
Influence of BoLA-DRB3 Polymorphism and Bovine Leukemia Virus (BLV) Infection on Dairy Cattle Productivity

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Blood Sample Collection and Genomic DNA Isolation
2.2. Diagnosis of BLV Infection
2.3. BoLA-DRB3 Allele Typing
2.4. Assessment of Dairy Cattle Productivity
2.5. Staristical Analysis
3. Results
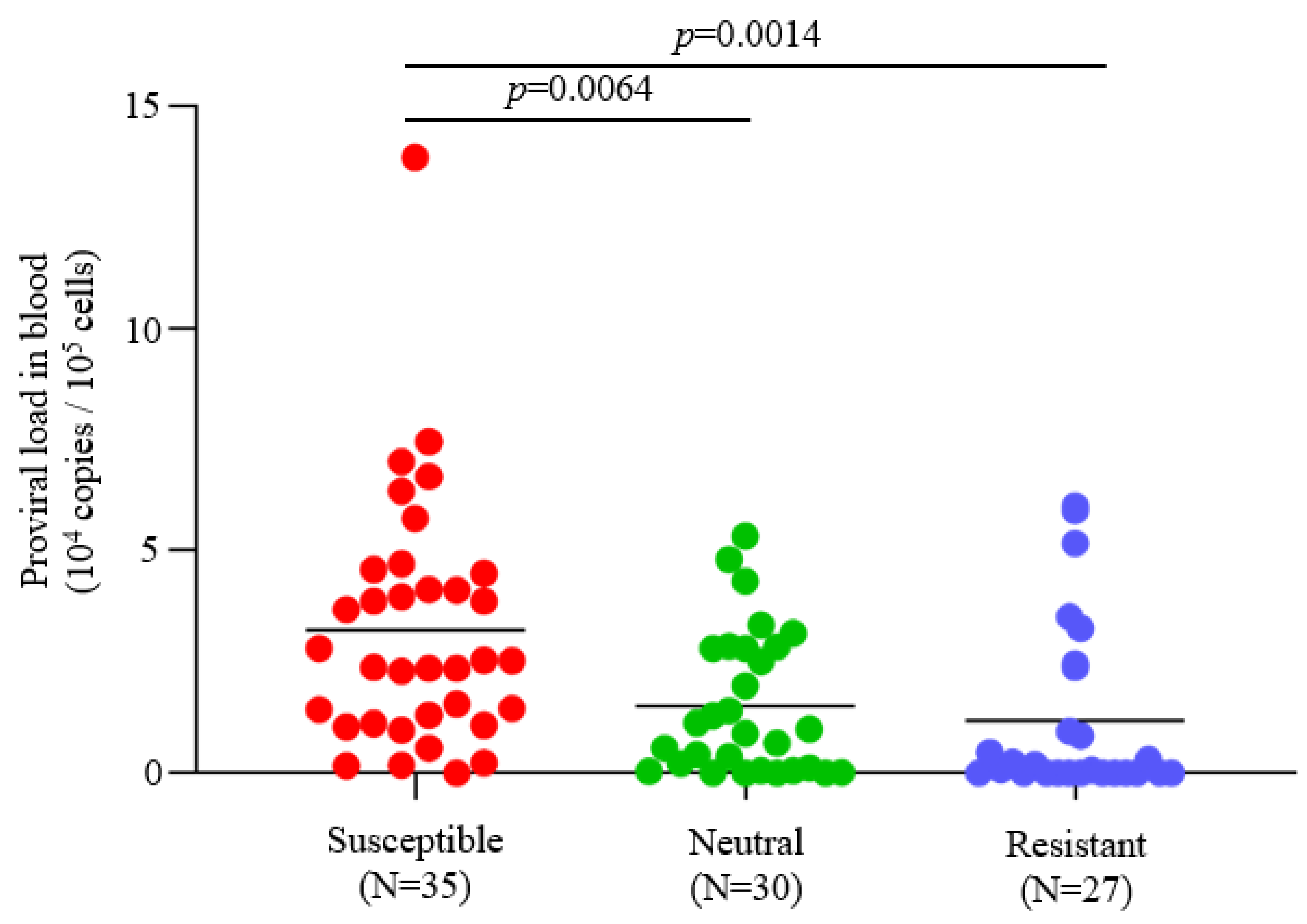
3.1. Determination of the BoLA-DRB3 Alleles and BLV Infection
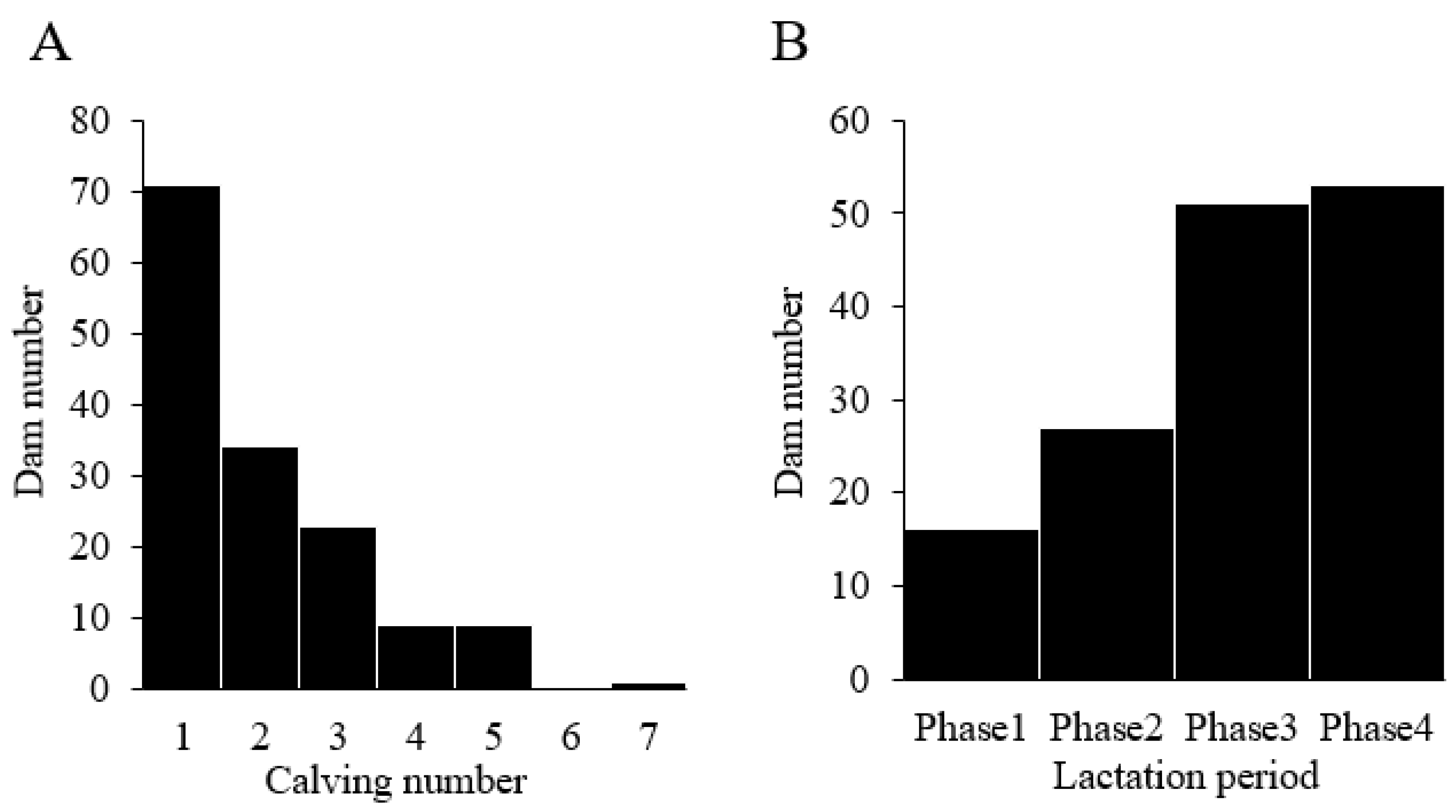
3.2. Distribution of Dams Based on Calving Number and Lactation Period
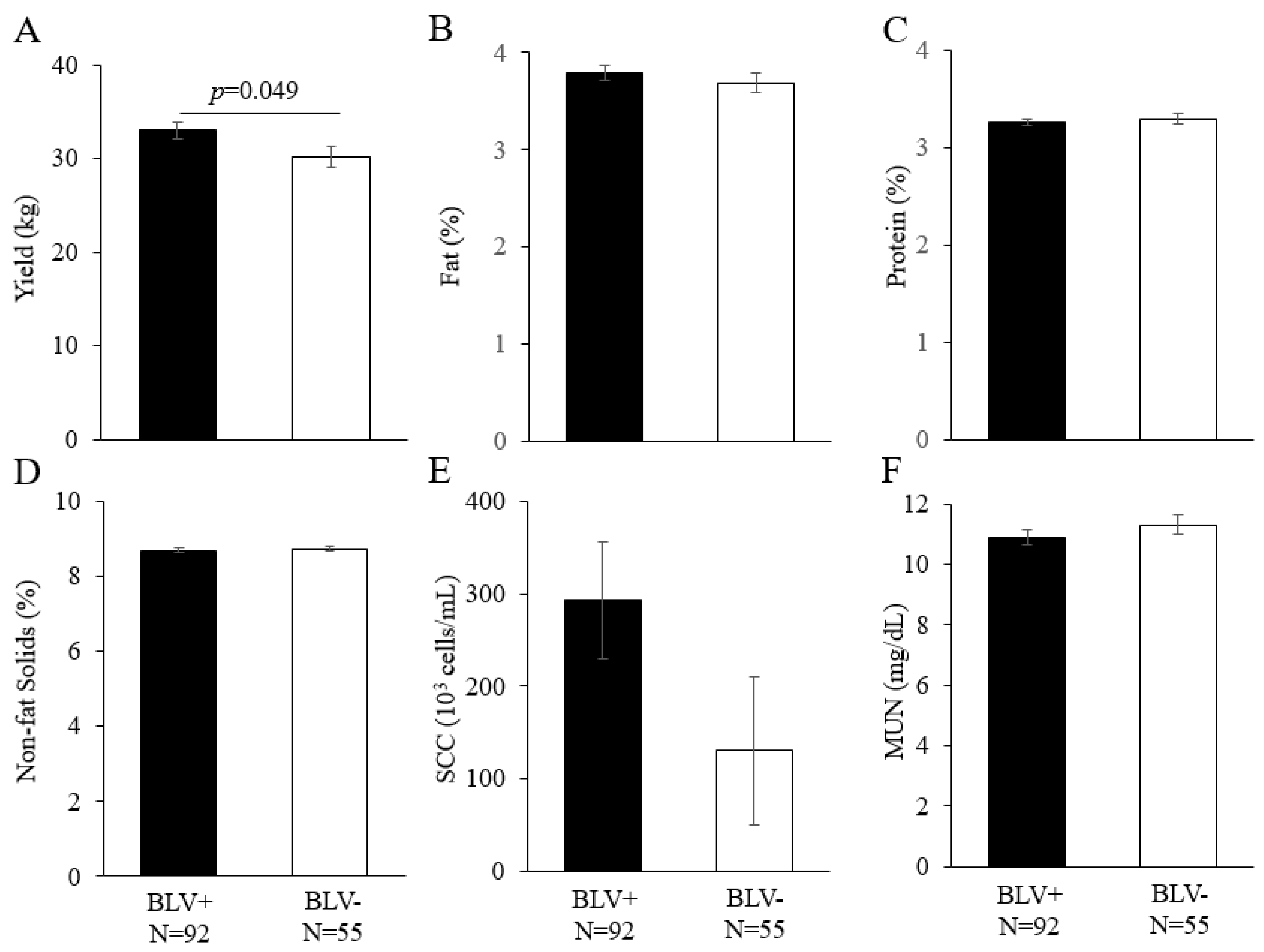
3.3. Effect of BLV Infection on Dairy Cattle Productivity
3.4. BoLA-DRB3 Polymorphism Does Not Affect Dairy Cattle Productivity
