
Molecular Profile of Canine Hemangiosarcoma and Potential Novel Therapeutic Targets
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
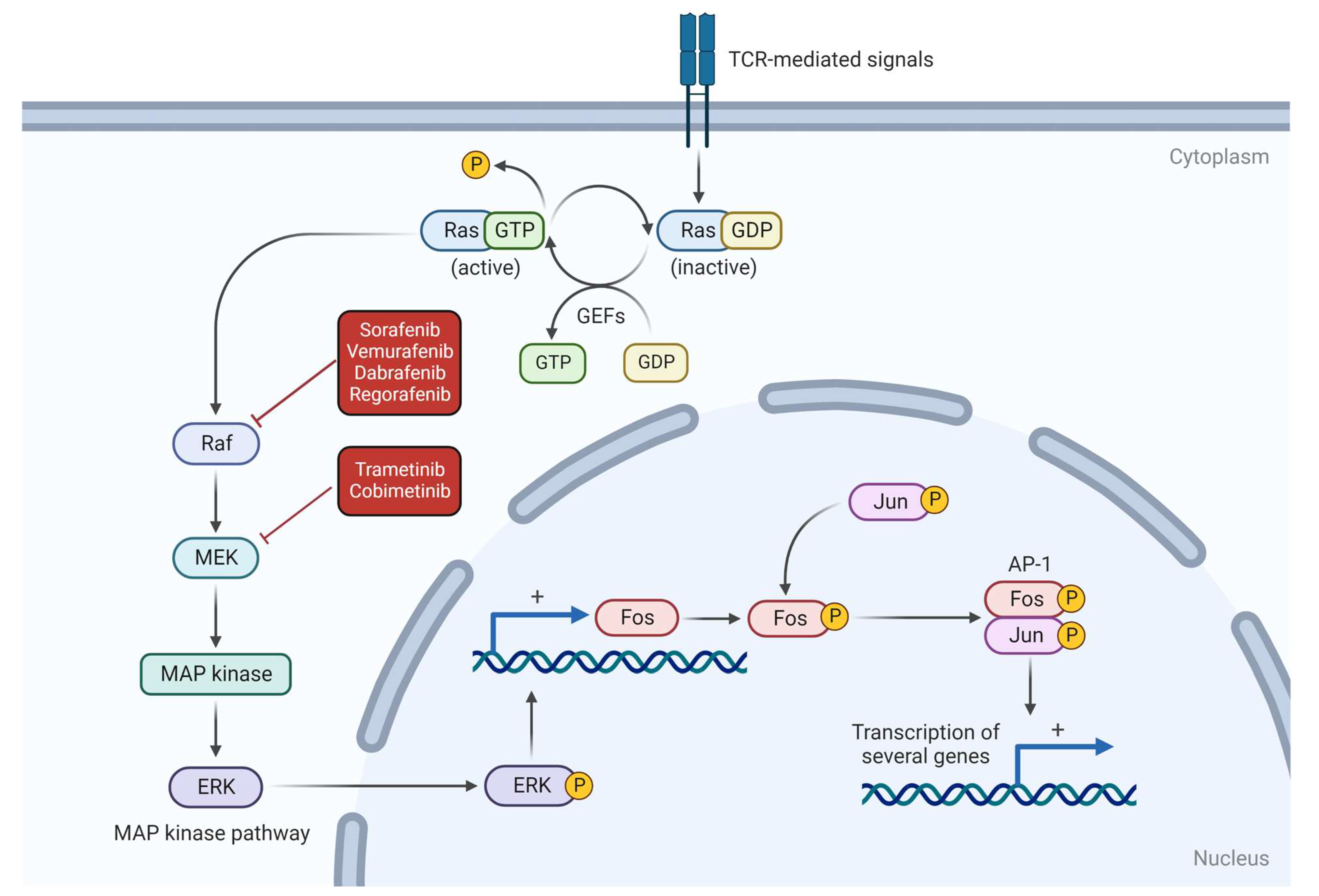
2. Hemangiosarcoma Carcinogenesis
3. Mutations and Potential Therapeutic Targets
3.1. NRAS
3.2. PIK3CA
3.3. PTEN
3.4. TP53
3.5. CDKN2A
3.6. VEGF, Angiogenesis and Hypoxia
3.7. PD-1/PD-L1 Complex
4. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naka, N.; Ohsawa, M.; Tomita, Y.; Kanno, H.; Uchida, A.; Myoui, A.; Aozasa, K. Prognostic Factors in Angiosarcoma: A Multivariate Analysis of 55 Cases. J. Surg. Oncol. 1996, 61, 170–176. [Google Scholar] [CrossRef]
- Rocha, M.S.T.; Lucci, C.M.; dos Santos, J.A.M.; Longo, J.P.F.; Muehlmann, L.A.; Azevedo, R.B. Photodynamic Therapy for Cutaneous Hemangiosarcoma in Dogs. Photodiagnosis Photodyn. Ther. 2019, 27, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Carnio, A.; Eleni, C.; Cocumelli, C.; Bartolomé Del Pino, L.E.; Simeoni, S.; Spallucci, V.; Scaramozzino, P. Evaluation of Intrinsic and Extrinsic Risk Factors for Dog Visceral Hemangiosarcoma: A Retrospective Case-Control Study Register-Based in Lazio Region, Italy. Prev. Vet. Med. 2020, 181, 105074. [Google Scholar] [CrossRef]
- Baretta, L.T.; De Oliveira Dhein, J.; Lupion, C.G.; Figueiredo, C.D.; Gerardi, D.G. Occurrence of Cutaneous Neoplasia in Dogs with Actinic Dermatitis in a Veterinary Medical Teaching Hospital—UFRGS, Brazil. Acta Sci. Vet. 2021, 49. [Google Scholar] [CrossRef]
- Ward, H.; Fox, L.E.; Calderwood-Mays, M.B.; Hammer, A.S.; Couto, C.G. Cutaneous Hemangiosarcoma in 25 Dogs: A Retrospective Study. J. Vet. Intern. Med. 1994, 8, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Hoshi, K.; Hirakawa, A.; Chimura, S.; Kobayashi, M.; Machida, N. Epidemiological, Clinical and Pathological Features of Primary Cardiac Hemangiosarcoma in Dogs: A Review of 51 Cases. J. Vet. Med. Sci. 2013, 75, 1433–1441. [Google Scholar] [CrossRef] [Green Version]
- Wendelburg, K.M.; Price, L.L.; Burgess, K.E.; Lyons, J.A.; Lew, F.H.; Berg, J. Survival Time of Dogs with Splenic Hemangiosarcoma Treated by Splenectomy with or without Adjuvant Chemotherapy: 208 Cases (2001–2012). J. Am. Vet. Med. Assoc. 2015, 247, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Story, A.L.; Wavreille, V.; Abrams, B.; Egan, A.; Cray, M.; Selmic, L.E. Outcomes of 43 Small Breed Dogs Treated for Splenic Hemangiosarcoma. Vet. Surg. 2020, 49, 1154–1163. [Google Scholar] [CrossRef] [PubMed]
- Carloni, A.; Terragni, R.; Morselli-Labate, A.M.; Paninarova, M.; Graham, J.; Valenti, P.; Alberti, M.; Albarello, G.; Millanta, F.; Vignoli, M. Prevalence, Distribution, and Clinical Characteristics of Hemangiosarcoma-Associated Skeletal Muscle Metastases in 61 Dogs: A Whole Body Computed Tomographic Study. J. Vet. Intern. Med. 2019, 33, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Fleming, J.; Giuffrida, M.A.; Runge, J.J.; Balsa, I.M.; Culp, W.T.N.; Mayhew, P.D.; Steffey, M.; Epstein, S.E. Anatomic Site and Etiology of Hemorrhage in Small versus Large Dogs with Spontaneous Hemoperitoneum. Vet. Surg. 2018, 47, 1031–1038. [Google Scholar] [CrossRef]
- Wright, L.V.; Renwick, M.; Soh, R.W.Y.; Fan, N.R.; Tebb, A.J.; Indrawirawan, Y.H. Outcomes and Blood Product Use in 89 Surgically Managed and 79 Medically Managed Cases of Acute Spontaneous Hemoperitoneum in the Dog. Front. Vet. Sci. 2021, 8, 736329. [Google Scholar] [CrossRef]
- Kim, S.E.; Liptak, J.M.; Gall, T.T.; Monteith, G.J.; Woods, J.P. Paul Woods Epirubicin in the Adjuvant Treatment of Splenic Hemangiosarcoma in Dogs: 59 Cases (1997–2004). JAVMA 2007, 231, 1550–1557. [Google Scholar]
- Matsuyama, A.; Poirier, V.J.; Mantovani, F.; Foster, R.A.; Mutsaers, A.J. Adjuvant Doxorubicin with or without Metronomic Cyclophosphamide for Canine Splenic Hemangiosarcoma. J. Am. Anim. Hosp. Assoc. 2017, 53, 304–312. [Google Scholar] [CrossRef]
- Masyr, A.R.; Rendahl, A.K.; Winter, A.L.; Borgatti, A.; Modiano, J.F. Retrospective Evaluation of Thrombocytopenia and Tumor Stage as Prognostic Indicators in Dogs with Splenic Hemangiosarcoma. J. Am. Vet. Med. Assoc. 2021, 258, 630–637. [Google Scholar] [CrossRef]
- Lamerato-Kozicki, A.R.; Helm, K.M.; Jubala, C.M.; Cutter, G.C.; Modiano, J.F. Canine Hemangiosarcoma Originates from Hematopoietic Precursors with Potential for Endothelial Differentiation. Exp. Hematol. 2006, 34, 870–878. [Google Scholar] [CrossRef]
- Kim, J.H.; Graef, A.J.; Dickerson, E.B.; Modiano, J.F. Pathobiology of Hemangiosarcoma in Dogs: Research Advances and Future Perspectives. Vet. Sci. 2015, 2, 388–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, B.D.C.; Torres, B.B.J.; Rodriguez, A.A.M.; Gamba, C.O.; Cassali, G.D.; Lavalle, G.E.; Martins, G.D.C.; Melo, E.G. Clinical and Pathological Aspects of Multicentric Hemangiosarcoma in a Pinscher Dog. Arq. Bras. Med. Vet. Zootec. 2013, 65, 322–328. [Google Scholar] [CrossRef] [Green Version]
