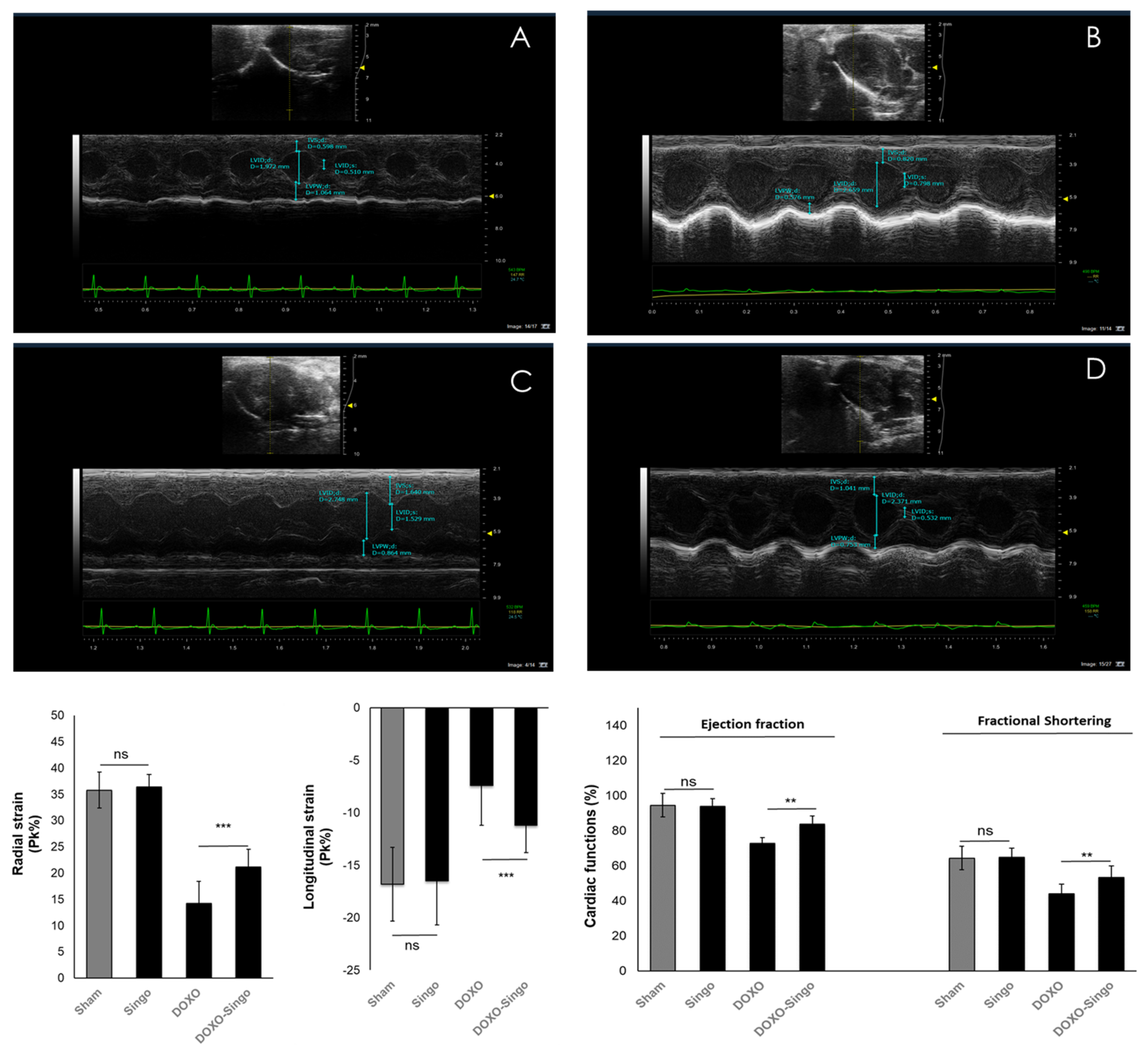
2.5.1. Echocardiographic Evaluation of Ventricular Functions
2.5.2. Cardiac Inflammation and DAMPs
2.5.3. Haematoxylin and Eosin (H&E) and Masson’s Trichrome Staining in Myocardial Tissues
2.5.4. Terminal dUTP Nick End-Labelling (TUNEL) Assay
2.6. Cell Studies: Cell Viability
2.6.1. Lipid Peroxidation
2.6.2. p65/NF-kB Expression
2.6.3. NLRP3 Expression
2.6.4. Intracellular Cytokines
2.7. Statistics
3. Results
3.1. Chemical and Physical Analysis of Plant Extract
3.2. Cardiac Function Studies
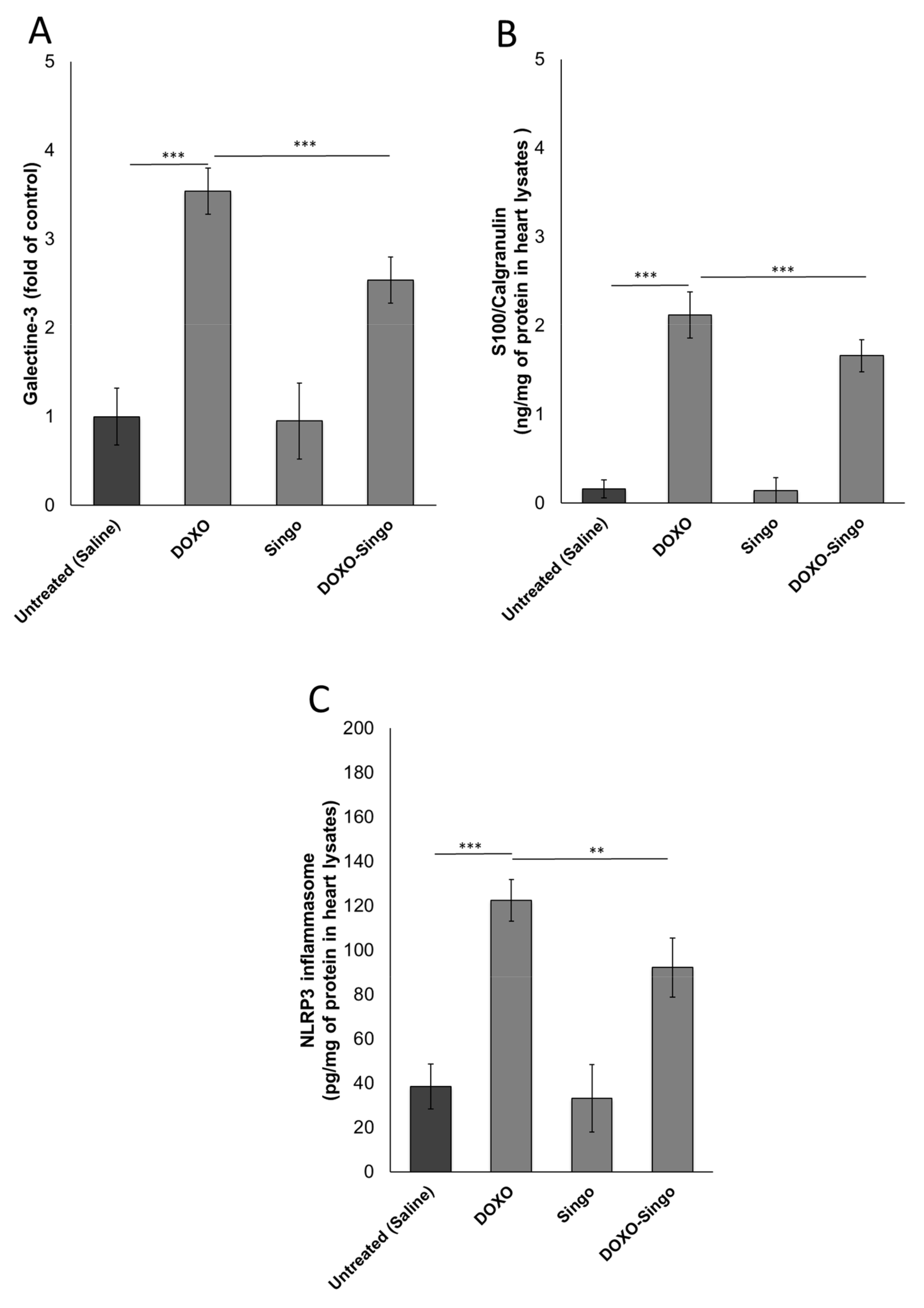
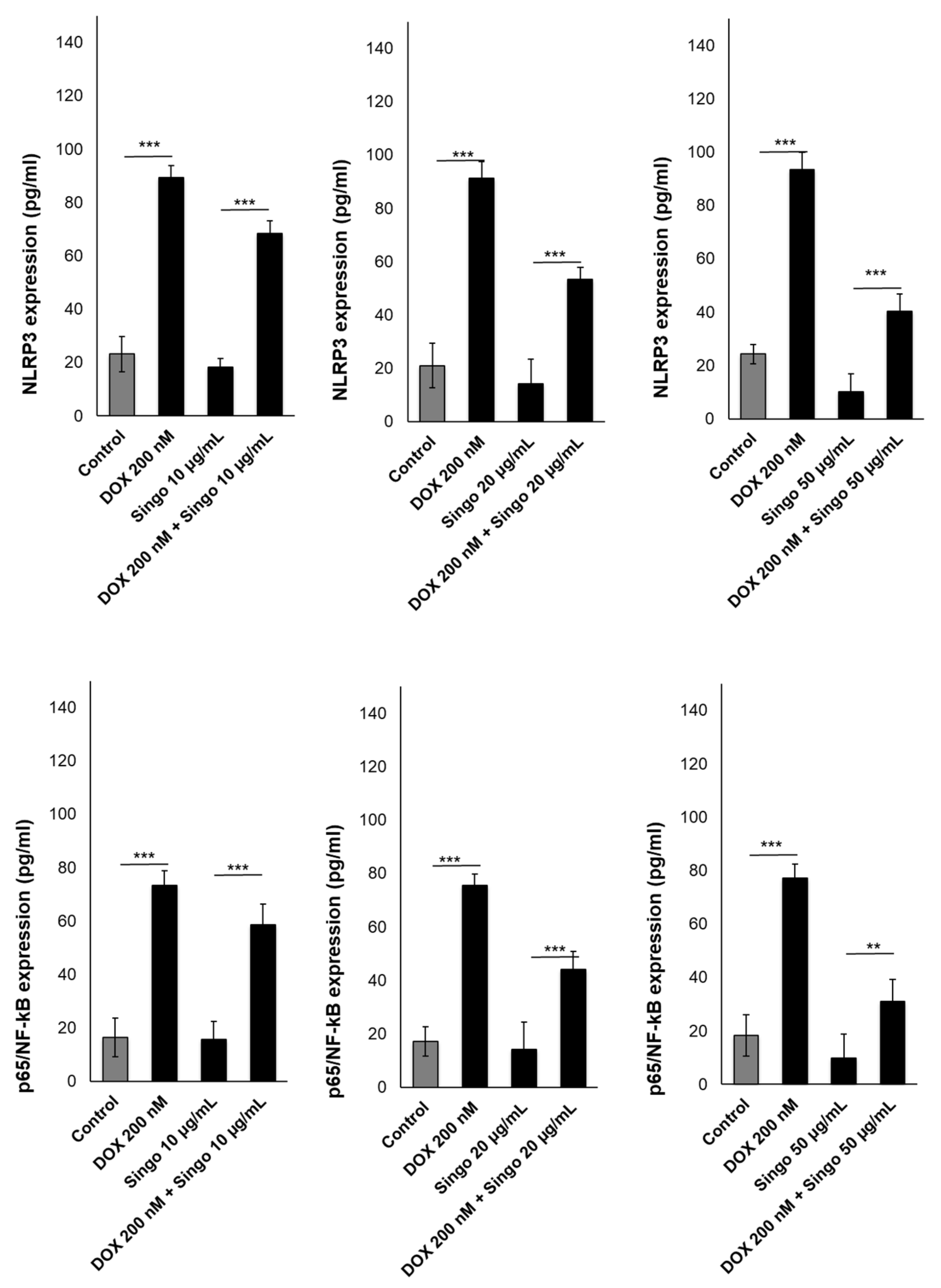
3.3. Myocardial DAMPs and Inflammasome Expression
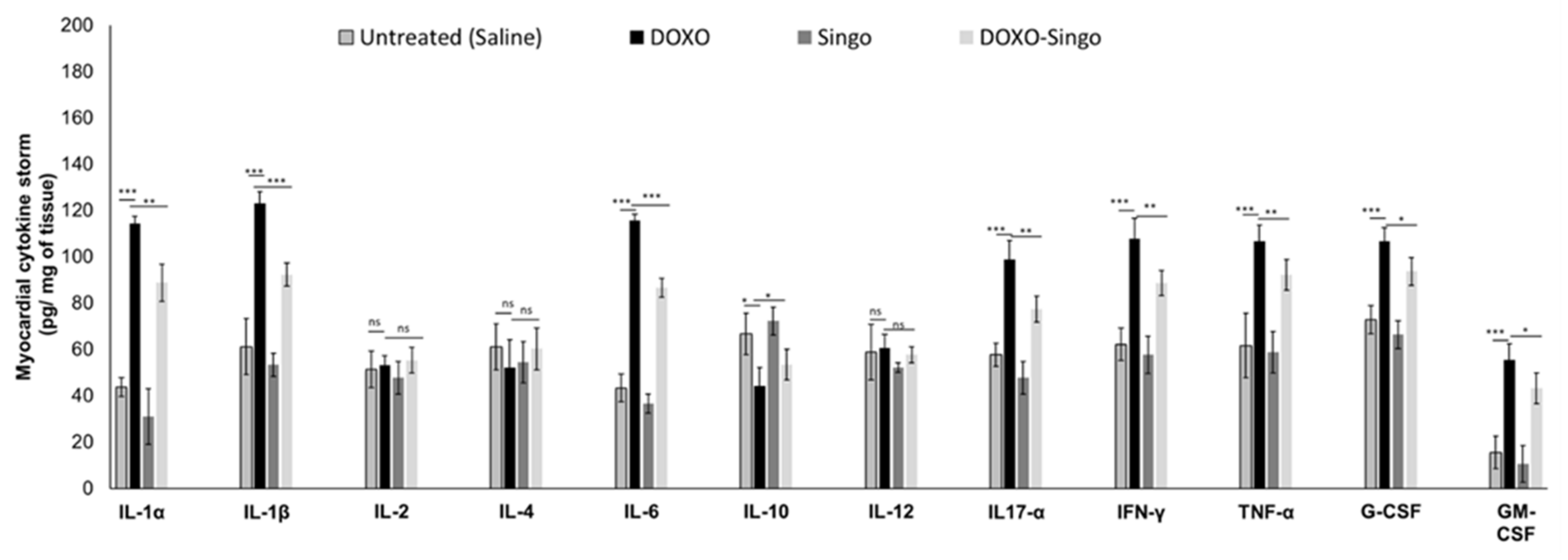
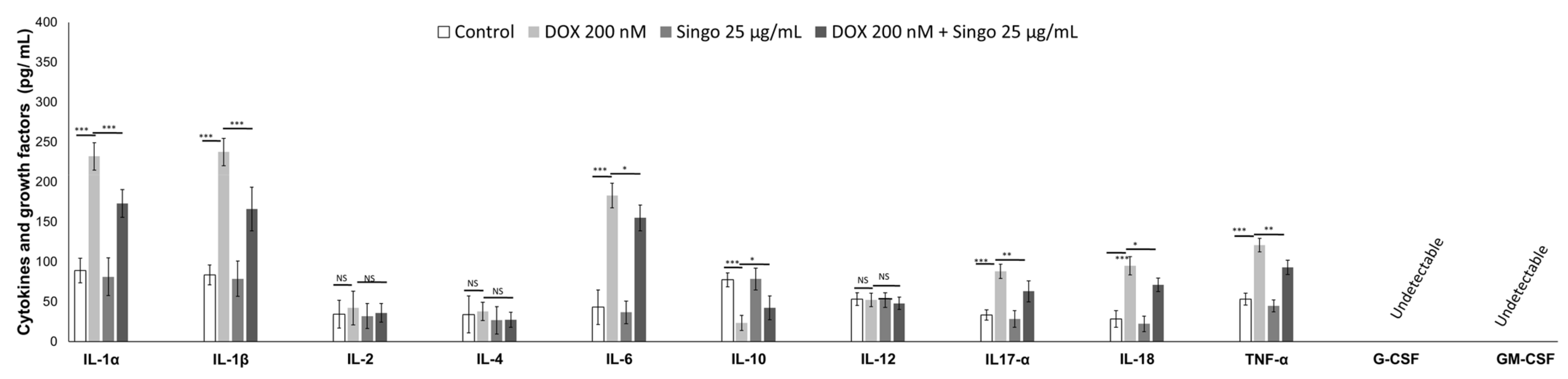
3.4. Cytokinome Profile Evaluation in Myocardial Tissue
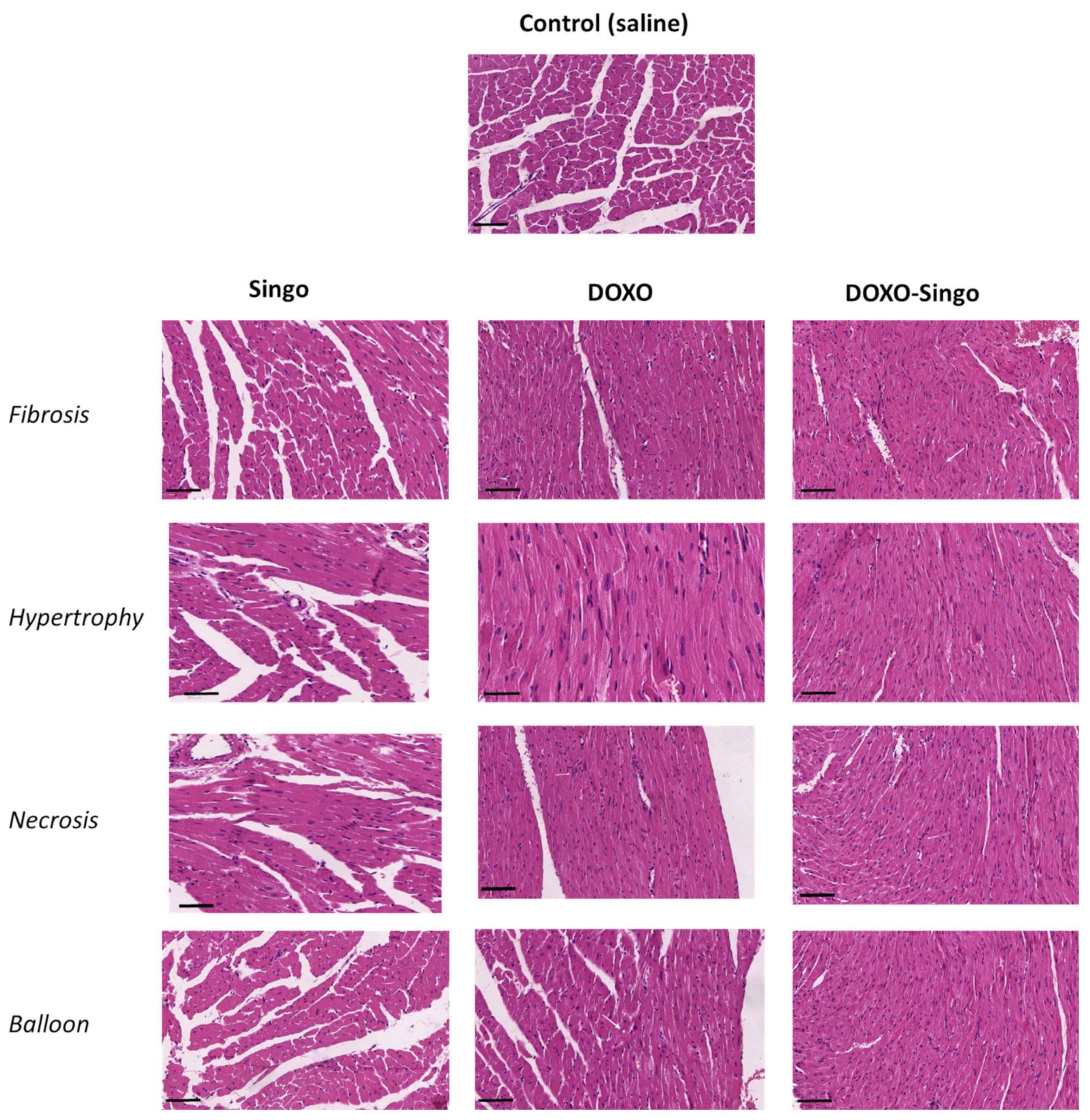
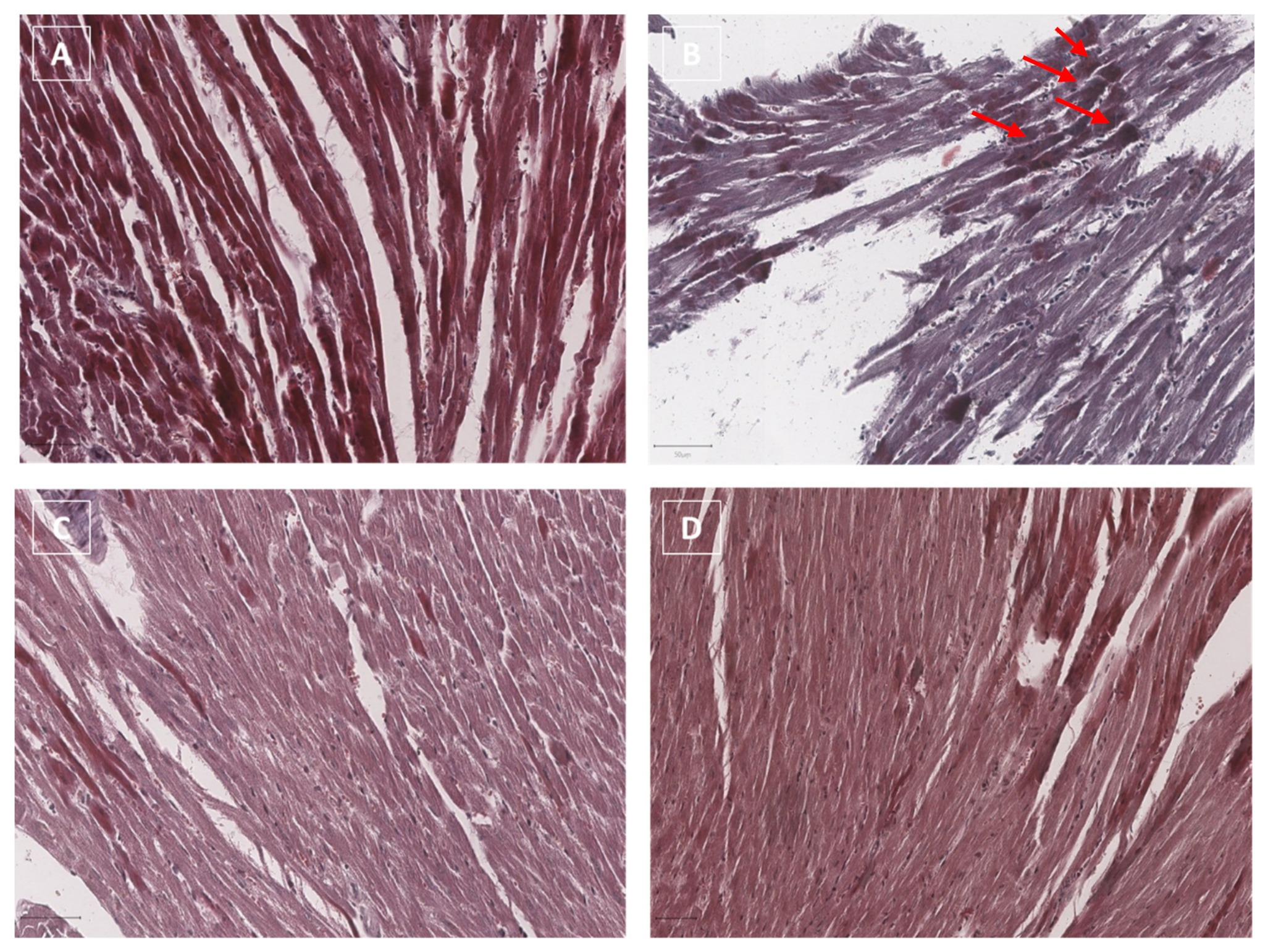
3.5. Histology Evaluations: H&E and Masson’s Trichrome Staining in Myocardial Tissues
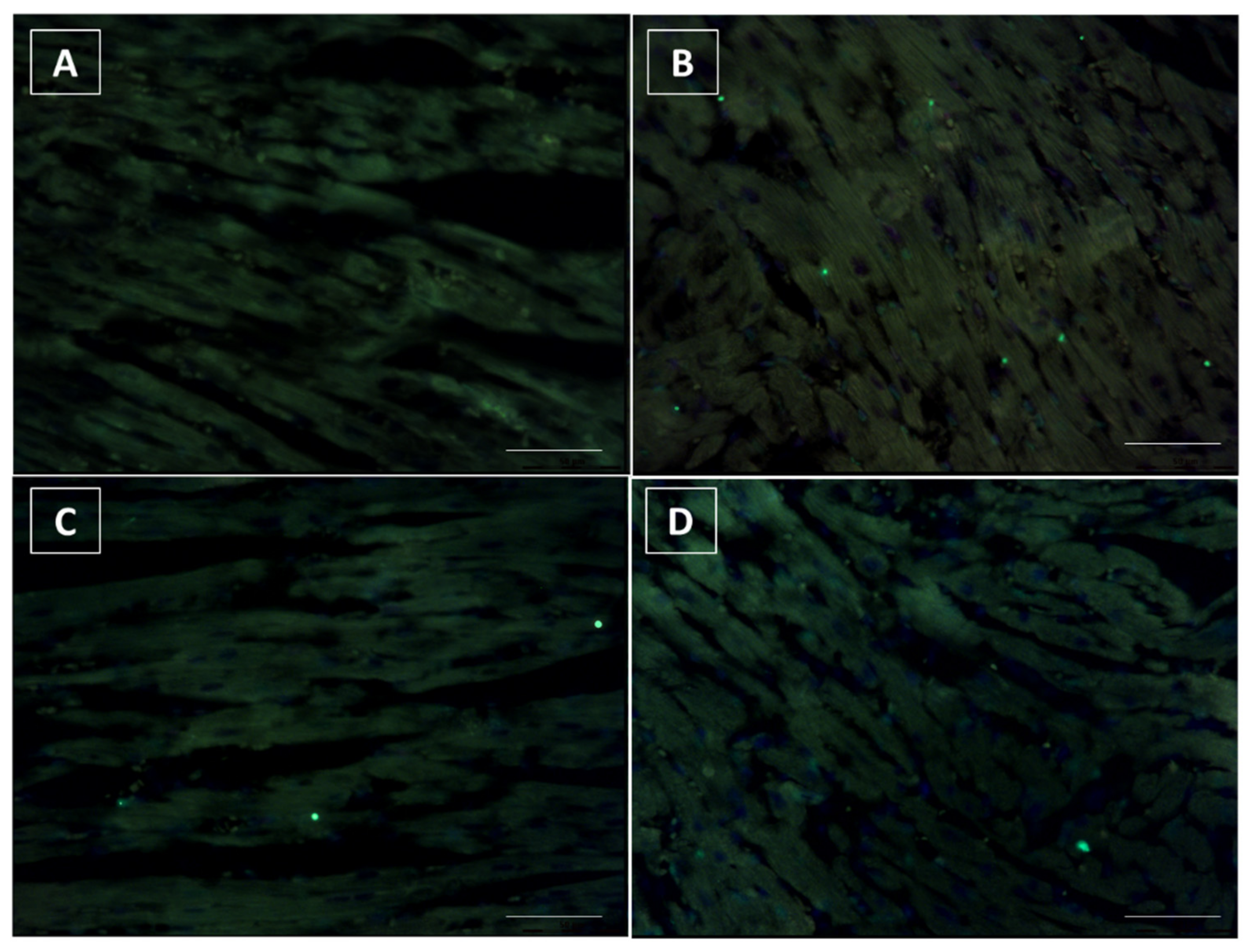
3.6. Terminal dUTP Nick End-Labeling (TUNEL) Assay
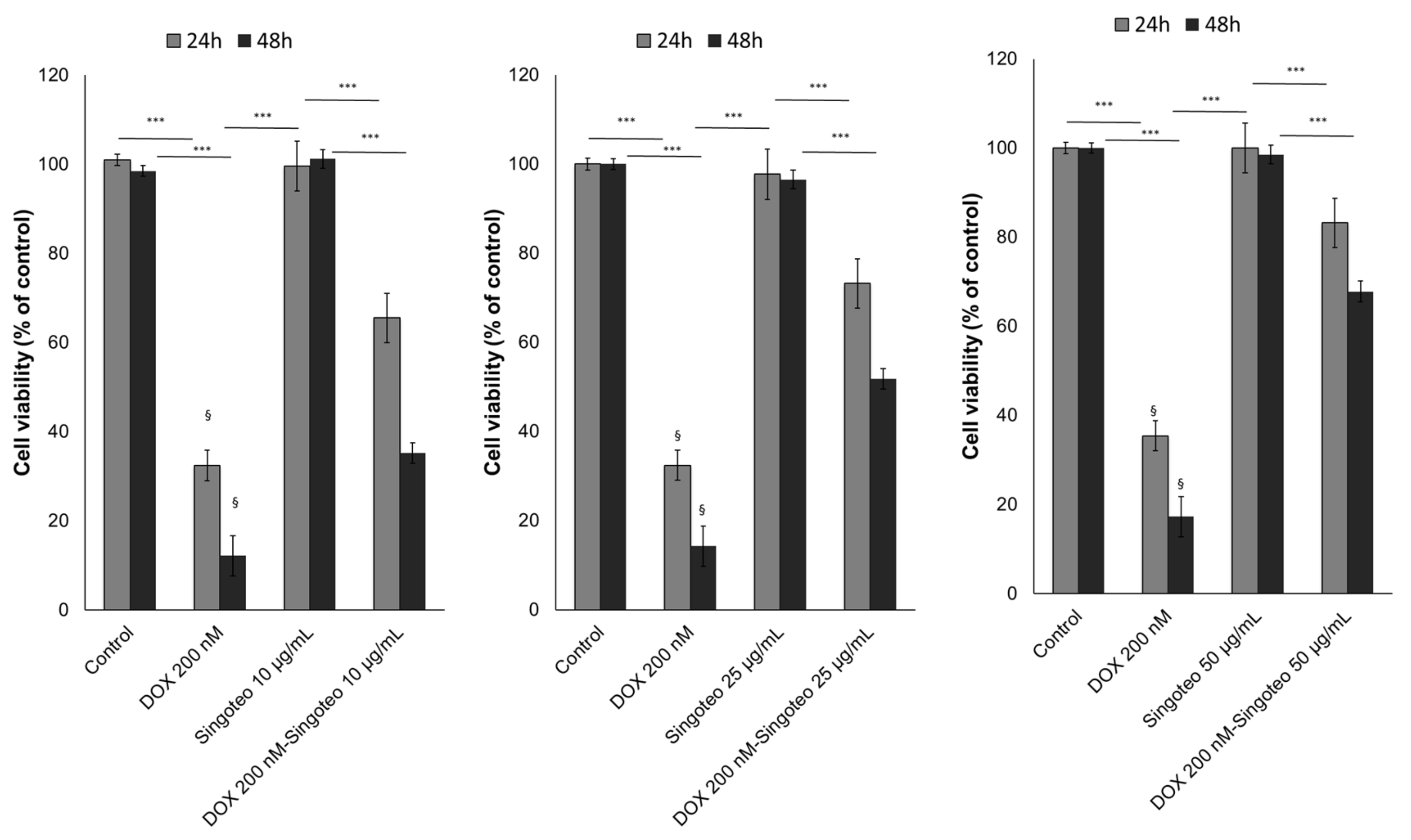
3.7. Cell Viability
3.8. Lipid Peroxidation
3.9. Inflammasome and p65/NF-kB Expression in Human Cardiomyocytes
Cytokinome Profile Evaluation in Human Cardiomyocytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nicoletto, R.E.; Ofner, C.M., III. Cytotoxic mechanisms of doxorubicin at clinically relevant concentrations in breast cancer cells. Cancer Chemother. Pharmacol. 2022, 89, 285–311. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Wang, D. Analysis of Cardiac Adverse Reactions Caused by Different Doses of Adriamycin Chemotherapy in Patients with Breast Cancer. J. Healthc. Eng. 2022, 2022, 1642244. [Google Scholar] [CrossRef] [PubMed]
- Sheibani, M.; Azizi, Y.; Shayan, M.; Nezamoleslami, S.; Eslami, F.; Farjoo, M.H.; Dehpour, A.R. Doxorubicin-Induced Cardiotoxicity: An Overview on Pre-clinical Therapeutic Approaches. Cardiovasc. Toxicol. 2022, 22, 292–310. [Google Scholar] [CrossRef] [PubMed]
- Tadokoro, T.; Ikeda, M.; Ide, T.; Deguchi, H.; Ikeda, S.; Okabe, K.; Ishikita, A.; Matsushima, S.; Koumura, T.; Yamada, K.I.; et al. Mitochondria-dependent ferroptosis plays a pivotal role in doxorubicin cardiotoxicity. JCI Insight 2020, 5, e132747. [Google Scholar] [CrossRef] [PubMed]
- Kitakata, H.; Endo, J.; Ikura, H.; Moriyama, H.; Shirakawa, K.; Katsumata, Y.; Sano, M. Therapeutic Targets for DOX-Induced Cardiomyopathy: Role of Apoptosis vs. Ferroptosis. Int. J. Mol. Sci. 2022, 23, 1414. [Google Scholar] [CrossRef]
- Syukri, A.; Budu Hatta, M.; Amir, M.; Rohman, M.S.; Mappangara, I.; Kaelan, C.; Wahyuni, S.; Bukhari, A.; Junita, A.R.; Primaguna, M.R.; et al. Doxorubicin induced immune abnormalities and inflammatory responses via HMGB1, HIF1-α and VEGF pathway in progressive of cardiovascular damage. Ann. Med. Surg. 2022, 76, 103501. [Google Scholar] [CrossRef]
