
Retrospective Investigation and Genetic Variation Analysis of Chicken Infectious Anemia in Shandong Province, 2020–2022
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Treatment
2.2. Viral DNA Detection by PCR
2.3. VP1 Gene Sequencing and Analysis
3. Results
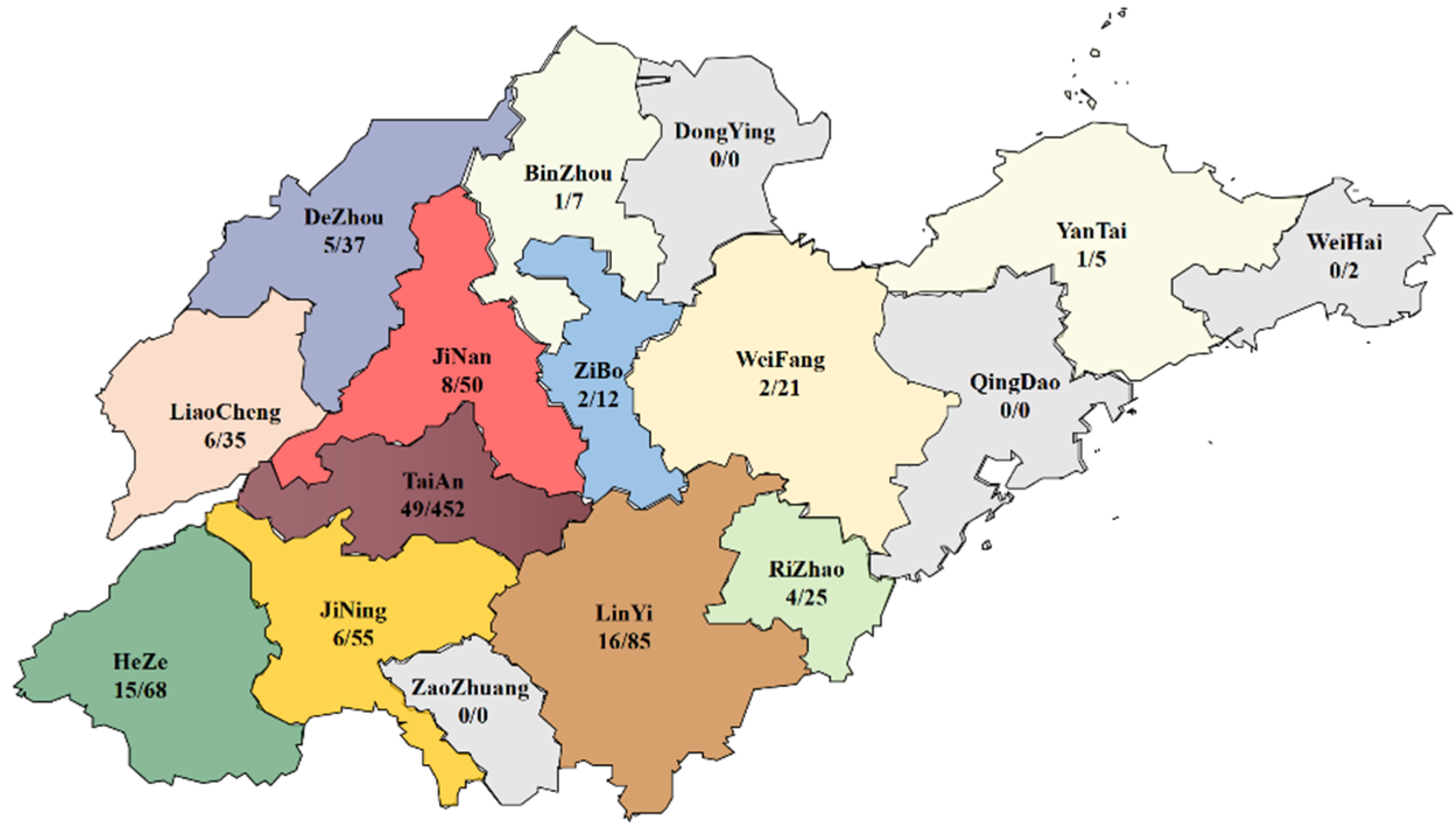
3.1. PCR Results and Geographical Distribution of CAV-Positive Samples
3.2. Mixed Infections of Samples
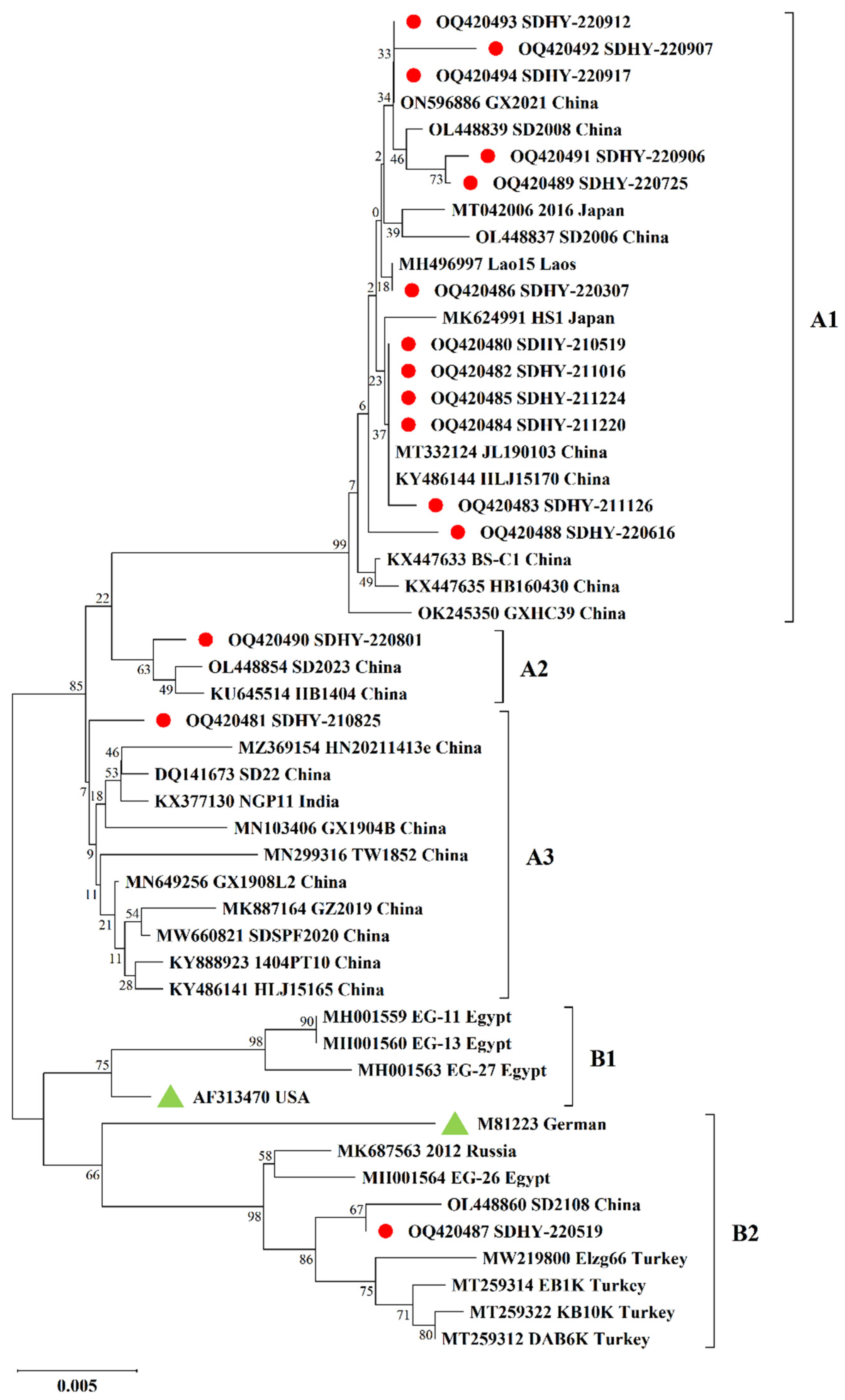
3.3. Molecular Characterization of CAV VP1 Gene
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gimeno, I.M.; Schat, K.A. Virus-Induced Immunosuppression in Chickens. Avian Dis. 2018, 62, 272–285. [Google Scholar] [CrossRef] [PubMed]
- Haridy, M.; Goryo, M.; Sasaki, J.; Okada, K. Pathological and immunohistochemical study of chickens with co-infection of Marek’s disease virus and chicken anaemia virus. Avian Pathol. 2009, 38, 469–483. [Google Scholar] [CrossRef] [PubMed]
- Adair, B.M. Immunopathogenesis of chicken anemia virus infection. Dev. Comp. Immunol. 2000, 24, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hu, Y.; Cui, S.; Fu, J.; Wang, Y.; Cui, Z.; Fang, L.; Chang, S.; Zhao, P. Molecular characterization of chicken infectious anemia virus from contaminated live-virus vaccines. Poult. Sci. 2017, 96, 1045–1051. [Google Scholar] [CrossRef] [PubMed]
- Kraberger, S.; Opriessnig, T.; Celer, V.; Maggi, F.; Okamoto, H.; Blomström, A.L.; Cadar, D.; Harrach, B.; Biagini, P.; Varsani, A. Taxonomic updates for the genus Gyrovirus (family Anelloviridae): Recognition of several new members and establishment of species de-marcation criteria. Arch. Virol. 2021, 166, 2937–2942. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Song, X.; Gao, H.; Wang, X.; Hu, Y.; Gao, Y.; Qi, X.; Qin, L.; Lin, H.; Gao, L.; et al. C-terminal region of apoptin affects chicken anemia virus replication and virulence. Virol. J. 2017, 14, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lien, Y.Y.; Huang, C.H.; Sun, F.C.; Sheu, S.C.; Lu, T.C.; Lee, M.S.; Hsueh, S.C.; Chen, H.J.; Lee, M.S. Development and characterization of a potential diagnostic monoclonal antibody against capsid protein VP1 of the chicken anemia virus. J. Vet. Sci. 2012, 13, 73–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Fang, L.; Cui, S.; Fu, J.; Li, X.; Zhang, H.; Cui, Z.; Chang, S.; Shi, W.; Zhao, P. Genomic Characterization of Recent Chicken Anemia Virus Isolates in China. Front. Microbiol. 