
Rottlerin-Liposome Inhibits the Endocytosis of Feline Coronavirus Infection
 , , and
, , and 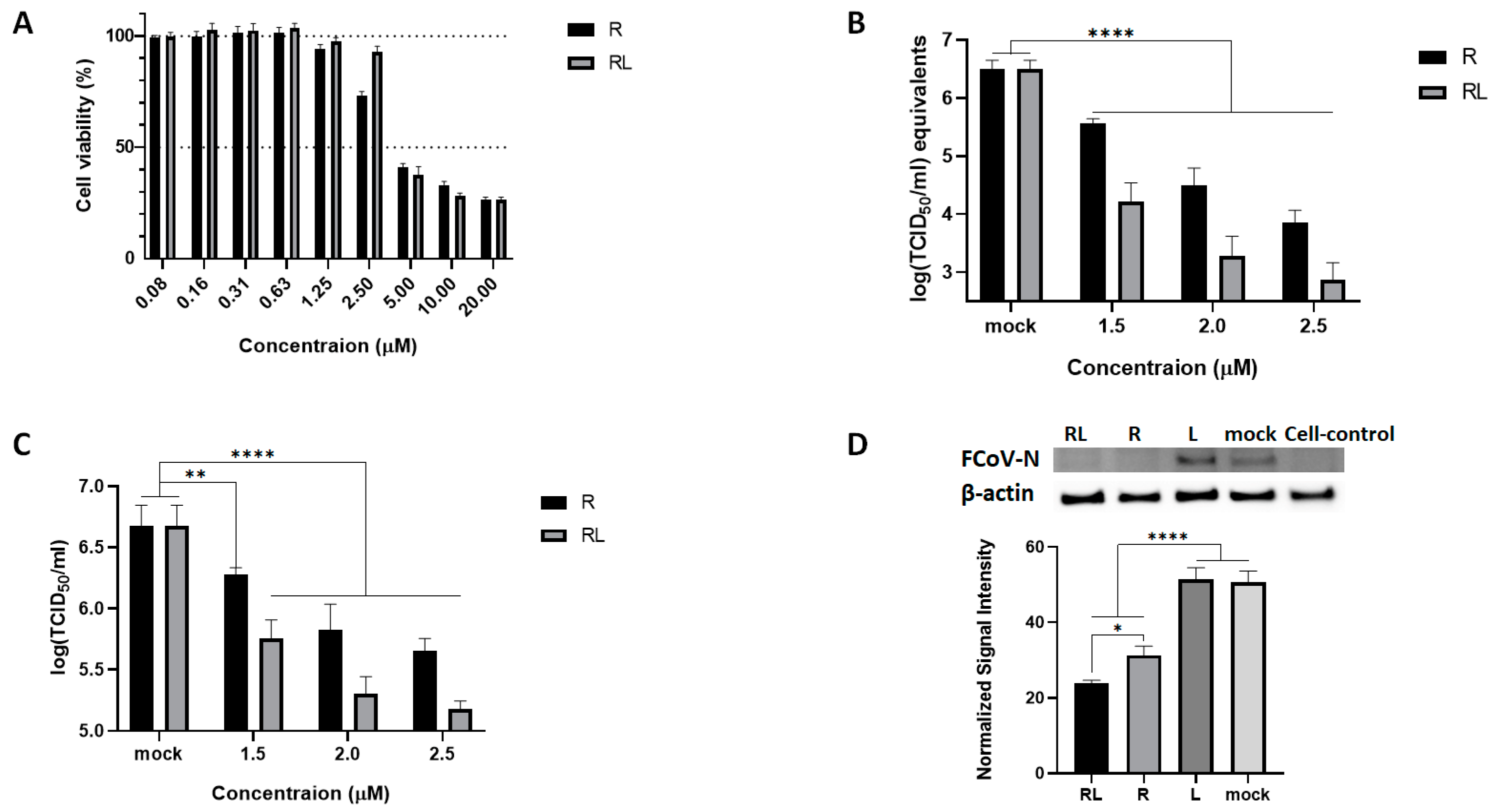{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells, Virus, and Antibodies
2.2. Preparation of R and RL
2.3. Characterization of RL
2.4. Cell Viability Assay
2.5. Dose-Dependent Inhibition Assay
2.6. RNA Extraction and RT-qPCR Assay
2.7. Western Blot
2.8. Time of Drug Addition Assay
2.9. Immunofluorescent Assay
2.10. Internalization Assay
2.11. Statistical Analysis
3. Results
3.1. RL Inhibits FCoV Replication
3.2. RL Decreased PKC Delta Phosphorylation Induced by FCoV Infection at an Early Stage of Infection
3.3. RL Inhibited FCoV Replication at Both Early and Late Stages of Infection
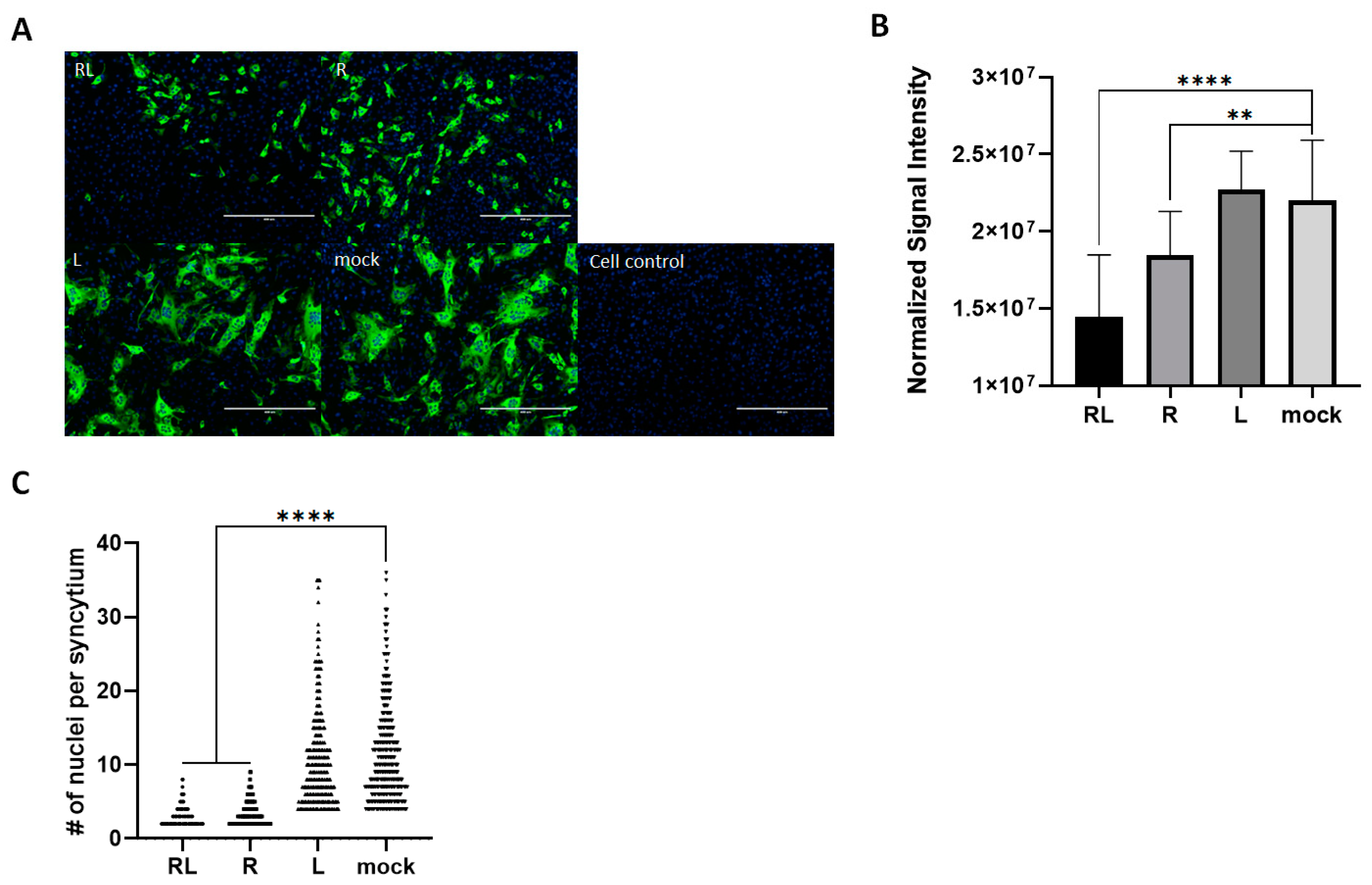
3.4. RL Decreased FCoV-Mediated Syncytia in Cell Cultures
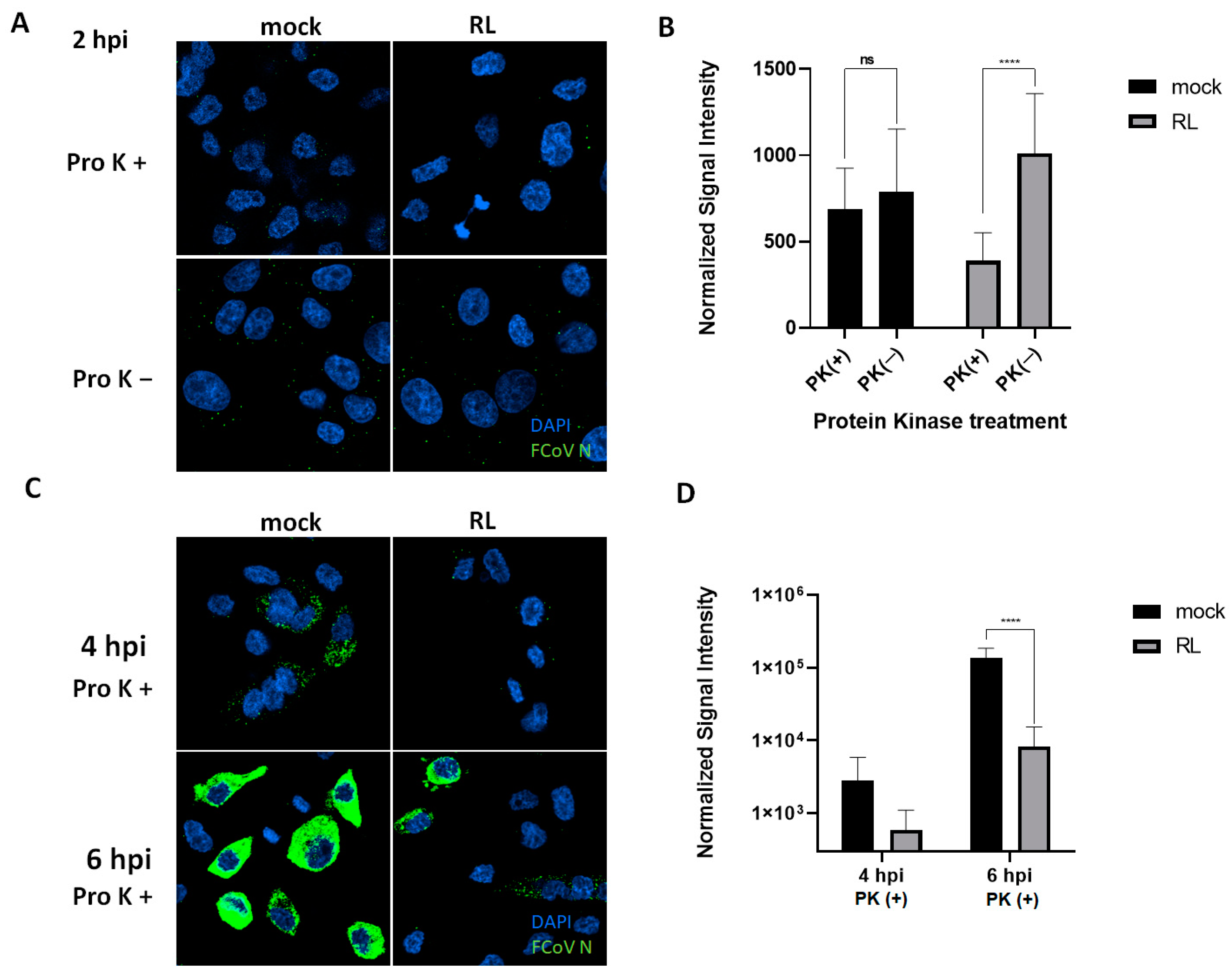
3.5. RL Inhibits FCoV Endocytosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, A.; Patil, M.; Kumar, P.; Bhatti, R.C.; Kaur, R.; Sharma, N.K.; Singh, A.N. Mallotus philippensis (Lam.) Müll. Arg.: A Review on Its Pharmacology and Phytochemistry. J. Herbmed. Pharmacol. 2020, 10, 31–50. [Google Scholar] [CrossRef]
- Rivière, C.; Nguyen Thi Hong, V.; Tran Hong, Q.; Chataigné, G.; Nguyen Hoai, N.; Dejaegher, B.; Tistaert, C.; Nguyen Thi Kim, T.; Vander Heyden, Y.; Chau Van, M.; et al. Mallotus Species from Vietnamese Mountainous Areas: Phytochemistry and Pharmacological Activities. Phytochem. Rev. 2010, 9, 217–253. [Google Scholar] [CrossRef]
- Gschwendt, M.; Muller, H.J.; Kielbassa, K.; Zang, R.; Kittstein, W.; Rincke, G.; Marks, F. Rottlerin, a Novel Protein Kinase Inhibitor. Biochem. Biophys. Res. Commun. 1994, 199, 93–98. [Google Scholar] [CrossRef]
- Kumar, V.; Kotrannavar, S.; Shivappa Angadi, S. A Review of Kampillaka (Mallotus Philippinensis Muell) from Ayurvedic Perspective. Ann. Ayurvedic Med. 2013, 2, 89–98. [Google Scholar]
- Manhas, D.; Gour, A.; Bhardwaj, N.; Sharma, D.K.; Sharma, K.; Vij, B.; Jain, S.K.; Singh, G.; Nandi, U. Pharmacokinetic Assessment of Rottlerin from Mallotus Philippensis Using a Highly Sensitive Liquid Chromatography–Tandem Mass Spectrometry-Based Bioanalytical Method. ACS Omega 2021, 6, 32637–32646. [Google Scholar] [CrossRef]