- Quagliariello, V.; Vecchione, R.; De Capua, A.; Lagreca, E.; Iaffaioli, R.V.; Botti, G.; Netti, P.A.; Maurea, N. Nano-Encapsulation of Coenzyme Q10 in Secondary and Tertiary Nano-Emulsions for Enhanced Cardioprotection and Hepatoprotection in Human Cardiomyocytes and Hepatocytes During Exposure to Anthracyclines and Trastuzumab. Int. J. Nanomed. 2020, 15, 4859–4876. [Google Scholar] [CrossRef]
- Osataphan, N.; Phrommintikul, A.; Chattipakorn, S.C.; Chattipakorn, N. Effects of doxorubicin-induced cardiotoxicity on cardiac mitochondrial dynamics and mitochondrial function: Insights for future interventions. J. Cell Mol. Med. 2020, 24, 6534–6557. [Google Scholar] [CrossRef]
- Totzeck, M.; Mincu, R.I.; Heusch, G.; Rassaf, T. Heart failure from cancer therapy: Can we prevent it? ESC Heart Fail. 2019, 6, 856–862. [Google Scholar] [CrossRef] [Green Version]
- Quagliariello, V.; De Laurentiis, M.; Rea, D.; Barbieri, A.; Monti, M.G.; Carbone, A.; Paccone, A.; Altucci, L.; Conte, M.; Canale, M.L.; et al. The SGLT-2 inhibitor empagliflozin improves myocardial strain, reduces cardiac fibrosis and pro-inflammatory cytokines in non-diabetic mice treated with doxorubicin. Cardiovasc. Diabetol. 2021, 20, 150. [Google Scholar] [CrossRef]
- Schreckenberg, R.; Wolf, A.; Szabados, T.; Gömöri, K.; Szabó, I.A.; Ágoston, G.; Brenner, G.; Bencsik, P.; Ferdinandy, P.; Schulz, R.; et al. Proprotein Convertase Subtilisin Kexin Type 9 (PCSK9) Deletion but Not Inhibition of Extracellular PCSK9 Reduces Infarct Sizes Ex Vivo but Not In Vivo. Int. J. Mol. Sci. 2022, 23, 6512. [Google Scholar] [CrossRef]
- Butt, J.H.; Dewan, P.; Jhund, P.S.; Anand, I.S.; Atar, D.; Ge, J.; Desai, A.S.; Echeverria, L.E.; Køber, L.; Lam, C.S.P.; et al. Sacubitril/Valsartan and Frailty in Patients with Heart Failure and Preserved Ejection Fraction. J. Am. Coll Cardiol. 2022, 80, 1130–1143. [Google Scholar] [CrossRef]
- Badimon, L.; Vilahur, G.; Padro, T. Nutraceuticals and atherosclerosis: Human trials. Cardiovasc. Ther. 2010, 28, 202–215. [Google Scholar] [CrossRef]
- Vilahur, G.; Casaní, L.; Peña, E.; Crespo, J.; Juan-Babot, O.; Ben-Aicha, S.; Mendieta, G.; Béjar, M.T.; Borrell, M.; Badimon, L. Silybum marianum provides cardioprotection and limits adverse remodeling post-myocardial infarction by mitigating oxidative stress and reactive fibrosis. Int. J. Cardiol. 2018, 270, 28–35. [Google Scholar] [CrossRef]
- Carrizzo, A.; Izzo, C.; Forte, M.; Sommella, E.; Di Pietro, P.; Venturini, E.; Ciccarelli, M.; Galasso, G.; Rubattu, S.; Campiglia, P.; et al. A Novel Promising Frontier for Human Health: The Beneficial Effects of Nutraceuticals in Cardiovascular Diseases. Int. J. Mol. Sci. 2020, 21, 8706. [Google Scholar] [CrossRef]
- Costa, F.O.; Cortelli, S.C.; Costa, A.A.; Cyrino, R.M.; Cortelli, J.R.; Miranda Cota, L.O. Impact of compliance during periodontal maintenance therapy on oral health-related quality of life: A 6-year follow-up. J. Dent. 2019, 83, 50–55. [Google Scholar] [CrossRef]
- Finamore, A.; Palmery, M.; Bensehaila, S.; Peluso, I. Antioxidant, Immunomodulating, and Microbial-Modulating Activities of the Sustainable and Ecofriendly Spirulina. Oxid. Med. Cell Longev. 2017, 2017, 3247528. [Google Scholar] [CrossRef] [Green Version]