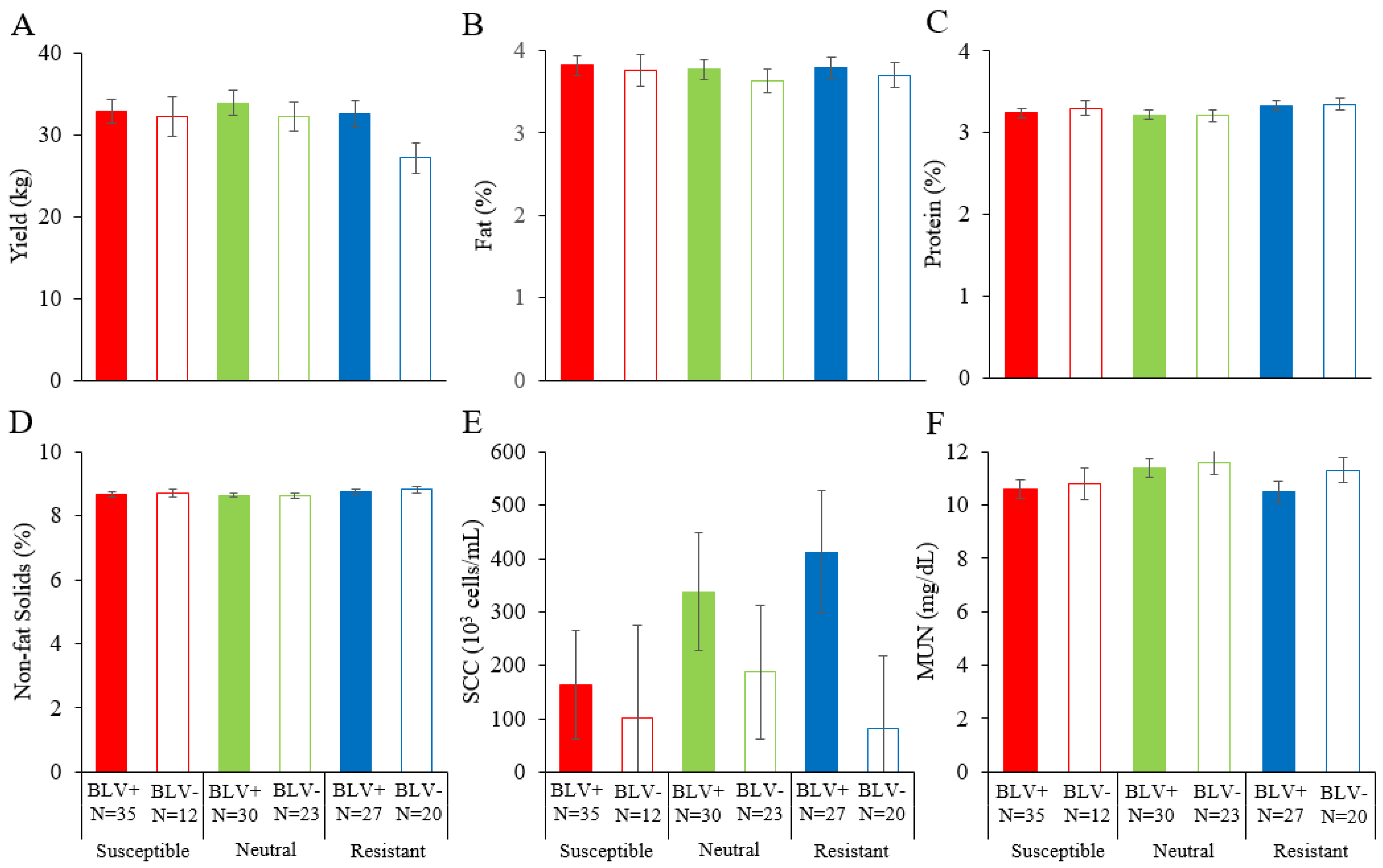
3.5. Combined Effect of BoLA-DRB Polymorphism and BLV Infection on Dairy Cattle Productivity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marawan, M.A.; Alouffi, A.; El Tokhy, S.; Badawy, S.; Shirani, I.; Dawood, A.; Guo, A.; Almutairi, M.M.; Alshammari, F.A.; Selim, A. Bovine Leukaemia Virus: Current Epidemiological Circumstance and Future Prospective. Viruses 2021, 13, 2167. [Google Scholar] [CrossRef]
- OIE. World Animal Health Infromation Database-Version: 1.4; World Organisation for Animal Health: Paris, France, 2009; Available online: https://scholar.google.com/scholar_lookup?title=World+Animal+Health+Infromation+Database-Version:+1.4&author=OIE&publication_year=2009 (accessed on 4 January 2022).
