

Effect of Procyanidin on Canine Sperm Quality during Chilled Storage
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Sources
2.2. Animals and Semen Collection
2.3. Sperm Processing
2.4. Sperm Motility and Kinetic Parameters Analysis
2.5. Sperm Plasma Membrane Integrity
2.6. Sperm Acrosome Integrity
2.7. Sperm Mitochondrial Membrane Potential
2.8. Measurement of Sperm T-AOC
2.9. Gene Expression Analysis
2.10. Statistical Analysis
3. Results
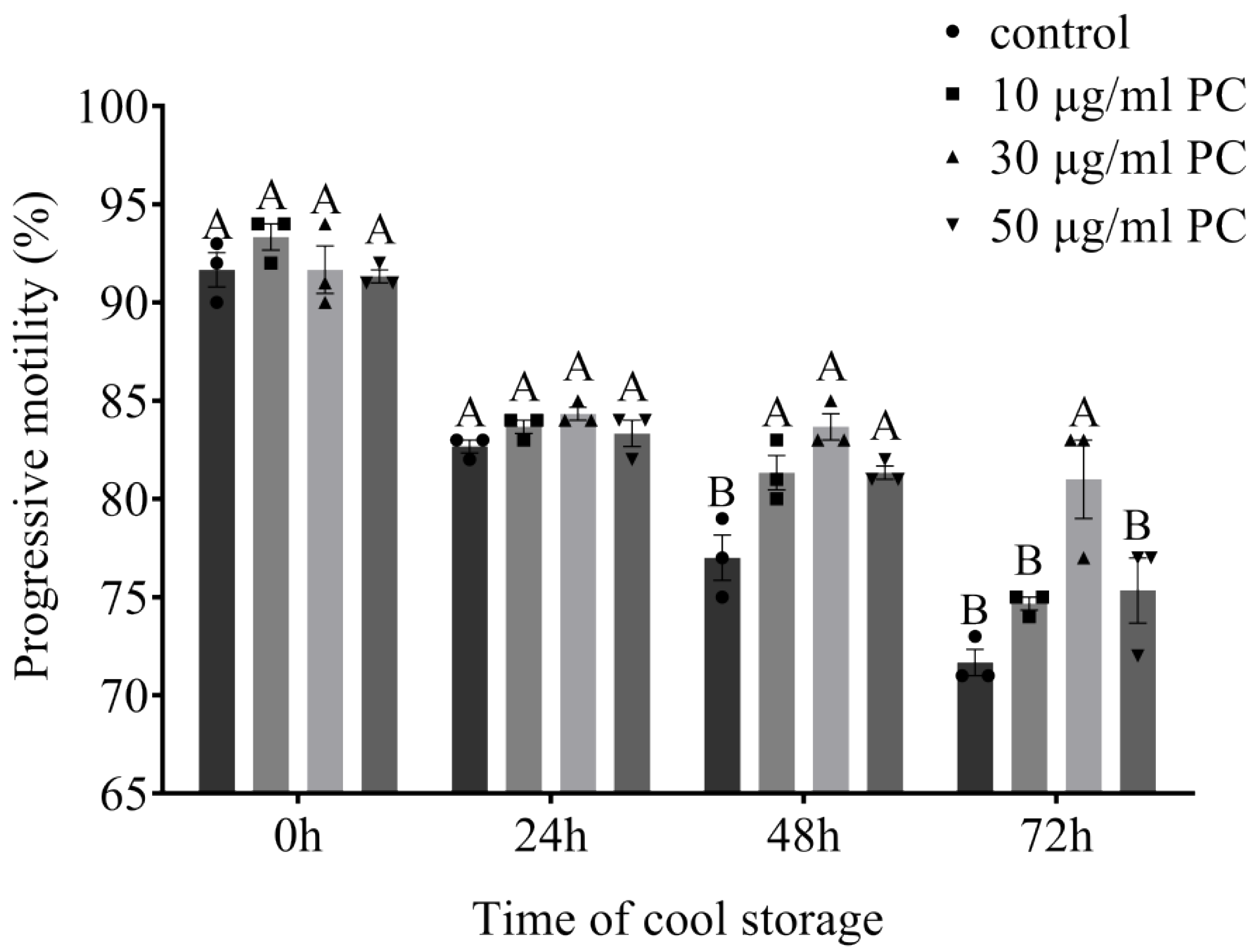
3.1. Effects of Procyanidin on Sperm Motility and Kinetic Parameters
3.2. Effects of Procyanidin on Sperm Plasma Membrane Integrity
3.3. Effects of Procyanidin on Sperm Acrosome Integrity
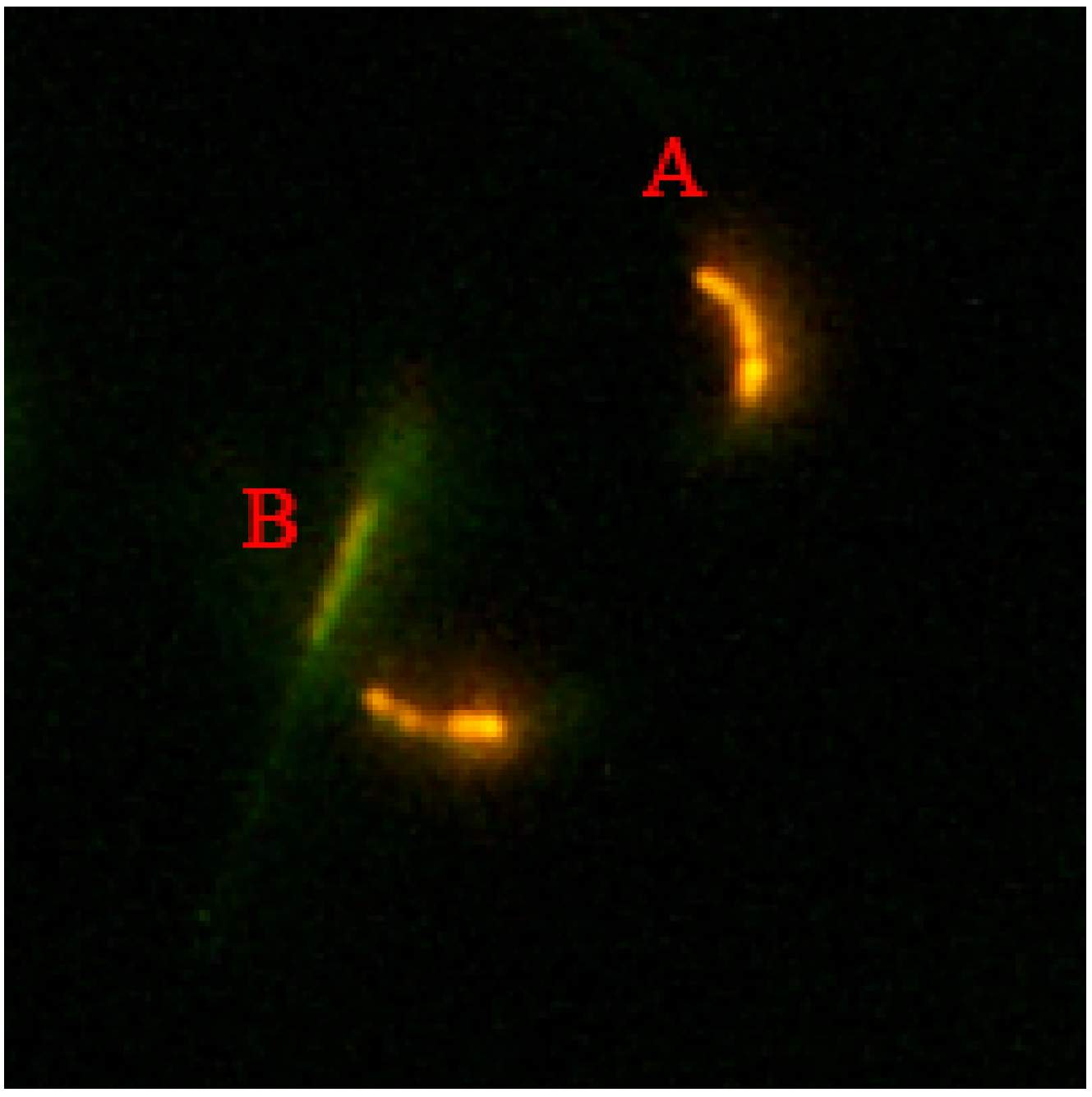
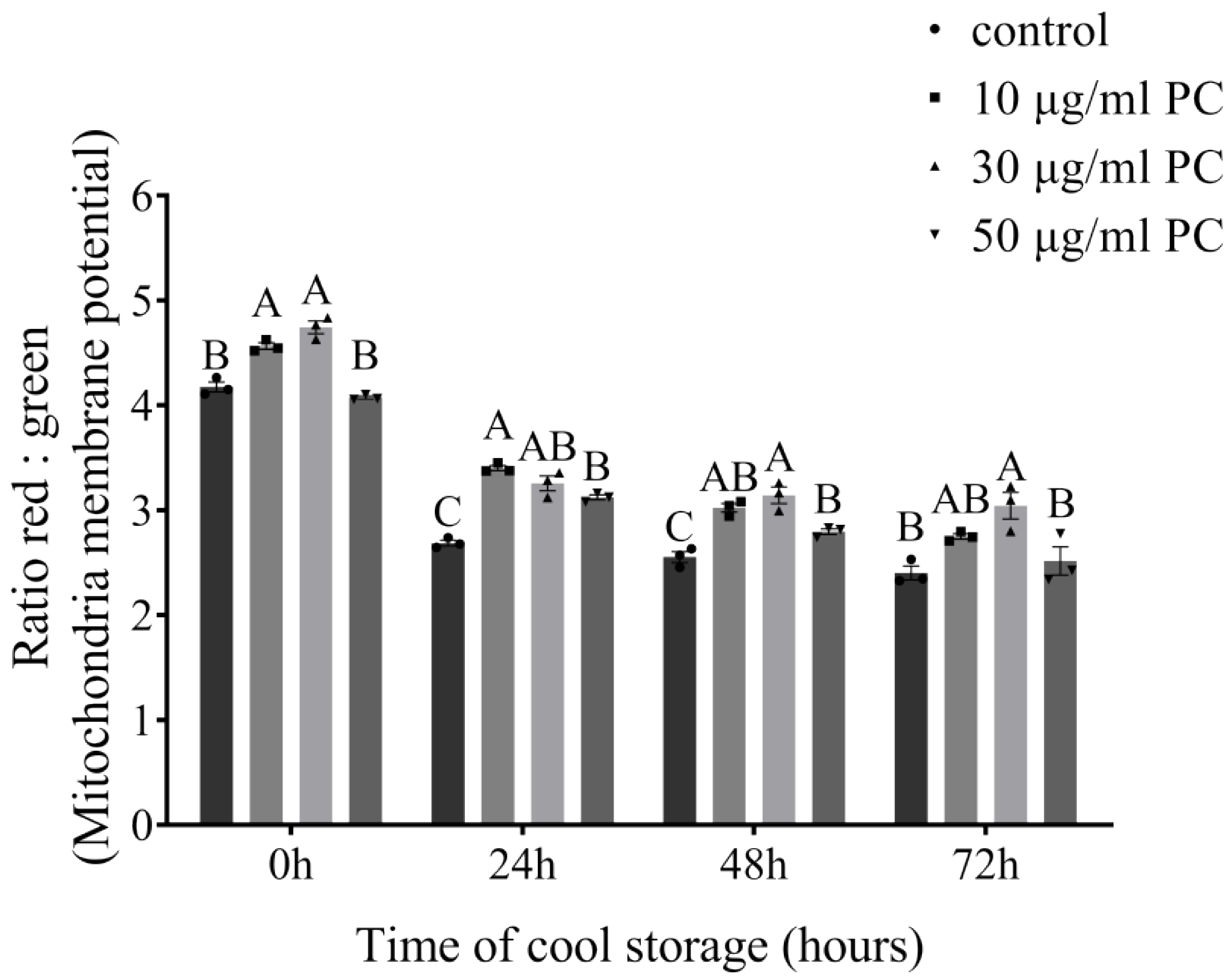
3.4. Effects of Procyanidin on Sperm Mitochondrial Membrane Potential
3.5. Effect of Procyandin on Sperm T-AOC
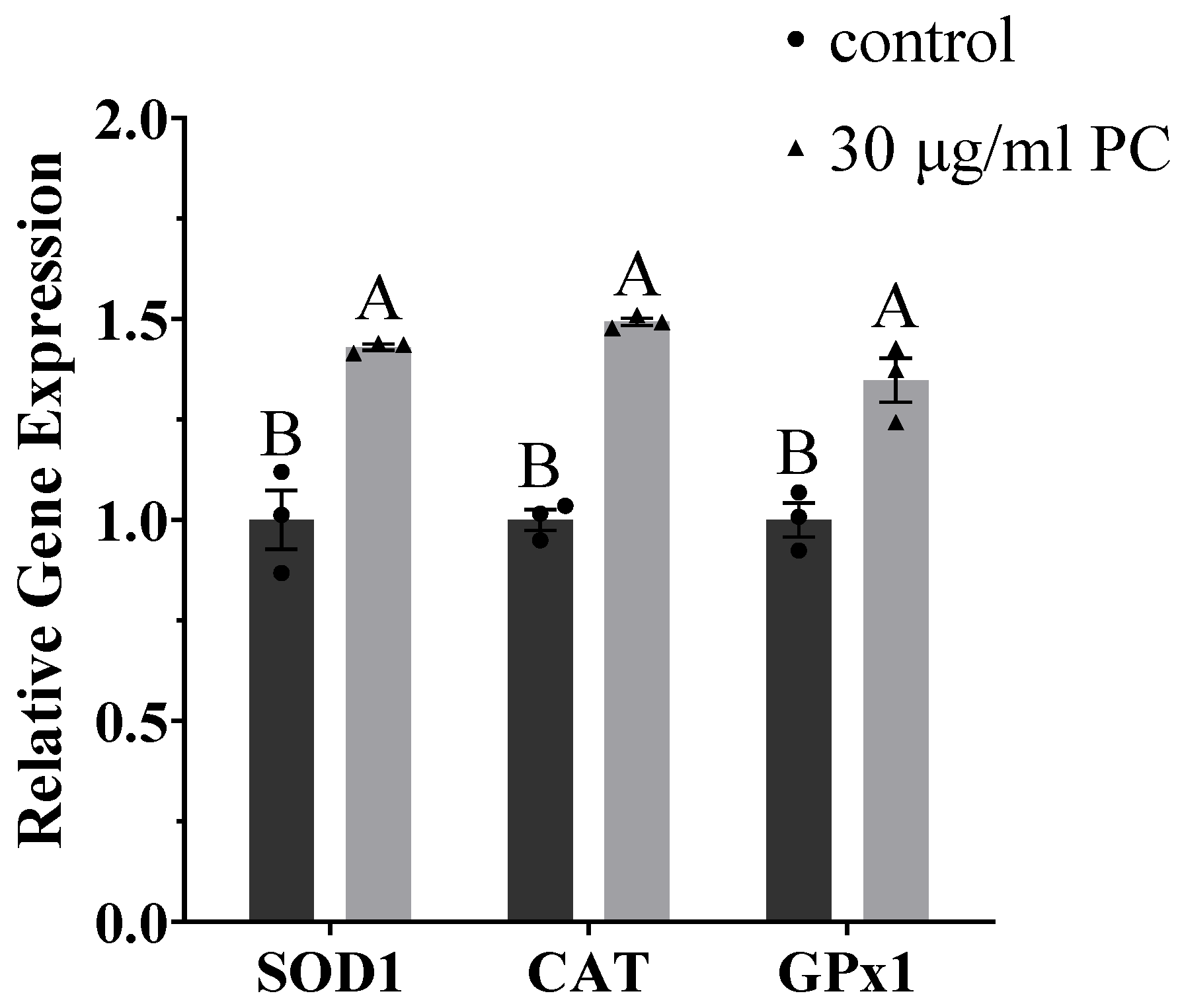
3.6. Effects of Procyandin on Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Suzuki, H.; Watanabe, H.; Abe, Y. Assisted reproductive techniques for canines: Preservation of genetic material in domestic dogs. J. Reprod. Dev. 2022, 68, 1–11. [Google Scholar] [CrossRef]
- Luno, V.; Gonzalez, N.; Martinez, F.; Gonzalez, B.; Olaciregui, M.; Gil, L. Specific phosphodiesterase type-10 inhibitor, papaverine, added after the cooling period improves canine sperm quality. Anim. Biotechnol. 2020, 33, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Linde-Forsberg, C. Artificial insemination with fresh, chilled extended, and frozen-thawed semen in the dog. Semin. Vet. Med. Surg. Small Anim. 1995, 10, 48–58. [Google Scholar] [PubMed]
- Moustafa, M.H.; Sharma, R.K.; Thornton, J.; Mascha, E.; Abdel-Hafez, M.A.; Thomas, A.J.; Agarwal, A. Relationship between ROS production, apoptosis and DNA denaturation in spermatozoa from patients examined for infertility. Hum. Reprod. 2004, 19, 129–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovalski, N.N.; de Lamirande, E.; Gagnon, C. Reactive oxygen species generated by human neutrophils inhibit sperm motility: Protective effect of seminal plasma and scavengers. Fertil. Steril. 1992, 58, 809–816. [Google Scholar] [CrossRef]