- DiNicolantonio, J.J.; Bhat, A.G.; OKeefe, J. Effects of spirulina on weight loss and blood lipids: A review. Open Heart 2020, 7, e001003. [Google Scholar] [CrossRef] [Green Version]
- Yousefi, O.S.; Günther, M.; Hörner, M.; Chalupsky, J.; Wess, M.; Brandl, S.M.; Smith, R.W.; Fleck, C.; Kunkel, T.; Zurbriggen, M.D.; et al. Optogenetic control shows that kinetic proofreading regulates the activity of the T cell receptor. eLife 2019, 8, e42475. [Google Scholar] [CrossRef]
- DiNicolantonio, J.J.; Barroso-Aranda, J.; McCarty, M.F. Anti-inflammatory activity of ivermectin in late-stage COVID-19 may reflect activation of systemic glycine receptors. Open Heart 2021, 8, e001655. [Google Scholar] [CrossRef]
- Grosshagauer, S.; Kraemer, K.; Somoza, V. The True Value of Spirulina. J. Agric. Food Chem. 2020, 68, 4109–4115. [Google Scholar] [CrossRef] [PubMed]
- Park, I.H.; Zhao, R.; West, J.A.; Yabuuchi, A.; Huo, H.; Ince, T.A.; Lerou, P.H.; Lensch, M.W.; Daley, G.Q. Reprogramming of human somatic cells to pluripotency with defined factors. Nature 2008, 451, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Mazokopakis, E.E.; Starakis, I.K.; Papadomanolaki, M.G.; Mavroeidi, N.G.; Ganotakis, E.S. The hypolipidaemic effects of Spirulina (Arthrospira platensis) supplementation in a Cretan population: A prospective study. J. Sci. Food Agric. 2014, 94, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Serban, M.C.; Sahebkar, A.; Dragan, S.; Stoichescu-Hogea, G.; Ursoniu, S.; Andrica, F.; Banach, M. A systematic review and meta-analysis of the impact of Spirulina supplementation on plasma lipid concentrations. Clin. Nutr. 2016, 35, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.S.; Kim, S.K. Down-regulation of histamine-induced endothelial cell activation as potential anti-atherosclerotic activity of peptides from Spirulina maxima. Eur. J. Pharm. Sci. 2013, 50, 198–207. [Google Scholar] [CrossRef]
- Ren, J.; Friedmann, D.; Xiong, J.; Liu, C.D.; Ferguson, B.R.; Weerakkody, T.; DeLoach, K.E.; Ran, C.; Pun, A.; Sun, Y.; et al. Anatomically Defined and Functionally Distinct Dorsal Raphe Serotonin Sub-systems. Cell 2018, 175, 472–487.e20. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Kolts, R.L.; Rapaport, M.H.; Krishnan, K.R.; Brodhead, A.E.; Browns, W.A. Magnitude of placebo response and drug-placebo differences across psychiatric disorders. Psychol. Med. 2005, 35, 743–749. [Google Scholar] [CrossRef]
- Khan, R.J.; Fick, D.; Yao, F.; Tang, K.; Hurworth, M.; Nivbrant, B.; Wood, D. A comparison of three methods of wound closure following arthroplasty: A prospective, randomised, controlled trial. J. Bone Joint Surg. Br. 2006, 88, 238–242. [Google Scholar] [CrossRef]
- Kou, X.; Li, B.; Olayanju, J.B.; Drake, J.M.; Chen, N. Nutraceutical or Pharmacological Potential of Moringa oleifera Lam. Nutrients 2018, 10, 343. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Wang, J.; Meng, C.; Liang, W.; Wang, T.; Zhou, B.; Wang, Y.; Luo, X.; Gao, L.; Zhang, L. Moringa oleifera Lam and its Therapeutic Effects in Immune Disorders. Front Pharmacol. 2020, 11, 566783. [Google Scholar] [CrossRef]
