
Honey Bee Pathogen Prevalence and Interactions within the Marmara Region of Turkey


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Sample Processing
2.3. Pathogen Screening
2.3.1. Stonebrood (SB) Screening
2.3.2. Chalkbrood (CB) Screening
2.3.3. American Foulbrood (AFB) Screening
2.3.4. European Foulbrood (EFB) Screening
2.3.5. Nosema Screening and Semi-Quantification of Nosema Infection
2.3.6. RNA Extraction, Quality Control (QC), and Sequencing for Viral Screening
2.3.7. Quality Control and Pre-Processing of RNA-seq Reads for Viral Identification and Quantification
2.4. Statistical Analysis and Visualization
3. Results
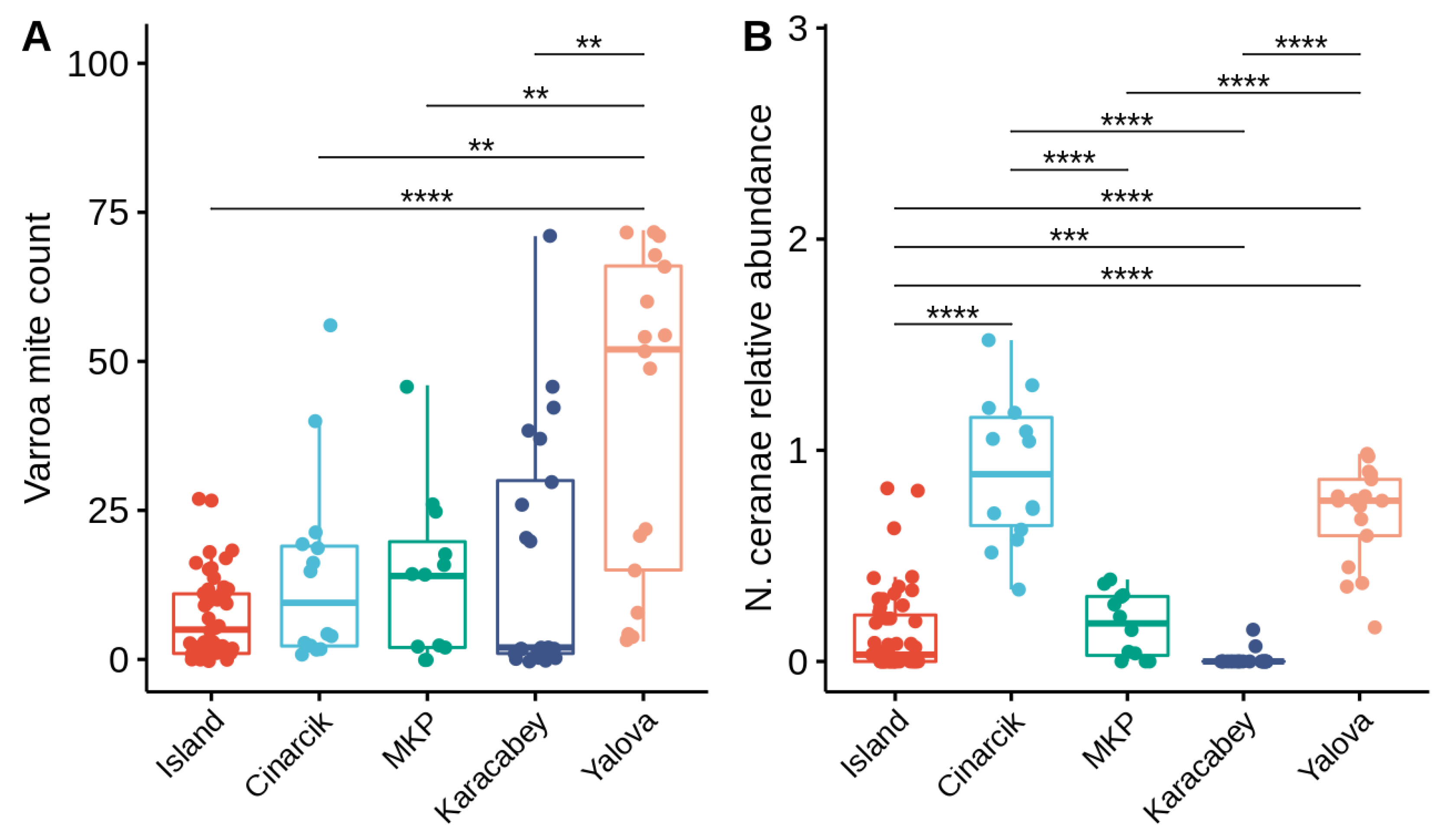
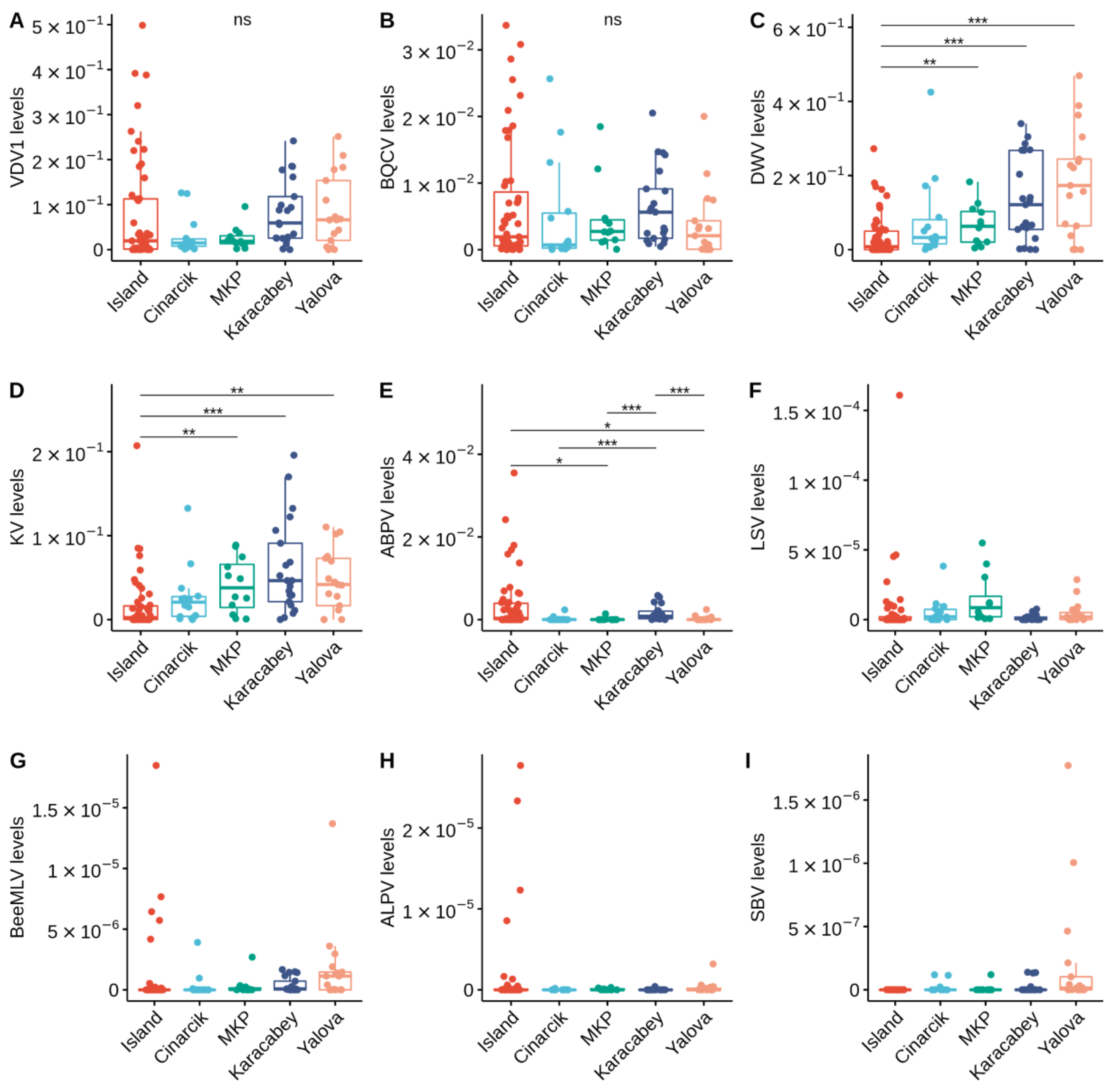
3.1. Prevalence of Pathogens
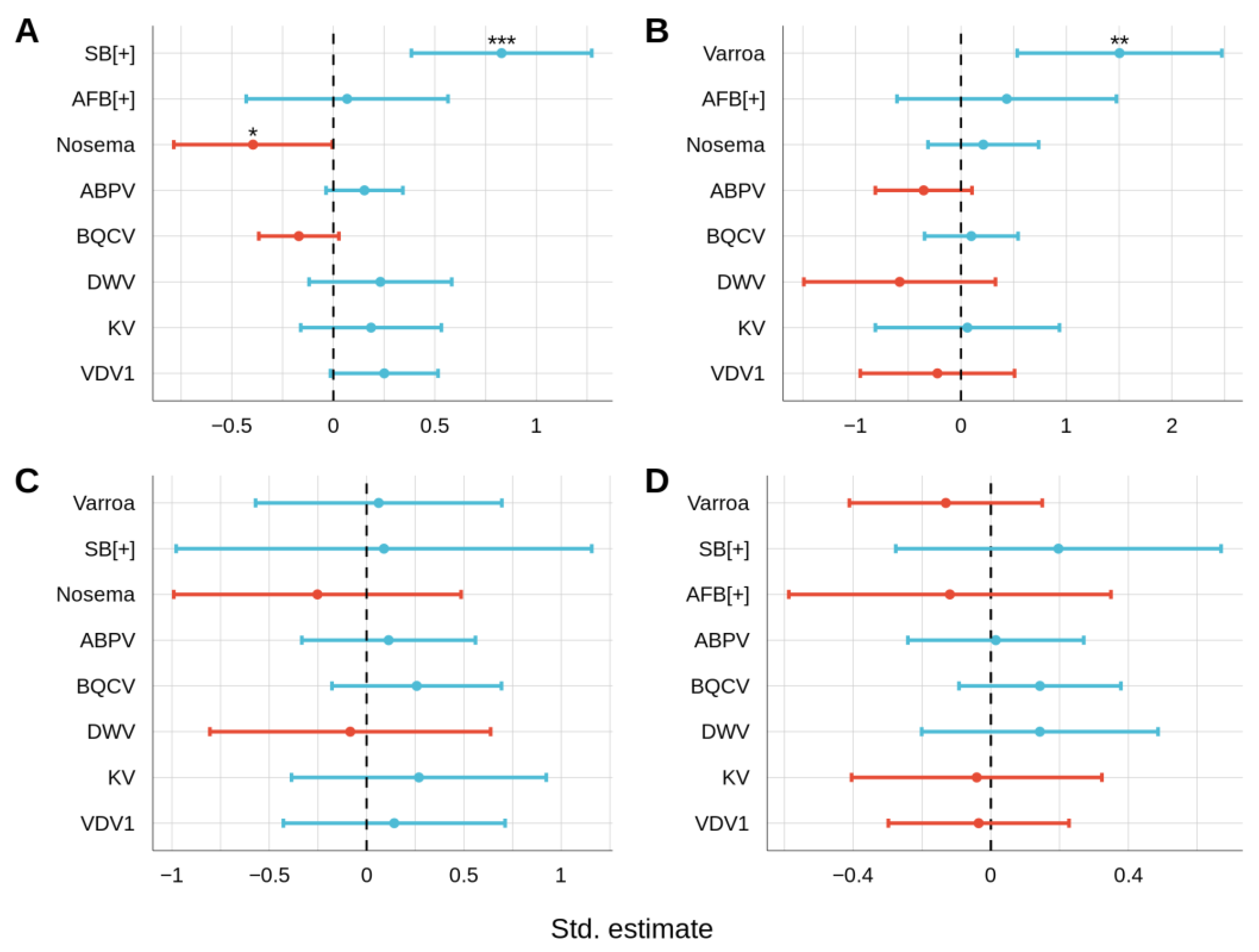
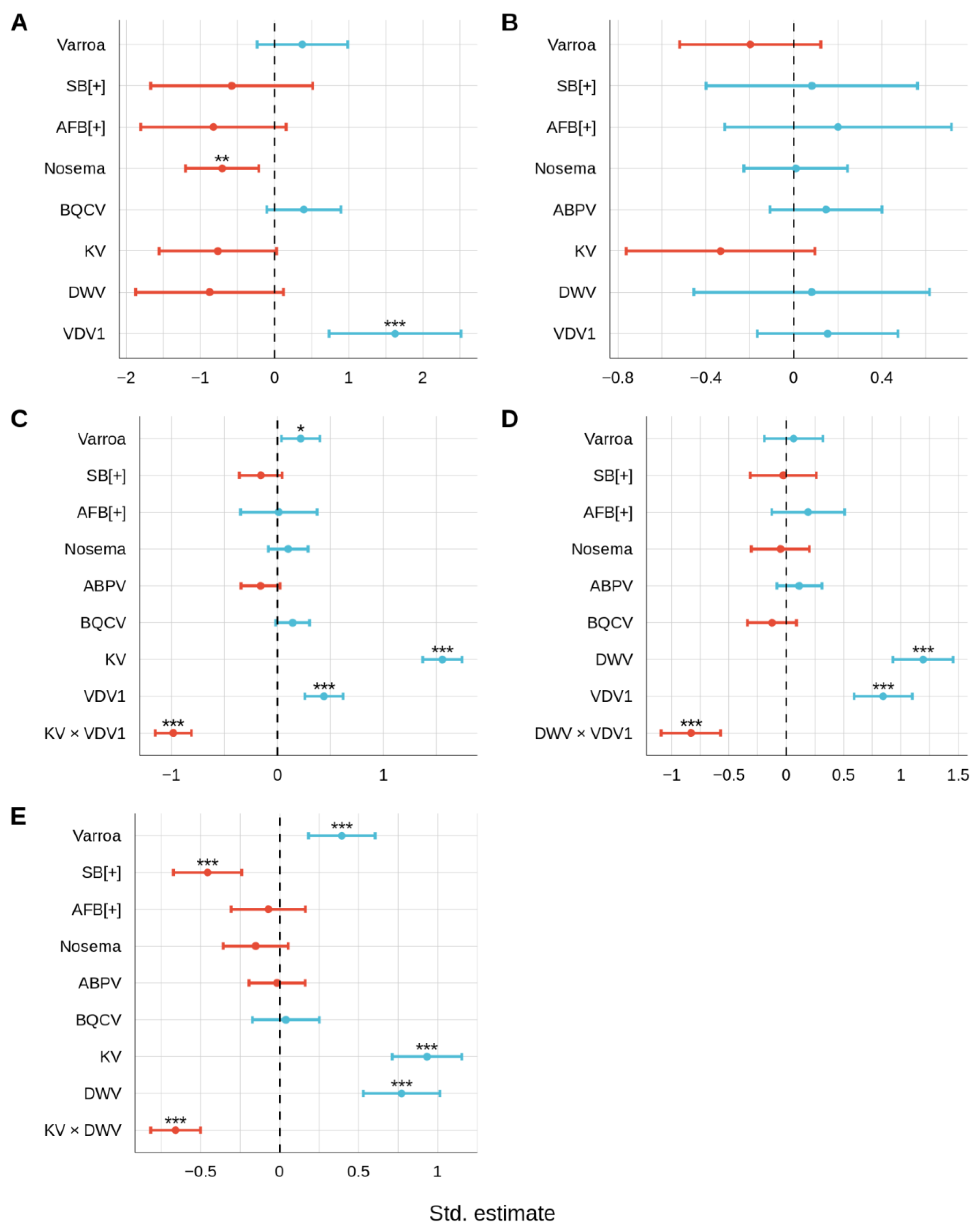
3.2. Relationships between Honey Bee Pathogens
3.2.1. Non-Viral Infections
3.2.2. Viral Infections
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of Pollinators in Changing Landscapes for World Crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potts, S.G.; Imperatriz-Fonseca, V.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; et al. Safeguarding Pollinators and Their Values to Human Well-Being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandemir, I.; Kence, M.; Sheppard, W.S.; Kence, A. Mitochondrial DNA Variation in Honey Bee (Apis mellifera L.) Populations from Turkey. J. Apic. Res. 2006, 45, 33–38. [Google Scholar] [CrossRef]
- Kandemir, I.; Kence, M.; Kence, A. Genetic and Morphometric Variation in Honeybee (Apis mellifera L.) Populations of Turkey. Apidologie 2000, 31, 343–356. [Google Scholar] [CrossRef] [Green Version]
- Turkish Statistical Institute Data Portal of Statistics, Agriculture. Available online: https://data.tuik.gov.tr/Kategori/GetKategori?p=Agriculture-111 (accessed on 25 August 2022).
