
Host PTX3 Protein and Bacterial Capsule Coordinately Regulate the Inflammatory Response during Streptococcus suis Infection
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Bacterial Strains and Cells
2.3. RNA Isolation and qRT-PCR Analysis
2.4. Production, Purification, and Identification of Recombinant Murine PTX3
2.5. Preparation of Capsular Polysaccharide from S. suis 2
2.6. Regulating Phagocytosis of S. suis 2 HA9801 by Ana-1
2.7. Mouse Air Pouch Model
2.8. Inflammatory Cells and IL-6 Detection
2.9. Mouse Infection Assays
2.10. Statistical Analysis
3. Results
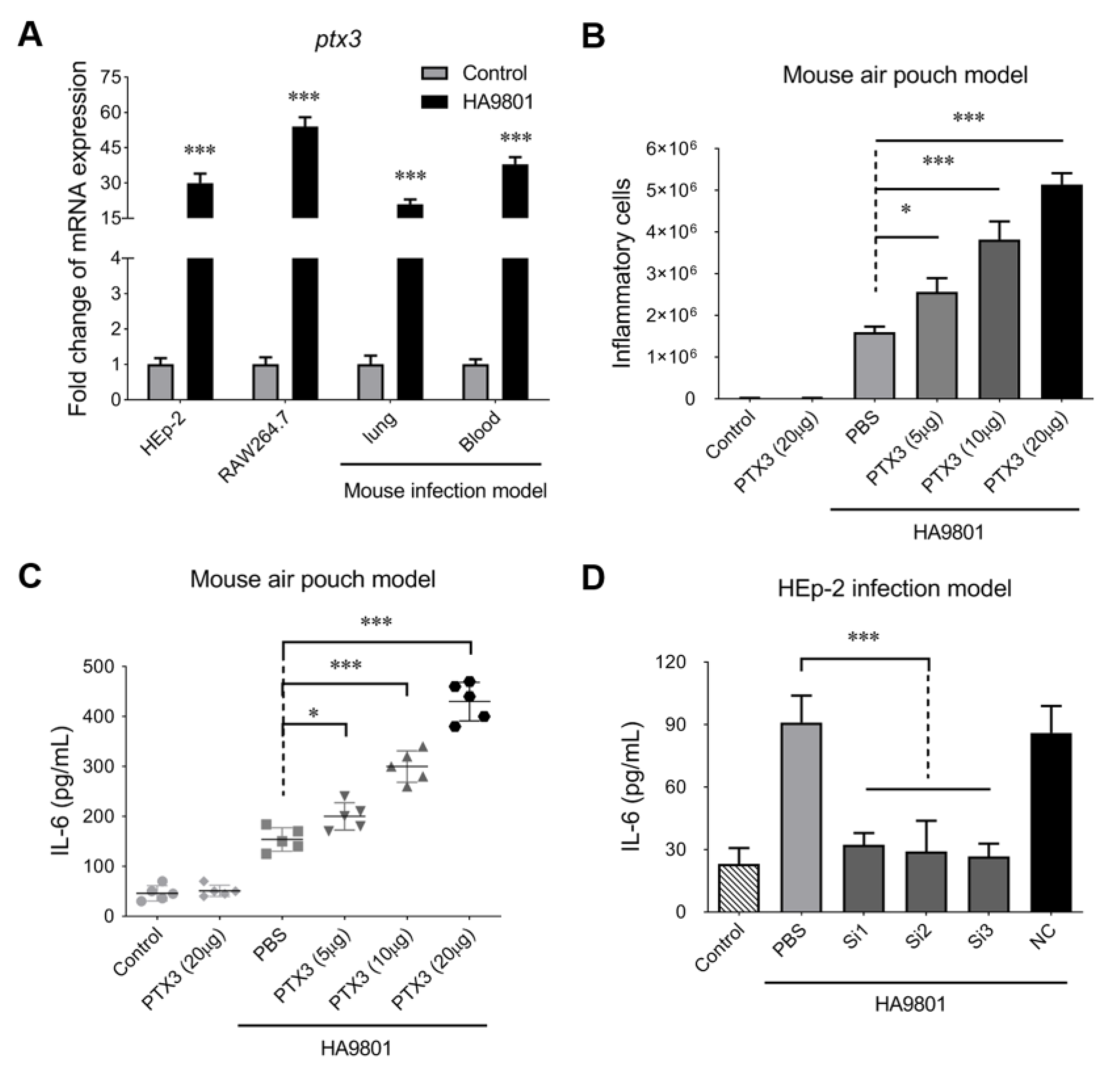
3.1. PTX3 Protein Is Responsible for the Significant Inflammatory Response during SS Infection
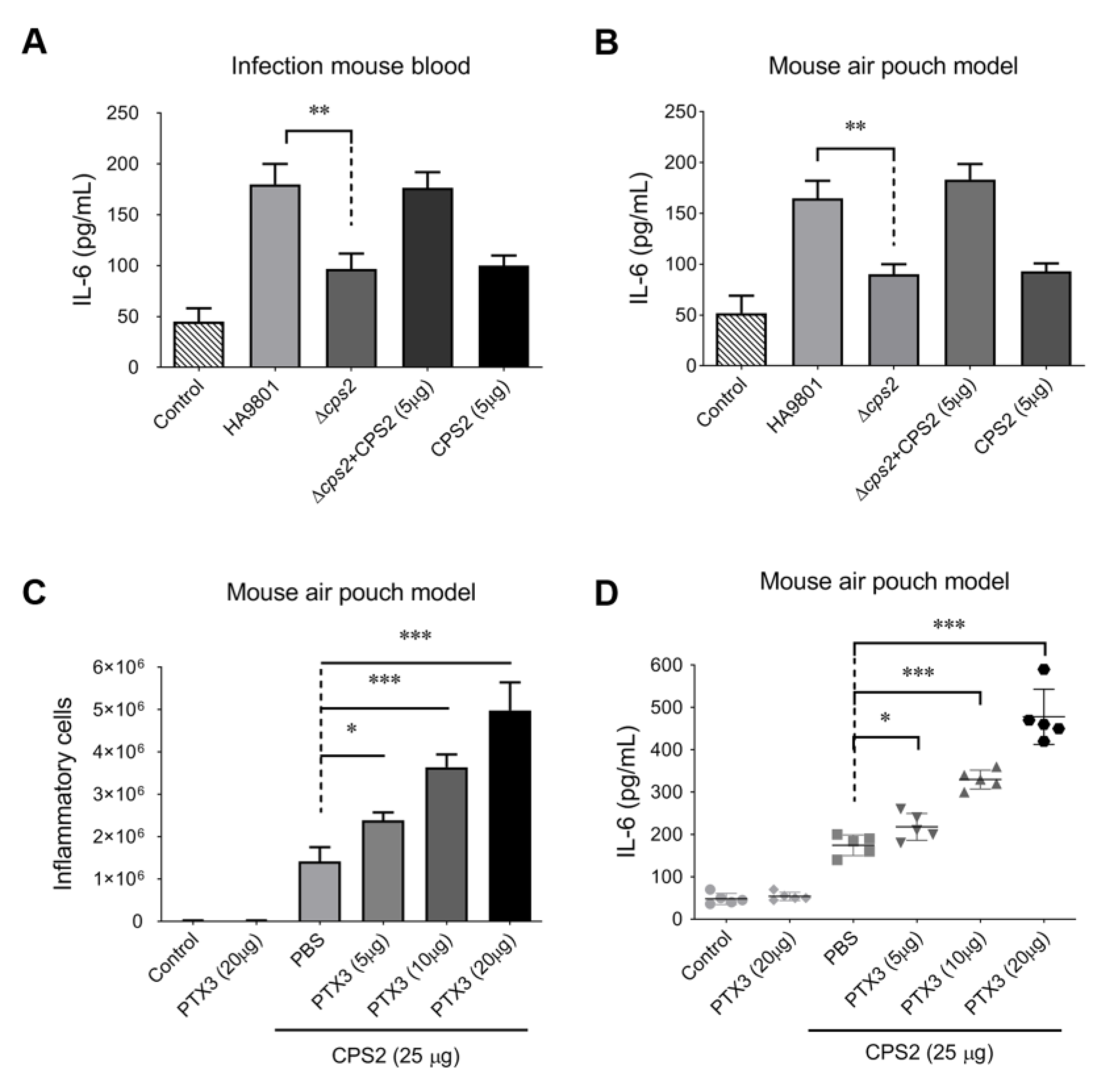
3.2. The CPS2 Is Required for Triggering the Host Inflammatory Response during SS2 Infection with or without Exogenous PTX3
3.3. PTX3 Enhances the Phagocytosis of Ana-1 cells to SS2 Strain HA9801
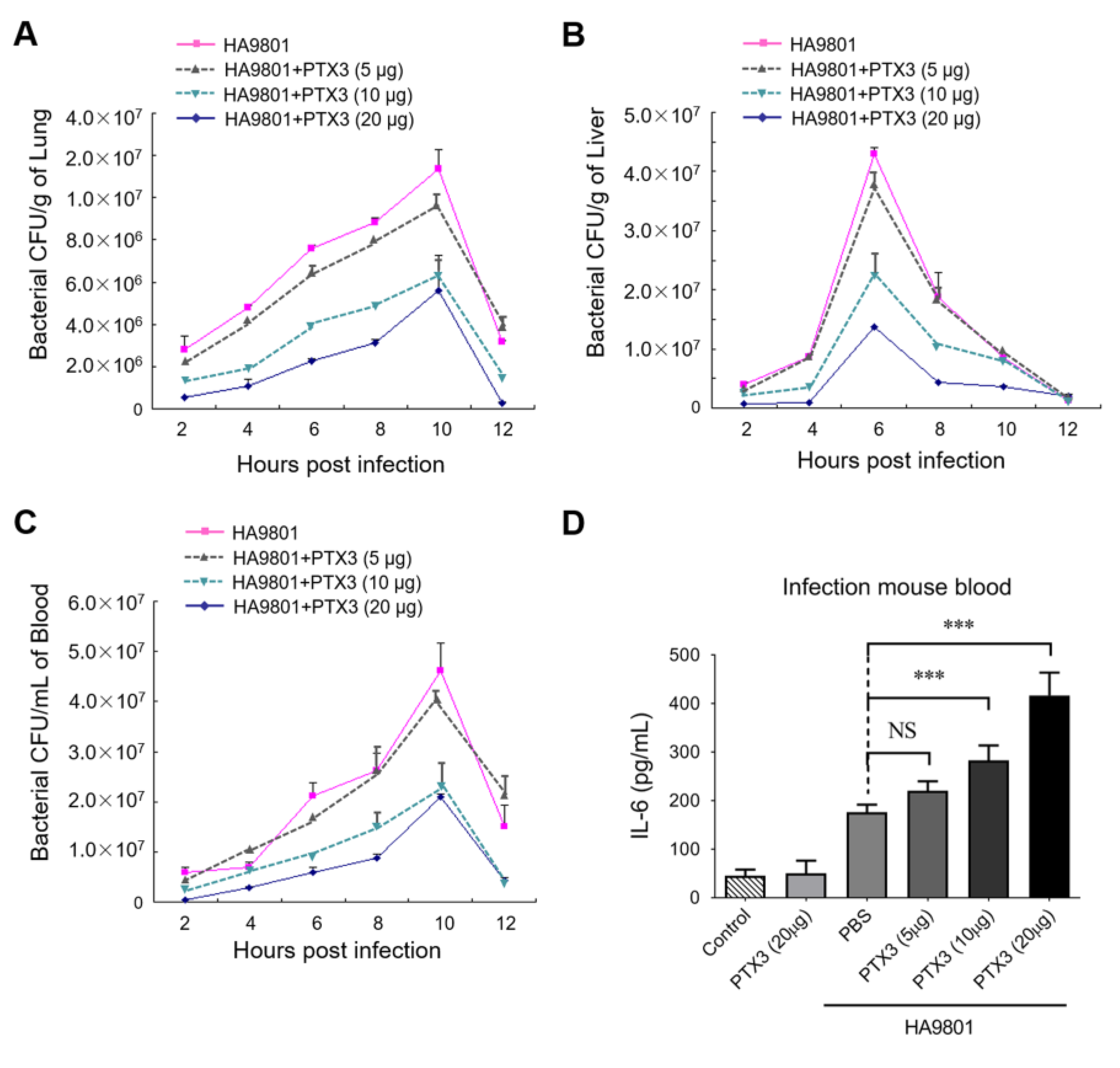
3.4. PTX3 Contributes to Attenuate the Bacterial Colonization/Proliferation in Multiple Organs during the HA9801 Infection
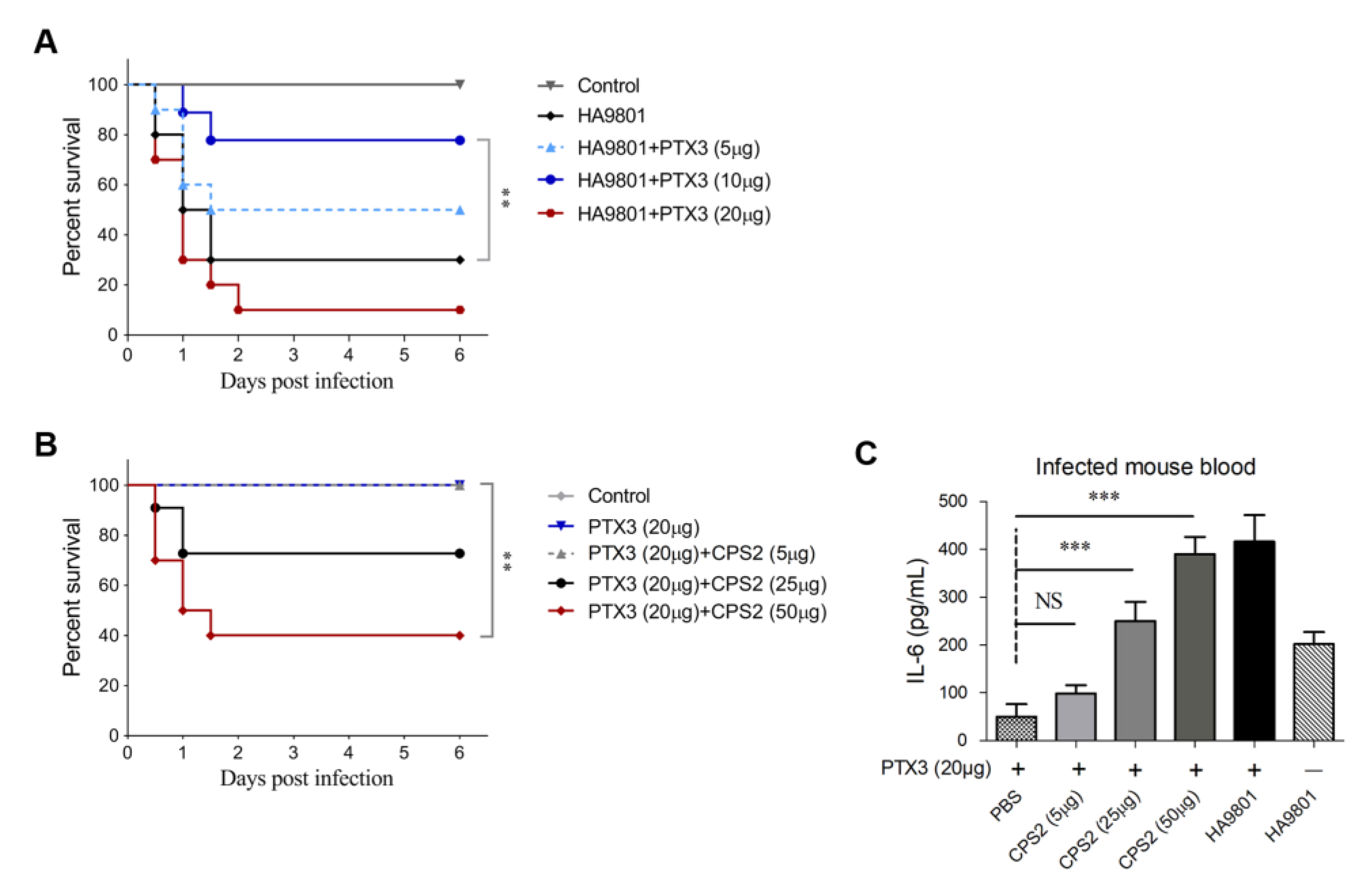
3.5. Intravenous Injection of PTX3 at an Appropriate Dose Significantly Improves Mouse Survival after SS2 Challenge
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bonifait, L.; Veillette, M.; Létourneau, V.; Grenier, D.; Duchaine, C. Detection of Streptococcus suis in bioaerosols of swine confinement buildings. Appl. Environ. Microbiol. 