- Zhou, S.; Lin, Q.; Huang, C.; Luo, X.; Tian, X.; Liu, C.; Zhang, P. Rottlerin Plays an Antiviral Role at Early and Late Steps of Zika Virus Infection. Virol. Sin. 2022, 37, 685–694. [Google Scholar] [CrossRef]
- Ietta, F.; Maioli, E.; Daveri, E.; Gonzaga Oliveira, J.; da Silva, R.J.; Romagnoli, R.; Cresti, L.; Maria Avanzati, A.; Paulesu, L.; de Freitas Barbosa, B.; et al. Rottlerin-Mediated Inhibition of Toxoplasma gondii Growth in BeWo Trophoblast-like Cells. Sci. Rep. 2017, 7, 1279. [Google Scholar] [CrossRef]
- Pandey, S.; Chatterjee, A.; Jaiswal, S.; Kumar, S.; Ramachandran, R.; Srivastava, K.K. Protein Kinase C-δ Inhibitor, Rottlerin Inhibits Growth and Survival of Mycobacteria Exclusively through Shikimate Kinase. Biochem. Biophys. Res. Commun. 2016, 478, 721–726. [Google Scholar] [CrossRef]
- Shivshankar, P.; Lei, L.; Wang, J.; Zhong, G. Rottlerin Inhibits Chlamydial Intracellular Growth and Blocks Chlamydial Acquisition of Sphingolipids from Host Cells. Appl. Environ. Microbiol. 2008, 74, 1243–1249. [Google Scholar] [CrossRef]
- Contreras, X.; Mzoughi, O.; Gaston, F.; Peterlin, M.B.; Bahraoui, E. Protein Kinase C-Delta Regulates HIV-1 Replication at an Early Post-Entry Step in Macrophages. Retrovirology 2012, 9, 37. [Google Scholar] [CrossRef]
- Filone, C.M.; Hanna, S.L.; Caino, M.C.; Bambina, S.; Doms, R.W.; Cherry, S. Rift Valley Fever Virus Infection of Human Cells and Insect Hosts Is Promoted by Protein Kinase C Epsilon. PLoS ONE 2010, 5, e15483. [Google Scholar] [CrossRef]
- Kang, Y.L.; Oh, C.; Ahn, S.H.; Choi, J.C.; Choi, H.Y.; Lee, S.W.; Choi, I.S.; Song, C.S.; Lee, J.B.; Park, S.Y. Inhibition of Endocytosis of Porcine Reproductive and Respiratory Syndrome Virus by Rottlerin and Its Potential Prophylactic Administration in Piglets. Antiviral. Res. 2021, 195, 105191. [Google Scholar] [CrossRef] [PubMed]
- Mori, N.; Ishikawa, C.; Senba, M. Activation of PKC-δ in HTLV-1-Infected T Cells. Int. J. Oncol. 2015, 46, 1609–1618. [Google Scholar] [CrossRef]
