
Recasting Current Knowledge of Human Fetal Circulation: The Importance of Computational Models

Abstract
:1. Introduction
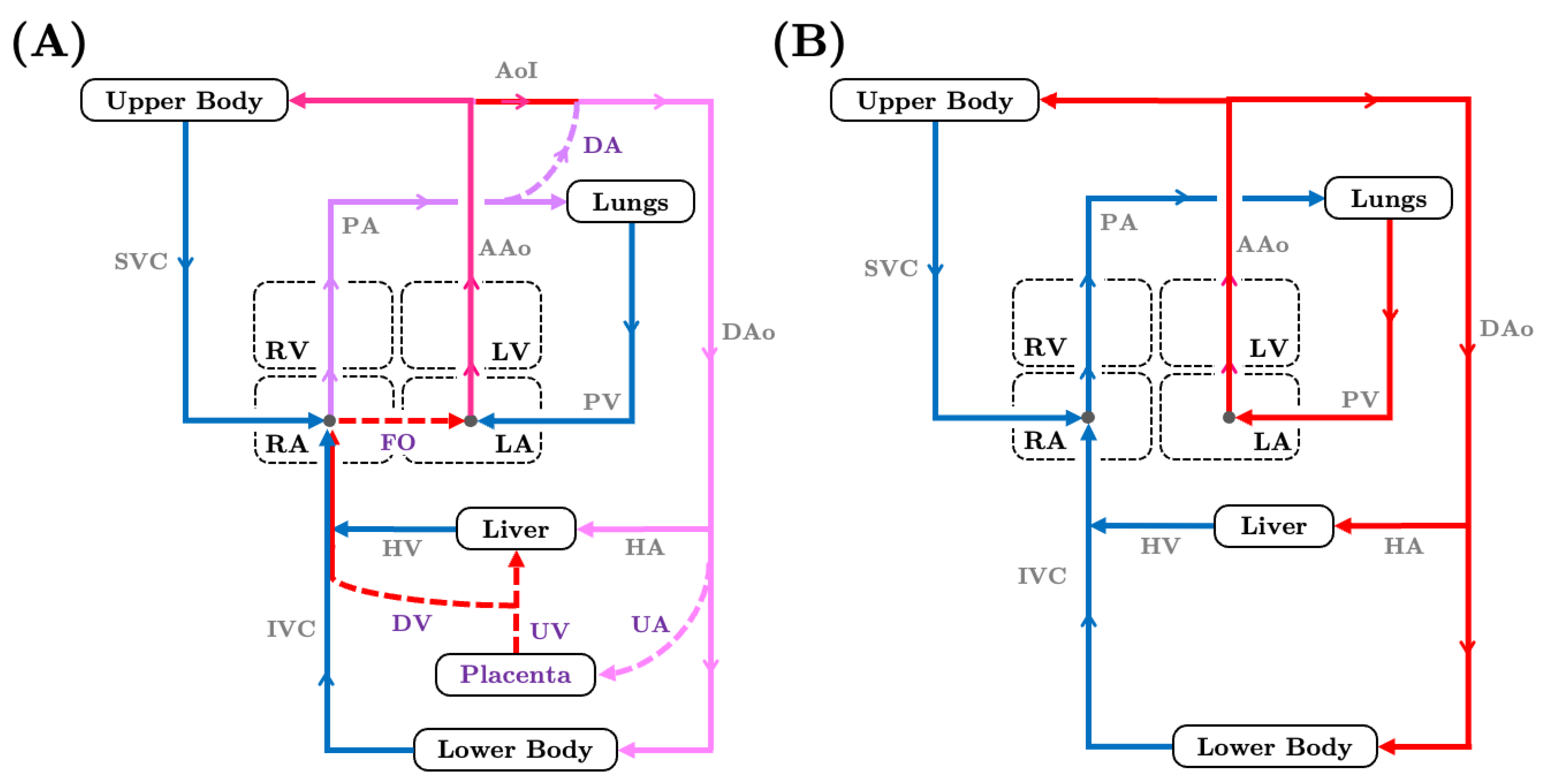
2. Fetal Cardiovascular Physiology
2.1. Landmark Experimental Investigations of Fetal Circulation
2.2. Characterizing Fetal Hemodynamics through Ultrasound and Magnetic Resonance Imaging
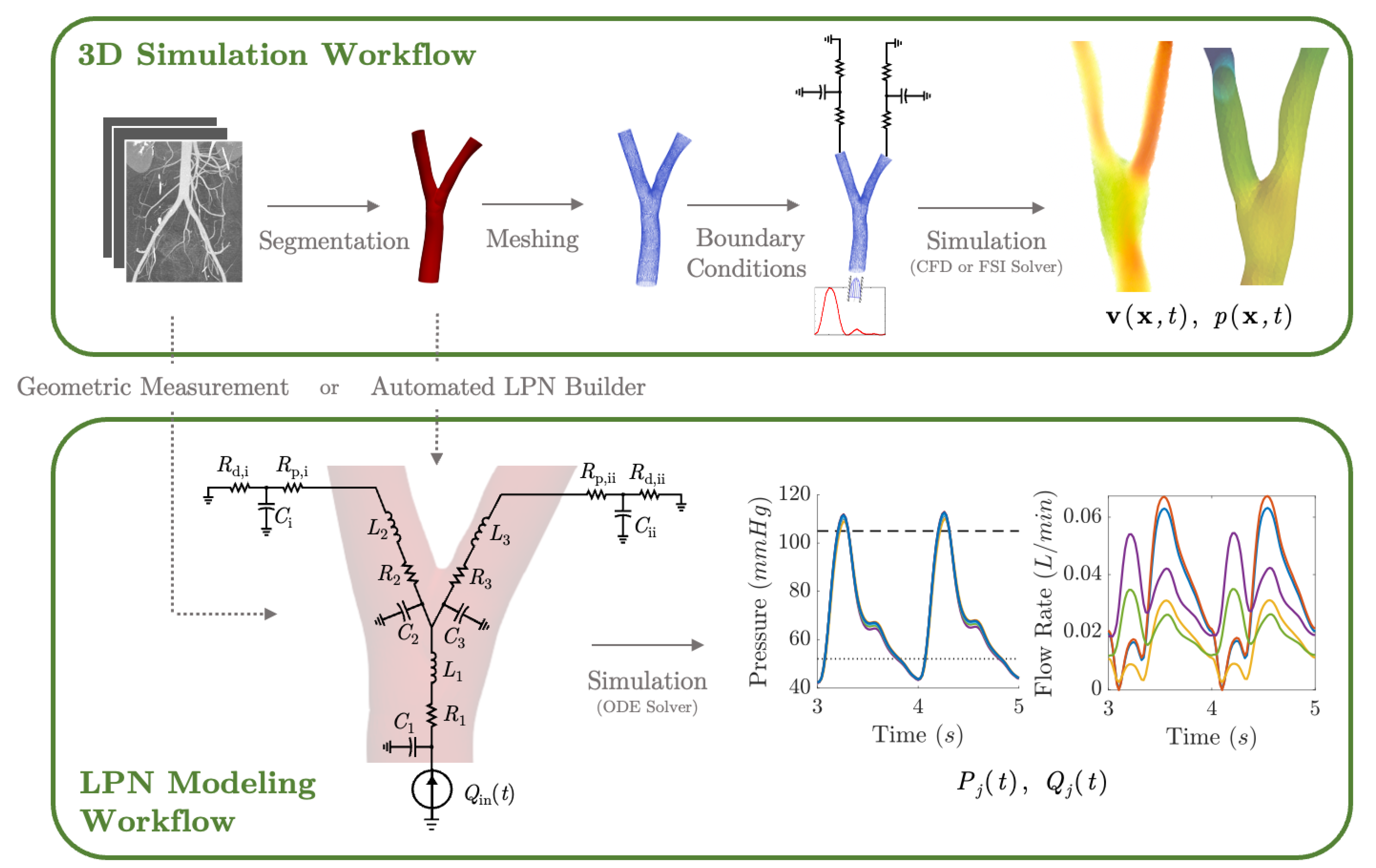
3. A Primer on Computational Hemodynamic Simulations
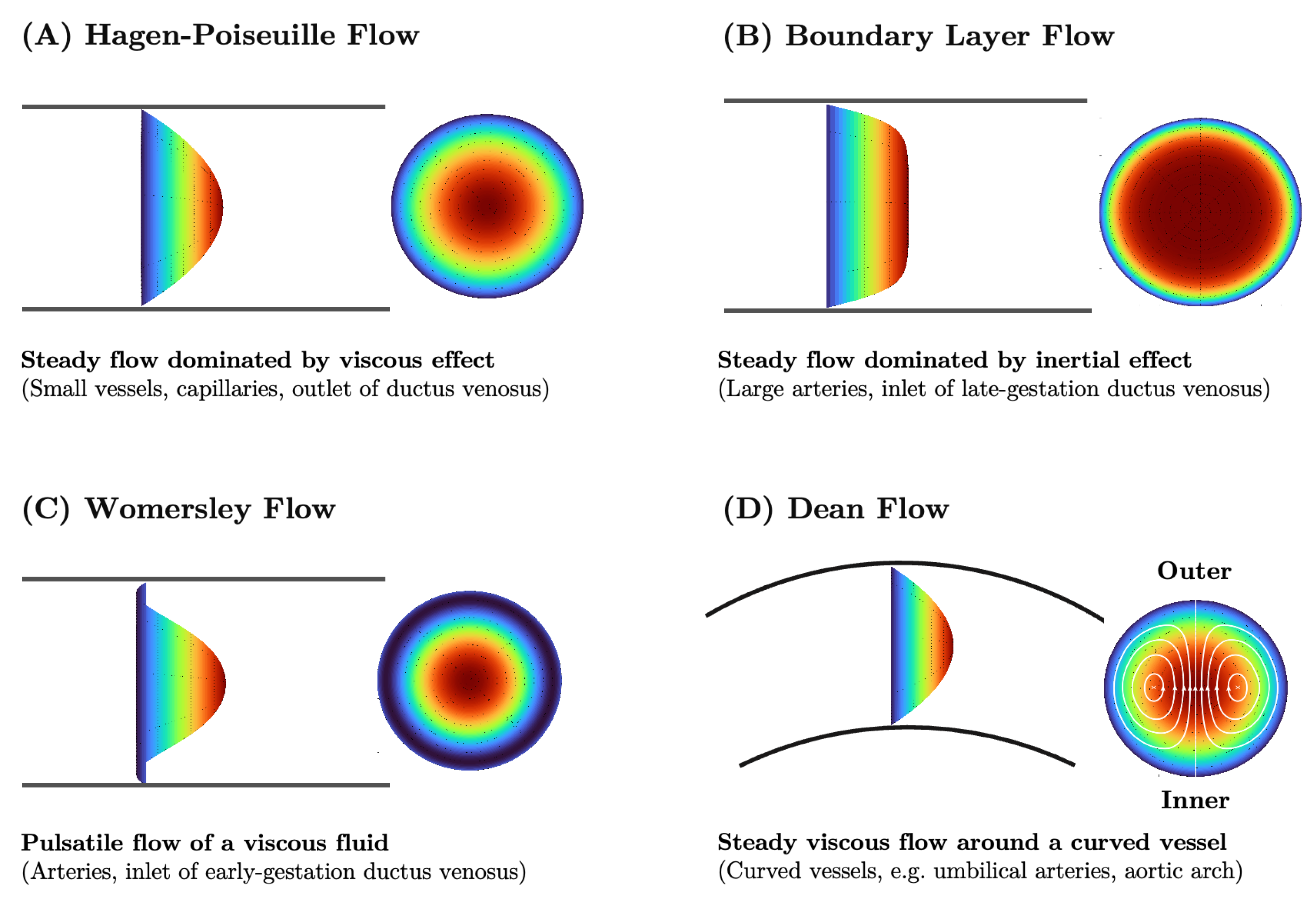
3.1. Fluid Mechanics Principles of Blood Flow
3.2. Three-Dimensional Hemodynamics Simulation
3.3. Lumped Parameter Networks
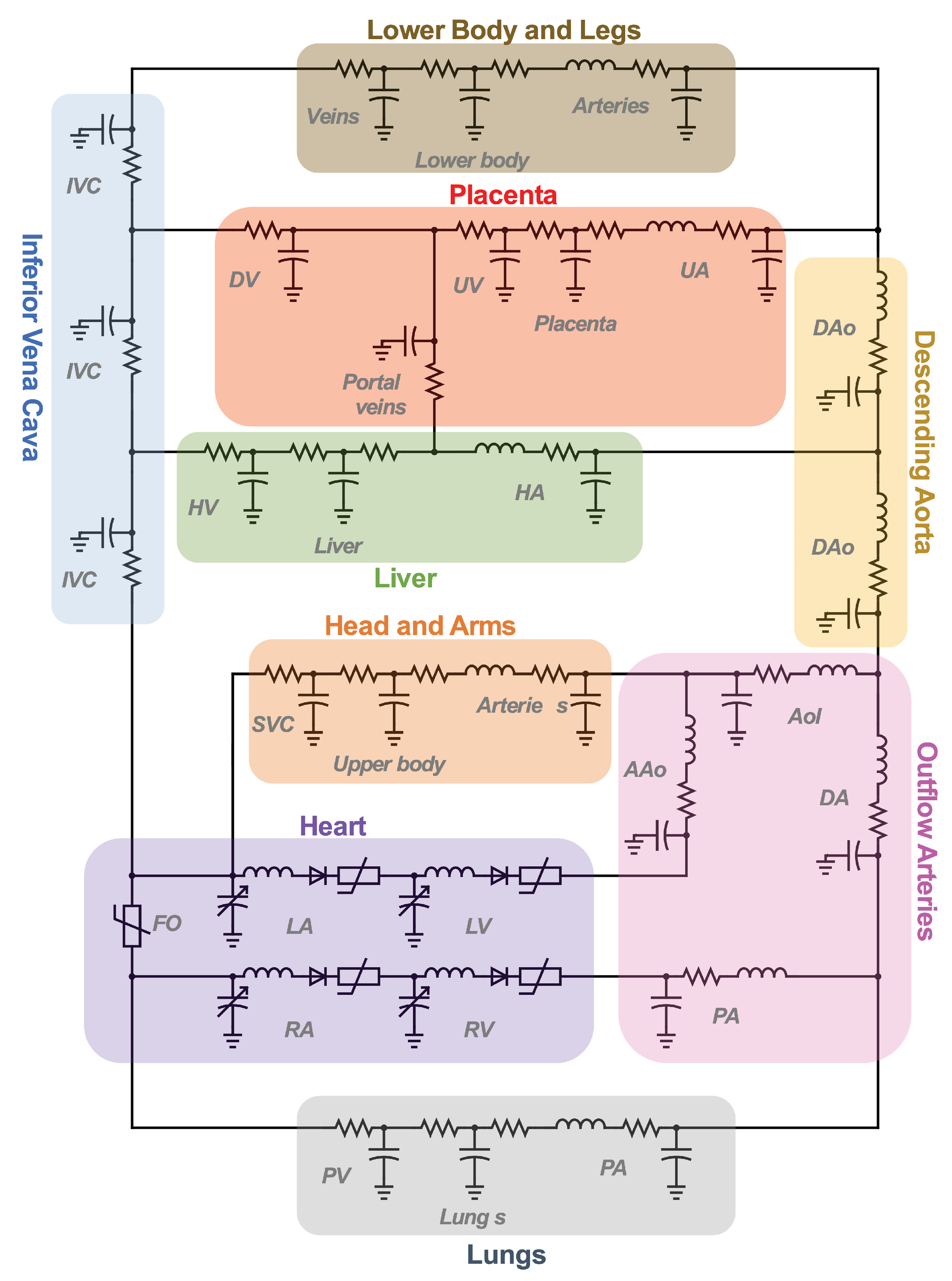
4. Lumped Parameter Models of Fetal Circulation
4.1. Evolution and Sophistication of Fetal Lumped Parameter Network Models
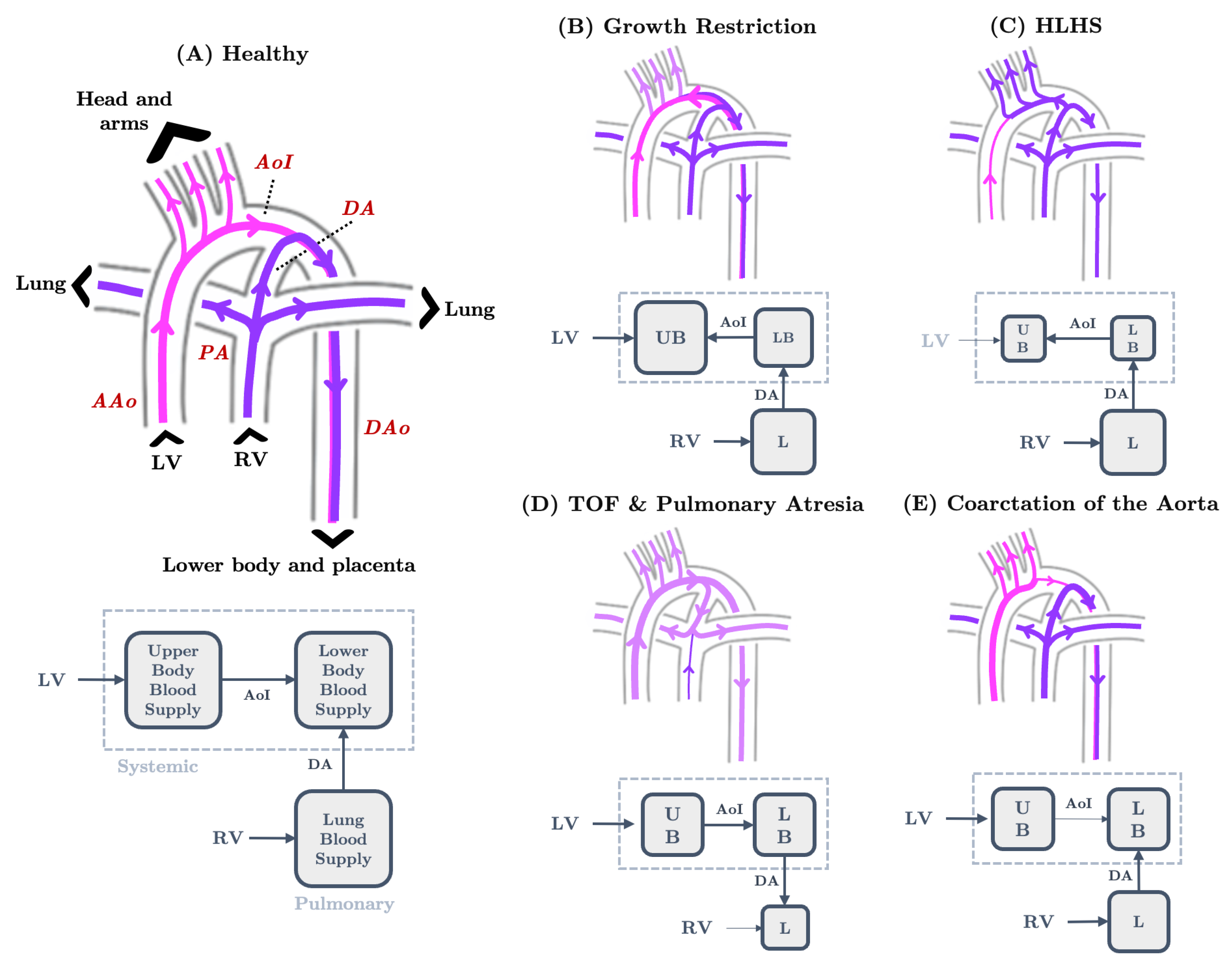
4.2. LPN Hemodynamic Models of Growth-Restricted Fetuses
4.3. Patient-Specific Parameter Estimation in LPN Models for Diagnosis
4.4. LPN Models of Transitional Hemodynamics in Neonates
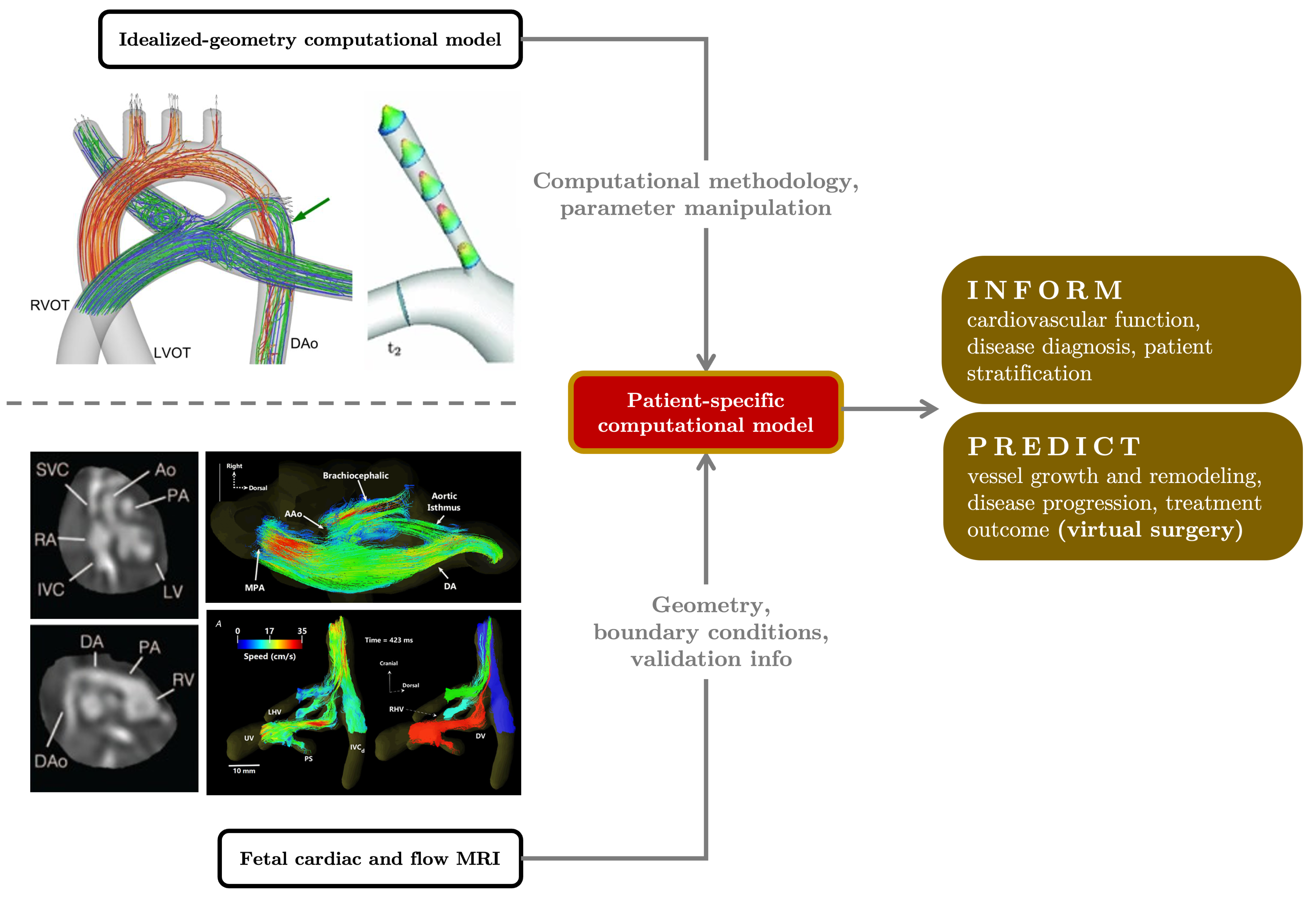
5. Local Hemodynamic Features of Fetal Circulation and 3D Simulations
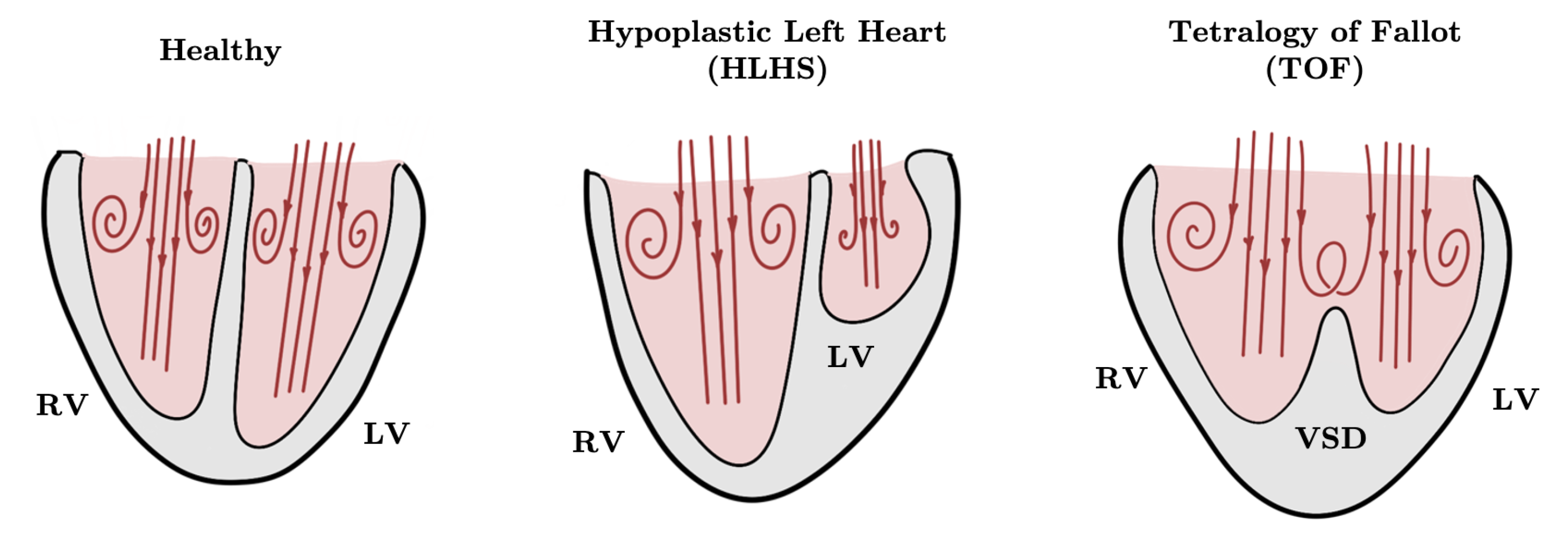
5.1. Blood Flow in Healthy Fetal Hearts and Fetal Hearts with CHDs
5.2. The Great Vessels and Cardiac Output Distribution
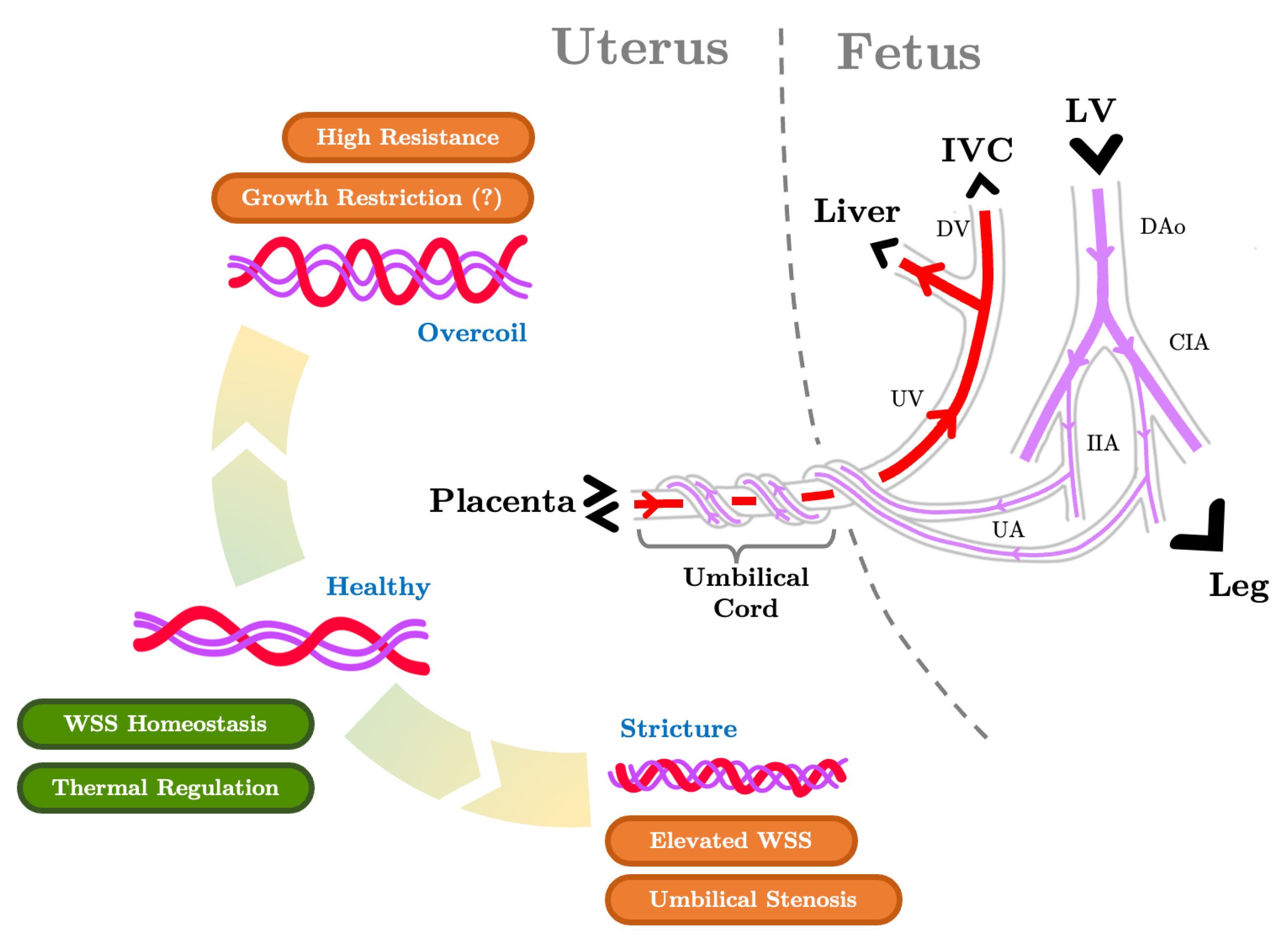
5.3. The Umbilical Arteries and the Role of their Helical Morphology
5.4. The Ductus Venosus and Placental Venous Return Distribution
6. Concluding Remarks and Future Directions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 3D | Three-dimensional |
| 4D | Four-dimensional |
| BC | Boundary condition |
| CCO | Combined cardiac output |
| CHD | Congenital heart defect |
| DCC | Delayed cord clamping |
| FSI | Fluid–structure interaction |
| FMDM | Fetus of mother with diabetes mellitus |
| HLHS | Hypoplastic left heart syndrome |
| ICC | Immediate cord clamping |
| IUGR | Intrauterine growth restriction |
| LPN | Lumped parameter network |
| MRI | Magnetic resonance imaging |
| PC-MRI | Phase contrast magnetic resonance imaging |
| TOF | Tetralogy of Fallot |
Appendix A. Oxygen Saturation in the Fetal Cardiovascular System

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oxygen Saturation (%) | Oxygen Saturation (%) | ||
|---|---|---|---|
| Left Atrium | 49 | Main Pulmonary Artery | 42 |
| Left Ventricle | 49 | Ductus Arteriosus | 42 |
| Right Atrium | 42 | Umbilical Arteries (Fetal End) | 43.7 |
| Right Ventricle | 42 | Umbilical Vein (Placental End) | 69.4 |
| Foramen Ovale | - | Ductus Venosus | 69.4 |
| Ascending Aorta | 49 | Thoracic Inferior Vena Cava | 49.9 |
| Carotid Arteries | 49 | ||
| Descending Aorta | 43.7 | Superior Vena Cava | 40.2 |
Appendix B. Characteristic Scales of Fetal Blood Flow
| Gestational Week | Peak Velocity (cm/s) | Diameter (cm) | Reynolds Number | |
|---|---|---|---|---|
| Mitral Valve 1 | 21 | 41 | 0.49 | 532 |
| 27 | 45 | 0.71 | 847 | |
| 35 | 47 | 0.93 | 1158 | |
| Tricuspic Valve 2 | 21 | 45 | 0.49 | 584 |
| 27 | 49 | 0.74 | 961 | |
| 35 | 51 | 1.00 | 1352 | |
| Ascending Aorta 3 | 20 | 55.6 | 0.33 | 487 |
| 24 | 71.0 | 0.43 | 809 | |
| 28 | 84.8 | 0.53 | 1191 | |
| 32 | 96.3 | 0.64 | 1633 | |
| 36 | 104.7 | 0.73 | 2026 | |
| 40 | 109.7 | 0.83 | 2413 | |
| Main Pulmonary Artery 4 | 20 | 55.1 | 0.36 | 526 |
| 24 | 64.6 | 0.48 | 822 | |
| 28 | 73.6 | 0.60 | 1170 | |
| 32 | 80.7 | 0.71 | 1518 | |
| 36 | 86.2 | 0.82 | 1873 | |
| 40 | 88.9 | 0.93 | 2190 | |
| Ductus Arteriosus 5 | 22 | 75.9 | 0.238 | 479 |
| 26 | 92.0 | 0.270 | 658 | |
| 30 | 107 | 0.304 | 862 | |
| 34 | 127 | 0.349 | 1175 | |
| Umbilical Artery 6 | 20 | 41.05 | 0.252 | 274 |
| 24 | 45.47 | 0.341 | 411 | |
| 28 | 48.92 | 0.402 | 521 | |
| 32 | 51.65 | 0.433 | 593 | |
| 36 | 53.93 | 0.447 | 639 | |
| 40 | 55.80 | 0.464 | 686 | |
| Ductus Venosus 7 | 20 | 49.3 | 0.143 | 187 |
| 24 | 55.7 | 0.152 | 224 | |
| 28 | 60.8 | 0.160 | 258 | |
| 32 | 62.8 | 0.168 | 280 | |
| 36 | 61.1 | 0.177 | 287 | |
| 40 | 55.2 | 0.189 | 276 |
References
- Rana, M.S.; Sizarov, A.; Christoffels, V.M.; Moorman, A.F. Development of the human aortic arch system captured in an interactive three-dimensional reference model. Am. J. Med. Genet. Part A 2014, 164, 1372–1383. [Google Scholar] [CrossRef] [PubMed]
- Yagel, S.; Kivilevitch, Z.; Cohen, S.M.; Valsky, D.V.; Messing, B.; Shen, O.; Achiron, R. The fetal venous system, Part I: Normal embryology, anatomy, hemodynamics, ultrasound evaluation and Doppler investigation. Ultrasound Obstet. Gynecol. 2010, 35, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Yee, K.; Lui, F. Anatomy, Thorax, Heart Foramen Ovale. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Lindsey, S.E.; Butcher, J.T.; Yalcin, H.C. Mechanical regulation of cardiac development. Front. Physiol. 2014, 5, 318. [Google Scholar] [CrossRef] [PubMed]
- Chew, L.C.; Verma, R.P. Fetal Growth Restriction; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Donofrio, M.; Bremer, Y.; Schieken, R.; Gennings, C.; Morton, L.; Eidem, B.; Cetta, F.; Falkensammer, C.; Huhta, J.; Kleinman, C. Autoregulation of cerebral blood flow in fetuses with congenital heart disease: The brain sparing effect. Pediatr. Cardiol. 2003, 24, 436–443. [Google Scholar] [CrossRef]
- Itsukaichi, M.; Kikuchi, A.; Yoshihara, K.; Serikawa, T.; Takakuwa, K.; Tanaka, K. Changes in Fetal Circulation Associated with Congenital Heart Disease and Their Effects on Fetal Growth. Fetal Diagn. Ther. 2011, 30, 219–224. [Google Scholar] [CrossRef]
- Krishna, U.; Bhalerao, S. Placental insufficiency and fetal growth restriction. J. Obstet. Gynecol. India 2011, 61, 505–511. [Google Scholar] [CrossRef]
- Miller, S.L.; Huppi, P.S.; Mallard, C. The consequences of fetal growth restriction on brain structure and neurodevelopmental outcome. J. Physiol. 2016, 594, 807–823. [Google Scholar] [CrossRef]
- McBride, K.L.; Zender, G.A.; Fitzgerald-Butt, S.M.; Koehler, D.; Menesses-Diaz, A.; Fernbach, S.; Lee, K.; Towbin, J.A.; Leal, S.; Belmont, J.W. Linkage analysis of left ventricular outflow tract malformations (aortic valve stenosis, coarctation of the aorta, and hypoplastic left heart syndrome). Eur. J. Hum. Genet. 2009, 17, 811–819. [Google Scholar] [CrossRef]