- Özkırım, A. Beekeeping in Turkey: Bridging Asia and Europe. In Asian Beekeeping in the 21st Century; Chantawannakul, P., Williams, G., Neumann, P., Eds.; Springer: Singapore, 2018; pp. 41–69. [Google Scholar]
- Food and Agriculture Organization of the United Nations. FAOSTAT, Crops and Livestock Products. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 25 August 2022).
- Çakmak, S.S.; Çakmak, İ. Beekeeping and Recent Colony Losses in Turkey. Uludağ Arıcılık Derg. 2016, 16, 31–48. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee Declines Driven by Combined Stress from Parasites, Pesticides, and Lack of Flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global Pollinator Declines: Trends, Impacts and Drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Naug, D. Nutritional Stress Due to Habitat Loss May Explain Recent Honeybee Colony Collapses. Biol. Conserv. 2009, 142, 2369–2372. [Google Scholar] [CrossRef]
- Broadrup, R.L.; Mayack, C.; Schick, S.J.; Eppley, E.J.; White, H.K.; Macherone, A. Honey Bee (Apis mellifera) Exposomes and Dysregulated Metabolic Pathways Associated with Nosema Ceranae Infection. PLoS ONE 2019, 14, e0213249. [Google Scholar] [CrossRef] [Green Version]
- Mayack, C.; Boff, S. LD50 Values May Be Misleading Predictors of Neonicotinoid Toxicity across Different Bee Species. Uludağ Arıcılık Derg. 2019, 19, 19–33. [Google Scholar] [CrossRef]
- Mayack, C.; Macherone, A.; Zaki, A.G.; Filiztekin, E.; Özkazanç, B.; Koperly, Y.; Schick, S.J.; Eppley, E.J.; Deb, M.; Ambiel, N.; et al. Environmental Exposures Associated with Honey Bee Health. Chemosphere 2022, 286, 131948. [Google Scholar] [CrossRef] [PubMed]
- Siviter, H.; Bailes, E.J.; Martin, C.D.; Oliver, T.R.; Koricheva, J.; Leadbeater, E.; Brown, M.J.F. Agrochemicals Interact Synergistically to Increase Bee Mortality. Nature 2021, 596, 389–392. [Google Scholar] [CrossRef] [PubMed]
- Tozkar, C.Ö.; Kence, M.; Kence, A.; Huang, Q.; Evans, J.D. Metatranscriptomic Analyses of Honey Bee Colonies. Front. Genet. 2015, 6, 100. [Google Scholar] [CrossRef] [Green Version]
- Brettell, L.E.; Schroeder, D.C.; Martin, S.J. RNAseq of Deformed Wing Virus and Other Honey Bee-Associated Viruses in Eight Insect Taxa with or without Varroa Infestation. Viruses 2020, 12, 1229. [Google Scholar] [CrossRef]
- Evans, J.D.; Schwarz, R.S. Bees Brought to Their Knees: Microbes Affecting Honey Bee Health. Trends Microbiol. 2011, 19, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Çakmak, İ.; Çakmak, S.S.; Fuchs, S.; Yenïnar, H. Balarısı Kolonilerinde Varroa Bulaşıklık Seviyesinin Belirlenmesinde Pudra Şekeri ve Deterjan Yönteminin Karşılaştırılması. Uludağ Arıcılık Derg. 2011, 11, 63–68. [Google Scholar]
- Nasri, T.; Hedayati, M.T.; Abastabar, M.; Pasqualotto, A.C.; Armaki, M.T.; Hoseinnejad, A.; Nabili, M. PCR-RFLP on β-Tubulin Gene for Rapid Identification of the Most Clinically Important Species of Aspergillus. J. Microbiol. Methods 2015, 117, 144–147. [Google Scholar] [CrossRef]
- Jeon, J.H.; Moon, K.; Kim, Y.; Kim, Y.H. Reference Gene Selection for QRT-PCR Analysis of Season- and Tissue-Specific Gene Expression Profiles in the Honey Bee Apis mellifera. Sci. Rep. 2020, 10, 13935. [Google Scholar] [CrossRef]
- Thompson, G.J.; Yockey, H.; Lim, J.; Oldroyd, B.P. Experimental Manipulation of Ovary Activation and Gene Expression in Honey Bee (Apis mellifera) Queens and Workers: Testing Hypotheses of Reproductive Regulation. J. Exp. Zool. Part Ecol. Genet. Physiol. 2007, 307, 600–610. [Google Scholar] [CrossRef]
- Jensen, A.B.; Welker, D.L.; Kryger, P.; James, R.R. Polymorphic DNA Sequences of the Fungal Honey Bee Pathogen Ascosphaera Apis. FEMS Microbiol. Lett. 2012, 330, 17–22. [Google Scholar] [CrossRef]