2014, 80, 3296–3304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Jing, H.; Chen, Z.; Zheng, H.; Zhu, X.; Wang, H.; Wang, S.; Liu, L.; Zu, R.; Luo, L.; et al. Human Streptococcus suis outbreak, Sichuan, China. Emerg. Infect. Infect. Dis. 2006, 12, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; Du, D.; Yu, Y.; Zhang, Y.; Qian, Y.; Zhang, W. Pan-genome analysis of Streptococcus suis serotype 2 revealed genomic diversity among strains of different virulence. Transbound. Emerg. Dis. 2021, 68, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Kerdsin, A.; Takeuchi, D.; Akeda, Y.; Nakamura, S.; Gottschalk, M.; Oishi, K. Genomic differences between sequence types 1 and 104 of Streptococcus suis Serotype 2. PeerJ 2022, 10, e14144. [Google Scholar] [CrossRef]
- Lapointe, L.; D’Allaire, S.; Lebrun, A.; Lacouture, S.; Gottschalk, M. Antibody response to an autogenous vaccine and serologic profile for Streptococcus suis capsular type 1/2. Can. J. Vet. Res. Rev. Can. De Rech. Vet. 2002, 66, 8–14. [Google Scholar]
- Smith, H.E.; Damman, M.; van der Velde, J.; Wagenaar, F.; Wisselink, H.J.; Stockhofe-Zurwieden, N.; Smits, M.A. Identification and characterization of the cps locus of Streptococcus suis serotype 2: The capsule protects against phagocytosis and is an important virulence factor. Infect. Immun. 1999, 67, 1750–1756. [Google Scholar] [CrossRef]
- Auger, J.P.; Dolbec, D.; Roy, D.; Segura, M.; Gottschalk, M. Role of the Streptococcus suis serotype 2 capsular polysaccharide in the interactions with dendritic cells is strain-dependent but remains critical for virulence. PLoS ONE 2018, 13, e0200453. [Google Scholar] [CrossRef]
- Kock, C.; Beineke, A.; Seitz, M.; Ganter, M.; Waldmann, K.H.; Valentin-Weigand, P.; Baums, C.G. Intranasal immunization with a live Streptococcus suis isogenic ofs mutant elicited suilysin-neutralization titers but failed to induce opsonizing antibodies and protection. Vet. Immunol. Immunopathol. 2009, 132, 135–145. [Google Scholar] [CrossRef]