- Gorden, B.H.; Kim, J.H.; Sarver, A.L.; Frantz, A.M.; Breen, M.; Lindblad-Toh, K.; O’Brien, T.D.; Sharkey, L.C.; Modiano, J.F.; Dickerson, E.B. Identification of Three Molecular and Functional Subtypes in Canine Hemangiosarcoma through Gene Expression Profiling and Progenitor Cell Characterization. Am. J. Pathol. 2014, 184, 985–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, K.S.; Graef, A.J.; Breen, M.; Lindblad-Toh, K.; Modiano, J.F.; Kim, J.H. Interactions between CXCR4 and CXCL12 Promote Cell Migration and Invasion of Canine Hemangiosarcoma. Vet. Comp. Oncol. 2017, 15, 315–327. [Google Scholar] [CrossRef]
- Kim, J.H.; Frantz, A.M.; Anderson, K.L.; Graef, A.J.; Scott, M.C.; Robinson, S.; Sharkey, L.C.; O’Brien, T.D.; Dickerson, E.B.; Modiano, J.F. Interleukin-8 Promotes Canine Hemangiosarcoma Growth by Regulating the Tumor Microenvironment. Exp. Cell. Res. 2014, 323, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Herman, E.J.; Stern, A.W.; Fox, R.J.; Dark, M.J. Understanding the Efficiency of Splenic Hemangiosarcoma Diagnosis Using Monte Carlo Simulations. Vet. Pathol. 2019, 56, 856–859. [Google Scholar] [CrossRef] [PubMed]
- Tecilla, M.; Gambini, M.; Forlani, A.; Caniatti, M.; Ghisleni, G.; Roccabianca, P. Evaluation of Cytological Diagnostic Accuracy for Canine Splenic Neoplasms: An Investigation in 78 Cases Using STARD Guidelines. PLoS ONE 2019, 14, e0224945. [Google Scholar] [CrossRef] [PubMed]
- Flory, A.; Kruglyak, K.M.; Tynan, J.A.; McLennan, L.M.; Rafalko, J.M.; Fiaux, P.C.; Hernandez, G.E.; Marass, F.; Nakashe, P.; Ruiz-Perez, C.A.; et al. Clinical Validation of a Next-Generation Sequencing-Based Multi-Cancer Early Detection “Liquid Biopsy” Blood Test in over 1000 Dogs Using an Independent Testing Set: The CANcer Detection in Dogs (CANDiD) Study. PLoS ONE 2022, 17, e0266623. [Google Scholar] [CrossRef] [PubMed]
- Wilson-Robles, H.M.; Bygott, T.; Kelly, T.K.; Miller, T.M.; Miller, P.; Matsushita, M.; Terrell, J.; Bougoussa, M.; Butera, T. Evaluation of Plasma Nucleosome Concentrations in Dogs with a Variety of Common Cancers and in Healthy Dogs. BMC Vet. Res. 2022, 18. [Google Scholar] [CrossRef] [PubMed]
- Scarff, D. Solar (Actinic) Dermatoses in the Dog and Cat. Companion Anim. 2017, 22, 188–196. [Google Scholar] [CrossRef]
- Nóbrega, D.F.; Sehaber, V.F.; Madureira, R.; Bracarense, A.P.F.R.L. Canine Cutaneous Haemangiosarcoma: Biomarkers and Survival. J. Comp. Pathol. 2019, 166, 87–96. [Google Scholar] [CrossRef]
- Moore, A.S.; Rassnick, K.M.; Frimberger, A.E. Evaluation of Clinical and Histologic Factors Associated with Survival Time in Dogs with Stage II Splenic Hemangiosarcoma Treated by Splenectomy. J. Am. Vet. Med. Assoc. 2017, 251, 559–565. [Google Scholar] [CrossRef]
- Lyles, S.E.; Milner, R.J.; Kow, K.; Salute, M.E. In Vitro Effects of the Tyrosine Kinase Inhibitor, Masitinib Mesylate, on Canine Hemangiosarcoma Cell Lines. Vet. Comp. Oncol. 2012, 10, 223–235. [Google Scholar] [CrossRef]
- Gardner, H.L.; London, C.A.; Portela, R.A.; Nguyen, S.; Rosenberg, M.P.; Klein, M.K.; Clifford, C.; Thamm, D.H.; Vail, D.M.; Bergman, P.; et al. Maintenance Therapy with Toceranib Following Doxorubicin-Based Chemotherapy for Canine Splenic Hemangiosarcoma. BMC Vet. Res. 2015, 11, 131. [Google Scholar] [CrossRef] [Green Version]
- Alexander, J.E.; Colyer, A.; Haydock, R.M.; Hayek, M.G.; Park, J. Understanding How Dogs Age: Longitudinal Analysis of Markers of Inflammation, Immune Function, and Oxidative Stress. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2018, 73, 720–728. [Google Scholar] [CrossRef] [Green Version]
- De Campos, C.B.; Lavalle, G.E.; Monteiro, L.N.; Arantes Pêgas, G.R.; Fialho, S.L.; Balabram, D.; Cassali, G.D. Adjuvant Thalidomide and Metronomic Chemotherapy for the Treatment of Canine Malignant Mammary Gland Neoplasms. In Vivo (Brooklyn) 2018, 32, 1659–1666. [Google Scholar] [CrossRef] [Green Version]
- Treggiari, E.; Borrego, J.F.; Gramer, I.; Valenti, P.; Harper, A.; Finotello, R.; Toni, C.; Laomedonte, P.; Romanelli, G. Retrospective Comparison of First-Line Adjuvant Anthracycline vs Metronomic-Based Chemotherapy Protocols in the Treatment of Stage I and II Canine Splenic Haemangiosarcoma. Vet. Comp. Oncol. 2020, 18, 43–51. [Google Scholar] [CrossRef]
- Finotello, R.; Stefanello, D.; Zini, E.; Marconato, L. Comparison of Doxorubicin–Cyclophosphamide with Doxorubicin–Dacarbazine for the Adjuvant Treatment of Canine Hemangiosarcoma. Vet. Comp. Oncol. 2017, 15, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Bray, J.P.; Munday, J.S. Thalidomide Reduces Vascular Endothelial Growth Factor Immunostaining in Canine Splenic Hemangiosarcoma. Vet. Sci. 2020, 7, 67. [Google Scholar] [CrossRef]
- Musser, M.L.; Coto, G.M.; Lingnan, Y.; Mochel, P.; Johannes, C.M. Pilot Safety Evaluation of Doxorubicin Chemotherapy Combined with Non-Specific Immunotherapy (Immunocidin®) for Canine Splenic Hemangiosarcoma. PLoS ONE 2022, 17, e0279594. [Google Scholar] [CrossRef]