- Zhao, H.; Guo, X.; Bi, Y.; Zhu, Y.; Feng, W. PKCδ Is Required for Porcine Reproductive and Respiratory Syndrome Virus Replication. Virology 2014, 468–470, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Soltoff, S.P. Rottlerin: An Inappropriate and Ineffective Inhibitor of PKCδ. Trends Pharmacol. Sci. 2007, 28, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Lama, Z.; Gaudin, Y.; Blondel, D.; Lagaudrière-Gesbert, C. Kinase Inhibitors Tyrphostin 9 and Rottlerin Block Early Steps of Rabies Virus Cycle. Antivir. Res. 2019, 168, 51–60. [Google Scholar] [CrossRef]
- Wallerstedt, E.; Smith, U.; Andersson, C.X. Protein Kinase C-δ Is Involved in the Inflammatory Effect of IL-6 in Mouse Adipose Cells. Diabetologia 2010, 53, 946–954. [Google Scholar] [CrossRef]
- Lee; Kim; Lee; Kwon PKCδ Mediates NF-ΚB Inflammatory Response and Downregulates SIRT1 Expression in Liver Fibrosis. Int. J. Mol. Sci. 2019, 20, 4607. [CrossRef]
- Parihar, S.P.; Ozturk, M.; Marakalala, M.J.; Loots, D.T.; Hurdayal, R.; Maasdorp, D.B.; Van Reenen, M.; Zak, D.E.; Darboe, F.; Penn-Nicholson, A.; et al. Protein Kinase C-Delta (PKCδ), a Marker of Inflammation and Tuberculosis Disease Progression in Humans, Is Important for Optimal Macrophage Killing Effector Functions and Survival in Mice. Mucosal Immunol. 2018, 11, 496–511. [Google Scholar] [CrossRef]
- Gao, A.; Hu, X.; Saeed, M.; Chen, B.; Li, Y.; Yu, H. Overview of Recent Advances in Liposomal Nanoparticle-Based Cancer Immunotherapy. Acta Pharmacol. Sin. 2019, 40, 1129–1137. [Google Scholar] [CrossRef]
- Gheibi Hayat, S.M.; Darroudi, M. Nanovaccine: A Novel Approach in Immunization. J. Cell Physiol. 2019, 234, 12530–12536. [Google Scholar] [CrossRef]
- Hu, C.-M.J.; Chang, W.-S.; Fang, Z.-S.; Chen, Y.-T.; Wang, W.-L.; Tsai, H.-H.; Chueh, L.-L.; Takano, T.; Hohdatsu, T.; Chen, H.-W. Nanoparticulate Vacuolar ATPase Blocker Exhibits Potent Host-Targeted Antiviral Activity against Feline Coronavirus. Sci. Rep. 2017, 7, 13043. [Google Scholar] [CrossRef] [PubMed]
- Malam, Y.; Loizidou, M.; Seifalian, A.M. Liposomes and Nanoparticles: Nanosized Vehicles for Drug Delivery in Cancer. Trends Pharmacol. Sci. 2009, 30, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhuang, S.; Qi, X.-R. Comparative Study of the in Vitro and in Vivo Characteristics of Cationic and Neutral Liposomes. Int. J. Nanomed. 2011, 6, 3087–3098. [Google Scholar] [CrossRef]