- Luetragoon, T.; Pankla Sranujit, R.; Noysang, C.; Thongsri, Y.; Potup, P.; Suphrom, N.; Nuengchamnong, N.; Usuwanthim, K. Bioactive Compounds in Moringa oleifera Lam. Leaves Inhibit the Pro-Inflammatory Mediators in Lipopolysaccharide-Induced Human Monocyte-Derived Macrophages. Molecules 2020, 25, 191. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Ruiz Beguerie, J.; Sze, D.M.; Chan, G.C. Ganoderma lucidum (Reishi mushroom) for cancer treatment. Cochrane Database Syst. Rev. 2016, 4, CD007731. [Google Scholar] [CrossRef]
- Barbieri, A.; Quagliariello, V.; Del Vecchio, V.; Falco, M.; Luciano, A.; Amruthraj, N.J.; Nasti, G.; Ottaiano, A.; Berretta, M.; Iaffaioli, R.V.; et al. Anticancer and Anti-Inflammatory Properties of Ganoderma lucidum Extract Effects on Melanoma and Triple-Negative Breast Cancer Treatment. Nutrients 2017, 9, 210. [Google Scholar] [CrossRef] [Green Version]
- Klupp, N.L.; Chang, D.; Hawke, F.; Kiat, H.; Cao, H.; Grant, S.J.; Bensoussan, A. Ganoderma lucidum mushroom for the treatment of cardiovascular risk factors. Cochrane Database Syst. Rev. 2015, 2015, CD007259. [Google Scholar] [CrossRef]
- Cör, D.; Knez, Ž.; Knez Hrnčič, M. Antitumour, Antimicrobial, Antioxidant and Antiacetylcholinesterase Effect of Ganoderma Lucidum Terpenoids and Polysaccharides: A Review. Molecules. 2018, 23, 649. [Google Scholar] [CrossRef] [Green Version]
- Quagliariello, V.; Passariello, M.; Coppola, C.; Rea, D.; Barbieri, A.; Scherillo, M.; Monti, M.G.; Iaffaioli, R.V.; De Laurentiis, M.; Ascierto, P.A.; et al. Cardiotoxicity and pro-inflammatory effects of the immune checkpoint inhibitor Pembrolizumab associated with Trastuzumab. Int. J. Cardiol. 2019, 292, 171–179. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254, ISSN 0003-2697. [Google Scholar] [CrossRef]
- Medzihradszky, K.F. In-solution digestion of proteins for mass spectrometry. Methods Enzymol. 2005, 405, 50–65. [Google Scholar] [CrossRef]
- Carrizzo, A.; Conte, G.M.; Sommella, E.; Damato, A.; Ambrosio, M.; Sala, M.; Scala, M.C.; Aquino, R.P.; De Lucia, M.; Madonna, M.; et al. Novel Potent Decameric Peptide of Spirulina platensis Reduces Blood Pressure Levels Through a PI3K/AKT/eNOS-Dependent Mechanism. Hypertension 2018, 73, 449–457. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, X.; Huang, A. Proteomic analysis and food-grade enzymes of Moringa oleifer Lam. a Lam. flower. Int. J. Biol. Macromol. 2018, 115, 883–890, ISSN 0141-8130. [Google Scholar] [CrossRef]
- Nielsen, S.S. Total carbohydrate by phenol-sulfuric acid method. In Food Analysis Laboratory Manual; Food Science Text Series; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Kan, Y.; Chen, T.; Wu, Y.; Wu, J.; Wu, J. Antioxidant activity of polysaccharide extracted from Ganoderma lucidum using response surface methodology. Int. J. Biol. Macromol. 