- Kiserud, T.; Acharya, G. The fetal circulation. Prenat. Diagn. 2004, 24, 1049–1059. [Google Scholar] [CrossRef] [PubMed]
- Kiserud, T. Physiology of the fetal circulation. Semin. Fetal Neonatal Med. 2005, 10, 493–503. [Google Scholar] [CrossRef]
- Lasheras, J.C. The biomechanics of arterial aneurysms. Annu. Rev. Fluid Mech. 2007, 39, 293–319. [Google Scholar] [CrossRef]
- Kanter, K.R.; Haggerty, C.M.; Restrepo, M.; de Zelicourt, D.A.; Rossignac, J.; Parks, W.J.; Yoganathan, A.P. Preliminary clinical experience with a bifurcated Y-graft Fontan procedure—A feasibility study. J. Thorac. Cardiovasc. Surg. 2012, 144, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.A.; Fonte, T.A.; Min, J.K. Computational Fluid Dynamics Applied to Cardiac Computed Tomography for Noninvasive Quantification of Fractional Flow Reserve. J. Am. Coll. Cardiol. 2013, 61, 2233–2241. [Google Scholar] [CrossRef] [PubMed]
- Poelma, C.; Watton, P.N.; Ventikos, Y. Transitional flow in aneurysms and the computation of haemodynamic parameters. J. R. Soc. Interface 2015, 12, 20141394. [Google Scholar] [CrossRef] [PubMed]
- Grande Gutiérrez, N.; Mathew, M.; McCrindle, B.W.; Tran, J.S.; Kahn, A.M.; Burns, J.C.; Marsden, A.L. Hemodynamic variables in aneurysms are associated with thrombotic risk in children with Kawasaki disease. Int. J. Cardiol. 2019, 281, 15–21. [Google Scholar] [CrossRef]
- Schwarz, E.L.; Kelly, J.M.; Blum, K.M.; Hor, K.N.; Yates, A.R.; Zbinden, J.C.; Verma, A.; Lindsey, S.E.; Ramachandra, A.B.; Szafron, J.M.; et al. Hemodynamic performance of tissue-engineered vascular grafts in Fontan patients. NPJ Regen. Med. 2021, 6, 38. [Google Scholar] [CrossRef]
- Taylor, C.A.; Draney, M.T.; Ku, J.P.; Parker, D.; Steele, B.N.; Wang, K.; Zarins, C.K. Predictive medicine: Computational techniques in therapeutic decision-making. Comput. Aided Surg. 1999, 4, 231–247. [Google Scholar] [CrossRef]
- Gundert, T.J.; Marsden, A.L.; Yang, W.; LaDisa, J.F., Jr. Optimization of Cardiovascular Stent Design Using Computational Fluid Dynamics. J. Biomech. Eng. 2012, 134, 011002. [Google Scholar] [CrossRef]
- Lindsey, S.E.; Menon, P.G.; Kowalski, W.J.; Shekhar, A.; Yalcin, H.C.; Nishimura, N.; Schaffer, C.B.; Butcher, J.T.; Pekkan, K. Growth and hemodynamics after early embryonic aortic arch occlusion. Biomech. Model. Mechanobiol. 2015, 14, 735–751. [Google Scholar] [CrossRef]
- Lashkarinia, S.S.; Piskin, S.; Bozkaya, T.A.; Salihoglu, E.; Yerebakan, C.; Pekkan, K. Computational pre-surgical planning of arterial patch reconstruction: Parametric limits and in vitro validation. Ann. Biomed. Eng. 2018, 46, 1292–1308. [Google Scholar] [CrossRef]
- Anbazhakan, S.; Rios Coronado, P.E.; Sy-Quia, A.N.L.; Seow, L.W.; Hands, A.M.; Zhao, M.; Dong, M.L.; Pfaller, M.R.; Amir, Z.A.; Raftrey, B.C.; et al. Blood flow modeling reveals improved collateral artery performance during the regenerative period in mammalian hearts. Nat. Cardiovasc. Res. 2022, 1, 775–790. [Google Scholar] [CrossRef]
- Garcia-Cañadilla, P.; Rudenick, P.A.; Crispi, F.; Cruz-Lemini, M.; Palau, G.; Camara, O.; Gratacos, E.; Bijens, B.H. A Computational Model of the Fetal Circulation to Quantify Blood Redistribution in Intrauterine Growth Restriction. PLoS Comput. Biol. 2014, 10, e1003667. [Google Scholar] [CrossRef]
- Wong, H.S.; Wiputra, H.; Tulzer, A.; Tulzer, G.; Yap, C.H. Fluid Mechanics of Fetal Left Ventricle During Aortic Stenosis with Evolving Hypoplastic Left Heart Syndrome. Ann. Biomed. Eng. 2022, 50, 1158–1172. [Google Scholar] [CrossRef]
- Wiputra, H.; Chen, C.K.; Talbi, E.; Lim, G.L.; Soomar, S.M.; Biswas, A.; Mattar, C.N.Z.; Bark, D.; Leo, H.L.; Yap, C.H. Human fetal hearts with tetralogy of Fallot have altered fluid dynamics and forces. Am. J. Physiol.-Heart Circ. Physiol. 2018, 315, H1649–H1659. [Google Scholar] [CrossRef] [PubMed]
- Pekkan, K.; Dasi, L.P.; Nourparvar, P.; Yerneni, S.; Tobita, K.; Fogel, M.A.; Keller, B.; Yoganathan, A. In vitro hemodynamic investigation of the embryonic aortic arch at late gestation. J. Biomech. 2008, 41, 1697–1706. [Google Scholar] [CrossRef] [PubMed]
- Langille, B.L. Arterial remodeling: Relation to hemodynamics. Can. J. Physiol. Pharmacol. 1996, 74, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Sedmera, D.; Hu, N.; Weiss, K.M.; Keller, B.B.; Denslow, S.; Thompson, R.P. Cellular changes in experimental left heart hypoplasia. Anat. Rec. 2002, 267, 137–145. [Google Scholar] [CrossRef]
- Li, Y.S.J.; Haga, J.H.; Chien, S. Molecular basis of the effects of shear stress on vascular endothelial cells. J. Biomech. 2005, 38, 1949–1971. [Google Scholar] [CrossRef]
- Lindsey, S.E.; Butcher, J.T.; Vignon-Clementel, I.E. Cohort-based multiscale analysis of hemodynamic-driven growth and remodeling of the embryonic pharyngeal arch arteries. Development 2018, 145, dev162578. [Google Scholar] [CrossRef]
- Salman, H.E.; Alser, M.; Shekhar, A.; Gould, R.A.; Benslimane, F.M.; Butcher, J.T.; Yalcin, H.C. Effect of left atrial ligation-driven altered inflow hemodynamics on embryonic heart development: Clues for prenatal progression of hypoplastic left heart syndrome. Biomech. Model. Mechanobiol. 2021, 20, 733–750. [Google Scholar] [CrossRef]
- Swanson, A.; David, A. Animal models of fetal growth restriction: Considerations for translational medicine. Placenta 2015, 36, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Everett, N.B.; Johnson, R.J. Use of Radioactive Phosphorus in Studies of Fetal Circulation. Am. J. Physiol.-Leg. Content 1950, 162, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.Q.; Cahill, L.S.; Wong, M.D.; Seed, M.; Macgowan, C.K.; Sled, J.G. Assessment of flow distribution in the mouse fetal circulation at late gestation by high-frequency Doppler ultrasound. Physiol. Genom. 2014, 46, 602–614. [Google Scholar] [CrossRef] [PubMed]
- Salman, H.E.; Yalcin, H.C. Computational Modeling of Blood Flow Hemodynamics for Biomechanical Investigation of Cardiac Development and Disease. J. Cardiovasc. Dev. Dis. 2021, 8, 14. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, W.J.; Teslovich, N.C.; Menon, P.G.; Tinney, J.P.; Keller, B.B.; Pekkan, K. Left atrial ligation alters intracardiac flow patterns and the biomechanical landscape in the chick embryo. Dev. Dyn. 2014, 243, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, S.E.; Vignon-Clementel, I.E.; Butcher, J.T. Assessing early cardiac outflow tract adaptive responses through combined experimental-computational manipulations. Ann. Biomed. Eng. 2021, 49, 3227–3242. [Google Scholar] [CrossRef]