- Lucroy, M.D.; Clauson, R.M.; Suckow, M.A.; El-Tayyeb, F.; Kalinauskas, A. Evaluation of an Autologous Cancer Vaccine for the Treatment of Metastatic Canine Hemangiosarcoma: A Preliminary Study. BMC Vet. Res. 2020, 16, 447. [Google Scholar] [CrossRef]
- Konduri, V.; Halpert, M.M.; Baig, Y.C.; Coronado, R.; Rodgers, J.R.; Levitt, J.M.; Cerroni, B.; Piscoya, S.; Wilson, N.; DiBernardi, L.; et al. Dendritic Cell Vaccination plus Low-Dose Doxorubicin for the Treatment of Spontaneous Canine Hemangiosarcoma. Cancer Gene Ther. 2019, 26, 282–291. [Google Scholar] [CrossRef] [Green Version]
- Faulhaber, E.A.; Janik, E.; Thamm, D.H. Adjuvant Carboplatin for Treatment of Splenic Hemangiosarcoma in Dogs: Retrospective Evaluation of 18 Cases (2011-2016) and Comparison with Doxorubicin-Based Chemotherapy. J. Vet. Intern. Med. 2021, 35, 1929–1934. [Google Scholar] [CrossRef] [PubMed]
- Gedney, A.; Salah, P.; Mahoney, J.A.; Krick, E.; Martins, R.; Scavello, H.; Lenz, J.A.; Atherton, M.J. Evaluation of the Anti-Tumour Activity of Coriolus Versicolor Polysaccharopeptide (I’m-Yunity) Alone or in Combination with Doxorubicin for Canine Splenic Hemangiosarcoma. Vet. Comp. Oncol. 2022, 20, 688–696. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.; Ehrhart, E.J.; Stewart, S.; Zismann, V.; Cawley, J.; Halperin, R.; Briones, N.; Richter, K.; Sivaprakasam, K.; Perdigones, N.; et al. Genomic Landscapes of Canine Splenic Angiosarcoma (Hemangiosarcoma) Contain Extensive Heterogeneity within and between Patients. PLoS ONE 2022, 17, e0264986. [Google Scholar] [CrossRef] [PubMed]
- Alsaihati, B.A.; Ho, K.L.; Watson, J.; Feng, Y.; Wang, T.; Dobbin, K.K.; Zhao, S. Canine Tumor Mutational Burden Is Correlated with TP53 Mutation across Tumor Types and Breeds. Nat. Commun. 2021, 12, 4670. [Google Scholar] [CrossRef]
- Wu, K.; Rodrigues, L.; Post, G.; Harvey, G.; White, M.; Miller, A.; Lambert, L.; Lewis, B.; Lopes, C.; Zou, J. Analyses of Canine Cancer Mutations and Treatment Outcomes Using Real-World Clinico-Genomics Data of 2119 Dogs. NPJ Precis. Oncol. 2023, 7, 8. [Google Scholar] [CrossRef]
- Wang, G.; Wu, M.; Durham, A.C.; Radaelli, E.; Mason, N.J.; Xu, X.W.; Roth, D.B. Molecular Subtypes in Canine Hemangiosarcoma Reveal Similarities with Human Angiosarcoma. PLoS ONE 2020, 15, e0229728. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Megquier, K.; Thomas, R.; Sarver, A.L.; Song, J.M.; Kim, Y.T.; Cheng, N.; Schulte, A.J.; Linden, M.A.; Murugan, P.; et al. Genomically Complex Human Angiosarcoma and Canine Hemangiosarcoma Establish Convergent Angiogenic Transcriptional Programs Driven by Novel Gene Fusions. Mol. Cancer Res. 2021, 19, 847–861. [Google Scholar] [CrossRef]
- Neganova, M.E.; Klochkov, S.G.; Aleksandrova, Y.R.; Aliev, G. Histone Modifications in Epigenetic Regulation of Cancer: Perspectives and Achieved Progress. Semin. Cancer Biol. 2022, 83, 452–471. [Google Scholar] [CrossRef]
- Suzuki, T.; Aoshima, K.; Yamazaki, J.; Kobayashi, A.; Kimura, T. Manipulating Histone Acetylation Leads to Antitumor Effects in Hemangiosarcoma Cells. Vet. Comp. Oncol. 2022, 20, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.; Borst, L.; Rotroff, D.; Motsinger-Reif, A.; Lindblad-Toh, K.; Modiano, J.F.; Breen, M. Genomic Profiling Reveals Extensive Heterogeneity in Somatic DNA Copy Number Aberrations of Canine Hemangiosarcoma. Chromosom. Res. 2014, 22, 305–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penel, N.; Ray-Coquard, I.; Bal-Mahieu, C.; Chevreau, C.; Le Cesne, A.; Italiano, A.; Bompas, E.; Clisant, S.; Baldeyrou, B.; Lansiaux, A.; et al. Low Level of Baseline Circulating VEGF-A Is Associated with Better Outcome in Patients with Vascular Sarcomas Receiving Sorafenib: An Ancillary Study from a Phase II Trial. Target Oncol. 2014, 9, 273–277. [Google Scholar] [CrossRef]
- Wan, H.; Zhang, D.; Hu, W.; Xie, Z.; Du, Q.; Xia, Q.; Wen, T.; Jia, H. Aberrant PTEN, PIK3CA, PMAPK, and TP53 Expression in Human Scalp and Face Angiosarcoma. Medicine 2021, 100, e26779. [Google Scholar] [CrossRef]
- Conciatori, F.; Bazzichetto, C.; Falcone, I.; Ciuffreda, L.; Ferretti, G.; Vari, S.; Ferraresi, V.; Cognetti, F.; Milella, M. Pten Function at the Interface between Cancer and Tumor Microenvironment: Implications for Response to Immunotherapy. Int. J. Mol. Sci. 2020, 21, 5337. [Google Scholar] [CrossRef]
- Papa, A.; Pandolfi, P.P. The PTEN–PI3K Axis in Cancer. Biomolecules 2019, 9, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamburini, B.A.; Trapp, S.; Phang, T.L.; Schappa, J.T.; Hunter, L.E.; Modiano, J.F. Gene Expression Profiles of Sporadic Canine Hemangiosarcoma Are Uniquely Associated with Breed. PLoS ONE 2009, 4, e5549. [Google Scholar] [CrossRef]
- Wang, G.; Wu, M.; Maloneyhuss, M.A.; Wojcik, J.; Durham, A.C.; Mason, N.J.; Roth, D.B. Actionable Mutations in Canine Hemangiosarcoma. PLoS ONE 2017, 12, e0188667. [Google Scholar] [CrossRef] [Green Version]