2015, 72, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Tocchetti, C.G.; Carpi, A.; Coppola, C.; Quintavalle, C.; Rea, D.; Campesan, M.; Arcari, A.; Piscopo, G.; Cipresso, C.; Monti, M.G.; et al. Ranolazine protects from doxorubicin-induced oxidative stress and cardiac dysfunction. Eur. J. Heart Fail. 2014, 16, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Kong, S.; Wu, M.; Niu, Y.; Wang, K.; Zhu, H.; Yuan, J. Impact high fat diet on myocardial strain in mice by 2D speckle tracking imaging. Obes. Res. Clin. Pract. 2021, 15, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.; Cheng, S.; Jain, M.; Ngoy, S.; Theodoropoulos, C.; Trujillo, A.; Lin, F.C.; Liao, R. Echocardiographic speckle-tracking based strain imaging for rapid cardiovascular phenotyping in mice. Circ. Res. 2011, 108, 908–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cihakova, D.; Barin, J.G.; Afanasyeva, M.; Kimura, M.; Fairweather, D.; Berg, M.; Talor, M.V.; Baldeviano, G.C.; Frisancho, S.; Gabrielson, K.; et al. Interleukin-13 protects against experimental autoimmune myocarditis by regulating macrophage differentiation. Am. J. Pathol. 2008, 172, 1195–1208. [Google Scholar] [CrossRef] [Green Version]
- Quagliariello, V.; Passariello, M.; Di Mauro, A.; Cipullo, C.; Paccone, A.; Barbieri, A.; Palma, G.; Luciano, A.; Buccolo, S.; Bisceglia, I.; et al. Immune checkpoint inhibitor therapy increases systemic SDF-1, cardiac DAMPs Fibronectin-EDA, S100/Calgranulin, galectin-3, and NLRP3-MyD88-chemokine pathways. Front. Cardiovasc. Med. 2022, 9, 930797. [Google Scholar] [CrossRef]
- Quagliariello, V.; Berretta, M.; Buccolo, S.; Iovine, M.; Paccone, A.; Cavalcanti, E.; Taibi, R.; Montopoli, M.; Botti, G.; Maurea, N. Polydatin Reduces Cardiotoxicity and Enhances the Anticancer Effects of Sunitinib by Decreasing Pro-Oxidative Stress, Pro-Inflammatory Cytokines, and NLRP3 Inflammasome Expression. Front. Oncol. 2021, 11, 680758. [Google Scholar] [CrossRef]
- Soulage, C.O.; Pelletier, C.C.; Florens, N.; Lemoine, S.; Dubourg, L.; Juillard, L.; Guebre-Egziabher, F. Two Toxic Lipid Aldehydes, 4-hydroxy-2-hexenal (4-HHE) and 4-hydroxy-2-nonenal (4-HNE), Accumulate in Patients with Chronic Kidney Disease. Toxins 2020, 12, 567. [Google Scholar] [CrossRef]
- Quagliariello, V.; Passariello, M.; Rea, D.; Barbieri, A.; Iovine, M.; Bonelli, A.; Caronna, A.; Botti, G.; De Lorenzo, C.; Maurea, N. Evidences of CTLA-4 and PD-1 Blocking Agents-Induced Cardiotoxicity in Cellular and Preclinical Models. J. Pers. Med. 2020, 10, 179. [Google Scholar] [CrossRef]
- Sakalyte, R.; Denkovskij, J.; Bernotiene, E.; Stropuviene, S.; Mikulenaite, S.O.; Kvederas, G.; Porvaneckas, N.; Tutkus, V.; Venalis, A.; Butrimiene, I. The Expression of Inflammasomes NLRP1 and NLRP3, Toll-Like Receptors, and Vitamin D Receptor in Synovial Fibroblasts from Patients with Different Types of Knee Arthritis. Front. Immunol. 2022, 12, 767512. [Google Scholar] [CrossRef]
- Mubarak, A.; Alrfaei, B.; Aljurayyan, A.; Alqafil, M.M.; Farrag, M.A.; Hamed, M.E.; Alosaimi, B.; Almajhdi, F.; Alturaiki, W. In vivo and in vitro Evaluation of Cytokine Expression Profiles During Middle East Respiratory Syndrome Coronavirus (MERS-CoV) Infection. J. Inflamm. Res. 2021, 14, 2121–2131. [Google Scholar] [CrossRef]