- Sogut, O.; Aydemir Sezer, U.; Sezer, S. Liposomal Delivery Systems for Herbal Extracts. J. Drug Deliv. Sci. Technol. 2021, 61, 102147. [Google Scholar] [CrossRef]
- Olusanya, T.; Haj Ahmad, R.; Ibegbu, D.; Smith, J.; Elkordy, A. Liposomal Drug Delivery Systems and Anticancer Drugs. Molecules 2018, 23, 907. [Google Scholar] [CrossRef]
- Chang, Y.; Meng, F.-C.; Wang, R.; Wang, C.M.; Lu, X.-Y.; Zhang, Q.-W. Chemistry, Bioactivity, and the Structure-Activity Relationship of Cephalotaxine-Type Alkaloids From Cephalotaxus sp. Stud. Nat. Prod. Chem. 2017, 53, 339–373. [Google Scholar] [CrossRef]
- De Rosa, G.; Campani, V.; Marchese, D.; Pitaro, M.T.; Pitaro, M.; Grieco, P. Development of a Liposome-Based Formulation for Vitamin K1 Nebulization on the Skin. Int. J. Nanomed. 2014, 9, 1823. [Google Scholar] [CrossRef]
- Zou, L.; Liu, W.; Liu, W.; Liang, R.; Li, T.; Liu, C.; Cao, Y.; Niu, J.; Liu, Z. Characterization and Bioavailability of Tea Polyphenol Nanoliposome Prepared by Combining an Ethanol Injection Method with Dynamic High-Pressure Microfluidization. J. Agric. Food Chem. 2014, 62, 934–941. [Google Scholar] [CrossRef]
- Kim, Y.; Shivanna, V.; Narayanan, S.; Prior, A.M.; Weerasekara, S.; Hua, D.H.; Kankanamalage, A.C.G.; Groutas, W.C.; Chang, K.-O. Broad-Spectrum Inhibitors against 3C-Like Proteases of Feline Coronaviruses and Feline Caliciviruses. J. Virol. 2015, 89, 4942–4950. [Google Scholar] [CrossRef]
- Rottier, P.J.M.; Nakamura, K.; Schellen, P.; Volders, H.; Haijema, B.J. Acquisition of Macrophage Tropism during the Pathogenesis of Feline Infectious Peritonitis Is Determined by Mutations in the Feline Coronavirus Spike Protein. J. Virol. 2005, 79, 14122–14130. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, N.C. An Update on Feline Infectious Peritonitis: Diagnostics and Therapeutics. Vet. J. 2014, 201, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, N.C.; Eckstrand, C.; Liu, H.; Leutenegger, C.; Murphy, B. Levels of Feline Infectious Peritonitis Virus in Blood, Effusions, and Various Tissues and the Role of Lymphopenia in Disease Outcome Following Experimental Infection. Vet. Microbiol. 2015, 175, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Liu, H.; Galasiti Kankanamalage, A.C.; Weerasekara, S.; Hua, D.H.; Groutas, W.C.; Chang, K.O.; Pedersen, N.C. Reversal of the Progression of Fatal Coronavirus Infection in Cats by a Broad-Spectrum Coronavirus Protease Inhibitor. PLoS Pathog. 2016, 12, e1005650. [Google Scholar] [CrossRef]