- Clendaniel, D.C.; Sivacolundhu, R.K.; Sorenmo, K.U.; Donovan, T.A.; Turner, A.; Arteaga, T.; Bergman, P.J. Association between Macroscopic Appearance of Liver Lesions and Liver Histology in Dogs with Splenic Hemangiosarcoma: 79 Cases (2004–2009). J. Am. Anim. Hosp. Assoc. 2014, 50, e6–e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batschinski, K.; Nobre, A.; Vargas-Mendez, E.; Tedardi, M.V.; Cirillo, J.; Cestari, G.; Ubukata, R.; Dagli, M.L.Z. Canine Visceral Hemangiosarcoma Treated with Surgery Alone or Surgery and Doxorubicin: 37 Cases (2005–2014). Can. Vet. J. 2018, 59, 967–972. [Google Scholar] [PubMed]
- García-Iglesias, M.J.; Cuevas-Higuera, J.L.; Bastida-Sáenz, A.; De Garnica-García, M.G.; Polledo, L.; Perero, P.; González-Fernández, J.; Fernández-Martínez, B.; Pérez-Martínez, C. Immunohistochemical Detection of P53 and Pp53 Ser392in Canine Hemangiomas and Hemangiosarcomas Located in the Skin. BMC Vet. Res. 2020, 16, 239. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.L.; Bryan, M.E.; Atkinson, E.S.; Keeler, M.R.; Hahn, A.W.; Bryan, J.N. Neutering Is Associated with Developing Hemangiosarcoma in Dogs in the Veterinary Medical Database: An Age and Time-Period Matched Case-Control Study (1964–2003). Can. Vet. J. 2020, 61, 499–504. [Google Scholar]
- Sherwood, J.M.; Haynes, A.M.; Klocke, E.; Higginbotham, M.L.; Thomson, E.M.; Weng, H.Y.; Millard, H.A.T. Occurrence and Clinicopathologic Features of Splenic Neoplasia Based on Body Weight: 325 Dogs (2003–2013). J. Am. Anim. Hosp. Assoc. 2016, 52, 220–226. [Google Scholar] [CrossRef]
- Torres de la Riva, G.; Hart, B.L.; Farver, T.B.; Oberbauer, A.M.; Messam, L.L.M.V.; Willits, N.; Hart, L.A. Neutering Dogs: Effects on Joint Disorders and Cancers in Golden Retrievers. PLoS ONE 2013, 8, 55937. [Google Scholar] [CrossRef]
- Hart, B.L.; Hart, L.A.; Thigpen, A.P.; Willits, N.H. Neutering of German Shepherd Dogs: Associated Joint Disorders, Cancers and Urinary Incontinence. Vet. Med. Sci. 2016, 2, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Maeda, M.; Ochiai, K.; Michishita, M.; Morimatsu, M.; Sakai, H.; Kinoshita, N.; Sakaue, M.; Onozawa, E.; Azakami, D.; Yamamoto, M.; et al. In Vitro Anticancer Effects of Alpelisib against PIK3CA-mutated Canine Hemangiosarcoma Cell Lines. Oncol. Rep. 2022, 47, 84. [Google Scholar] [CrossRef] [PubMed]
- Igase, M.; Fujiki, N.; Shibutani, S.; Sakai, H.; Noguchi, S.; Nemoto, Y.; Mizuno, T. Tenovin-6 Induces the SIRT-Independent Cell Growth Suppression and Blocks Autophagy Flux in Canine Hemangiosarcoma Cell Lines. Exp. Cell. Res. 2020, 388. [Google Scholar] [CrossRef] [PubMed]
- Megquier, K.; Turner-Maier, J.; Swofford, R.; Kim, J.H.; Sarver, A.L.; Wang, C.; Sakthikumar, S.; Johnson, J.; Koltookian, M.; Lewellen, M.; et al. Comparative Genomics Reveals Shared Mutational Landscape in Canine Hemangiosarcoma and Human Angiosarcoma. Mol. Cancer Res. 2019, 17, 2410–2421. [Google Scholar] [CrossRef] [Green Version]
- Dickerson, E.B.; Thomas, R.; Fosmire, S.P.; Lamerato-Kozicki, A.R.; Bianco, S.R.; Wojcieszyn, J.W.; Breen, M.; Helfand, S.C.; Modiano, J.F. Mutations of Phosphatase and Tensin Homolog Deleted from Chromosome 10 in Canine Hemangiosarcoma. Vet. Pathol. 2005, 42, 618–632. [Google Scholar] [CrossRef]
- Wong, K.; Ludwig, L.; Krijgsman, O.; Adams, D.J.; Wood, G.A.; Van Der Weyden, L. Comparison of the Oncogenomic Landscape of Canine and Feline Hemangiosarcoma Shows Novel Parallels with Human Angiosarcoma. DMM Dis. Models Mechan. 2021, 14, dmm049044. [Google Scholar] [CrossRef]
- Hasbullah, H.H.; Musa, M. Gene Therapy Targeting P53 and Kras for Colorectal Cancer Treatment: A Myth or the Way Forward? Int. J. Mol. Sci. 2021, 22, 11941. [Google Scholar] [CrossRef]
- Zhang, W.W.; Li, L.; Li, D.; Liu, J.; Li, X.; Li, W.; Xu, X.; Zhang, M.J.; Chandler, L.A.; Lin, H.; et al. The First Approved Gene Therapy Product for Cancer Ad-P53 (Gendicine): 12 Years in the Clinic. Hum. Gene Ther. 2018, 29, 160–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igase, M.; Inanaga, S.; Tani, K.; Nakaichi, M.; Sakai, Y.; Sakurai, M.; Kato, M.; Tsukui, T.; Mizuno, T. Long-term Survival of Dogs with Stage 4 Oral Malignant Melanoma Treated with Anti-canine Therapeutic Antibody: A Follow-up Case Report. Vet. Comp. Oncol. 2022, 20, 901–905. [Google Scholar] [CrossRef]
- Pantelyushin, S.; Ranninger, E.; Guerrera, D.; Hutter, G.; Maake, C.; Markkanen, E.; Bettschart-Wolfensberger, R.; Bley, C.R.; Läubli, H.; Vom Berg, J. Cross-Reactivity and Functionality of Approved Human Immune Checkpoint Blockers in Dogs. Cancers 2021, 13, 785. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, M.; Nie, H.; Yuan, Y. PD-1 and PD-L1 in Cancer Immunotherapy: Clinical Implications and Future Considerations. Hum. Vaccin. Immunother. 2019, 15, 1111–1122. [Google Scholar] [CrossRef]
- Boston, S.E.; Lu, X.; Culp, W.T.N.; Montinaro, V.; Romanelli, G.; Dudley, R.M.; Liptak, J.M.; Mestrinho, L.A.; Buracco, P. Efficacy of Systemic Adjuvant Therapies Administered to Dogs after Excision of Oral Malignant Melanomas: 151 Cases (2001–2012). J. Am. Vet. Med. Assoc. 2014, 245, 401–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cawley, J.R.; Stewart, S.D.; Mochel, J.P.; Veluvolu, S.; Khanna, C.; Fenger, J.M. Pharmacokinetic Exposures Associated With Oral Administration of Sorafenib in Dogs With Spontaneous Tumors. Front. Vet. Sci. 2022, 9, 888483. [Google Scholar] [CrossRef] [PubMed]