- Cocco, L.D.; Chiaparini, A.F.; Saffi, M.A.L.; Leiria, T.L.L. Global Longitudinal Strain for the Early Detection of Chemotherapy-Induced Cardiotoxicity: A Systematic Review and Meta-analysis. Clin. Oncol. 2022, 34, 514–525. [Google Scholar] [CrossRef]
- Kato, J.; Svensson, C.I. Role of extracellular damage-associated molecular pattern molecules (DAMPs) as mediators of persistent pain. Prog. Mol. Biol. Transl. Sci. 2015, 131, 251–279. [Google Scholar] [CrossRef]
- Hancq, S.; Salmon, I.; Brotchi, J.; Gabius, H.J.; Heizmann, C.W.; Kiss, R.; Decaestecker, C. Detection of S100B, S100A6 and galectin-3 ligands in meningiomas as markers of aggressiveness. Int. J. Oncol. 2004, 25, 1233–1240. [Google Scholar] [CrossRef]
- Rubio-Infante, N.; Ramírez-Flores, Y.A.; Castillo, E.C.; Lozano, O.; García-Rivas, G.; Torre-Amione, G. A Systematic Review of the Mechanisms Involved in Immune Checkpoint Inhibitors Cardiotoxicity and Challenges to Improve Clinical Safety. Front. Cell Dev. Biol. 2022, 10, 851032. [Google Scholar] [CrossRef]
- Haupt, L.P.; Rebs, S.; Maurer, W.; Hübscher, D.; Tiburcy, M.; Pabel, S.; Maus, A.; Köhne, S.; Tappu, R.; Haas, J.; et al. Doxorubicin induces cardiotoxicity in a pluripotent stem cell model of aggressive B cell lymphoma cancer patients. Basic Res. Cardiol. 2022, 117, 13. [Google Scholar] [CrossRef]
- Li, D.; Liu, X.; Pi, W.; Zhang, Y.; Yu, L.; Xu, C.; Sun, Z.; Jiang, J. Fisetin Attenuates Doxorubicin-Induced Cardiomyopathy In Vivo and In Vitro by Inhibiting Ferroptosis Through SIRT1/Nrf2 Signaling Pathway Activation. Front. Pharmacol. 2022, 12, 808480. [Google Scholar] [CrossRef]
- Russell, C.; Keshavamurthy, S.; Saha, S. Nutraceuticals in the Management of Cardiovascular Risk Factors: Where is the Evidence? Cardiovasc. Hematol. Disord. Drug Targets 2021, 21, 150–161. [Google Scholar] [CrossRef]
- Wei, S.; Ma, W.; Li, X.; Jiang, C.; Sun, T.; Li, Y.; Zhang, B.; Li, W. Involvement of ROS/NLRP3 Inflammasome Signaling Pathway in Doxorubicin-Induced Cardiotoxicity. Cardiovasc. Toxicol. 2020, 20, 507–519. [Google Scholar] [CrossRef]
- Szekely, Y.; Arbel, Y. A Review of Interleukin-1 in Heart Disease: Where Do We Stand Today? Cardiol. Ther. 2018, 7, 25–44. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Lakdawalla, D.N.; Shafrin, J.; Hou, N.; Peneva, D.; Vine, S.; Park, J.; Zhang, J.; Brookmeyer, R.; Figlin, R.A. Predicting Real-World Effectiveness of Cancer Therapies Using Overall Survival and Progression-Free Survival from Clinical Trials: Empirical Evidence for the ASCO Value Framework. Value Health 2017, 20, 866–875. [Google Scholar] [CrossRef] [PubMed]
- Lyon, A.R.; López-Fernández, T.; Couch, L.S.; Asteggiano, R.; Aznar, M.C.; Bergler-Klein, J.; Boriani, G.; Cardinale, D.; Cordoba, R.; Cosyns, B.; et al. 2022 ESC Guidelines on cardio-oncology developed in collaboration with the European Hematology Association (EHA), the European Society for Therapeutic Radiology and Oncology (ESTRO) and the International Cardio-Oncology Society (IC-OS). Eur. Heart J. 2022, 43, ehac244. [Google Scholar] [CrossRef] [PubMed]