2017, 8, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Liu, Y.; Wu, B.; Sun, B.; Chen, F.; Ji, J.; Ma, J.; Xie, Q. Phylogenetic and molecular characterization of chicken anemia virus in southern China from 2011 to 2012. Sci. Rep. 2013, 3, 3519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhang, K.; Pei, Y.; Xue, J.; Ruan, S.; Zhang, G. Development and Application of an MRT-qPCR Assay for Detecting Coinfection of Six Vertically Transmitted or Immunosuppressive Avian Viruses. Front. Microbiol. 2020, 11, 1581. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, Y.; Yin, M.; Zhao, P.; Guo, L.; Wang, Y. Genomic Characterization of Chicken Anemia Virus in Broilers in Shandong Province, China, 2020–2021. Front. Vet. Sci. 2022, 9, 816860. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Zhao, Y.; Zhao, J.; Geng, N.; Meng, F.; Wang, S.; Li, J.; Zhong, Z.; Zhu, L.; Liu, S.; et al. The diagnosis and molecular epidemiology investigation of avian hepatitis E in Shandong province, China. BMC Vet. Res. 2022, 18, 56. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Feng, J.; Jin, J.X.; Zhu, X.J.; Sun, A.J.; Liu, H.Y.; Wang, J.J.; Wang, R.; Yang, X.; Chen, L.; et al. Molecular Epidemiology and Pathogenic Characterization of Novel Chicken Infectious Anemia Viruses in Henan Province of China. Front. Vet. Sci. 2022, 9, 871826. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Deng, X.; Xie, Z.; Zhang, Y.; Xie, Z.; Xie, L.; Luo, S.; Fan, Q.; Zeng, T.; Huang, J.; et al. Molecular characterization of chicken anemia virus in Guangxi Province, southern China, from 2018 to 2020. J. Vet. Sci. 2022, 23, e63. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Dong, G.; Zhang, Y.; Tian, S.; Cui, Z.; Chang, S.; Zhao, P. Co-infection of fowl adenovirus with different immunosuppressive viruses in a chicken flock. Poult. Sci. 2018, 97, 1699–1705. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Wang, Z.; Lei, X.; Lu, J.; Yan, Z.; Qin, J.; Chen, F.; Xie, Q.; Lin, W. Epidemiology, molecular characterization, and recombination analysis of chicken anemia virus in Guangdong province, China. Arch. Virol. 2020, 165, 1409–1417. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Su, Q.; Li, Y.; Wang, J.; Zhang, Y.; Chang, S.; Wang, Y.; Zhao, P. Genomic Characteristics of a Chicken Infectious Anemia Virus in Contaminated Attenuated Vaccine. Front. Vet. Sci. 2022, 9, 925935. [Google Scholar] [CrossRef]
- Erfan, A.M.; Selim, A.A.; Naguib, M.M. Characterization of full genome sequences of chicken anemia viruses circulating in Egypt reveals distinct genetic diversity and evidence of recombination. Virus Res. 2018, 251, 78–85. [Google Scholar] [CrossRef]
- Renshaw, R.W.; Soiné, C.; Weinkle, T.; O’Connell, P.H.; Ohashi, K.; Watson, S.; Lucio, B.; Harrington, S.; Schat, K.A. A hypervariable region in VP1 of chicken infectious anemia virus mediates rate of spread and cell tropism in tissue culture. J. Virol. 1996, 70, 8872–8878. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Genes | Primer Sequence (5′-3′) | Tm (°C) | Sizes (bp) |
|---|---|---|---|
| CAV-F | CAGAATTCCCACCTCAAGCGACTTCGAC | 55 | 582 |
| CAV-R | ATGTCGACGGGGCTGAAGGAT | ||
| FAdV-F | AATTTCGACCCCATGACGCGCCAGG | 56 | 508 |
| FAdV-R | TGGCGAAAGGCGTACGGAAGTAAGC | ||
| REV-F | CATACGAGCCAATGGTT | 54 | 300 |
| REV-R | AATGTTGTAGCGAAGTACT | ||
| ALV-J-F | GGATGAGGTGACTAAGA | 56 | 512 |
| ALV-J-R | CGAACCAAAGGTAACACACG | ||
| MDV-F | TCATCAGGGTCTCCCGTCACCT | 58 | 1005 |
| MDV-R | AGAGATGTCTCAGGAGCCAGAG | ||
| HEV-F1 | TCGCCT(C)GGTAAT(C)ACA(T)AATGC | 60 | 278 |
| HEV-R1 | GCGTTC(G)CCG(C)ACAGGT(C)CGGCC | ||
| HEV-F2 | ACA(T)AATGCT(C)AGGGTCACCCG | 56 | 242 |
| HEV-R2 | ATGTACTGA(G)CCA(G)CTG(C)GCCGC |
| Group | Strain | VP1 Amino Acid Site | Genotype | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 75 | 97 | 125 | 139 | 141 | 144 | 157 | 287 | 290 | 294 | |||
| 1 | on596886 | V | L | L | K | Q | E | M | N | P | H | A1 |