- Marconato, L.; Sabattini, S.; Marisi, G.; Rossi, F.; Leone, V.F.; Casadei-Gardini, A. Sorafenib for the Treatment of Unresectable Hepatocellular Carcinoma: Preliminary Toxicity and Activity Data in Dogs. Cancers 2020, 12, 1272. [Google Scholar] [CrossRef]
- Foskett, A.; Manley, C.; Naramore, R.; Gordon, I.; Stewart, B.; Khanna, C. Tolerability of Oral Sorafenib in Pet Dogs with a Diagnosis of Cancer. Vet. Med. Res. Rep. 2017, 8, 97–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murase, Y.; Hosoya, K.; Sato, T.; Kim, S.; Okumura, M. Antitumor Activity of the Dual PI3K/MTOR Inhibitor Gedatolisib and the Involvement of ABCB1 in Gedatolisib Resistance in Canine Tumor Cells. Oncol. Rep. 2022, 47, 61. [Google Scholar] [CrossRef]
- Wilson, G.D.; Wilson, T.G.; Hanna, A.; Dabjan, M.; Buelow, K.; Torma, J.; Marples, B.; Galoforo, S. Dacomitinib and Gedatolisib in Combination with Fractionated Radiation in Head and Neck Cancer. Clin. Transl. Radiat. Oncol. 2021, 26, 15–23. [Google Scholar] [CrossRef]
- Liu, C.; Xing, W.; Yu, H.; Zhang, W.; Si, T. ABCB1 and ABCG2 Restricts the Efficacy of Gedatolisib (PF-05212384), a PI3K Inhibitor in Colorectal Cancer Cells. Cancer Cell Int. 2021, 21, 108. [Google Scholar] [CrossRef]
- Markham, A. Alpelisib: First Global Approval. Drugs 2019, 79, 1249–1253. [Google Scholar] [CrossRef]
- Prior, I.A.; Hood, F.E.; Hartley, J.L. The Frequency of Ras Mutations in Cancer. Cancer Res. 2020, 80, 2669–2974. [Google Scholar] [CrossRef] [Green Version]
- Usher, S.G.; Radford, A.D.; Villiers, E.J.; Blackwood, L. RAS, FLT3, and C-KIT Mutations in Immunophenotyped Canine Leukemias. Exp. Hematol. 2009, 37, 65–77. [Google Scholar] [CrossRef]
- Gong, B.; Liu, W.W.; Nie, W.J.; Li, D.F.; Xie, Z.J.; Liu, C.; Liu, Y.H.; Mei, P.; Li, Z.J. MiR-21/RASA1 Axis Affects Malignancy of Colon Cancer Cells via RAS Pathways. World J. Gastroenterol. 2015, 21, 1488–1497. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, H.; Breen, M. Sequence Analysis of RAS and RAF Mutation Hot Spots in Canine Carcinoma. Vet. Comp. Oncol. 2017, 15, 1598–1605. [Google Scholar] [CrossRef] [PubMed]
- Mayr, B.; Schaffner, G.; Reifinger, M. K-Ras Mutations in Canine Pancreatic Cancers. Vet. Rec. 2003, 153, 87–89. [Google Scholar] [CrossRef]
- Griffey, S.M.; Kraegel, S.A.; Madewell, B.R. Rapid Detection of K-Ras Gene Mutations in Canine Lung Cancer Using Single-Strand Conformational Polymorphism Analysis. Carcinogenesis 1998, 19, 959–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degirmenci, U.; Wang, M.; Hu, J. Targeting Aberrant RAS/RAF/MEK/ERK Signaling for Cancer Therapy. Cells 2020, 9, 198. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, Y.; Wang, Q.; Su, B.; Xu, H.; Sun, Y.; Sun, P.; Li, R.; Peng, X.; Cai, J. Role of RASA1 in Cancer: A Review and Update (Review). Oncol. Rep. 2020, 44, 2386–2396. [Google Scholar] [CrossRef]
- Xiao, W.; Zheng, S.; Zou, Y.; Yang, A.; Xie, X.; Tang, H.; Xie, X. CircAHNAK1 Inhibits Proliferation and Metastasis of Triple-Negative Breast Cancer by Modulating MiR-421 and RASA1. Aging 2019, 11, 12043–12056. [Google Scholar] [CrossRef]
- Pottier, C.; Fresnais, M.; Gilon, M.; Jérusalem, G.; Longuespée, R.; Sounni, N.E. Tyrosine Kinase Inhibitors in Cancer: Breakthrough and Challenges of Targeted Therapy. Cancers 2020, 12, 731. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Huang, J.; Ma, L.; Shan, J.; Shen, J.; Yang, Z.; Liu, L.; Luo, Y.; Yao, C.; Qian, C. MicroRNA-122 Confers Sorafenib Resistance to Hepatocellular Carcinoma Cells by Targeting IGF-1R to Regulate RAS/RAF/ERK Signaling Pathways. Cancer Lett. 2016, 371, 171–181. [Google Scholar] [CrossRef]
- Andersen, N.J.; Nickoloff, B.J.; Dykema, K.J.; Boguslawski, E.A.; Krivochenitser, R.I.; Froman, R.E.; Dawes, M.J.; Baker, L.H.; Thomas, D.G.; Kamstock, D.A.; et al. Pharmacologic Inhibition of MEK Signaling Prevents Growth of Canine Hemangiosarcoma. Mol. Cancer Ther. 2013, 12, 1701–1714. [Google Scholar] [CrossRef] [Green Version]
- Signorelli, J.; Shah Gandhi, A. Cobimetinib: A Novel MEK Inhibitor for Metastatic Melanoma. Ann. Pharmacother. 2017, 51, 146–153. [Google Scholar] [CrossRef]
- Morales, M.L.; Arenas, A.; Ortiz-Ruiz, A.; Leivas, A.; Rapado, I.; Rodríguez-García, A.; Castro, N.; Zagorac, I.; Quintela-Fandino, M.; Gómez-López, G.; et al. MEK Inhibition Enhances the Response to Tyrosine Kinase Inhibitors in Acute Myeloid Leukemia. Sci. Rep. 2019, 9, 18630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cathomas, G. PIK3CA in Colorectal Cancer. Front. Oncol. 2014, 4, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arafeh, R.; Samuels, Y. PIK3CA in Cancer: The Past 30 Years. Semin. Cancer Biol. 2019, 59, 36–49. [Google Scholar] [CrossRef]