- James, R.R.; Skinner, J.S. PCR Diagnostic Methods for Ascosphaera Infections in Bees. J. Invertebr. Pathol. 2005, 90, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Jensen, A.B.; Aronstein, K.; Flores, J.M.; Vojvodic, S.; Palacio, M.A.; Spivak, M. Standard Methods for Fungal Brood Disease Research. J. Apic. Res. 2013, 52, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govan, V.A.; Allsopp, M.H.; Davison, S. A PCR Detection Method for Rapid Identification of Paenibacillus Larvae. Appl. Environ. Microbiol. 1999, 65, 2243–2245. [Google Scholar] [CrossRef] [Green Version]
- Rossi, F.; Amadoro, C.; Ruberto, A.; Ricchiuti, L. Evaluation of Quantitative PCR (QPCR) Paenibacillus Larvae Targeted Assays and Definition of Optimal Conditions for Its Detection/Quantification in Honey and Hive Debris. Insects 2018, 9, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forsgren, E.; Budge, G.E.; Charrière, J.-D.; Hornitzky, M.A.Z. Standard Methods for European Foulbrood Research. J. Apic. Res. 2013, 52, 1–14. [Google Scholar] [CrossRef]
- Hamiduzzaman, M.M.; Guzman-Novoa, E.; Goodwin, P.H. A Multiplex PCR Assay to Diagnose and Quantify Nosema Infections in Honey Bees (Apis mellifera). J. Invertebr. Pathol. 2010, 105, 151–155. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Aranda, P.S.; LaJoie, D.M.; Jorcyk, C.L. Bleach Gel: A Simple Agarose Gel for Analyzing RNA Quality. Electrophoresis 2012, 33, 366–369. [Google Scholar] [CrossRef] [Green Version]
- Winnebeck, E.C.; Millar, C.D.; Warman, G.R. Why Does Insect RNA Look Degraded? J. Insect Sci. Online 2010, 10, 159. [Google Scholar] [CrossRef]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize Analysis Results for Multiple Tools and Samples in a Single Report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-Based Genome Alignment and Genotyping with HISAT2 and HISAT-Genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup the Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Hartig, F. DHARMa-Residual Diagnostics for HierARchical Models 2022. Available online: https://github.com/florianhartig/DHARMa (accessed on 4 July 2022).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. Artic. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. GlmmTMB Balances Speed and Flexibility Among Packages for Zero-Inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Venables, B.; Ripley, B. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Lüdecke, D. SjPlot: Data Visualization for Statistics in Social Science 2021. Available online: https://CRAN.R-project.org/package=sjPlot (accessed on 27 July 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Kassambara, A. ggpubr: ‘ggplot2’ Based Publication Ready Plots. 2020. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 22 September 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Xiao, N. ggsci: Scientific Journal and Sci-Fi Themed Color Palettes for ‘gplot2’. 2018. Available online: https://github.com/nanxstats/ggsci (accessed on 4 July 2022).
- Çakmak, İ.; Aydın, L.; Gulegen, E.; Wells, H. Varroa (Varroa Destructor) and Tracheal Mite (Acarapis Woodi) Incidence in the Republic of Turkey. J. Apic. Res. 2003, 42, 57–60. [Google Scholar] [CrossRef]
- Ütük, A.E.; Pïşkïn, F.Ç.; Denïz, A.; Balkaya, İ. Varroosis ve nosemosis üzerine retrospektif bir çalışma. Etlik Vet. Mikrobiyoloji Derg. 2011, 22, 11–15. [Google Scholar]
- Çakmak, İ.; Aydın, L.; Güleğen, A.E. Güney marmara bölgesinde balarısı zararlı ve hastalıkları. Uludag Bee J. 2003, 3, 33–35. [Google Scholar]
- Bayrakal, G.; Dümen, E.; Ekïcï, G.; Akkaya, H.; Sezgïn, F.H. Detection and Molecular Examination of Pathogens in Honey and Bees in the Northern Marmara Region, Turkey. Kafkas Üniversitesi Vet. Fakültesi Derg. 2020, 26, 313–319. [Google Scholar] [CrossRef]
- Muz, D.; Muz, M.N. Investigation of Some Pathogens “Colony Loss Syndrome” Resembled Losses Apiaries in Tekirdağ. Kocatepe Vet. J. 2017, 10, 21–28. [Google Scholar]
- Şahïnler, N.; Gül, A. Hatay Yöresinde Bulunan Arıcılık İşletmelerinde Arı Hastalıklarının Araştırılması. Uludağ Arıcılık Derg. 2005, 5, 27–31. [Google Scholar]