- Michael, A.J.; Alexopoulos, C.; Pontiki, E.A.; Hadjipavlou-Litina, D.J.; Saratsis, P.; Ververidis, H.N.; Boscos, C.M. Effect of antioxidant supplementation in semen extenders on semen quality and reactive oxygen species of chilled canine spermatozoa. Anim. Reprod. Sci. 2009, 112, 119–135. [Google Scholar] [CrossRef]
- Masoudi, R.; Sharafi, M.; Pourazadi, L.; Dadashpour Davachi, N.; Asadzadeh, N.; Esmaeilkhanian, S.; Dirandeh, E. Supplementation of chilling storage medium with glutathione protects rooster sperm quality. Cryobiology 2020, 92, 260–262. [Google Scholar] [CrossRef]
- Munsi, M.N.; Bhuiyan, M.; Majumder, S.; Alam, M. Effects of Exogenous Glutathione on the Quality of Chilled Bull Semen. Reprod. Domest. Anim. 2007, 42, 358–362. [Google Scholar] [CrossRef]
- Sheikholeslami, S.A.; Soleimanzadeh, A.; Rakhshanpour, A.; Shirani, D. The evaluation of lycopene and cysteamine supplementation effects on sperm and oxidative stress parameters during chilled storage of canine semen. Reprod. Domest. Anim. 2020, 55, 1229–1239. [Google Scholar] [CrossRef]
- Llopiz, N.; Puiggros, F.; Cespedes, E.; Arola, L.; Ardevol, A.; Blade, C.; Salvado, M.J. Antigenotoxic effect of grape seed procyanidin extract in Fao cells submitted to oxidative stress. J. Agric. Food Chem. 2004, 52, 1083–1087. [Google Scholar] [CrossRef]
- Bagchi, D.; Garg, A.; Krohn, R.L.; Bagchi, M.; Tran, M.X.; Stohs, S.J. Oxygen free radical scavenging abilities of vitamins C and E, and a grape seed proanthocyanidin extract in vitro. Res. Commun. Mol. Pathol. Pharmacol. 1997, 95, 179–189. [Google Scholar] [PubMed]