| 2 | KU645514 | · | M | · | · | · | · | V | · | · | · | A2 |
| 3 | DQ141673 | · | M | · | · | · | · | V | · | · | · | A3 |
| 4 | AF311470 | · | M | I | · | · | · | V | S | A | Q | B1 |
| 5 | MH001564 | I | · | I | Q | · | Q | V | A | · | Q | B2 |
| 6 | SDHY-210519 | · | · | · | · | · | · | · | T | · | · | A1 |
| 7 | SDHY-211016 | · | · | · | · | · | · | V | S | A | Q | A1 |
| 8 | SDHY-211126 | · | · | · | · | · | · | · | T | · | · | A1 |
| 9 | SDHY-211220 | · | · | · | · | · | · | · | T | · | · | A1 |
| 10 | SDHY-211224 | · | · | · | · | · | · | · | T | · | · | A1 |
| 11 | SDHY-220307 | · | · | · | · | · | · | · | S | · | · | A1 |
| 12 | SDHY-220616 | · | · | · | · | · | · | · | T | · | Q | A1 |
| 13 | SDHY-220725 | · | · | · | · | · | · | · | · | · | · | A1 |
| 14 | SDHY-220906 | · | · | · | · | · | · | · | · | · | Q | A1 |
| 15 | SDHY-220907 | · | · | · | · | · | · | · | · | · | · | A1 |
| 16 | SDHY-220912 | · | · | · | · | · | · | · | · | · | · | A1 |
| 17 | SDHY-220917 | · | · | · | · | · | · | · | · | · | · | A1 |
| 18 | SDHY-220801 | · | M | · | · | · | · | · | S | A | Q | A2 |
| 19 | SDHY-210825 | · | · | · | · | · | · | V | S | A | Q | A3 |
| 20 | SDHY-220519 | I | · | I | Q | · | Q | V | A | A | Q | B2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Lou, Y.; Li, P.; Wang, T.; Lv, Z.; Guo, Z.; Geng, N.; Meng, F.; Liu, S.; Li, N. Retrospective Investigation and Genetic Variation Analysis of Chicken Infectious Anemia in Shandong Province, 2020–2022. Vet. Sci. 2023, 10, 263. https://doi.org/10.3390/vetsci10040263
Li J, Lou Y, Li P, Wang T, Lv Z, Guo Z, Geng N, Meng F, Liu S, Li N. Retrospective Investigation and Genetic Variation Analysis of Chicken Infectious Anemia in Shandong Province, 2020–2022. Veterinary Sciences. 2023; 10(4):263. https://doi.org/10.3390/vetsci10040263
Chicago/Turabian StyleLi, Jing, Yufei Lou, Peixun Li, Tailong Wang, Zehao Lv, Zhiyun Guo, Ningwei Geng, Fanliang Meng, Sidang Liu, and Ning Li. 2023. "Retrospective Investigation and Genetic Variation Analysis of Chicken Infectious Anemia in Shandong Province, 2020–2022" Veterinary Sciences 10, no. 4: 263. https://doi.org/10.3390/vetsci10040263