- Segura, M. Streptococcus suis vaccines: Candidate antigens and progress. Expert Rev. Vaccines 2015, 14, 1587–1608. [Google Scholar] [CrossRef]
- Baums, C.G.; Kock, C.; Beineke, A.; Bennecke, K.; Goethe, R.; Schröder, C.; Waldmann, K.H.; Valentin-Weigand, P. Streptococcus suis bacterin and subunit vaccine immunogenicities and protective efficacies against serotypes 2 and 9. Clin. Vaccine Immunol. CVI 2009, 16, 200–208. [Google Scholar] [CrossRef] [Green Version]
- Bottazzi, B.; Doni, A.; Garlanda, C.; Mantovani, A. An integrated view of humoral innate immunity: Pentraxins as a paradigm. Annu. Rev. Immunol. 2009, 28, 157–183. [Google Scholar] [CrossRef] [PubMed]
- Introna, M.; Alles, V.V.; Castellano, M.; Picardi, G.; De Gioia, L.; Bottazzai, B.; Peri, G.; Breviario, F.; Salmona, M.; De Gregorio, L.; et al. Cloning of mouse ptx3, a new member of the pentraxin gene family expressed at extrahepatic sites. Blood 1996, 87, 1862–1872. [Google Scholar] [CrossRef] [Green Version]
- Bottazzi, B.; Vouret-Craviari, V.; Bastone, A.; De Gioia, L.; Matteucci, C.; Peri, G.; Spreafico, F.; Pausa, M.; D’Ettorre, C.; Gianazza, E.; et al. Multimer formation and ligand recognition by the long pentraxin PTX3. Similarities and differences with the short pentraxins C-reactive protein and serum amyloid P component. J. Biol. Chem. 1997, 272, 32817–32823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garlanda, C.; Jaillon, S.; Doni, A.; Bottazzi, B.; Mantovani, A. PTX3, a humoral pattern recognition molecule at the interface between microbe and matrix recognition. Curr. Opin. Immunol. 