- Martinez-Micaelo, N.; Gonzalez-Abuin, N.; Ardevol, A.; Pinent, M.; Blay, M.T. Procyanidins and inflammation: Molecular targets and health implications. Biofactors 2012, 38, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Ribas-Latre, A.; Baselga-Escudero, L.; Casanova, E.; Arola-Arnal, A.; Salvado, M.J.; Blade, C.; Arola, L. Dietary proanthocyanidins modulate BMAL1 acetylation, Nampt expression and NAD levels in rat liver. Sci. Rep. 2015, 5, 10954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, L.; Deng, Y.; Zhang, Y.; Li, C.; Zhang, R.; Sun, Y.; Zhang, K.; Li, J.; Yao, S. Protective effects of grape seed procyanidin extract against nickel sulfate-induced apoptosis and oxidative stress in rat testes. Toxicol. Mech. Methods 2011, 21, 487–494. [Google Scholar] [CrossRef]
- Decorde, K.; Teissedre, P.L.; Sutra, T.; Ventura, E.; Cristol, J.P.; Rouanet, J.M. Chardonnay grape seed procyanidin extract supplementation prevents high-fat diet-induced obesity in hamsters by improving adipokine imbalance and oxidative stress markers. Mol. Nutr. Food Res. 2009, 53, 659–666. [Google Scholar] [CrossRef]
- Guo, L.; Wang, L.H.; Sun, B.; Yang, J.Y.; Zhao, Y.Q.; Dong, Y.X.; Spranger, M.I.; Wu, C.F. Direct in vivo evidence of protective effects of grape seed procyanidin fractions and other antioxidants against ethanol-induced oxidative DNA damage in mouse brain cells. J. Agric. Food Chem. 2007, 55, 5881–5891. [Google Scholar] [CrossRef]
- Awaga, H.A.; Lymperi, S.; Bosdou, J.K.; Makedos, A.; Mitsoli, A.; Bazioti, M.G.; Savvaidou, D.; Goulis, D.G.; Chatzimeletiou, K.; Salem, M.N.; et al. Addition of procyanidine to semen preserves progressive sperm motility up to three hours of incubation. Reprod. Biol. 2019, 19, 255–260. [Google Scholar] [CrossRef]
- Li, Q.; Shaoyong, W.; Li, Y.; Chen, M.; Hu, Y.; Liu, B.; Yang, G.; Hu, J. Effects of oligomeric proanthocyanidins on quality of boar semen during liquid preservation at 17 degrees C. Anim. Reprod. Sci. 2018, 198, 47–56. [Google Scholar] [CrossRef]