- Murakami, K.; Kobayashi, S.; Konishi, M.; Kameyama, K.-I.; Tsutsui, T. Nationwide Survey of Bovine Leukemia Virus Infec-tion among Dairy and Beef Breeding Cattle in Japan from 2009–2011. J. Vet. Med. Sci. 2013, 75, 1123–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakada, S.; Fujimoto, Y.; Kohara, J.; Adachi, Y.; Makita, K. Estimation of economic loss by carcass weight reduction of Japanese dairy cows due to infection with bovine leukemia virus. Prev. Vet. Med. 2021, 198, 105528. [Google Scholar] [CrossRef] [PubMed]
- Benitez, O.J.; Norby, B.; Bartlett, P.C.; Maeroff, J.E.; Grooms, D.L. Impact of bovine leukemia virus infection on beef cow longevity. Prev. Vet. Med. 2020, 181, 105055. [Google Scholar] [CrossRef] [PubMed]
- Nekouei, O.; VanLeeuwen, J.; Stryhn, H.; Kelton, D.; Keefe, G. Lifetime effects of infection with bovine leukemia virus on longevity and milk production of dairy cows. Prev. Vet. Med. 2016, 133, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Norby, B.; Bartlett, P.; Byrem, T.; Erskine, R. Effect of infection with bovine leukemia virus on milk production in Michigan dairy cows. J. Dairy Sci. 2016, 99, 2043–2052. [Google Scholar] [CrossRef] [Green Version]
- Bartlett, P.C.; Norby, B.; Byrem, T.M.; Parmelee, A.; Ledergerber, J.T.; Erskine, R.J. Bovine leukemia virus and cow longevity in Michigan dairy herds. J. Dairy Sci. 2013, 96, 1591–1597. [Google Scholar] [CrossRef] [Green Version]
- Neefjes, J.; Jongsma, M.L.; Paul, P.; Bakke, O. Towards a systems understanding of MHC class I and MHC class II antigen presentation. Nat. Rev. Immunol. 2011, 11, 823–836. [Google Scholar] [CrossRef]
- Loat, S.; Kumari, N.; Saini, S.; Dige, M.S.; Kumar, A.; Dhilor, N.; Dang, A.K.; Lathwal, S.S.; Sodhi, M.; Kataria, R.S. Allelic diversity at BoLA DRB3 locus and association with predisposition to clinical mastitis in indicus and crossbred cattle. Anim. Biotechnol. 2021, 12, 1–10. [Google Scholar] [CrossRef]
- Duangjinda, M.; Jindatajak, Y.; Tipvong, W.; Sriwarothai, J.; Pattarajinda, V.; Katawatin, S.; Boonkum, W. Association of BoLA-DRB3 alleles with tick-borne disease tolerance in dairy cattle in a tropical environment. Vet Parasitol. 2013, 196, 314–320. [Google Scholar] [CrossRef]
- Martinez, M.L.; Machado, M.A.; Nascimento, C.S.; Silva, M.V.; Teodoro, R.L.; Furlong, J.; Prata, M.C.; Campos, A.L. Association of BoLA-DRB3.2 alleles with tick (Boophilus microplus) resistance in cattle. Genet. Mol. Res. 2006, 5, 513–524. [Google Scholar] [PubMed]
- Othman, O.E.; Khodary, M.G.; El-Deeb, A.H.; Hussein, H.A. Five BoLA-DRB3 genotypes detected in Egyptian buffalo infected with Foot and Mouth disease virus serotype O. J. Genet. Eng. Biotechnol. 2018, 16, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Morales, J.P.A.; López-Herrera, A.; Zuluaga, J.E. Association of BoLA DRB3 gene polymorphisms with BoHV-1 infection and zootechnical traits. Open Vet. J. 2020, 10, 331–339. [Google Scholar] [CrossRef]
- Longeri, M.; Russo, V.; Strillacci, M.G.; Perillo, A.; Carisetti, M.; Cozzi, M.C.; Neola, B.; Roperto, S. Association Between BoLA-DRB3.2 Polymorphism and Bovine Papillomavirus Infection for Bladder Tumor Risk in Podolica Cattle. Front. Vet. Sci. 2021, 8, 630089. [Google Scholar] [CrossRef]
- Zambrano, J.A.; Echeverri, J.Z.; López-Herrera, A. Association of gene BoLA DRB3.2 with production traits in a dairy herd of Antioquia, Colombia. Rev. MVZ Cordoba. 2014, 19, 4116–4129. [Google Scholar] [CrossRef] [Green Version]