2016, 38, 39–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nauta, A.J.; Bottazzi, B.; Mantovani, A.; Salvatori, G.; Kishore, U.; Schwaeble, W.J.; Gingras, A.R.; Tzima, S.; Vivanco, F.; Egido, J.; et al. Biochemical and functional characterization of the interaction between pentraxin 3 and C1q. Eur. J. Immunol. 2003, 33, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Garlanda, C.; Hirsch, E.; Bozza, S.; Salustri, A.; De Acetis, M.; Nota, R.; Maccagno, A.; Riva, F.; Bottazzi, B.; Peri, G.; et al. Non-redundant role of the long pentraxin PTX3 in anti-fungal innate immune response. Nature 2002, 420, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Bozza, S.; Bistoni, F.; Gaziano, R.; Pitzurra, L.; Zelante, T.; Bonifazi, P.; Perruccio, K.; Bellocchio, S.; Neri, M.; Iorio, A.M.; et al. Pentraxin 3 protects from MCMV infection and reactivation through TLR sensing pathways leading to IRF3 activation. Blood 2006, 108, 3387–3396. [Google Scholar] [CrossRef] [Green Version]
- Bonavita, E.; Gentile, S.; Rubino, M.; Maina, V.; Papait, R.; Kunderfranco, P.; Greco, C.; Feruglio, F.; Molgora, M.; Laface, I.; et al. PTX3 is an extrinsic oncosuppressor regulating complement-dependent inflammation in cancer. Cell 2015, 160, 700–714. [Google Scholar] [CrossRef] [Green Version]
- Jeon, H.; Lee, S.; Lee, W.H.; Suk, K. Analysis of glial secretome: The long pentraxin PTX3 modulates phagocytic activity of microglia. J. Neuroimmunol. 2010, 229, 63–72. [Google Scholar] [CrossRef]
- Deban, L.; Russo, R.C.; Sironi, M.; Moalli, F.; Scanziani, M.; Zambelli, V.; Cuccovillo, I.; Bastone, A.; Gobbi, M.; Valentino, S.; et al. Regulation of leukocyte recruitment by the long pentraxin PTX3. Nat. Immunol. 2010, 11, 328–334. [Google Scholar] [CrossRef]