- Jensen, O.E.; Chernyavsky, I.L. Blood Flow and Transport in the Human Placenta. Annu. Rev. Fluid Mech. 2019, 51, 25–47. [Google Scholar] [CrossRef]
- Kiserud, T. The ductus venosus. Semin. Perinatol. 2001, 25, 11–20. [Google Scholar] [CrossRef]
- Markl, M.; Frydrychowicz, A.; Kozerke, S.; Hope, M.; Wieben, O. 4D flow MRI. J. Magn. Reson. Imaging 2012, 36, 1015–1036. [Google Scholar] [CrossRef]
- Sun, L.; Macgowan, C.K.; Portnoy, S.; Sled, J.G.; Yoo, S.J.; Grosse-Wortmann, L.; Jaeggi, E.; Kingdom, J.; Seed, M. New advances in fetal cardiovascular magnetic resonance imaging for quantifying the distribution of blood flow and oxygen transport: Potential applications in fetal cardiovascular disease diagnosis and therapy. Echocardiography 2017, 34, 1799–1803. [Google Scholar] [CrossRef]
- Mielke, G.; Benda, N. Cardiac Output and Central Distribution of Blood Flow in the Human Fetus. Circulation 2001, 103, 1662–1668. [Google Scholar] [CrossRef] [PubMed]
- Murphy, P.J. The fetal circulation. Contin. Educ. Anaesth. Crit. Care Pain 2005, 5, 107–112. [Google Scholar] [CrossRef]
- Strong, T.H.; Jarles, D.L.; Vega, J.S.; Feldman, D.B. The umbilical coiling index. Am. J. Obstet. Gynecol. 1994, 170, 29–32. [Google Scholar] [CrossRef]
- Predanic, M.; Perni, S.C.; Chervenak, F.A. Antenatal umbilical coiling index and Doppler flow characteristics. Ultrasound Obstet. Gynecol. 2006, 28, 699–703. [Google Scholar] [CrossRef]
- Sharma, B.; Bhardwaj, N.; Gupta, S.; Gupta, P.K.; Verma, A.; Malviya, K. Association of umbilical coiling index by colour Doppler ultrasonography at 18–22 weeks of gestation and perinatal outcome. J. Obstet. Gynecol. India 2012, 62, 650–654. [Google Scholar] [CrossRef] [PubMed]
- Devaru, D.; Thusoo, M. Umbilical coiling index & the perinatal outcome. J. Obstet. Gynecol. India 2012, 62, 43–46. [Google Scholar]
- Pergialiotis, V.; Kotrogianni, P.; Koutaki, D.; Christopoulos-Timogiannakis, E.; Papantoniou, N.; Daskalakis, G. Umbilical cord coiling index for the prediction of adverse pregnancy outcomes: A meta-analysis and sequential analysis. J. Matern.-Fetal Neonatal Med. 2020, 33, 4022–4029. [Google Scholar] [CrossRef]
- Peng, H.Q.; Smith-Levitin, M.; Rochelson, B.; Kahn, E. Umbilical cord stricture and overcoiling are common causes of fetal demise. Pediatr. Dev. Pathol. 2006, 9, 14–19. [Google Scholar] [CrossRef]
- Gilbert, R.D.; Schroder, H.; Kawamura, T.; Dale, P.S.; Power, G.G. Heat transfer pathways between fetal lamb and ewe. J. Appl. Physiol. 1985, 59, 634–638. [Google Scholar] [CrossRef]
- Schrauben, E.M.; Saini, B.S.; Darby, J.R.; Soo, J.Y.; Lock, M.C.; Stirrat, E.; Stortz, G.; Sled, J.G.; Morrison, J.L.; Seed, M.; et al. Fetal hemodynamics and cardiac streaming assessed by 4D flow cardiovascular magnetic resonance in fetal sheep. J. Cardiovasc. Magn. Reson. 2019, 21, 8. [Google Scholar] [CrossRef]
- Finnemore, A.; Groves, A. Physiology of the fetal and transitional circulation. Semin. Fetal Neonatal Med. 2015, 20, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Mott, J.C. Control of the foetal circulation. J. Exp. Biol. 1982, 100, 129–146. [Google Scholar] [CrossRef]
- Nuwayhid, B.; Brinkman, C., III; Su, C.; Bevan, J.; Assali, N. Development of autonomic control of fetal circulation. Am. J. Physiol.-Leg. Content 1975, 228, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E.; Baerts, W.; van Bel, F. Brain-sparing in intrauterine growth restriction: Considerations for the neonatologist. Neonatology 2015, 108, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Baschat, A.A. The fetal circulation and essential organs—a new twist to an old tale. Ultrasound Obstet. Gynecol. 2006, 27, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Peleg, D.; Kennedy, C.M.; Hunter, S.K. Intrauterine growth restriction: Identification and management. Am. Fam. Physician 1998, 58, 453. [Google Scholar] [PubMed]
- Pohlman, A.G. The course of the blood through the heart of the fetal mammal, with a note on the reptilian and amphibian circulations. Anat. Rec. 1909, 3, 75–109. [Google Scholar] [CrossRef]
- Rudolph, A.M.; Heymann, M.A. The Circulation of the Fetus in Utero. Circ. Res. 1967, 21, 163–184. [Google Scholar] [CrossRef]
- Barclay, A.E.; Barcroft, J.; Barron, D.H.; Franklin, K.J. A Radiographic Demonstration of the Circulation through the Heart in the Adult and in the Fœtus, and the Identification of the Ductus Arteriosus. Br. J. Radiol. 1939, 12, 505–517. [Google Scholar] [CrossRef]
- Harada, K.; Rice, M.J.; Shiota, T.; Ishii, M.; McDonald, R.W.; Reller, M.D.; Sahn, D.J. Gestational age-and growth-related alterations in fetal right and left ventricular diastolic filling patterns. Am. J. Cardiol. 1997, 79, 173–177. [Google Scholar] [CrossRef]
- Bahlmann, F.; Wellek, S.; Reinhardt, I.; Krummenauer, F.; Merz, E.; Welter, C. Reference values of fetal aortic flow velocity waveforms and associated intra-observer reliability in normal pregnancies. Ultrasound Obstet. Gynecol. Off. J. Int. Soc. Ultrasound Obstet. Gynecol. 2001, 17, 42–49. [Google Scholar] [CrossRef]
- Mielke, G.; Benda, N. Blood flow velocity waveforms of the fetal pulmonary artery and the ductus arteriosus: Reference ranges from 13 weeks to term. Ultrasound Obstet. Gynecol. 2000, 15, 213–218. [Google Scholar] [CrossRef]
- Acharya, G.; Wilsgaard, T.; Berntsen, G.; Maltau, J.; Kiserud, T. Reference ranges for serial measurements of blood velocity and pulsatility index at the intra-abdominal portion, and fetal and placental ends of the umbilical artery. Ultrasound Obstet. Gynecol. Off. J. Int. Soc. Ultrasound Obstet. Gynecol. 2005, 26, 162–169. [Google Scholar] [CrossRef]
- Zytoon, A.A.; El-Abakawy, N.N.A.; Hassanein, S.A.h. Reference values for ductus venosus flow in normal gestation among an Egyptian population. Egypt. J. Radiol. Nucl. Med. 2020, 51, 7. [Google Scholar] [CrossRef]