- Murai, A.; Asa, S.A.; Kodama, A.; Hirata, A.; Yanai, T.; Sakai, H. Constitutive Phosphorylation of the MTORC2/Akt/4E-BP1 Pathway in Newly Derived Canine Hemangiosarcoma Cell Lines. BMC Vet. Res. 2012, 8, 128. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langdon, S.P.; Kay, C.; Um, I.H.; Dodds, M.; Muir, M.; Sellar, G.; Kan, J.; Gourley, C.; Harrison, D.J. Evaluation of the Dual MTOR/PI3K Inhibitors Gedatolisib (PF-05212384) and PF-04691502 against Ovarian Cancer Xenograft Models. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Pyuen, A.A.; Meuten, T.; Rose, B.J.; Thamm, D.H. In Vitro Effects of PI3K/MTOR Inhibition in Canine Hemangiosarcoma. PLoS ONE 2022, 13, e0200634. [Google Scholar] [CrossRef] [Green Version]
- Oda, K.; Okada, J.; Timmerman, L.; Rodriguez-Viciana, P.; Stokoe, D.; Shoji, K.; Taketani, Y.; Kuramoto, H.; Knight, Z.A.; Shokat, K.M.; et al. PIK3CA Cooperates with Other Phosphatidylinositol 3′-Kinase Pathway Mutations to Effect Oncogenic Transformation. Cancer Res. 2008, 68, 8127–8136. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Seung, B.J.; Cho, S.H.; Lim, H.Y.; Bae, M.K.; Sur, J.H. Dysregulation of Pi3k/Akt/Pten Pathway in Canine Mammary Tumor. Animals 2021, 11, 2079. [Google Scholar] [CrossRef]
- Yu, L.; Wei, J.; Liu, P. Attacking the PI3K/Akt/MTOR Signaling Pathway for Targeted Therapeutic Treatment in Human Cancer. Semin. Cancer Biol. 2022, 85, 69–94. [Google Scholar] [CrossRef]
- Jamaspishvili, T.; Berman, D.M.; Ross, A.E.; Scher, H.I.; De Marzo, A.M.; Squire, J.A.; Lotan, T.L. Clinical Implications of PTEN Loss in Prostate Cancer. Nat. Rev. Urol. 2018, 15, 222–234. [Google Scholar] [CrossRef]
- Álvarez-Garcia, V.; Tawil, Y.; Wise, H.M.; Leslie, N.R. Mechanisms of PTEN Loss in Cancer: It’s All about Diversity. Semin. Cancer Biol. 2019, 59, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.; Lin, D.; Wang, J.; Wang, L. Expression and Significance of PTEN in Canine Mammary Gland Tumours. Res. Vet. Sci. 2008, 85, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Wyeth Pharmaceuticals. TORISELTMKit (Temsirolimus). U.S Patent No. 5,362,718, 2 January 2007.
- Motzer, R.J.; Escudier, B.; Oudard, S.; Hutson, T.E.; Porta, C.; Bracarda, S.; Grünwald, V.; Thompson, J.A.; Figlin, R.A.; Hollaender, N.; et al. Efficacy of Everolimus in Advanced Renal Cell Carcinoma: A Double-Blind, Randomised, Placebo-Controlled Phase III Trial. Lancet 2008, 372, 449–456. [Google Scholar] [CrossRef]
- Yoo, C.; Lee, J.; Rha, S.Y.; Park, K.H.; Kim, T.M.; Kim, Y.J.; Lee, H.J.; Lee, K.H.; Ahn, J.H. Multicenter Phase II Study of Everolimus in Patients with Metastatic or Recurrent Bone and Soft-Tissue Sarcomas after Failure of Anthracycline and Ifosfamide. Invest. New Drugs 2013, 31, 1602–1608. [Google Scholar] [CrossRef] [PubMed]
- Michishita, M.; Ochiai, K.; Nakahira, R.; Azakami, D.; Machida, Y.; Nagashima, T.; Nakagawa, T.; Ishiwata, T. MTOR Pathway as a Potential Therapeutic Target for Cancer Stem Cells in Canine Mammary Carcinoma. Front. Oncol. 2023, 13, 1100602. [Google Scholar] [CrossRef]
- LeBlanc, A.K.; Mazcko, C.N.; Cherukuri, A.; Berger, E.P.; Kisseberth, W.C.; Brown, M.E.; Lana, S.E.; Weishaar, K.; Flesner, B.K.; Bryan, J.N.; et al. Adjuvant Sirolimus Does Not Improve Outcome in Pet Dogs Receiving Standard-of-Care Therapy for Appendicular Osteosarcoma: A Prospective, Randomized Trial of 324 Dogs. Clin. Cancer Res. 2021, 27, 3005–3016. [Google Scholar] [CrossRef]
- Giuliano, A.; Horta, R.S.; Vieira, R.A.M.; Hume, K.R.; Dobson, J. Repurposing Drugs in Small Animal Oncology Repurposing Drugs in Small Animal Oncology. Animals 2023, 13, 139. [Google Scholar] [CrossRef]
- Marei, H.E.; Althani, A.; Afifi, N.; Hasan, A.; Caceci, T.; Pozzoli, G.; Morrione, A.; Giordano, A.; Cenciarelli, C. p53 Signaling in Cancer Progression and Therapy. Cancer Cell Int. 2021, 21, 703. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, C.E.; High, K.A.; Joung, J.K.; Kohn, D.B.; Ozawa, K.; Sadelain, M. Gene Therapy Comes of Age. Science 2018, 359, eaan4672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Simon, M.; Seluanov, A.; Gorbunova, V. DNA Damage and Repair in Age-Related Inflammation. Nat. Rev. Immunol. 2023, 23, 75–89. [Google Scholar] [CrossRef]
- Sargen, M.R.; Merrill, S.L.; Chu, E.Y.; Nathanson, K.L. CDKN2A Mutations with P14 Loss Predisposing to Multiple Nerve Sheath Tumours, Melanoma, Dysplastic Naevi and Internal Malignancies: A Case Series and Review of the Literature. British J. Dermatol. 2016, 175, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Choi, B.Y.; Lee, M.H.; Bode, A.M.; Dong, Z. Implications of Genetic and Epigenetic Alterations of CDKN2A (P16INK4a) in Cancer. EBioMedicine 2016, 8, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Cicenas, J.; Kvederaviciute, K.; Meskinyte, I.; Meskinyte-Kausiliene, E.; Skeberdyte, A. KRAS, TP53, CDKN2A, SMAD4, BRCA1, and BRCA2 Mutations in Pancreatic Cancer. Cancers 2017, 9, 42. [Google Scholar] [CrossRef] [Green Version]