- Brunetta, E.; Folci, M.; Bottazzi, B.; De Santis, M.; Gritti, G.; Protti, A.; Mapelli, S.N.; Bonovas, S.; Piovani, D.; Leone, R.; et al. Macrophage expression and prognostic significance of the long pentraxin PTX3 in COVID-19. Nat. Immunol. 2021, 22, 19–24. [Google Scholar] [CrossRef]
- Zhang, Y.; Tedrow, J.; Nouraie, M.; Li, X.; Chandra, D.; Bon, J.; Kass, D.J.; Fuhrman, C.R.; Leader, J.K.; Duncan, S.R.; et al. Elevated plasma level of Pentraxin 3 is associated with emphysema and mortality in smokers. Thorax 2021, 76, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Rivieccio, V.; Esposito, A.; Bellofiore, P.; Palladino, P.; Sassano, M.; Colombo, M.; Verdoliva, A. High-level expression and efficient purification of recombinant human long pentraxin PTX3 in Chinese hamster ovary cells. Protein Expr. Purif. 2007, 51, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Gaikwad, W.K.; Jana, S.K.; Dhere, R.M.; Ravenscroft, N.; Kodam, K.M. Purification of capsular polysaccharides isolated from S. pneumoniae serotype 2 by hydrogen peroxide and endonuclease. Carbohydr. Polym. 2022, 294, 119783. [Google Scholar] [CrossRef]
- He, J.; Shi, Q.; Chen, Z.; Zhang, W.; Lan, P.; Xu, Q.; Hu, H.; Chen, Q.; Fan, J.; Jiang, Y.; et al. Opposite evolution of pathogenicity driven by in vivo wzc and wcaJ mutations in ST11-KL64 carbapenem-resistant Klebsiella pneumoniae. Drug Resist. Updates Rev. Comment. Antimicrob. Anticancer. Chemother. 2022, 66, 100891. [Google Scholar] [CrossRef]