- Muz, M.N.; Yaman, M.; Karakavuk, M.; Solmaz, H. Kış Salkımı erken bozulan Arı Kolonilerinde Paraziter ve Bakteriyel Patojenler. Üzüncü Il Üniversitesi Vet. Fakültesi Derg. 2012, 23, 147–150. [Google Scholar]
- Yalçınkaya, A.; Keskin, N. The Investigation of Honey Bee Diseases after Colony Losses in Hatay and Adana Provinces of Turkey. Mellifera 2010, 10, 24–31. [Google Scholar]
- Şïmşek, H. Elazığ yöresi bal arılarında bazı parazit ve mantar hastalıklarının araştırılması. Ank. Üniversitesi Vet. Fakültesi Derg. 2005, 52, 123–126. [Google Scholar]
- Balkaya, İ. Erzurum Yöresi Arıcılarının Karşılaştıkları Bal Arısı Hastalıkları. Atatürk Üniversitesi Vet. Bilim. Derg. 2016, 11, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Aydın, A. Hakkari Yöresinde Varroasis’in Yaygınlığı. Üzüncü Il Üniversitesi Vet. Fakültesi Derg. 2012, 23, 129–130. [Google Scholar]
- Önk, K.; Kılıç, Y. Kars yöresindeki bal arılarında varroosis’in yaygınlığı. Uludağ Arıcılık Derg. 2014, 14, 69–73. [Google Scholar] [CrossRef]
- Mõtus, K.; Raie, A.; Orro, T.; Chauzat, M.-P.; Viltrop, A. Epidemiology, Risk Factors and Varroa Mite Control in the Estonian Honey Bee Population. J. Apic. Res. 2016, 55, 396–412. [Google Scholar] [CrossRef]
- Dahle, B. The Role of Varroa Destructor for Honey Bee Colony Losses in Norway. J. Apic. Res. 2010, 49, 124–125. [Google Scholar] [CrossRef]
- Anido, M.; Branchiccela, B.; Castelli, L.; Harriet, J.; Campá, J.; Zunino, P.; Antúnez, K. Prevalence and Distribution of Honey Bee Pests and Pathogens in Uruguay. J. Apic. Res. 2015, 54, 532–540. [Google Scholar] [CrossRef]
- Antúnez, K.; Invernizzi, C.; Mendoza, Y.; vanEngelsdorp, D.; Zunino, P. Honeybee Colony Losses in Uruguay during 2013–2014. Apidologie 2017, 48, 364–370. [Google Scholar] [CrossRef] [Green Version]
- Kulhanek, K.; Steinhauer, N.; Rennich, K.; Caron, D.M.; Sagili, R.R.; Pettis, J.S.; Ellis, J.D.; Wilson, M.E.; Wilkes, J.T.; Tarpy, D.R.; et al. A National Survey of Managed Honey Bee 2015–2016 Annual Colony Losses in the USA. J. Apic. Res. 2017, 56, 328–340. [Google Scholar] [CrossRef] [Green Version]
- Seitz, N.; Traynor, K.S.; Steinhauer, N.; Rennich, K.; Wilson, M.E.; Ellis, J.D.; Rose, R.; Tarpy, D.R.; Sagili, R.R.; Caron, D.M.; et al. A National Survey of Managed Honey Bee 2014–2015 Annual Colony Losses in the USA. J. Apic. Res. 2015, 54, 292–304. [Google Scholar] [CrossRef]
- Traynor, K.S.; Rennich, K.; Forsgren, E.; Rose, R.; Pettis, J.; Kunkel, G.; Madella, S.; Evans, J.; Lopez, D.; vanEngelsdorp, D. Multiyear Survey Targeting Disease Incidence in US Honey Bees. Apidologie 2016, 47, 325–347. [Google Scholar] [CrossRef] [Green Version]
- Fujiyuki, T.; Ohka, S.; Takeuchi, H.; Ono, M.; Nomoto, A.; Kubo, T. Prevalence and Phylogeny of Kakugo Virus, a Novel Insect Picorna-Like Virus That Infects the Honeybee (Apis mellifera L.), under Various Colony Conditions. J. Virol. 2006, 80, 11528–11538. [Google Scholar] [CrossRef] [Green Version]
- Çağirgan, A.A.; Yazıcı, Z. The Prevalence of Seven Crucial Honeybee Viruses Using Multiplex RT-PCR and Theirphylogenetic Analysis. Turk. J. Vet. Anim. Sci. 2021, 45, 44–55. [Google Scholar] [CrossRef]
- Kalayci, G.; Çağirgan, A.A.; Kaplan, M.; Pekmez, K.; Beyazit, A.; Ozkan, B.; Yesıloz, H.; Arslan, F. The Role of Viral and Parasitic Pathogens Affected by Colony Losses in Turkish Apiaries. Kafkas Üniversitesi Vet. Fakültesi Derg. 2020, 26, 671–677. [Google Scholar] [CrossRef]
- Gumusova, S.O.; Albayrak, H.; Kurt, M.; Yazici, Z. Prevalence of Three Honey Bee Viruses in Turkey. Vet. Arh. 2010, 80, 779–785. [Google Scholar]
- Rüstemoğlu, M.; Sïpahïoğlu, H.M. Occurrence and Molecular Characterization of Acute Bee Paralysis Virus (ABPV) in Honeybee (Apis mellifera) Colonies in Hakkari Province. Üzüncü Il Üniversitesi Tarım Bilim. Derg. 2016, 26, 174–182. [Google Scholar]