- Tsai, H.-Y.; Chueh, L.-L.; Lin, C.-N.; Su, B.-L. Clinicopathological Findings and Disease Staging of Feline Infectious Peritonitis: 51 Cases from 2003 to 2009 in Taiwan. J. Feline Med. Surg. 2011, 13, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Legendre, A.M.; Bartges, J.W. Effect of Polyprenyl Immunostimulant on the Survival Times of Three Cats with the Dry Form of Feline Infectious Peritonitis. J. Feline Med. Surg. 2009, 11, 624–626. [Google Scholar] [CrossRef]
- Addie, D.D.; Paltrinieri, S.; Pedersen, N.C. Recommendations from Workshops of the Second International Feline Coronavirus/Feline Infectious Peritonitis Symposium. J. Feline Med. Surg. 2004, 6, 125–130. [Google Scholar] [CrossRef]
- Addie, D.; Belák, S.; Boucraut-Baralon, C.; Egberink, H.; Frymus, T.; Gruffydd-Jones, T.; Hartmann, K.; Hosie, M.J.; Lloret, A.; Lutz, H.; et al. Feline Infectious Peritonitis: ABCD Guidelines on Prevention and Management. J. Feline Med. Surg. 2009, 11, 594–604. [Google Scholar] [CrossRef]
- Takano, T.; Katoh, Y.; Doki, T.; Hohdatsu, T. Effect of Chloroquine on Feline Infectious Peritonitis Virus Infection in Vitro and in Vivo. Antivir. Res. 2013, 99, 100–107. [Google Scholar] [CrossRef]
- Pedersen, N.C.; Perron, M.; Bannasch, M.; Montgomery, E.; Murakami, E.; Liepnieks, M.; Liu, H. Efficacy and Safety of the Nucleoside Analog GS-441524 for Treatment of Cats with Naturally Occurring Feline Infectious Peritonitis. J. Feline Med. Surg. 2019, 21, 271–281. [Google Scholar] [CrossRef]
- Weiss, R.C.; Oostrom-Ram, T. Inhibitory Effects of Ribavirin Alone or Combined with Human Alpha Interferon on Feline Infectious Peritonitis Virus Replication in Vitro. Vet. Microbiol. 1989, 20, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Doki, T.; Tarusawa, T.; Hohdatsu, T.; Takano, T. In Vivo Antiviral Effects of U18666A Against Type I Feline Infectious Peritonitis Virus. Pathogens 2020, 9, 67. [Google Scholar] [CrossRef] [PubMed]