- Wen, F.; Li, Y.; Feng, T.; Du, Y.; Ren, F.; Zhang, L.; Han, N.; Ma, S.; Li, F.; Wang, P.; et al. Grape Seed Procyanidin Extract (GSPE) Improves Goat Sperm Quality When Preserved at 4 °C. Animals 2019, 9, 810. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Dai, J.; Zhang, S.; Sun, L.; Liu, Y.; Zhang, D. Effect of Thawing Rates and Antioxidants on Semen Cryopreservation in Hu Sheep. Biopreserv. Biobank. 2021, 19, 204–209. [Google Scholar] [CrossRef]
- Wang, H.; Lu, P.; Li, Z.; Yuan, C.; Liu, H.; Zhao, J.; Lu, W.; Wang, J. Oligomeric Proanthocyanidins and Bamboo Leaf Flavonoids Improve the Quality of Bull Semen Cryopreservation. Molecules 2022, 27, 1144. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, M.; Urbano, M.; Ortiz, I.; Demyda-Peyras, S.; Murabito, M.R.; Gálvez, M.J.; Dorado, J. DNA integrity of canine spermatozoa during chill storage assessed by the sperm chromatin dispersion test using bright-field or fluorescence microscopy. Theriogenology 2015, 84, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Drevet, J.R. The Importance of Oxidative Stress in Determining the Functionality of Mammalian Spermatozoa: A Two-Edged Sword. Antioxidants 2020, 9, 111. [Google Scholar] [CrossRef] [Green Version]
- Dutta, S.; Majzoub, A.; Agarwal, A. Oxidative stress and sperm function: A systematic review on evaluation and management. Arab. J. Urol. 2019, 17, 87–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betarelli, R.P.; Rocco, M.; Yeste, M.; Fernández-Novell, J.M.; Placci, A.; Azevedo, P.B.; Castillo-Martín, M.; Estrada, E.; Peña, A.; Zangeronimo, M.G.; et al. The achievement of boar sperm in vitro capacitation is related to an increase of disrupted disulphide bonds and intracellular reactive oxygen species levels. Andrology 2018, 6, 781–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khosrowbeygi, A.; Zarghami, N. Fatty acid composition of human spermatozoa and seminal plasma levels of oxidative stress biomarkers in subfertile males. Prostag. Leukot. Essent. Fatty Acids 2007, 77, 117–121. [Google Scholar] [CrossRef]
- Bromfield, E.G.; Aitken, R.J.; Anderson, A.L.; McLaughlin, E.A.; Nixon, B. The impact of oxidative stress on chaperone-mediated human sperm–egg interaction. Hum. Reprod. 2015, 30, 2597–2613. [Google Scholar] [CrossRef] [Green Version]