- Çağirgan, A.A.; Kaplan, M.; Pekmez, K.; Arslan, F. Türkiye’de Varroa Akarlarında Lake Sinai Virus (LSV)’ un İlk Tespiti. Atatürk Üniversitesi Vet. Bilim. Derg. 2022, 17, 16–19. [Google Scholar]
- Chen, Y.P.; Pettis, J.S.; Collins, A.; Feldlaufer, M.F. Prevalence and Transmission of Honeybee Viruses. Appl. Environ. Microbiol. 2006, 72, 606–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, L. Honey Bee Pathology. Annu. Rev. Entomol. 1968, 13, 191–212. [Google Scholar] [CrossRef]
- Sıralı, R.; Doğaroğlu, M. Survey Results on Honeybee Pests and Diseases in Thracian Region of Turkey. Uludag Bee J. 2005, 33, 56–62. [Google Scholar]
- Dümen, E.; Akkaya, H.; Öz, G.; Sezgin, F. Microbiological and Parasitological Quality of Honey Produced in İstanbul. Turk. J. Vet. Anim. Sci. 2013, 37, 602–607. [Google Scholar] [CrossRef]
- Foley, K.; Fazio, G.; Jensen, A.B.; Hughes, W.O.H. The Distribution of Aspergillus Spp. Opportunistic Parasites in Hives and Their Pathogenicity to Honey Bees. Vet. Microbiol. 2014, 169, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Evison, S.E. Chalkbrood: Epidemiological Perspectives from the Host–Parasite Relationship. Curr. Opin. Insect Sci. 2015, 10, 65–70. [Google Scholar] [CrossRef]
- Borum, A.E.; Ülgen, M. Güney Marmara Bölgesindeki Bal Arılarının Chalkbrood (Ascosphaera apis) İnfeksiyonunda Predispozisyon Faktörleri. Uludağ Arıcılık Derg. 2010, 10, 56–69. [Google Scholar]
- Maxfield-Taylor, S.A.; Mujic, A.B.; Rao, S. First Detection of the Larval Chalkbrood Disease Pathogen Ascosphaera Apis (Ascomycota: Eurotiomycetes: Ascosphaerales) in Adult Bumble Bees. PLoS ONE 2015, 10, e0124868. [Google Scholar] [CrossRef]
- Sevim, A.; Akpınar, R.; Karaoğlu, Ş.A.; Bozdeveci, A.; Sevim, E. Prevalence and Phylogenetic Analysis of Ascosphaera Apis (Maassen Ex Claussen) LS Olive & Spiltoir (1955) Isolates from Honeybee Colonies in Turkey. Biologia 2022, 1, 1–11. [Google Scholar] [CrossRef]
- Aronstein, K.A.; Murray, K.D. Chalkbrood Disease in Honey Bees. J. Invertebr. Pathol. 2010, 103, S20–S29. [Google Scholar] [CrossRef]
- Gillard, M.; Charriere, J.D.; Belloy, L. Distribution of Paenibacillus Larvae Spores inside Honey Bee Colonies and Its Relevance for Diagnosis. J. Invertebr. Pathol. 2008, 99, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Roetschi, A.; Berthoud, H.; Kuhn, R.; Imdorf, A. Infection Rate Based on Quantitative Real-Time PCR of Melissococcus Plutonius, the Causal Agent of European Foulbrood, in Honeybee Colonies before and after Apiary Sanitation. Apidologie 2008, 39, 362–371. [Google Scholar] [CrossRef]
- Genersch, E. American Foulbrood in Honeybees and Its Causative Agent, Paenibacillus Larvae. J. Invertebr. Pathol. 2010, 103, S10–S19. [Google Scholar] [CrossRef]
- Aydin, L.; Cakmak, I.; Gulegen, E.; Wells, H. Honey Bee Nosema Disease in the Republic of Turkey. J. Apic. Res. 2005, 44, 196–197. [Google Scholar] [CrossRef]
- Kartal, S.; Tunca, R.İ.; Özgül, O.; Karabağ, K.; Koç, H. Microscopic and Molecular Detection of Nosema sp. In the southwest aegean region. Uludağ Arıcılık Derg. 2021, 21, 8–20. [Google Scholar] [CrossRef]
- Yilmaz, F.; Öztürk, S.H.; Kuvanci, A.; Kayaboynu, Ü.; Karataş, Ü.; Kaya, S.; Derebaşi, E.; Buldağ, M. Doğu Karadeniz Bölgesinde Nosema apis ve Nosema ceranae’nın Epidemiyolojisi. Arıcılık Araşt. Derg. 2018, 10, 34–44. [Google Scholar]
- Oğuz, B.; Karapinar, Z.; Dïnçer, E.; Değer, M.S. Molecular Detection of Nosema Spp. and Black Queen-Cell Virus in Honeybees in Van Province, Turkey. Turk. J. Vet. Anim. Sci. 2017, 41, 221–227. [Google Scholar] [CrossRef]
- Matthijs, S.; De Waele, V.; Vandenberge, V.; Verhoeven, B.; Evers, J.; Brunain, M.; Saegerman, C.; De Winter, P.J.J.; Roels, S.; de Graaf, D.C.; et al. Nationwide Screening for Bee Viruses and Parasites in Belgian Honey Bees. Viruses 2020, 12, 890. [Google Scholar] [CrossRef]
- Mohammadian, B.; Bokaie, S.; Moharrami, M.; Nabian, S.; Forsi, M. Distribution of Nosema Spp. in Climatic Regions of Iran. Vet. Res. Forum 2018, 9, 259–263. [Google Scholar] [CrossRef]