- Derakhshani, H.; Plaizier, J.C.; De Buck, J.; Barkema, H.W.; Khafipour, E. Association of bovine major histocompatibility complex (BoLA) gene polymorphism with colostrum and milk microbiota of dairy cows during the first week of lactation. Microbiome 2018, 6, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwab, A.E.; Geary, T.G.; Baillargeon, P.; Schwab, A.J.; Fecteau, G. Association of BoLA DRB3 and DQA1 alleles with susceptibility to Neospora caninum and reproductive outcome in Quebec Holstein cattle. Vet Parasitol. 2009, 165, 136–140. [Google Scholar] [CrossRef]
- Rupp, R.; Hernandez, A.; Mallard, B.A. Association of bovine leukocyte antigen (BoLA) DRB3.2 with immune response, mastitis, and production and type traits in Canadian Holsteins. J. Dairy Sci. 2007, 90, 1029–1038. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Furuta, H.; Kondo, Y.; Mukoyama, H. Association of BoLA-DRB3 alleles with mastitis resistance and susceptibility in Japanese Holstein cows. Anim. Sci. J. 2012, 83, 359–366. [Google Scholar] [CrossRef]
- Pashmi, M.; Qanbari, S.; Ghorashi, S.A.; Sharifi, A.R.; Simianer, H. Analysis of relationship between bovine lymphocyte antigen DRB3.2 alleles, somatic cell count and milk traits in Iranian Holstein population. J. Anim. Breed Genet. 2009, 126, 296–303. [Google Scholar] [CrossRef]
- Sharif, S.; Mallard, B.A.; Wilkie, B.N.; Sargeant, J.M.; Scott, H.M.; Dekkers, J.C.; Leslie, K.E. Associations of the bovine major histocompatibility complex DRB3 (BoLA-DRB3) alleles with occurrence of disease and milk somatic cell score in Canadian dairy cattle. Anim. Genet. 1998, 29, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.X.; Yang, Z.P.; Wang, X.L.; Mao, Y.J.; Li, S.C.; Shi, X.K.; Chen, Y. Restriction fragment length polymorphism in the exon 2 of the BoLA-DRB3 gene in Chinese Holstein of the south China. J. Biomed Sci. Eng. 2010, 3, 221–225. [Google Scholar] [CrossRef] [Green Version]
- Zanotti, M.; Strillacci, M.G.; Taboni, I.; Samorè, A.B.; Longeri, M. Histocompatibility genes and somatic cell count (SCC) in Italian Holstein Friesian. Ital. J. Anim Sci. 2003, 2, 85–87. [Google Scholar] [CrossRef]
- Lo, C.-W.; Borjigin, L.; Saito, S.; Fukunaga, K.; Saitou, E.; Okazaki, K.; Mizutani, T.; Wada, S.; Takeshima, S.-N.; Aida, Y. BoLA-DRB3 Polymorphism is Associated with Differential Susceptibility to Bovine Leukemia Virus-Induced Lymphoma and Proviral Load. Viruses 2020, 12, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyasaka, T.; Takeshima, S.N.; Jimba, M.; Matsumoto, Y.; Kobayashi, N.; Matsuhashi, T.; Sentsui, H.; Aida, Y. Identification of bovine leukocyte antigen class II haplotypes associated with variations in bovine leukemia virus proviral load in Japanese Black cattle. Tissue Antigens 2013, 81, 72–82. [Google Scholar] [CrossRef]
- Borjigin, L.; Lo, C.-W.; Bai, L.; Hamada, R.; Sato, H.; Yoneyama, S.; Yasui, A.; Yasuda, S.; Yamanaka, R.; Mimura, M.; et al. Risk Assessment of Bovine Major Histocompatibility Complex Class II DRB3 Alleles for Perinatal Transmission of Bovine Leukemia Virus. Pathogens 2021, 10, 502. [Google Scholar] [CrossRef]
- Bai, L.; Borjigin, L.; Sato, H.; Takeshima, S.-N.; Asaji, S.; Ishizaki, H.; Kawashima, K.; Obuchi, Y.; Sunaga, S.; Ando, A.; et al. Kinetic Study of BLV Infectivity in BLV Susceptible and Resistant Cattle in Japan from 2017 to 2019. Pathogens 2021, 10, 1281. [Google Scholar] [CrossRef]