- Diniz, S.N.; Nomizo, R.; Cisalpino, P.S.; Teixeira, M.M.; Brown, G.D.; Mantovani, A.; Gordon, S.; Reis, L.F.; Dias, A.A. PTX3 function as an opsonin for the dectin-1-dependent internalization of zymosan by macrophages. J. Leukoc. Biol. 2004, 75, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Cotena, A.; Maina, V.; Sironi, M.; Bottazzi, B.; Jeannin, P.; Vecchi, A.; Corvaia, N.; Daha, M.R.; Mantovani, A.; Garlanda, C. Complement dependent amplification of the innate response to a cognate microbial ligand by the long pentraxin PTX3. J. Immunol. 2007, 179, 6311–6317. [Google Scholar] [CrossRef] [Green Version]
- Bai, Q.; Ma, J.; Zhang, Z.; Zhong, X.; Pan, Z.; Zhu, Y.; Zhang, Y.; Wu, Z.; Liu, G.; Yao, H. YSIRK-G/S-directed translocation is required for Streptococcus suis to deliver diverse cell wall anchoring effectors contributing to bacterial pathogenicity. Virulence 2020, 11, 1539–1556. [Google Scholar] [CrossRef]
- Rong, J.; Zhang, W.; Wang, X.; Fan, H.; Lu, C.; Yao, H. Identification of candidate susceptibility and resistance genes of mice infected with Streptococcus suis type 2. PLoS ONE 2012, 7, e32150. [Google Scholar] [CrossRef]
- Xu, J.; Mu, Y.; Zhang, Y.; Dong, W.; Zhu, Y.; Ma, J.; Song, W.; Pan, Z.; Lu, C.; Yao, H. Antibacterial effect of porcine PTX3 against Streptococcus suis type 2 infection. Microb. Pathog. 2015, 89, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, R.; Wang, Z.; Wu, W.; Zhang, N.; Zhang, L.; Hu, J.; Luo, P.; Zhang, J.; Liu, Z.; et al. Molecular insight into pentraxin-3: Update advances in innate immunity, inflammation, tissue remodeling, diseases, and drug role. Biomed. Pharmacother. Biomed. Pharmacother. 2022, 156, 113783. [Google Scholar] [CrossRef] [PubMed]