- Sharland, G.; Allan, L. Normal fetal cardiac measurements derived by cross-sectional echocardiography. Ultrasound Obstet. Gynecol. Off. J. Int. Soc. Ultrasound Obstet. Gynecol. 1992, 2, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Szpinda, M.; Szwesta, A.; Szpinda, E. Morphometric study of the ductus arteriosus during human development. Ann. Anat.-Anat. Anz. 2007, 189, 47–52. [Google Scholar] [CrossRef]
- Barbieri, C.; Cecatti, J.; Surita, F.; Marussi, E.; Costa, J. Sonographic measurement of the umbilical cord area and the diameters of its vessels during pregnancy. J. Obstet. Gynaecol. 2012, 32, 230–236. [Google Scholar] [CrossRef]
- FitzGerald, D.E.; Drumm, J.E. Non-invasive measurement of human fetal circulation using ultrasound: A new method. BMJ 1977, 2, 1450–1451. [Google Scholar] [CrossRef]
- Campbell, S.; Vyas, S.; Nicolaides, K.H. Doppler investigation of the fetal circulation. J. Perinat. Med. 1991, 19, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Molina, F.; Faro, C.; Sotiriadis, A.; Dagklis, T.; Nicolaides, K. Heart stroke volume and cardiac output by four-dimensional ultrasound in normal fetuses. Ultrasound Obstet. Gynecol. 2008, 32, 181–187. [Google Scholar] [CrossRef]
- Abduljalil, K.; Pan, X.; Clayton, R.; Johnson, T.N.; Jamei, M. Fetal physiologically based pharmacokinetic models: Systems information on fetal cardiac output and its distribution to different organs during development. Clin. Pharmacokinet. 2021, 60, 741–757. [Google Scholar] [CrossRef]
- Hecher, K.; Campbell, S.; Doyle, P.; Harrington, K.; Nicolaides, K. Assessment of Fetal Compromise by Doppler Ultrasound Investigation of the Fetal Circulation. Circulation 1995, 91, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Harrington, K.; Carpenter, R.G.; Nguyen, M.; Campbell, S. Changes observed in Doppler studies of the fetal circulation in pregnancies complicated by pre-eclampsia or the delivery of a small-for-gestational-age baby. I. Cross-sectional analysis. Ultrasound Obstet. Gynecol. 1995, 6, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Harrington, K.; Thompson, M.O.; Carpenter, R.G.; Nguyen, M.; Campbell, S. Doppler fetal circulation in pregnancies complicated by pre-eclampsia or delivery of a small for gestational age baby: 2. Longitudinal analysis. BJOG Int. J. Obstet. Gynaecol. 1999, 106, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Hecher, K.; Ville, Y.; Snijders, R.; Nicolaides, K. Doppler studies of the fetal circulation in twin–twin transfusion syndrome. Ultrasound Obstet. Gynecol. 1995, 5, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Transition from fetal to neonatal circulation: Modeling the effect of umbilical cord clamping. J. Biomech. 2015, 48, 1662–1670. [CrossRef]
- Ranke, C.; Hendrickx, P.; Roth, U.; Brassel, F.; Creutzig, A.; Alexander, K. Color and conventional image-directed Doppler ultrasonography: Accuracy and sources of error in quantitative blood flow measurements. J. Clin. Ultrasound 1992, 20, 187–193. [Google Scholar] [CrossRef]
- Saleem, S.N. Feasibility of MRI of the Fetal Heart with Balanced Steady-State Free Precession Sequence Along Fetal Body and Cardiac Planes. Am. J. Roentgenol. 2008, 191, 1208–1215. [Google Scholar] [CrossRef] [PubMed]
- Dashe, J.S.; McIntire, D.D.; Twickler, D.M. Maternal obesity limits the ultrasound evaluation of fetal anatomy. J. Ultrasound Med. 2009, 28, 1025–1030. [Google Scholar] [CrossRef]
- Lloyd, D.F.; Pushparajah, K.; Simpson, J.M.; Van Amerom, J.F.; Van Poppel, M.P.; Schulz, A.; Kainz, B.; Deprez, M.; Lohezic, M.; Allsop, J.; et al. Three-dimensional visualisation of the fetal heart using prenatal MRI with motion-corrected slice-volume registration: A prospective, single-centre cohort study. Lancet 2019, 393, 1619–1627. [Google Scholar] [CrossRef]
- van Amerom, J.F.; Lloyd, D.F.; Deprez, M.; Price, A.N.; Malik, S.J.; Pushparajah, K.; van Poppel, M.P.; Rutherford, M.A.; Razavi, R.; Hajnal, J.V. Fetal whole-heart 4D imaging using motion-corrected multi-planar real-time MRI. Magn. Reson. Med. 2019, 82, 1055–1072. [Google Scholar] [CrossRef] [PubMed]
- Roy, C.W.; van Amerom, J.F.; Marini, D.; Seed, M.; Macgowan, C.K. Fetal cardiac MRI: A review of technical advancements. Top. Magn. Reson. Imaging 2019, 28, 235. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Marini, D.; Saini, B.; Schrauben, E.; Macgowan, C.K.; Seed, M. Understanding Fetal Hemodynamics Using Cardiovascular Magnetic Resonance Imaging. Fetal Diagn. Ther. 2020, 47, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Seed, M.; van Amerom, J.F.P.; Yoo, S.J.; Bahiyah Al Nafisi, L.G.W.; Jaeggi, E.; Jansz, M.S.; Macgowan, C.K. Feasibility of quantification of the distribution of blood flow in the normal human fetal circulation using CMR: A cross-sectional study. J. Cardiovasc. Magn. Reson. 2012, 14, 79. [Google Scholar] [CrossRef]
- Goolaub, D.S.; Xu, J.; Schrauben, E.M.; Marini, D.; Kingdom, J.C.; Sled, J.G.; Seed, M.; Macgowan, C.K. Volumetric Fetal Flow Imaging with Magnetic Resonance Imaging. IEEE Trans. Med. Imaging 2022, 41, 2941–2952. [Google Scholar] [CrossRef]
- Prsa, M.; Sun, L.; van Amerom, J.; Yoo, S.J.; Grosse-Wortmann, L.; Jaeggi, E.; Macgowan, C.; Seed, M. Reference Ranges of Blood Flow in the Major Vessels of the Normal Human Fetal Circulation at Term by Phase-Contrast Magnetic Resonance Imaging. Circ. Cardiovasc. Imaging 2014, 7, 663–670. [Google Scholar] [CrossRef]
- Saini, B.S.; Darby, J.R.; Portnoy, S.; Sun, L.; van Amerom, J.; Lock, M.C.; Soo, J.Y.; Holman, S.L.; Perumal, S.R.; Kingdom, J.C.; et al. Normal human and sheep fetal vessel oxygen saturations by T2 magnetic resonance imaging. J. Physiol. 2020, 598, 3259–3281. [Google Scholar] [CrossRef]
- Batchelor, G.K. An Introduction to Fluid Dynamics; Cambridge Mathematical Library, Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Secomb, T.W. Blood Flow in the Microcirculation. Annu. Rev. Fluid Mech. 2017, 49, 443–461. [Google Scholar] [CrossRef]
- Al-Roubaie, S.; Jahnsen, E.D.; Mohammed, M.; Henderson-Toth, C.; Jones, E.A.V. Rheology of embryonic avian blood. Am. J. Physiol.-Heart Circ. Physiol. 2011, 301, H2473–H2481. [Google Scholar] [CrossRef]
- Gonzalo, A.; García-Villalba, M.; Rossini, L.; Durán, E.; Vigneault, D.; Martínez-Legazpi, P.; Flores, O.; Bermejo, J.; McVeigh, E.; Kahn, A.M.; et al. Non-Newtonian blood rheology impacts left atrial stasis in patient-specific simulations. Int. J. Numer. Methods Biomed. Eng. 2022, 38, e3597. [Google Scholar] [CrossRef]