- Morelli, M.B.; Bongiovanni, C.; Da Pra, S.; Miano, C.; Sacchi, F.; Lauriola, M.; D’Uva, G. Cardiotoxicity of Anticancer Drugs: Molecular Mechanisms and Strategies for Cardioprotection. Front. Cardiovasc. Med. 2022, 9, 847012. [Google Scholar] [CrossRef] [PubMed]
- Vitfell-Rasmussen, J.; Krarup-Hansen, A.; Vaage-Nilsen, M.; Kümler, T.; Zerahn, B. Real-life incidence of cardiotoxicity and associated risk factors in sarcoma patients receiving doxorubicin. Acta Oncol. 2022, 61, 801–808. [Google Scholar] [CrossRef]
- Carvalho, G.; Pelletier, P.; Albacker, T.; Lachapelle, K.; Joanisse, D.R.; Hatzakorzian, R.; Lattermann, R.; Sato, H.; Marette, A.; Schricker, T. Cardioprotective effects of glucose and insulin administration while maintaining normoglycemia (GIN therapy) in patients undergoing coronary artery bypass grafting. J. Clin. Endocrinol. Metab. 2011, 96, 1469–1477. [Google Scholar] [CrossRef] [Green Version]
- Escolà-Gil, J.C.; Cedó, L.; Blanco-Vaca, F. High-density lipoprotein cholesterol targeting for novel drug discovery: Where have we gone wrong? Expert Opin Drug Discov. 2014, 9, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Campana, C.; Dariolli, R.; Boutjdir, M.; Sobie, E.A. Inflammation as a Risk Factor in Cardiotoxicity: An Important Consideration for Screening During Drug Development. Front. Pharmacol. 2021, 12, 598549. [Google Scholar] [CrossRef]
- Quagliariello, V.; Paccone, A.; Iovine, M.; Cavalcanti, E.; Berretta, M.; Maurea, C.; Canale, M.L.; Maurea, N. Interleukin-1 blocking agents as promising strategy for prevention of anticancer drug-induced cardiotoxicities: Possible implications in cancer patients with COVID-19. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 6797–6812. [Google Scholar] [CrossRef]
- Sun, W.; Xu, J.; Wang, L.; Jiang, Y.; Cui, J.; Su, X.; Yang, F.; Tian, L.; Si, Z.; Xing, Y. Non-coding RNAs in cancer therapy-induced cardiotoxicity: Mechanisms, biomarkers, and treatments. Front Cardiovasc. Med. 2022, 9, 946137. [Google Scholar] [CrossRef]
- Afsheen, N.; Khalil-Ur-Rehman Jahan, N.; Ijaz, M.; Manzoor, A.; Khan, K.M.; Hina, S. Cardioprotective and Metabolomic Profiling of Selected Medicinal Plants against Oxidative Stress. Oxid. Med. Cell Longev. 2018, 2018, 9819360. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A.; Mukerjee, N.; Sharma, B.; Pant, A.; Kishore Mohanta, Y.; Jawarkar, R.D.; Bakal, R.L.; Terefe, E.M.; Batiha, G.E.; Mostafa-Hedeab, G.; et al. Target Specific Inhibition of Protein Tyrosine Kinase in Conjunction with Cancer and SARS-COV-2 by Olive Nutraceuticals. Front. Pharmacol. 2022, 12, 812565. [Google Scholar] [CrossRef]
- Narayanankutty, A. Inhibitory Potential of Dietary Nutraceuticals on Cellular PI3K/Akt Signaling: Implications in Cancer Prevention and Therapy. Curr. Top Med. Chem. 2021, 21, 1816–1831. [Google Scholar] [CrossRef]