- Takano, T.; Akiyama, M.; Doki, T.; Hohdatsu, T. Antiviral Activity of Itraconazole against Type I Feline Coronavirus Infection. Vet. Res. 2019, 50, 5. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.; Jacque, N.; Novicoff, W.; Li, E.; Negash, R.; Evans, S.J.M. Unlicensed Molnupiravir Is an Effective Rescue Treatment Following Failure of Unlicensed GS-441524-like Therapy for Cats with Suspected Feline Infectious Peritonitis. Pathogens 2022, 11, 1209. [Google Scholar] [CrossRef]
- Yu, J.; Kimble, B.; Norris, J.M.; Govendir, M. Pharmacokinetic Profile of Oral Administration of Mefloquine to Clinically Normal Cats: A Preliminary In-Vivo Study of a Potential Treatment for Feline Infectious Peritonitis (FIP). Animals 2020, 10, 1000. [Google Scholar] [CrossRef]
- Pedersen, N.C.; Evermann, J.F.; McKeirnan, A.J.; Ott, R.L. Pathogenicity Studies of Feline Coronavirus Isolates 79-1146 and 79-1683. Am. J. Vet. Res. 1984, 45, 2580–2585. [Google Scholar]
- Stranieri, A.; Giordano, A.; Paltrinieri, S.; Giudice, C.; Cannito, V.; Lauzi, S. Comparison of the Performance of Laboratory Tests in the Diagnosis of Feline Infectious Peritonitis. J. Vet. Diagn. Investig. 2018, 30, 459–463. [Google Scholar] [CrossRef]
- Wang, R.; Yang, L.; Zhang, Y.; Li, J.; Xu, L.; Xiao, Y.; Zhang, Q.; Bai, L.; Zhao, S.; Liu, E.; et al. Porcine Reproductive and Respiratory Syndrome Virus Induces HMGB1 Secretion via Activating PKC-Delta to Trigger Inflammatory Response. Virology 2018, 518, 172–183. [Google Scholar] [CrossRef]
- Mahapatra, A.; Kumar, P.; Bhansare, J.; Surapaneni, S.M.; Sen, A.; Pradhan, B. Development of Dye-Sensitized Solar Cell Using M. Philippensis (Kamala Tree) Fruit Extract: A Combined Experimental and Theoretical Study. Int. J. Energy Res. 2021, 45, 21509–21515. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Sisk, J.M.; Frieman, M.B.; Machamer, C.E. Coronavirus S Protein-Induced Fusion Is Blocked Prior to Hemifusion by Abl Kinase Inhibitors. J. Gen. Virol. 2018, 99, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Abdelnabi, R.; Amrun, S.N.; Ng, L.F.P.; Leyssen, P.; Neyts, J.; Delang, L. Protein Kinases C as Potential Host Targets for the Inhibition of Chikungunya Virus Replication. Antivir. Res. 2017, 139, 79–87. [Google Scholar] [CrossRef]