- Baker, M.A.; Weinberg, A.; Hetherington, L.; Villaverde, A.I.; Velkov, T.; Baell, J.; Gordon, C.P. Defining the mechanisms by which the reactive oxygen species by-product, 4-hydroxynonenal, affects human sperm cell function. Biol. Reprod. 2015, 92, 108. [Google Scholar] [CrossRef]
- Gharagozloo, P.; Aitken, R.J. The role of sperm oxidative stress in male infertility and the significance of oral antioxidant therapy. Hum. Reprod. 2011, 26, 1628–1640. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Kawai, T.; Umehara, T.; Hoque, S.; Zeng, W.; Shimada, M. Negative effects of ROS generated during linear sperm motility on gene expression and ATP generation in boar sperm mitochondria. Free Radic. Biol. Med. 2019, 141, 159–171. [Google Scholar] [CrossRef]
- de Lamirande, E.; Gagnon, C. Impact of reactive oxygen species on spermatozoa: A balancing act between beneficial and detrimental effects. Hum. Reprod. 1995, 10 (Suppl. S1), 15–21. [Google Scholar] [CrossRef] [PubMed]
- Divar, M.R.; Azari, M.; Mogheiseh, A.; Ghahramani, S. Supplementation of melatonin to cooling and freezing extenders improves canine spermatozoa quality measures. BMC Vet. Res. 2022, 18, 86. [Google Scholar] [CrossRef]
- Yang, L.; Xian, D.; Xiong, X.; Lai, R.; Song, J.; Zhong, J. Proanthocyanidins against Oxidative Stress: From Molecular Mechanisms to Clinical Applications. Biomed. Res. Int. 2018, 2018, 8584136. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.Q.; Wang, X.W.; Chen, J.F.; Ren, Q.L.; Wang, J.; Gao, B.W.; Shi, Z.H.; Zhang, Z.J.; Bai, X.X.; Xing, B.S. Grape Seed Procyanidin B2 Protects Porcine Ovarian Granulosa Cells against Oxidative Stress-Induced Apoptosis by Upregulating let-7a Expression. Oxid. Med. Cell. Longev. 2019, 2019, 1076512. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Gao, B.; Wang, J.; Ren, Q.; Chen, J.; Ma, Q.; Zhang, Z.; Xing, B. Critical Role of FoxO1 in Granulosa Cell Apoptosis Caused by Oxidative Stress and Protective Effects of Grape Seed Procyanidin B2. Oxid. Med. Cell. Longev. 2016, 2016, 6147345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Li, Q.; Wu, W.; Li, Y.; Hou, D.X.; Xu, H.; Zheng, B.; Zeng, S.; Shan, Y.; Lu, X.; et al. Lotus seed skin proanthocyanidin extract exhibits potent antioxidant property via activation of the Nrf2-ARE pathway. Acta Biochem. Biophys. Sin. 2019, 51, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Rajput, S.A.; Sun, L.; Zhang, N.Y.; Khalil, M.M.; Ling, Z.; Chong, L.; Wang, S.; Rajput, I.R.; Bloch, D.M.; Khan, F.A.; et al. Grape Seed Proanthocyanidin Extract Alleviates AflatoxinB(1)-Induced Immunotoxicity and Oxidative Stress via Modulation of NF-kappaB and Nrf2 Signaling Pathways in Broilers. Toxins 2019, 11, 23. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Cheng, Z.; Wang, K.; Zhu, X.; Ali, Y.; Shu, W.; Bao, X.; Zhu, L.; Fan, X.; Murray, M.; et al. Procyanidin B2 and rutin in Ginkgo biloba extracts protect human retinal pigment epithelial (RPE) cells from oxidative stress by modulating Nrf2 and Erk1/2 signalling. Exp. Eye Res. 2021, 207, 108586. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′–3′) | Product Size (bp) | NCBI Accession No. |
|---|---|---|---|
| ACTB | F: TGGCCGAGGACTTTGATTGT | 171 | NM_001195845.3 |
| R: CTCTTTCCCTCCCCTGTGTG | |||
| SOD1 | F: GTTGGAGACCTGGGCAATGT | 211 | NM_001003035.1 |
| R: TCCCAATGACACCACAAGCC | |||
| CAT | F: CCTATCCTGACACTCACCGC | 219 | NM_001002984.1 |
| R: GCACATCTGGCGAACATTGG | |||
| GPx1 | F: CGGGCATCAGGAAAACGCTA | 209 | NM_001115119.1 |
| R: TGATGAACTTGGGGTCGGTC |
| Motility Parameters | Time (h) | Groups | |||
|---|---|---|---|---|---|
| Control | 10 μg/mL PC | 30 μg/mL PC | 50 μg/mL PC | ||
| TM (%) | 0 | 96.60 ± 0.24 Aa | 97.80 ± 0.58 Aa | 97.00 ± 0.63 Aa | 97.20 ± 0.37 Aa |
| 24 | 93.20 ± 0.37 Bb | 95.80 ± 0.58 Aa | 96.20 ± 0.58 Aab | 96.00 ± 0.45 Aab | |
| 48 | 92.20 ± 0.58 Bb | 94.00 ± 0.55 Aab | 95.00 ± 0.32 Ab | 94.60 ± 0.51 Ab | |