- Reymond, P.; Crosetto, P.; Deparis, S.; Quarteroni, A.; Stergiopulos, N. Physiological simulation of blood flow in the aorta: Comparison of hemodynamic indices as predicted by 3-D FSI, 3-D rigid wall and 1-D models. Med. Eng. Phys. 2013, 35, 784–791. [Google Scholar] [CrossRef]
- Sengupta, D.; Kahn, A.M.; Kung, E.; Esmaily Moghadam, M.; Shirinsky, O.; Lyskina, G.A.; Burns, J.C.; Marsden, A.L. Thrombotic risk stratification using computational modeling in patients with coronary artery aneurysms following Kawasaki disease. Biomech. Model. Mechanobiol. 2014, 13, 1261–1276. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Zhu, L.; Huo, Y.; Liu, Y.; Kassab, G.S. Fluid–structure interaction (FSI) modeling in the cardiovascular system. In Computational Cardiovascular Mechanics; Springer: Berlin/Heidelberg, Germany, 2010; pp. 141–157. [Google Scholar]
- Hirschhorn, M.; Tchantchaleishvili, V.; Stevens, R.; Rossano, J.; Throckmorton, A. Fluid–structure interaction modeling in cardiovascular medicine – A systematic review 2017–2019. Med Eng. Phys. 2020, 78, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.J.R.; Liu, W.K.; Zimmermann, T.K. Lagrangian-Eulerian finite element formulation for incompressible viscous flows. Comput. Methods Appl. Mech. Eng. 1981, 29, 329–349. [Google Scholar] [CrossRef]
- Figueroa, C.A.; Vignon-Clementel, I.E.; Jansen, K.E.; Hughes, T.J.R.; Taylor, C.A. A coupled momentum method for modeling blood flow in three-dimensional deformable arteries. Comput. Methods Appl. Mech. Eng. 2006, 195, 5685–5706. [Google Scholar] [CrossRef]
- Womersley, J.R. An Elastic Tube Theory of Pulse Transmission and Oscillatory Flow in Mammalian Arteries; Technical Report; Aerospace Research Labs: Wright-Patterson AFB, OH, USA, 1957. [Google Scholar]
- Cheynet, E. Pcolor in Polar Coordinates: Version 3.11. 2021. MATLAB Central File Exchange. Available online: https://www.mathworks.com/matlabcentral/fileexchange/49040-pcolor-in-polar-coordinates (accessed on 14 February 2023).
- Alastruey, J.; Siggers, J.H.; Peiffer, V.; Doorly, D.J.; Sherwin, S.J. Reducing the data: Analysis of the role of vascular geometry on blood flow patterns in curved vessels. Phys. Fluids 2012, 24, 031902. [Google Scholar] [CrossRef]
- Larsson, S.; Thomée, V. Partial Differential Equations with Numerical Methods; Springer: Berlin/Heidelberg, Germany, 2003; Volume 45. [Google Scholar]
- Pfaller, M.R.; Pham, J.; Verma, A.; Pegolotti, L.; Wilson, N.M.; Parker, D.W.; Yang, W.; Marsden, A.L. Automated generation of 0D and 1D reduced-order models of patient-specific blood flow. Int. J. Numer. Methods Biomed. Eng. 2022, 38, e3639. [Google Scholar] [CrossRef]
- Wilson, N.M.; Arko, F.R.; Taylor, C.A. Predicting changes in blood flow in patient-specific operative plans for treating aortoiliac occlusive disease. Comput. Aided Surg. 2005, 10, 257–277. [Google Scholar] [CrossRef]
- Updegrove, A.; Wilson, N.M.; Merkow, J.; Lan, H.; Marsden, A.L.; Shadden, S.C. SimVascular: An open source pipeline for cardiovascular simulation. Ann. Biomed. Eng. 2017, 45, 525–541. [Google Scholar] [CrossRef]
- Vignon-Clementel, I.E.; Figueroa, C.A.; Jansen, K.E.; Taylor, C.A. Outflow boundary conditions for three-dimensional finite element modeling of blood flow and pressure in arteries. Comput. Methods Appl. Mech. Eng. 2006, 195, 3776–3796. [Google Scholar] [CrossRef]
- Vignon-Clementel, I.E.; Figueroa, C.A.; Jansen, K.E.; Taylor, C.A. Outflow boundary conditions for 3D simulations of non-periodic blood flow and pressure fields in deformable arteries. Comput. Methods Biomech. Biomed. Eng. 2010, 13, 625–640. [Google Scholar] [CrossRef] [PubMed]
- Coogan, J.S.; Humphrey, J.D.; Figueroa, C.A. Computational simulations of hemodynamic changes within thoracic, coronary, and cerebral arteries following early wall remodeling in response to distal aortic coarctation. Biomech. Model. Mechanobiol. 2013, 12, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, D.; Kahn, A.M.; Burns, J.C.; Sankaran, S.; Shadden, S.C.; Marsden, A.L. Image-based modeling of hemodynamics in coronary artery aneurysms caused by Kawasaki disease. Biomech. Model. Mechanobiol. 2012, 11, 915–932. [Google Scholar] [CrossRef] [PubMed]
- Miliić, V.; Quarteroni, A. Analysis of lumped parameter models for blood flow simulations and their relation with 1D models. Esaim Math. Model. Numer. Anal. 2004, 38, 613–632. [Google Scholar] [CrossRef]
- Karimi, A.; Sera, T.; Kudo, S.; Navidbakhsh, M. Experimental verification of the healthy and atherosclerotic coronary arteries incompressibility via Digital Image Correlation. Artery Res. 2016, 16, 1–7. [Google Scholar] [CrossRef]
- Pennati, G.; Migliavacca, F.; Dubini, G.; Pietrabissa, R.; de Leval, M.R. A mathematical model of circulation in the presence of the bidirectional cavopulmonary anastomosis in children with a univentricular heart. Med Eng. Phys. 1997, 19, 223–234. [Google Scholar] [CrossRef]
- Pennati, G.; Fumero, R. Scaling approach to study the changes through the gestation of human fetal cardiac and circulatory behaviors. Ann. Biomed. Eng. 2000, 28, 442–452. [Google Scholar] [CrossRef]
- Shimizu, S.; Une, D.; Kawada, T.; Hayama, Y.; Kamiya, A.; Shishido, T.; Sugimachi, M. Lumped parameter model for hemodynamic simulation of congenital heart diseases. J. Physiol. Sci. 2018, 68, 103–111. [Google Scholar] [CrossRef]
- Duanmu, Z.; Yin, M.; Fan, X.; Yang, X.; Luo, X. A patient-specific lumped-parameter model of coronary circulation. Sci. Rep. 2018, 8, 874. [Google Scholar] [CrossRef]
- Spilker, R.L.; Taylor, C.A. Tuning multidomain hemodynamic simulations to match physiological measurements. Ann. Biomed. Eng. 2010, 38, 2635–2648. [Google Scholar] [CrossRef]
- Ménigault, E.; Vieyres, P.; Lepoivre, B.; Durand, A.; Pourcelot, L.; Berson, M. Fetal heart modelling based on a pressure-volume relationship. Med Biol. Eng. Comput. 1997, 35, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Garber, L.; Khodaei, S.; Keshavarz-Motamed, Z. The critical role of lumped parameter models in patient-specific cardiovascular simulations. Arch. Comput. Methods Eng. 2022, 29, 2977–3000. [Google Scholar] [CrossRef]