- Ojha, D.; Winkler, C.W.; Leung, J.M.; Woods, T.A.; Chen, C.Z.; Nair, V.; Taylor, K.; Yeh, C.D.; Tawa, G.J.; Larson, C.L.; et al. Rottlerin Inhibits La Crosse Virus-Induced Encephalitis in Mice and Blocks Release of Replicating Virus from the Golgi Body in Neurons. Nat. Microbiol. 2021, 6, 1398–1409. [Google Scholar] [CrossRef]
- Chen, F.; Shi, Q.; Pei, F.; Vogt, A.; Porritt, R.A.; Garcia, G.; Gomez, A.C.; Cheng, M.H.; Schurdak, M.E.; Liu, B.; et al. A Systems-level Study Reveals Host-targeted Repurposable Drugs against SARS-CoV-2 Infection. Mol. Syst. Biol. 2021, 17, e10239. [Google Scholar] [CrossRef]
- Huang, C.; Feng, F.; Shi, Y.; Li, W.; Wang, Z.; Zhu, Y.; Yuan, S.; Hu, D.; Dai, J.; Jiang, Q.; et al. Protein Kinase C Inhibitors Reduce SARS-CoV-2 Replication in Cultured Cells. Microbiol. Spectr. 2022, 10, e01056-22. [Google Scholar] [CrossRef] [PubMed]
- Bauherr, S.; Larsberg, F.; Petrich, A.; Sperber, H.S.; Klose-Grzelka, V.; Luckner, M.; Azab, W.; Schade, M.; Höfer, C.T.; Lehmann, M.J.; et al. Macropinocytosis and Clathrin-Dependent Endocytosis Play Pivotal Roles for the Infectious Entry of Puumala Virus. J. Virol. 2020, 94, e00184-20. [Google Scholar] [CrossRef] [PubMed]
- Burkard, C.; Verheije, M.H.; Haagmans, B.L.; van Kuppeveld, F.J.; Rottier, P.J.M.; Bosch, B.-J.; de Haan, C.A.M. ATP1A1-Mediated Src Signaling Inhibits Coronavirus Entry into Host Cells. J. Virol. 2015, 89, 4434–4448. [Google Scholar] [CrossRef] [PubMed]
- Maioli, E.; Torricelli, C.; Valacchi, G. Rottlerin and Cancer: Novel Evidence and Mechanisms. Sci. World J. 2012, 2012, 350826. [Google Scholar] [CrossRef]
- Xia, S.; Yan, L.; Xu, W.; Agrawal, A.S.; Algaissi, A.; Tseng, C.-T.K.; Wang, Q.; Du, L.; Tan, W.; Wilson, I.A.; et al. A Pan-Coronavirus Fusion Inhibitor Targeting the HR1 Domain of Human Coronavirus Spike. Sci. Adv. 2019, 5, eaav4580. [Google Scholar] [CrossRef]
- Yoo, D.; Parker, M.D.; Babiuk, L.A. The S2 Subunit of the Spike Glycoprotein of Bovine Coronavirus Mediates Membrane Fusion in Insect Cells. Virology 1991, 180, 395–399. [Google Scholar] [CrossRef]
- Rahman, H.S.; Othman, H.H.; Hammadi, N.I.; Yeap, S.K.; Amin, K.M.; Abdul Samad, N.; Alitheen, N.B. Novel Drug Delivery Systems for Loading of Natural Plant Extracts and Their Biomedical Applications. Int. J. Nanomed. 2020, 15, 2439–2483. [Google Scholar] [CrossRef]