| 72 | 89.00 ± 1.00 Bc | 91.00 ± 2.83 ABb | 94.80 ± 0.86 Ab | 91.80 ± 1.07 ABc | |
| VCL (um/s) | 0 | 105.15 ± 2.78 Aa | 94.14 ± 4.20 Ba | 103.23 ± 2.05 ABa | 102.68 ± 3.97 ABa |
| 24 | 86.29 ± 1.07 Ab | 77.24 ± 1.09 Cb | 82.83 ± 1.96 ABb | 81.94 ± 1.19 Bb | |
| 48 | 74.78 ± 0.72 Ac | 77.38 ± 2.77 Ab | 77.96 ± 0.51 Ab | 75.83 ± 0.36 Abc | |
| 72 | 74.34 ± 2.85 Ac | 77.84 ± 2.86 Ab | 77.08 ± 2.89 Ab | 73.39 ± 3.48 Ac | |
| VSL (um/s) | 0 | 68.62 ± 1.03 Aa | 60.61 ± 3.06 Ba | 73.07 ± 2.07 Aa | 68.23 ± 3.00 Aa |
| 24 | 64.78 ± 0.88 Ab | 56.62 ± 1.24 Bab | 57.76 ± 1.88 Bb | 58.61 ± 1.42 Bb | |
| 48 | 52.70 ± 0.22 Ac | 53.34 ± 0.71 Ab | 54.44 ± 0.69 Ab | 53.51 ± 0.71 Abc | |
| 72 | 51.53 ± 1.06 ABc | 55.11 ± 2.11 Aab | 46.91 ± 1.52 Bc | 49.27 ± 3.13 ABc | |
| VAP (um/s) | 0 | 75.11 ± 1.17 ABa | 67.96 ± 3.52 Ba | 75.80 ± 1.21 Aa | 75.42 ± 3.25 ABa |
| 24 | 65.55 ± 1.10 Ab | 61.04 ± 1.03 BCb | 59.90 ± 1.73 Cb | 64.13 ± 1.27 ABb | |
| 48 | 56.73 ± 0.45 BCc | 60.13 ± 0.88 Ab | 56.16 ± 0.49 Cbc | 58.22 ± 0.52 Bbc | |
| 72 | 57.80 ± 1.62 ABc | 60.24 ± 2.32 Ab | 52.74 ± 1.62 Bc | 53.99 ± 3.01 ABc | |
| LIN (%) | 0 | 63.32 ± 0.71 ABa | 60.31 ± 1.89 Bb | 66.16 ± 0.46 Aa | 60.84 ± 0.54 Ba |
| 24 | 64.83 ± 0.63 ABa | 65.13 ± 0.75 Aa | 62.77 ± 0.39 BCb | 61.91 ± 1.04 Ca | |
| 48 | 59.93 ± 1.14 Ab | 59.68 ± 1.13 Ab | 60.61 ± 0.33 Ac | 59.65 ± 1.18 Aa | |
| 72 | 48.74 ± 0.73 Bc | 57.01 ± 1.59 Ab | 56.42 ± 0.92 Ad | 54.53 ± 0.88 Ab | |
| WOB (%) | 0 | 70.69 ± 0.47 Bb | 69.24 ± 1.09 Bb | 73.48 ± 0.38 Aa | 69.46 ± 0.77 Ba |
| 24 | 74.41 ± 0.34 Aa | 72.79 ± 0.65 Ba | 71.52 ± 0.13 BCb | 70.71 ± 0.55 Ca | |
| 48 | 70.94 ± 0.33 Ab | 71.31 ± 0.53 Aab | 70.26 ± 0.23 ABc | 69.32 ± 0.69 Ba | |
| 72 | 61.67 ± 1.24 Bc | 65.84 ± 1.62 Ac | 65.94 ± 0.48 Ad | 62.44 ± 0.62 Bb | |
| STR (%) | 0 | 88.39 ± 0.76 Aa | 85.89 ± 1.25 ABa | 88.28 ± 0.83 ABa | 85.48 ± 0.90 Ba |
| 24 | 86.72 ± 0.52 Aa | 84.88 ± 1.17 ABa | 86.39 ± 0.49 ABb | 83.87 ± 1.01 Bab | |