- Takeshima, S.H.; Matsumoto, Y.; Chen, J.; Yoshida, T.; Mukoyama, H.; Aida, Y. Evidence for cattle major histocompatibility complex (BoLA) class II DQA1 gene heterozygote advantage against clinical mastitis caused by streptococci and Escherichia species. Tissue Antigens 2008, 72, 525–531. [Google Scholar] [CrossRef]
- Forletti, A.; Juliarena, M.A.; Ceriani, C.; Amadio, A.F.; Esteban, E.; Gutiérrez, S.E. Identification of cattle carrying alleles associated with resistance and susceptibility to the Bovine Leukemia Virus progression by real-time PCR. Res. Vet. Sci. 2013, 95, 991–995. [Google Scholar] [CrossRef]
- Juliarena, M.A.; Poli, M.; Sala, L.; Ceriani, C.; Gutierrez, S.; Dolcini, G.; Rodríguez, E.M.; Mariño, B.; Rodríguez-Dubra, C.; Esteban, E.N. Association of BLV infection profiles with alleles of the BoLA-DRB3.2 gene. Anim Genet. 2008, 39, 432–438. [Google Scholar] [CrossRef]
- Lützelschwab, C.M.; Forletti, A.; Cepeda, R.; Esteban, E.N.; Confalonieri, O.; Gutiérrez, S.E. Co-infection with Mycobacterium bovis does not alter the response to bovine leukemia virus in BoLA DRB3*0902, genetically resistant cattle. Res. Vet. Sci. 2016, 109, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuchi, A.; Watanuki, S.; Borjigin, L.; Sato, H.; Bai, L.; Matsuura, R.; Kuroda, M.; Murakami, H.; Sato, R.; Asaji, S.; et al. BoLA-DRB3 Polymorphism Controls Proviral Load and Infectivity of Bovine Leukemia Virus (BLV) in Milk. Pathogens 2022, 11, 210. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuchi, A.; Bao, A.; Watanuki, S.; Matsuura, R.; Borjigin, L.; Bai, L.; Kuroda, M.; Matsumoto, Y.; Kohara, J.; Aida, Y. Anti-BLV antibodies in whey correlate with bovine leukemia virus disease progression and BoLA-DRB3 polymorphism. Front. Vet. Sci. 2022, 10, 3389. [Google Scholar] [CrossRef]
- Ray, D.E.; Halbach, T.J.; Armstrong, D.V. Season and lactation number effects on milk production and reproduction of dairy cattle in Arizona. J. Dairy Sci. 1992, 75, 2976–2983. [Google Scholar] [CrossRef] [PubMed]
- Vijayakumar, M.; Park, J.H.; Ki, K.S.; Lim, D.H.; Kim, S.B.; Park, S.M.; Jeong, H.Y.; Park, B.Y.; Kim, T.I. The effect of lactation number, stage, length, and milking frequency on milk yield in Korean Holstein dairy cows using automatic milking system. Asian-Australas J. Anim. Sci. 2017, 30, 1093–1098. [Google Scholar] [CrossRef] [PubMed]
- Strucken, E.M.; Laurenson, Y.C.; Brockmann, G.A. Go with the flow—Biology and genetics of the lactation cycle. Front. Genet. 2015, 6, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Gong, Z.; Lu, Y.; Lu, X.; Zhang, J.; Meng, Y.; Peng, Y.; Chu, S.; Cao, W.; Hao, X.; et al. Dairy Cows Experimentally Infected With Bovine Leukemia Virus Showed an Increased Milk Production in Lactation Numbers 3–4: A 4-Year Longitudinal Study. Front. Microbiol. 2022, 13, 946463. [Google Scholar] [CrossRef] [PubMed]
- Moriya, K.; Hirooka, H. Least squares analysis of variance and least squares means using the R package. Anim. Sci. J. 2018, 89, 1–8. [Google Scholar] [CrossRef]
- Abdalla, E.A.; Weigel, K.A.; Byrem, T.M.; Rosa, G.J.M. Short communication: Genetic correlation of bovine leukosis incidence with somatic cell score and milk yield in a US Holstein population. J. Dairy Sci. 2016, 99, 2005–2009. [Google Scholar] [CrossRef] [Green Version]