- Kelly, C.; Jefferies, C.; Cryan, S.-A. Targeted Liposomal Drug Delivery to Monocytes and Macrophages. J. Drug Deliv. 2011, 2011, 727241. [Google Scholar] [CrossRef]
- Shah, K.K.; Pritt, B.S.; Alexander, M.P. Histopathologic Review of Granulomatous Inflammation. J. Clin. Tuberc. Other Mycobact. Dis. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Epstein, W.L.; Fukuyama, K. Mechanisms of Granulomatous Inflammation. Immunol. Ser. 1989, 46, 687–721. [Google Scholar]
- Delaplace, M.; Huet, H.; Gambino, A.; Le Poder, S. Feline Coronavirus Antivirals: A Review. Pathogens 2021, 10, 1150. [Google Scholar] [CrossRef] [PubMed]
- Barlough, J.; Shacklett, B. Antiviral Studies of Feline Infectious Peritonitis Virus in Vitro. Vet. Rec. 1994, 135, 177–179. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, L.E.; Lin, C.N.; Su, B.L.; Jan, T.R.; Chen, C.M.; Wang, C.H.; Lin, D.S.; Lin, C.T.; Chueh, L.L. Synergistic Antiviral Effect of Galanthus Nivalis Agglutinin and Nelfinavir against Feline Coronavirus. Antivir. Res. 2010, 88, 25–30. [Google Scholar] [CrossRef]
- Takano, T.; Satoh, K.; Doki, T.; Tanabe, T.; Hohdatsu, T. Antiviral Effects of Hydroxychloroquine and Type I Interferon on In Vitro Fatal Feline Coronavirus Infection. Viruses 2020, 12, 576. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.-C.; Jung, S.-W.; Choi, I.-Y.; Kang, Y.-L.; Lee, D.-H.; Lee, S.-W.; Park, S.-Y.; Song, C.-S.; Choi, I.-S.; Lee, J.-B.; et al. Rottlerin-Liposome Inhibits the Endocytosis of Feline Coronavirus Infection. Vet. Sci. 2023, 10, 380. https://doi.org/10.3390/vetsci10060380
Choi J-C, Jung S-W, Choi I-Y, Kang Y-L, Lee D-H, Lee S-W, Park S-Y, Song C-S, Choi I-S, Lee J-B, et al. Rottlerin-Liposome Inhibits the Endocytosis of Feline Coronavirus Infection. Veterinary Sciences. 2023; 10(6):380. https://doi.org/10.3390/vetsci10060380
Chicago/Turabian StyleChoi, Jong-Chul, Sung-Won Jung, In-Yeong Choi, Yeong-Lim Kang, Dong-Hun Lee, Sang-Won Lee, Seung-Yong Park, Chang-Seon Song, In-Soo Choi, Joong-Bok Lee, and et al. 2023. "Rottlerin-Liposome Inhibits the Endocytosis of Feline Coronavirus Infection" Veterinary Sciences 10, no. 6: 380. https://doi.org/10.3390/vetsci10060380