- Moin, P. Fundamentals of Engineering Numerical Analysis; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Pennati, G.; Bellotti, M.; Fumero, R. Mathematical modelling of the human foetal cardiovascular system based on Doppler ultrasound data. Med Eng. Phys. 1997, 19, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Lingman, G.; Maršal, K. Fetal central blood circulation in the third trimester of normal pregnancy-a longitudinal study. I. Aortic and umbilical blood flow. Early Hum. Dev. 1986, 13, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Guettouche, A.; Papapanayotou, C.; Cherruault, Y.; Azancot-Benisty, A.; Challier, J. Optimization and resolution algorithm of the human fetal blood circulation model. Math. Comput. Model. 1993, 18, 1–8. [Google Scholar] [CrossRef]
- Sutton, M.; Groves, A.; MacNeill, A.; Sharland, G.; Allan, L. Assessment of changes in blood flow through the lungs and foramen ovale in the normal human fetus with gestational age: A prospective Doppler echocardiographic study. Heart 1994, 71, 232–237. [Google Scholar] [CrossRef]
- Capper, W.; Myers, L. Gestational age dependency of umbilical flow waveforms: An investigation using a lumped parameter model. In Proceedings of the 22nd Annual International Conference of the IEEE Engineering in Medicine and Biology Society (Cat. No.00CH37143), Chicago, IL, USA, 23–28 July 2000; Volume 2, pp. 1130–1133. [Google Scholar]
- Kenny, J.F.; Plappert, T.; Doubilet, P.; Saltzman, D.H.; Cartier, M.; Zollars, L.; Leatherman, G.; St John Sutton, M. Changes in intracardiac blood flow velocities and right and left ventricular stroke volumes with gestational age in the normal human fetus: A prospective Doppler echocardiographic study. Circulation 1986, 74, 1208–1216. [Google Scholar] [CrossRef]
- Ferrazzi, E.; Gementi, P.; Bellotti, M.; Rodolfi, M.; Della Peruta, S.; Barbera, A.; Pardi, G. Doppler velocimetry: Critical analysis of umbilical, cerebral and aortic reference values. Eur. J. Obstet. Gynecol. Reprod. Biol. 1991, 38, 189–196. [Google Scholar] [CrossRef]
- Hecher, K.; Campbell, S.; Snijders, R.; Nicolaides, K. Reference ranges for fetal venous and atrioventricular blood flow parameters. Ultrasound Obstet. Gynecol. Off. J. Int. Soc. Ultrasound Obstet. Gynecol. 1994, 4, 381–390. [Google Scholar] [CrossRef]
- Pennati, G.; Corno, C.; Costantino, M.L.; Bellotti, M. Umbilical flow distribution to the liver and the ductus venosus in human fetuses during gestation: An anatomy-based mathematical modeling. Med. Eng. Phys. 2003, 25, 229–238. [Google Scholar] [CrossRef]
- van den Wijngaard, J.P.H.M.; Westerhof, B.E.; Faber, D.J.; Ramsay, M.M.; Westerhof, N.; van Gemert, M.J.C. Abnormal arterial flows by a distributed model of the fetal circulation. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2006, 291, R1222–R1233. [Google Scholar] [CrossRef]
- Bellotti, M.; Pennati, G.; Pardi, G.; Fumero, R. Dilatation of the ductus venosus in human fetuses: Ultrasonographic evidence and mathematical modeling. Am. J. Physiol.-Heart Circ. Physiol. 1998, 275, H1759–H1767. [Google Scholar] [CrossRef] [PubMed]
- Grigioni, M.; Carotti, A.; Daniele, C.; D’Avenio, G.; Morbiducci, U.; Di Benedetto, G.; Albanese, S.; Di Donato, R.; Barbaro, V. A mathematical model of the fetal cardiovascular system based on genetic algorithms as identification technique. Int. J. Artif. Organs 2001, 24, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Struijk, P.C.; Mathews, V.J.; Loupas, T.; Stewart, P.A.; Clark, E.B.; Steegers, E.A.P.; Wladimiroff, J.W. Blood pressure estimation in the human fetal descending aorta. Ultrasound Obstet. Gynecol. 2008, 32, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Luria, O.; Bar, J.; Shalev, J.; Kovo, M.; Golan, A.; Barnea, O. Inverse solution of the fetal-circulation model based on ultrasound Doppler measurements. Cardiovasc. Eng. Technol. 2014, 5, 202–216. [Google Scholar] [CrossRef]
- Garcia-Cañadilla, P.; Crispi, F.; Cruz-Lemini, M.; Triunfo, S.; Nadal, A.; Valenzuela-Alcaraz, B.; Rudenick, P.A.; Gratacos, E.; Bijnens, B.H. Patient-specific estimates of vascular and placental properties in growth-restricted fetuses based on a model of the fetal circulation. Placenta 2015, 36, 981–989. [Google Scholar] [CrossRef]
- Kulkarni, A.; Garcia-Cañadilla, P.; Khan, A.; Lorenzo, J.M.; Beckerman, K.; Valenzuela-Alcaraz, B.; Cruz-Lemini, M.; Gomez, O.; Gratacos, E.; Crispi, F.; et al. Remodeling of the cardiovascular circulation in fetuses of mothers with diabetes: A fetal computational model analysis. Placenta 2018, 63, 1–6. [Google Scholar] [CrossRef]
- Hooper, S.; Pas, A.; Lang, J.; Van Vonderen, J.; Roehr, C.; Kluckow, M.; Gill, A.; Wallace, E.; Polglase, G. Cardiovascular transition at birth: A physiological sequence. Pediatr. Res. 2015, 77, 608–614. [Google Scholar] [CrossRef]
- Sá-Couto, C.D.; Andriessen, P.; Van Meurs, W.L.; Ayres-de Campos, D.; Sá-Couto, P.M. A model for educational simulation of hemodynamic transitions at birth. Pediatr. Res. 2010, 67, 158–165. [Google Scholar] [CrossRef]
- Yigit, B.; Tutsak, E.; Yıldırım, C.; Hutchon, D.; Pekkan, K. Transitional fetal hemodynamics and gas exchange in premature postpartum adaptation: Immediate vs. delayed cord clamping. Matern. Heal. Neonatol. Perinatol. 2019, 5, 5. [Google Scholar] [CrossRef]
- Rasanen, J.; Wood, D.C.; Weiner, S.; Ludomirski, A.; Huhta, J.C. Role of the pulmonary circulation in the distribution of human fetal cardiac output during the second half of pregnancy. Circulation 1996, 94, 1068–1073. [Google Scholar] [CrossRef] [PubMed]
- Wiputra, H.; Lai, C.Q.; Lim, G.L.; Heng, J.J.W.; Guo, L.; Soomar, S.M.; Leo, H.L.; Biwas, A.; Mattar, C.N.Z.; Yap, C.H. Fluid mechanics of human fetal right ventricles from image-based computational fluid dynamics using 4D clinical ultrasound scans. Am. J. Physiol.-Heart Circ. Physiol. 2016, 311, H1498–H1508. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.Q.; Lim, G.L.; Jamil, M.; Mattar, C.N.Z.; Biswas, A.; Yap, C.H. Fluid mechanics of blood flow in human fetal left ventricles based on patient-specific 4D ultrasound scans. Biomech. Model. Mechanobiol. 2016, 15, 1159–1172. [Google Scholar] [CrossRef] [PubMed]