- Miller, T.I.; Zoumberos, N.A.; Johnson, B.; Rhodes, D.R.; Tomlins, S.A.; Chan, M.P.; Andea, A.A.; Lucas, D.R.; McHugh, J.B.; Smith, N.; et al. A Genomic Survey of Sarcomas on Sun-Exposed Skin Reveals Distinctive Candidate Drivers and Potentially Targetable Mutations. Hum. Pathol. 2020, 102, 60–69. [Google Scholar] [CrossRef]
- Brandt, L.P.; Albers, J.; Hejhal, T.; Pfundstein, S.; Gonçalves, A.F.; Catalano, A.; Wild, P.J.; Frew, I.J. Mouse Genetic Background Influences Whether Hras G12V Expression plus Cdkn2a Knockdown Causes Angiosarcoma or Undifferentiated Pleomorphic Sarcoma. Oncotarget 2018, 9, 19753–19766. [Google Scholar] [CrossRef] [Green Version]
- Melincovici, C.S.; Boşca, A.B.; Şuşman, S.; Mărginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.-M.; Roman, A.L.; Mihu, C.M. Vascular Endothelial Growth Factor (VEGF)-Key Factor in Normal and Pathological Angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar]
- Nejad, A.E.; Najafgholian, S.; Rostami, A.; Sistani, A.; Shojaeifar, S.; Esparvarinha, M.; Nedaeinia, R.; Haghjooy Javanmard, S.; Taherian, M.; Ahmadlou, M.; et al. The Role of Hypoxia in the Tumor Microenvironment and Development of Cancer Stem Cell: A Novel Approach to Developing Treatment. Cancer Cell Int. 2021, 21, 62. [Google Scholar] [CrossRef]
- Li, X.; Gao, Y.; Li, J.; Zhang, K.; Han, J.; Li, W.; Hao, Q.; Zhang, W.; Wang, S.; Zeng, C.; et al. FOXP3 Inhibits Angiogenesis by Downregulating VEGF in Breast Cancer. Cell. Death Dis. 2018, 9, 744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, R.J.; Woll, P.J. Anti-Angiogenic Therapies for the Treatment of Angiosarcoma: A Clinical Update. Mag. Eur. Med. Oncol. 2017, 10, 190–193. [Google Scholar] [CrossRef] [Green Version]
- Hegde, P.S.; Wallin, J.J.; Mancao, C. Predictive Markers of Anti-VEGF and Emerging Role of Angiogenesis Inhibitors as Immunotherapeutics. Semin. Cancer Biol. 2018, 52, 117–124. [Google Scholar] [CrossRef]
- Maeda-Otsuka, S.; Kajihara, I.; Tasaki, Y.; Yamada-Kanazawa, S.; Sakamoto, R.; Sawamura, S.; Masuzawa, M.; Masuzawa, M.; Amoh, Y.; Hoshina, D.; et al. Hypoxia Accelerates the Progression of Angiosarcoma through the Regulation of Angiosarcoma Cells and Tumor Microenvironment. J. Dermatol. Sci. 2019, 93, 123–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moschetta, M.G.; Maschio, L.B.; Jardim-Perassi, B.V.; Gelaleti, G.B.; Lopes, J.R.; Leonel, C.; Do Nascimento Goncalves, N.; Ferreira, L.C.; Martins, G.R.; Borin, T.F.; et al. Prognostic Value of Vascular Endothelial Growth Factor and Hypoxia-Inducible Factor 1α in Canine Malignant Mammary Tumors. Oncol. Rep. 2015, 33, 2345–2353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.; Chen, Y.; Jiang, X.; Peng, M.; Liu, Y.; Mo, Y.; Ren, D.; Hua, Y.; Yu, B.; Zhou, Y.; et al. Mechanisms of Vasculogenic Mimicry in Hypoxic Tumor Microenvironments. Mol. Cancer 2021, 20, 7. [Google Scholar] [CrossRef]
- Walsh, J.C.; Lebedev, A.; Aten, E.; Madsen, K.; Marciano, L.; Kolb, H.C. The Clinical Importance of Assessing Tumor Hypoxia: Relationship of Tumor Hypoxia to Prognosis and Therapeutic Opportunities. Antioxid. Redox Signal. 2014, 21, 1516–1554. [Google Scholar] [CrossRef]
- Ghahremani, M.F.; Radaelli, E.; Haigh, K.; Bartunkova, S.; Haenebalcke, L.; Marine, J.C.; Goossens, S.; Haigh, J.J. Loss of Autocrine Endothelial-Derived VEGF Significantly Reduces Hemangiosarcoma Development in Conditional P53-Deficient Mice. Cell Cycle 2014, 13, 1502–1508. [Google Scholar] [CrossRef] [Green Version]
- Adachi, M.; Hoshino, Y.; Izumi, Y.; Takagi, S. Immunohistochemical Detection of a Potential Molecular Therapeutic Target for Canine Hemangiosarcoma. J. Vet. Med. Sci. 2016, 78, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Lebellec, L.; Bertucci, F.; Tresch-Bruneel, E.; Ray-Coquard, I.; Le Cesne, A.; Bompas, E.; Blay, J.Y.; Italiano, A.; Mir, O.; Ryckewaert, T.; et al. Prognostic and Predictive Factors for Angiosarcoma Patients Receiving Paclitaxel Once Weekly plus or Minus Bevacizumab: An Ancillary Study Derived from a Randomized Clinical Trial. BMC Cancer 2018, 18, 963. [Google Scholar] [CrossRef] [Green Version]
- Campos, G.A. Juliana Alvares Duarte Bonini Campos; Daniel Soares Sanches; Maria Lúcia Zaidan Dagli; Julia Maria Matera Immunohistochemical Evaluation of Vascular Endothelial Growth Factor (VEGF) in Splenic Hemangiomas and Hemangiosarcomas in Dogs. Open J. Vet. Med. 2012, 2, 191–195. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Wang, J.; He, C.; Fang, M. Angiosarcoma: A Review of Diagnosis and Current Treatment. Am. J. Cancer Res. 2019, 9, 2303–2313. [Google Scholar] [PubMed]
- Sturm, E.C.; Marasco, I.S.; Katz, S.C. Multidisciplinary Management of Angiosarcoma—A Review. J. Surg. Res. 2021, 257, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Verganti, S.; Berlato, D.; Blackwood, L.; Amores-Fuster, I.; Polton, G.A.; Elders, R.; Doyle, R.; Taylor, A.; Murphy, S. Use of Oncept Melanoma Vaccine in 69 Canine Oral Malignant Melanomas in the UK. J. Small Anim. Pract. 2017, 58, 10–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, J.C.C.; MacIel, M.C.; Ferrari, H.A.; Gonçalves, J.M.; Sasaki, B.R.S.; Cintra, A.A.; Durán, N.; Billis, A.; Fávaro, W.J. Safety and Efficacy of OncoTherad Nano-Immunotherapy in Patients with Non-Muscle Invasive Bladder Cancer. J. Phys. Conf. Ser. 2021, 1953, 012012. [Google Scholar] [CrossRef]