- Wu, X.; Liu, Z.; Liu, Y.; Yang, Y.; Shi, F.; Cheong, K.L.; Teng, B. Immunostimulatory Effects of Polysaccharides from Spirulina platensis In Vivo and Vitro and Their Activation Mechanism on RAW246.7 Macrophages. Mar. Drugs. 2020, 18, 538. [Google Scholar] [CrossRef]
- Kabel, A.M.; Salama, S.A.; Adwas, A.A.; Estfanous, R.S. Targeting Oxidative Stress, NLRP3 Inflammasome, and Autophagy by Fraxetin to Combat Doxorubicin-Induced Cardiotoxicity. Pharmaceuticals 2021, 14, 1188. [Google Scholar] [CrossRef]
- Wu, J.; Dong, E.; Zhang, Y.; Xiao, H. The Role of the Inflammasome in Heart Failure. Front Physiol. 2021, 12, 709703. [Google Scholar] [CrossRef]
- Whiteside, T.L. Immune suppression in cancer: Effects on immune cells, mechanisms and future therapeutic intervention. Semin. Cancer Biol. 2006, 16, 3–15. [Google Scholar] [CrossRef]
- Padegimas, A.; Clasen, S.; Ky, B. Cardioprotective strategies to prevent breast cancer therapy-induced cardiotoxicity. Trends Cardiovasc. Med. 2020, 30, 22–28. [Google Scholar] [CrossRef]
- Sheng, C.C.; Amiri-Kordestani, L.; Palmby, T.; Force, T.; Hong, C.C.; Wu, J.C.; Croce, K.; Kim, G.; Moslehi, J. 21st Century Cardio-Oncology: Identifying Cardiac Safety Signals in the Era of Personalized Medicine. JACC Basic Transl. Sci. 2016, 1, 386–398. [Google Scholar] [CrossRef]
- Peres Diaz, L.S.; Schuman, M.L.; Aisicovich, M.; Toblli, J.E.; Pirola, C.J.; Landa, M.S.; García, S.I. Short-term doxorubicin cardiotoxic effects: Involvement of cardiac Thyrotropin Releasing Hormone system. Life Sci. 2020, 261, 118346. [Google Scholar] [CrossRef]














Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quagliariello, V.; Basilicata, M.G.; Pepe, G.; De Anseris, R.; Di Mauro, A.; Scognamiglio, G.; Palma, G.; Vestuto, V.; Buccolo, S.; Luciano, A.; et al. Combination of Spirulina platensis, Ganoderma lucidum and Moringa oleifera Improves Cardiac Functions and Reduces Pro-Inflammatory Biomarkers in Preclinical Models of Short-Term Doxorubicin-Mediated Cardiotoxicity: New Frontiers in Cardioncology? J. Cardiovasc. Dev. Dis. 2022, 9, 423. https://doi.org/10.3390/jcdd9120423
Quagliariello V, Basilicata MG, Pepe G, De Anseris R, Di Mauro A, Scognamiglio G, Palma G, Vestuto V, Buccolo S, Luciano A, et al. Combination of Spirulina platensis, Ganoderma lucidum and Moringa oleifera Improves Cardiac Functions and Reduces Pro-Inflammatory Biomarkers in Preclinical Models of Short-Term Doxorubicin-Mediated Cardiotoxicity: New Frontiers in Cardioncology? Journal of Cardiovascular Development and Disease. 2022; 9(12):423. https://doi.org/10.3390/jcdd9120423
Chicago/Turabian StyleQuagliariello, Vincenzo, Manuela Giovanna Basilicata, Giacomo Pepe, Raffaele De Anseris, Annabella Di Mauro, Giosuè Scognamiglio, Giuseppe Palma, Vincenzo Vestuto, Simona Buccolo, Antonio Luciano, and et al. 2022. "Combination of Spirulina platensis, Ganoderma lucidum and Moringa oleifera Improves Cardiac Functions and Reduces Pro-Inflammatory Biomarkers in Preclinical Models of Short-Term Doxorubicin-Mediated Cardiotoxicity: New Frontiers in Cardioncology?" Journal of Cardiovascular Development and Disease 9, no. 12: 423. https://doi.org/10.3390/jcdd9120423