- Groenenberg, I.; Stijnen, T.; Wladimiroff, J. Blood flow velocity waveforms in the fetal cardiac outflow tract as a measure of fetal well-being in intrauterine growth retardation. Pediatr. Res. 1990, 27, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Arvidsson, P.M.; Kovács, S.J.; Töger, J.; Borgquist, R.; Heiberg, E.; Carlsson, M.; Arheden, H. Vortex ring behavior provides the epigenetic blueprint for the human heart. Sci. Rep. 2016, 6, 22021. [Google Scholar] [CrossRef] [PubMed]
- Sahn, D.J.; Lange, L.W.; Allen, H.D.; Goldberg, S.J.; Anderson, C.; Giles, H.; Haber, K. Quantitative real-time cross-sectional echocardiography in the developing normal humam fetus and newborn. Circulation 1980, 62, 588–597. [Google Scholar] [CrossRef]
- Salman, H.E.; Kamal, R.Y.; Yalcin, H.C. Numerical Investigation of the Fetal Left Heart Hemodynamics During Gestational Stages. Front. Physiol. 2021, 12, 731428. [Google Scholar] [CrossRef]
- Salman, H.E.; Kamal, R.Y.; Hijazi, Z.M.; Yalcin, H.C. Hemodynamic and Structural Comparison of Human Fetal Heart Development Between Normally Growing and Hypoplastic Left Heart Syndrome-Diagnosed Hearts. Front. Physiol. 2022, 13, 856879. [Google Scholar] [CrossRef]
- Wiputra, H.; Lim, G.L.; Chia, D.A.K.; Mattar, C.N.Z.; Biswas, A.; Yap, C.H. Methods for fluid dynamics simulations of human fetal cardiac chambers based on patient-specific 4D ultrasound scans. J. Biomech. Sci. Eng. 2016, 11, 15-00608. [Google Scholar] [CrossRef]
- Wiputra, H.; Lim, G.L.; Chua, K.C.; Nivetha, R.; Soomar, S.M.; Biwas, A.; Mattar, C.N.Z.; Leo, H.L.; Yap, C.H. Peristaltic-like motion of the human fetal right ventricle and its effects on fluid dynamics and energy dynamics. Ann. Biomed. Eng. 2017, 45, 2335–2347. [Google Scholar] [CrossRef]
- Vasudevan, V.; Wiputra, H.; Yap, C.H. Torsional motion of the left ventricle does not affect ventricular fluid dynamics of both foetal and adult hearts. J. Biomech. 2019, 96, 109357. [Google Scholar] [CrossRef] [PubMed]
- Zebhi, B.; Wiputra, H.; Howley, L.; Cuneo, B.; Park, D.; Hoffman, H.; Gilbert, L.; Yap, C.H.; Bark, D. Right ventricle in hypoplastic left heart syndrome exhibits altered hemodynamics in the human fetus. J. Biomech. 2020, 112, 110035. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhou, Y.; Wang, J.; Liu, X.; Ge, S.; He, Y. Modeling of coarctation of aorta in human fetuses using 3D/4D fetal echocardiography and computational fluid dynamics. Echocardiography 2017, 34, 1858–1866. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhao, H.; Zhao, Y.; Han, J.; Yang, X.; Throckmorton, A.; Wei, Z.; Ge, S.; He, Y. Retrograde flow in aortic isthmus in normal and fetal heart disease by principal component analysis and computational fluid dynamics. Echocardiography 2022, 39, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Dean, W.R., XVI. Note on the motion of fluid in a curved pipe. Lond. Edinb. Dublin Philos. Mag. J. Sci. 1927, 4, 208–223. [Google Scholar] [CrossRef]
- Germano, M. On the effect of torsion on a helical pipe flow. J. Fluid Mech. 1982, 125, 1–8. [Google Scholar] [CrossRef]
- Kaplan, A.D.; Jaffa, A.J.; Timor, I.E.; Elad, D. Hemodynamic analysis of arterial blood flow in the coiled umbilical cord. Reprod. Sci. 2010, 17, 258–268. [Google Scholar] [CrossRef]
- Shah, R.G.; Girardi, T.; Merz, G.; Necaise, P.; Salafia, C.M. Hemodynamic analysis of blood flow in umbilical artery using computational modeling. Placenta 2017, 57, 9–12. [Google Scholar] [CrossRef]
- Saw, S.N.; Poh, Y.W.; Chia, D.; Biswas, A.; Mattar, C.N.Z.; Yap, C.H. Characterization of the hemodynamic wall shear stresses in human umbilical vessels from normal and intrauterine growth restricted pregnancies. Biomech. Model. Mechanobiol. 2018, 17, 1107–1117. [Google Scholar] [CrossRef]
- Wen, J.; Tang, J.; Ran, S.; Ho, H. Computational modelling for the spiral flow in umbilical arteries with different systole/diastole flow velocity ratios. Med. Eng. Phys. 2020, 84, 96–102. [Google Scholar] [CrossRef]
- Saw, S.N.; Dawn, C.; Biswas, A.; Mattar, C.N.Z.; Yap, C.H. Characterization of the in vivo wall shear stress environment of human fetus umbilical arteries and veins. Biomech. Model. Mechanobiol. 2017, 16, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Wilke, D.; Denier, J.; Khong, T.; Mattner, T. Pressure and flow in the umbilical cord. J. Biomech. 2018, 79, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Tejada-Martínez, A.E.; Borberg, C.J.; Venugopal, R.; Carballo, C.; Moreno, W.A.; Quintero, R.A. Computational fluid dynamic analysis of flow velocity waveform notching in umbilical arteries. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2011, 300, R76–R84. [Google Scholar] [CrossRef] [PubMed]
- Kasiteropoulou, D.; Topalidou, A.; Downe, S. A computational fluid dynamics modelling of maternal-fetal heat exchange and blood flow in the umbilical cord. PLoS ONE 2020, 15, e0231997. [Google Scholar] [CrossRef]
- Chato, J.C. Heat transfer to blood vessels. J. Biomed. Eng. 1980, 102, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Kolios, M.C.; Sherar, M.; Hunt, J. Large blood vessel cooling in heated tissues: A numerical study. Phys. Med. Biol. 1995, 40, 477. [Google Scholar] [CrossRef]
- Amare, R.; Hodneland, E.; Roberts, J.A.; Bahadori, A.A.; Eckels, S. Modeling a 3-D multiscale blood-flow and heat-transfer framework for realistic vascular systems. Sci. Rep. 2022, 12, 14610. [Google Scholar] [CrossRef]
- Liu, X.; Chen, X.; Zhang, Y.; Xie, J.; Jia, X.; Deng, T.; Zheng, Y.; Davood, T.; Majid, Z. The thermal behavior of blood flow in the arteries with various radii and various stenosis angles using non-Newtonian Sisko model. Alex. Eng. J. 2022, 61, 7195–7201. [Google Scholar] [CrossRef]
- Jiji, L.M. Heat Transfer in Living Tissue. In Heat Conduction: Third Edition; Springer: Berlin/Heidelberg, Germany, 2009; pp. 302–346. [Google Scholar]
- Pennati, G.; Redaelli, A.; Bellotti, M.; Ferrazzi, E. Computational analysis of the ductus venosus fluid dynamics based on Doppler measurements. Ultrasound Med. Biol. 1996, 22, 1017–1029. [Google Scholar] [CrossRef]
- Leinan, P.R.; Degroote, J.; Kiserud, T.; Skallerud, B.; Vierendeels, J.; Hellevik, L.R. Velocity profiles in the human ductus venosus: A numerical fluid structure interaction study. Biomech. Model. Mechanobiol. 2013, 12, 1019–1035. [Google Scholar] [CrossRef]