- Shumkova, R.; Georgieva, A.; Radoslavov, G.; Sirakova, D.; Dzhebir, G.; Neov, B.; Bouga, M.; Hristov, P. The First Report of the Prevalence of Nosema Ceranae in Bulgaria. PeerJ 2018, 6, e4252. [Google Scholar] [CrossRef] [Green Version]
- Emsen, B.; De la Mora, A.; Lacey, B.; Eccles, L.; Kelly, P.G.; Medina-Flores, C.A.; Petukhova, T.; Morfin, N.; Guzman-Novoa, E. Seasonality of Nosema Ceranae Infections and Their Relationship with Honey Bee Populations, Food Stores, and Survivorship in a North American Region. Vet. Sci. 2020, 7, 131. [Google Scholar] [CrossRef] [PubMed]
- Emsen, B.; Guzman-Novoa, E.; Hamiduzzaman, M.M.; Eccles, L.; Lacey, B.; Ruiz-Pérez, R.A.; Nasr, M. Higher Prevalence and Levels of Nosema Ceranae than Nosema Apis Infections in Canadian Honey Bee Colonies. Parasitol. Res. 2016, 115, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Benoit, J.B.; Yoder, J.A.; Sammataro, D.; Zettler, L.W. Mycoflora and Fungal Vector Capacity of the Parasitic Mite Varroa Destructor (Mesostigmata: Varroidae) in Honey Bee (Hymenoptera: Apidae) Colonies. Int. J. Acarol. 2004, 30, 103–106. [Google Scholar] [CrossRef]
- Ball, B.V.; Allen, M.F. The Prevalence of Pathogens in Honey Bee (Apis mellifera) Colonies Infested with the Parasitic Mite Varroa Jacobsoni. Ann. Appl. Biol. 1988, 113, 237–244. [Google Scholar] [CrossRef]
- Highfield, A.C.; El Nagar, A.; Mackinder, L.C.M.; Noël, L.M.-L.J.; Hall, M.J.; Martin, S.J.; Schroeder, D.C. Deformed Wing Virus Implicated in Overwintering Honeybee Colony Losses. Appl. Environ. Microbiol. 2009, 75, 7212–7220. [Google Scholar] [CrossRef]
- Moore, J.; Jironkin, A.; Chandler, D.; Burroughs, N.; Evans, D.J.; Ryabov, E.V.Y. 2011 Recombinants between Deformed Wing Virus and Varroa Destructor Virus-1 May Prevail in Varroa Destructor-Infested Honeybee Colonies. J. Gen. Virol. 2011, 92, 156–161. [Google Scholar] [CrossRef]
- McMahon, D.P.; Natsopoulou, M.E.; Doublet, V.; Fürst, M.; Weging, S.; Brown, M.J.F.; Gogol-Döring, A.; Paxton, R.J. Elevated Virulence of an Emerging Viral Genotype as a Driver of Honeybee Loss. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160811. [Google Scholar] [CrossRef] [Green Version]
- Ryabov, E.V.; Wood, G.R.; Fannon, J.M.; Moore, J.D.; Bull, J.C.; Chandler, D.; Mead, A.; Burroughs, N.; Evans, D.J. A Virulent Strain of Deformed Wing Virus (DWV) of Honeybees (Apis mellifera) Prevails after Varroa Destructor-Mediated, or In Vitro, Transmission. PLOS Pathog. 2014, 10, e1004230. [Google Scholar] [CrossRef] [Green Version]
- Remnant, E.J.; Mather, N.; Gillard, T.L.; Yagound, B.; Beekman, M. Direct Transmission by Injection Affects Competition among RNA Viruses in Honeybees. Proc. R. Soc. B Biol. Sci. 2019, 286, 20182452. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Name of Location | GPS Coordinates | Sample Size |
|---|---|---|---|
| December 2020 | Marmara Island (Island) | 40.6227° N, 27.6175° E | 19 |
| July 2021 | Marmara Island (Island) | 40.6227° N, 27.6175° E | 32 |
| Karacabey | 40.2160° N, 28.3590° E | 21 | |
| Mustafakemalpasa (MKP) | 40.0394° N, 28.4052° E | 12 | |
| Cinarcik | 40.6452° N, 29.1192° E | 14 | |
| Yalova | 40.6549° N, 29.2842° E | 17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayack, C.; Hakanoğlu, H. Honey Bee Pathogen Prevalence and Interactions within the Marmara Region of Turkey. Vet. Sci. 2022, 9, 573. https://doi.org/10.3390/vetsci9100573
Mayack C, Hakanoğlu H. Honey Bee Pathogen Prevalence and Interactions within the Marmara Region of Turkey. Veterinary Sciences. 2022; 9(10):573. https://doi.org/10.3390/vetsci9100573
Chicago/Turabian StyleMayack, Christopher, and Haşim Hakanoğlu. 2022. "Honey Bee Pathogen Prevalence and Interactions within the Marmara Region of Turkey" Veterinary Sciences 9, no. 10: 573. https://doi.org/10.3390/vetsci9100573