- Böckelmann, P.K.; Tizziani, S.H.S.; Durán, N.; Fávaro, W.J. New Therapeutic Perspective for Bladder Cancer in Dogs: Toxicological and Clinical Effects of OncoTherad Nanostructured Immunotherapy. J. Phys. Conf. Ser. 2019, 1323, 012022. [Google Scholar] [CrossRef]
- Durán, N.; Dias, Q.C.; Fávaro, W.J. OncoTherad: A New Nanobiological Response Modifier, Its Toxicological and Anticancer Activities. J. Phys. Conf. Ser. 2019, 1323, 012018. [Google Scholar] [CrossRef]
- Rhea, L.P.; Mendez-Marti, S.; Kim, D.; Aragon-Ching, J.B. Role of Immunotherapy in Bladder Cancer. Cancer Treat Res. Commun. 2021, 26, 100296. [Google Scholar] [CrossRef]
- Gulay, K.C.M.; Aoshima, K.; Maekawa, N.; Suzuki, T.; Konnai, S.; Kobayashi, A.; Kimura, T. Hemangiosarcoma Cells Induce M2 Polarization and PD-L1 Expression in Macrophages. Sci. Rep. 2022, 12. [Google Scholar] [CrossRef]
- Maekawa, N.; Konnai, S.; Okagawa, T.; Nishimori, A.; Ikebuchi, R.; Izumi, Y.; Takagi, S.; Kagawa, Y.; Nakajima, C.; Suzuki, Y.; et al. Immunohistochemical Analysis of PD-L1 Expression in Canine Malignant Cancers and PD-1 Expression on Lymphocytes in Canine Oral Melanoma. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Reis, I.B.; Tibo, L.H.S.; de Souza, B.R.; Durán, N.; Fávaro, W.J. OncoTherad® Is an Immunomodulator of Biological Response That Downregulate RANK/RANKL Signaling Pathway and PD-1/PD-L1 Immune Checkpoint in Non-Muscle Invasive Bladder Cancer. J. Cancer Res. Clin. Oncol. 2022, 20–30. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene | Role | Reference |
|---|---|---|
| AKT1 | Oncogene | Wong et al. (2022) [40]. |
| CDKN2A | Tumoral supressor gene | Wong et al. (2022), Thomas et al. (2014) [40,47]. |
| EGFR | Oncogene | Wu et al. (2023), Wong et al. (2022) [40,42]. |
| NRAS | Oncogene | Wu et al. (2023), Wong et al. (2022), Alsaihati et al. (2021), Kim et al. (2021), Wang et al. (2020) [40,41,42,43,44]. |
| PIK3CA | Oncogene | Wu et al. (2023), Wong et al. (2022), Alsaihati et al. (2021), Kim et al. (2021), Wang et al. (2020), Megquier et al. (2019), Wang et al. (2017) [40,41,42,43,44,53,65]. |
| PTEN | Tumoral supressor gene | Wong et al. (2022), Wang et al. (2020), Megquier et al. (2019), Wang et al. (2017), Dickerson et al. (2005) [40,43,53,63,65]. |
| RASA1 | Oncogene | Wong et al. (2021), Megquier et al. (2019) [64,65]. |
| TP53 | Tumoral supressor gene | Wu et al. (2023), Wong et al. (2022), Alsaihati et al. (2021), Kim et al. (2021), García-Iglesias et al. (2020), Wang et al. (2020), Megquier et al. (2019), Wang et al. (2017) [40,41,42,44,53,56,65]. |
| Targets | Targeted Therapies, Experiment Conditions (Species) | Reference |
|---|---|---|
| Histone acetylation | JQ1/BETi, in vitro and in vivo (humans and dogs). | Neganova et al. (2022), Suzuki et al. (2022) [45,46]. |
| p53 | Gendicine/recombinant human p53 adenovirus, in vitro and in vivo (humans). | Hasbullah and Musa (2021), Zhang et al. (2018) [66,67]. |
| PD-1/PD-L1 complex | Pembrolizumab/ICI, in vitro and in vivo (humans). Atezolizumab/ICI, in vitro and in vivo (humans) and ca-4F12-E6 (dogs). | Igase et al. (2022), Pantelyushi et al. (2021), Jiang et al. (2019) [68,69,70,71]. |
| PDGFR | Sorafenib/TKI, in vitro and in vivo (humans and dogs). | Cawley et al. (2022), Marconato et al. (2020), Foskett et al. (2017) [72,73,74]. |
| PI3K/AKT/mTOR | Alpelisib/PIK3 inhibitor, in vitro (humans and dogs) and in vivo (humans), and Gedatolisib/PIK3 inhibitor, in vitro and in vivo (humans). | Maeda et al. (2022), Murase et al. (2022), Wilson et al. (2021), Liu et al. (2021), Markham (2019) [61,75,76,77,78]. |
| RAS-RAF-MEK | Sorafenib/TKI, in vitro and in vivo (humans, dogs). | Cawley et al. (2022), Marconato et al. (2020), Foskett et al. (2017) [72,73,74]. |
| VEGFR | Sorafenib and Toceranib/TKI, in vitro and in vivo (humans and dogs). | Cawley et al. (2022), Marconato et al. (2020), Foskett et al. (2017) [72,73,74]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pimentel, P.A.B.; Giuliano, A.; Bęczkowski, P.M.; Horta, R.D.S. Molecular Profile of Canine Hemangiosarcoma and Potential Novel Therapeutic Targets. Vet. Sci. 2023, 10, 387. https://doi.org/10.3390/vetsci10060387
Pimentel PAB, Giuliano A, Bęczkowski PM, Horta RDS. Molecular Profile of Canine Hemangiosarcoma and Potential Novel Therapeutic Targets. Veterinary Sciences. 2023; 10(6):387. https://doi.org/10.3390/vetsci10060387
Chicago/Turabian StylePimentel, Pedro Antônio Bronhara, Antonio Giuliano, Paweł Marek Bęczkowski, and Rodrigo Dos Santos Horta. 2023. "Molecular Profile of Canine Hemangiosarcoma and Potential Novel Therapeutic Targets" Veterinary Sciences 10, no. 6: 387. https://doi.org/10.3390/vetsci10060387