- Acharya, G.; Kiserud, T. Pulsations of the ductus venosus blood velocity and diameter are more pronounced at the outlet than at the inlet. Eur. J. Obstet. Gynecol. Reprod. Biol. 1999, 84, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Pennati, G.; Bellotti, M.; Ferrazzi, E.; Rigano, S.; Garberi, A. Hemodynamic changes across the human ductus venosus: A comparison between clinical findings and mathematical calculations. Ultrasound Obstet. Gynecol. 1997, 9, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Leinan, P.R.; Kiserud, T.; Hellevik, L.R. Human ductus venosus velocity profiles in the first trimester. Cardiovasc. Eng. Technol. 2013, 4, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Pennati, G.; Bellotti, M.; Ferrazzi, E.; Bozzo, M.; Pardi, G.; Fumero, R. Blood Flow Through the Ductus Venosus in Human Fetus: Calculation Using Doppler Velocimetry and Computational Findings. Ultrasound Med. Biol. 1998, 24, 477–487. [Google Scholar] [CrossRef]
- Haugen, G.; Kiserud, T.; Godfrey, K.; Crozier, S.; Hanson, M. Portal and umbilical venous blood supply to the liver in the human fetus near term. Ultrasound Obstet. Gynecol. 2004, 24, 599–605. [Google Scholar] [CrossRef]
- Poelmann, R.E.; Gittenberger-de Groot, A.C. Hemodynamics in Cardiac Development. J. Cardiovasc. Dev. Dis. 2018, 5, 54. [Google Scholar] [CrossRef]
- Kung, E.; Kahn, A.M.; Burns, J.C.; Marsden, A. In vitro validation of patient-specific hemodynamic simulations in coronary aneurysms caused by Kawasaki disease. Cardiovasc. Eng. Technol. 2014, 5, 189–201. [Google Scholar] [CrossRef]
- Roldán-Alzate, A.; García-Rodríguez, S.; Anagnostopoulos, P.V.; Srinivasan, S.; Wieben, O.; François, C.J. Hemodynamic study of TCPC using in vivo and in vitro 4D Flow MRI and numerical simulation. J. Biomech. 2015, 48, 1325–1330. [Google Scholar] [CrossRef]
- Sincomb, S.; Coenen, W.; Sánchez, A.L.; Lasheras, J.C. A model for the oscillatory flow in the cerebral aqueduct. J. Fluid Mech. 2020, 899, R1. [Google Scholar] [CrossRef]
- Kung, E.O.; Les, A.S.; Figueroa, C.A.; Medina, F.; Arcaute, K.; Wicker, R.B.; McConnell, M.V.; Taylor, C.A. In vitro validation of finite element analysis of blood flow in deformable models. Ann. Biomed. Eng. 2011, 39, 1947–1960. [Google Scholar] [CrossRef]
- Marquis, A.D.; Arnold, A.; Dean-Bernhoft, C.; Carlson, B.E.; Olufsen, M.S. Practical identifiability and uncertainty quantification of a pulsatile cardiovascular model. Math. Biosci. 2018, 304, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Bjørdalsbakke, N.L.; Sturdy, J.T.; Hose, D.R.; Hellevik, L.R. Parameter estimation for closed-loop lumped parameter models of the systemic circulation using synthetic data. Math. Biosci. 2022, 343, 108731. [Google Scholar] [CrossRef] [PubMed]
- Schrauben, E.M.; Darby, J.R.; Saini, B.S.; Holman, S.L.; Lock, M.C.; Perumal, S.R.; Seed, M.; Morrison, J.L.; Macgowan, C.K. Technique for comprehensive fetal hepatic blood flow assessment in sheep using 4D flow MRI. J. Physiol. 2020, 598, 3555–3567. [Google Scholar] [CrossRef] [PubMed]








| Mean Pressure (mmHg) | Flow Rate (%CCO) a | |
|---|---|---|
| Source | Yigit et al., 2015 [78] | Abduljalil et al., 2021 [73] |
| Methodology | Simulated using LPN | Compiled from ultrasound and MRI data |
| Gestational Age b | 40 weeks | weeks |
| Sample Size | - | Varies |
| Brain | 32 | |
| Lungs | 13 | 22.0 |
| Upper Limbs | 19 | - |
| Liver | 6 | |
| Intestine | 12 | - |
| Kidney | 34 | |
| Placenta | 26 |
| Oxygen Saturation (%) | Flow Rate (%CCO) | |
|---|---|---|
| Source | Saini et al., 2020 [89] | Prsa et al., 2014 [88] |
| Methodology | T2-MRI Oximetry | PC-MRI |
| Gestational Age | weeks | weeks |
| Sample Size | 40 fetuses | 30 fetuses |
| Ascending Aorta | ||
| Main Pulmonary Artery | ||
| Superior Vena Cava | - | |
| Ductus Arteriosus | - | |
| Descending Aorta | ||
| Umbilical Vein | ||
| Foramen Ovale | - |
| Circuit Element | Symbol | Hemodynamic Interpretation | Used for |
|---|---|---|---|
| Linear Resistor |  | Resistance to viscous flow | Vessels, peripheral vascular beds, heart chambers. |
| Nonlinear Resistor |  | Energy dissipation due to irregular local hemodynamics | Heart valves, vessels with abrupt changes in diameter (e.g., stenosis), etc. |
| Capacitor |  | Changes in flow due to vessel expansion and contraction | Compliant vessels and peripheral vascular beds. |
| Variable Capacitor |  | Time-varying changes in myocardial compliance | Contracting heart chambers. |
| Inductor |  | Inertia of flowing blood | Large arteries, heart chambers. |
| Diode |  | Unidirectional flow | Heart valves. |
| Current Source |  | Prescribed flow rate waveform | Inlets with available Doppler measurements. |
| Mean Pressure (mmHg) | Max Pressure (mmHg) | Min Pressure (mmHg) | Flow Rate (%CCO) | |
|---|---|---|---|---|
| Left Atrium | 3.0 | 4.0 | 1.5 | 57 |
| Left Ventricle | 25.0 | 55.0 | 1.9 | 57 |
| Right Atrium | 3.5 | 4.9 | 1.9 | 43 |
| Right Ventricle | 26.0 | 57.0 | 2.0 | 43 |
| Foramen Ovale | - | - | - | 21 |
| Ascending Aorta | 45.0 | 53.0 | 36.0 | 43 |
| Carotid Arteries | 42.0 | 48.0 | 35.0 | 21 |
| Descending Aorta | 42.0 | 48.0 | 36.0 | 47 |
| Main Pulmonary Artery | 46.0 | 54.0 | 37.0 | 57 |
| Ductus Arteriosus | - | - | - | 35 |
| Umbilical Arteries (Fetal End) | 41.0 | 47.0 | 35.0 | 22 |
| Umbilical Vein (Placental End) | 8.0 | - | - | 22 |
| Ductus Venosus | - | - | - | 8 |
| Thoracic Inferior Vena Cava | 4.9 | 5.5 | 4.4 | 47 |
| Superior Vena Cava | 5.0 | 5.5 | 4.8 | 31 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, D.; Lindsey, S.E. Recasting Current Knowledge of Human Fetal Circulation: The Importance of Computational Models. J. Cardiovasc. Dev. Dis. 2023, 10, 240. https://doi.org/10.3390/jcdd10060240
Zhang D, Lindsey SE. Recasting Current Knowledge of Human Fetal Circulation: The Importance of Computational Models. Journal of Cardiovascular Development and Disease. 2023; 10(6):240. https://doi.org/10.3390/jcdd10060240
Chicago/Turabian StyleZhang, Daibo, and Stephanie E. Lindsey. 2023. "Recasting Current Knowledge of Human Fetal Circulation: The Importance of Computational Models" Journal of Cardiovascular Development and Disease 10, no. 6: 240. https://doi.org/10.3390/jcdd10060240