- Erskine, R.J.; Bartlett, P.C.; Byrem, T.M.; Render, C.L.; Febvay, C.; Houseman, J.T. Association between bovine leukemia virus, production, and population age in Michigan dairy herds. J. Dairy Sci. 2012, 95, 727–734. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Fan, W.; Mao, Y.; Yang, Z.; Lu, G.; Zhang, R.; Zhang, H.; Szeto, C.; Wang, C. Bovine leukemia virus infection in cattle of China: Association with reduced milk production and increased somatic cell score. J. Dairy Sci. 2016, 99, 3688–3697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorge, U.S.; Lissemore, K.; Cantin, R.; Kelton, D.F. Short communication: Milk ELISA status for bovine leukosis virus infection is not associated with milk production in dairy cows. J. Dairy Sci. 2011, 94, 5062–5064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, C.W.; Takeshima, S.N.; Okada, K.; Saitou, E.; Fujita, T.; Matsumoto, Y.; Wada, S.; Inoko, H.; Aida, Y. Association of Bovine Leukemia Virus-Induced Lymphoma with BoLA-DRB3 Polymorphisms at DNA, Amino Acid, and Binding Pocket Property Levels. Pathogens 2021, 10, 437. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Farm | Total Dam | Susceptible Dam b | Neutral Dam c | Resistant Dam d | |||
|---|---|---|---|---|---|---|---|
| BLV+ a (%) | BLV− a (%) | BLV+ a (%) | BLV− a (%) | BLV+ a (%) | BLV− a (%) | ||
| A | 88 | 15 (17.0) | 10 (11.4) | 15 (17.0) | 20 (22.7) | 10 (11.4) | 18 (20.5) |
| B | 42 | 19 (45.2) | 0 (0.0) | 7 (16.7) | 1 (2.4) | 15 (35.7) | 0 (0.0) |
| C | 17 | 1 (5.8) | 2 (11.8) | 8 (47.0) | 2 (11.8) | 2 (11.8) | 2 (11.8) |
| Total | 147 | 35 (23.8) | 12 (8.2) | 30 (20.4) | 23 (15.6) | 27 (18.4) | 20 (13.6) |
| Milk Parameter | BoLA-DRB3 | Estimated Coefficients | Standard Error for the Coefficient | p-Value |
|---|---|---|---|---|
| Yield (kg) | Susceptible vs. Neutral | 0.271 | 1.66 | 0.9855 |
| Susceptible vs. Resistant | 2.432 | 1.73 | 0.3395 | |
| Neutral vs. Resistant | 2.703 | 1.67 | 0.2432 | |
| Fat (%) | Susceptible vs. Neutral | 0.0939 | 0.127 | 0.7421 |
| Susceptible vs. Resistant | 0.0500 | 0.127 | 0.9183 | |
| Neutral vs. Resistant | 0.0439 | 0.126 | 0.9355 | |
| Protein (%) | Susceptible vs. Neutral | 0.0421 | 0.0570 | 0.7421 |
| Susceptible vs. Resistant | 0.0831 | 0.0579 | 0.3264 | |
| Neutral vs. Resistant | 0.1252 | 0.0567 | 0.0735 | |
| Non-fat Solids (%) | Susceptible vs. Neutral | 0.0501 | 0.0706 | 0.7588 |
| Susceptible vs. Resistant | 0.0919 | 0.0717 | 0.4079 | |
| Neutral vs. Resistant | 0.1420 | 0.0702 | 0.1108 | |
| SCC (103 cells/mL) | Susceptible vs. Neutral | 125.2 | 121 | 0.5583 |
| Susceptible vs. Resistant | 124.3 | 125 | 0.5811 | |
| Neutral vs. Resistant | 0.837 | 121 | 1.000 | |
| MUN (mg/dL) | Susceptible vs. Neutral | 0.827 | 0.391 | 0.0910 |
| Susceptible vs. Resistant | 0.183 | 0.390 | 0.8863 | |
| Neutral vs. Resistant | 0.644 | 0.387 | 0.2234 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakatsuchi, A.; Matsumoto, Y.; Aida, Y. Influence of BoLA-DRB3 Polymorphism and Bovine Leukemia Virus (BLV) Infection on Dairy Cattle Productivity. Vet. Sci. 2023, 10, 250. https://doi.org/10.3390/vetsci10040250
Nakatsuchi A, Matsumoto Y, Aida Y. Influence of BoLA-DRB3 Polymorphism and Bovine Leukemia Virus (BLV) Infection on Dairy Cattle Productivity. Veterinary Sciences. 2023; 10(4):250. https://doi.org/10.3390/vetsci10040250
Chicago/Turabian StyleNakatsuchi, Ayumi, Yasunobu Matsumoto, and Yoko Aida. 2023. "Influence of BoLA-DRB3 Polymorphism and Bovine Leukemia Virus (BLV) Infection on Dairy Cattle Productivity" Veterinary Sciences 10, no. 4: 250. https://doi.org/10.3390/vetsci10040250