| 48 | 82.34 ± 0.68 Cb | 84.31 ± 0.38 ABa | 84.48 ± 0.09 Ac | 82.73 ± 0.75 BCb | |
| 72 | 80.80 ± 0.36 Bb | 84.15 ± 0.49 Aa | 83.47 ± 0.58 Ac | 81.79 ± 0.65 Bb | |
| ALH (μm) | 0 | 4.96 ± 0.23 Aa | 5.18 ± 0.18 Aa | 4.92 ± 0.06 Aa | 5.07 ± 0.12 Aa |
| 24 | 4.38 ± 0.09 Aab | 4.58 ± 0.13 Aab | 4.73 ± 0.14 Aab | 4.54 ± 0.16 Ab | |
| 48 | 3.84 ± 0.08 Bbc | 4.26 ± 0.13 ABbc | 4.44 ± 0.09 Ab | 3.94 ± 0.06 Bc | |
| 72 | 3.27 ± 0.11 Cc | 3.70 ± 0.08 ABc | 3.94 ± 0.06 Ac | 3.41 ± 0.11 BCd | |
| Analysis | Time (h) | Groups | |||
|---|---|---|---|---|---|
| Control | 10 μg/mL PC | 30 μg/mL PC | 50 μg/mL PC | ||
| Membrane integrity (%) | 0 h | 72.13 ± 0.04 Ba | 73.71 ± 0.49 ABa | 74.89 ± 0.70 Aa | 72.87 ± 0.72 Ba |
| 24 h | 65.45 ± 0.35 Cb | 67.76 ± 0.48 Bb | 70.94 ± 0.41 Ab | 69.87 ± 0.57 Ab | |
| 48 h | 59.34 ± 0.32 Dc | 62.73 ± 0.23 Cc | 65.49 ± 0.29 Ac | 64.39 ± 0.29 Bc | |
| 72 h | 54.12 ± 0.40 Cd | 56.83 ± 0.19 Bd | 59.83 ± 0.27 Ad | 57.63 ± 0.31 Bd | |
| Acrosome integrity (%) | 0 h | 74.97 ± 0.59 Aa | 73.69 ± 0.15 Aa | 74.10 ± 0.46 Aa | 73.48 ± 0.76 Aa |
| 24 h | 64.97 ± 0.56 Bb | 67.84 ± 0.25 Ab | 67.37 ± 0.31 Ab | 65.01 ± 0.24 Bb | |
| 48 h | 60.12 ± 0.36 Cc | 64.34 ± 0.32 Ac | 64.61 ± 0.41 Ac | 62.77 ± 0.20 Bc | |
| 72 h | 49.26 ± 0.38 Dd | 52.64 ± 0.23 Bd | 54.69 ± 0.23 Ad | 50.56 ± 0.56 Cd | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Zhao, Z.; Wang, R.; Ma, Y.; Bu, Y.; Hu, M.; Zhang, S. Effect of Procyanidin on Canine Sperm Quality during Chilled Storage. Vet. Sci. 2022, 9, 588. https://doi.org/10.3390/vetsci9110588
Huang X, Zhao Z, Wang R, Ma Y, Bu Y, Hu M, Zhang S. Effect of Procyanidin on Canine Sperm Quality during Chilled Storage. Veterinary Sciences. 2022; 9(11):588. https://doi.org/10.3390/vetsci9110588
Chicago/Turabian StyleHuang, Xiaogang, Zhihong Zhao, Ronggen Wang, Ying Ma, Yonghui Bu, Minhua Hu, and Shouquan Zhang. 2022. "Effect of Procyanidin on Canine Sperm Quality during Chilled Storage" Veterinary Sciences 9, no. 11: 588. https://doi.org/10.3390/vetsci9110588