- Paton, J.C.; Trappetti, C. Streptococcus pneumoniae Capsular Polysaccharide. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Uruén, C.; García, C.; Fraile, L.; Tommassen, J.; Arenas, J. How Streptococcus suis escapes antibiotic treatments. Vet. Res. 2022, 53, 91. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Ma, D.; Liu, Y.; Sun, W.; Cheng, D.; Li, G.; Zhou, S.; Wang, Y.; Wang, H.; Ni, C. PTX3 alleviates hard metal-induced acute lung injury through potentiating efferocytosis. Ecotoxicol. Environ. Saf. 2022, 230, 113139. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Vedi, S.; Garg, S.; Loo, E.; Li, J.; Kunimoto, D.Y.; Kumar, R.; Agrawal, B. Harnessing Innate Immunity to Treat Mycobacterium tuberculosis Infections: Heat-Killed Caulobacter crescentus as a Novel Biotherapeutic. Cells 2023, 12, 560. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Yasmeen, N.; Chaudhary, A.A.; Alawam, A.S.; Al-Zharani, M.; Suliman Basher, N.; Harikrishnan, S.; Goud, M.D.; Pandey, A.; Lakhawat, S.S.; et al. Specialized pro-resolving lipid mediators regulate inflammatory macrophages: A paradigm shift from antibiotics to immunotherapy for mitigating COVID-19 pandemic. Front. Mol. Biosci. 2023, 10, 1104577. [Google Scholar] [CrossRef]
- Tidswell, R.; Singer, M. Sepsis—Thoughtful management for the non-expert. Clin. Med. 2018, 18, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Zhang, Y.; Wang, Y. Immunotherapy strategies and prospects for acute lung injury: Focus on immune cells and cytokines. Front. Pharmacol. 2022, 13, 1103309. [Google Scholar] [CrossRef]
- Soares, A.C.; Souza, D.G.; Pinho, V.; Vieira, A.T.; Nicoli, J.R.; Cunha, F.Q.; Mantovani, A.; Reis, L.F.; Dias, A.A.; Teixeira, M.M. Dual function of the long pentraxin PTX3 in resistance against pulmonary infection with Klebsiella pneumoniae in transgenic mice. Microbes Infect. 2006, 8, 1321–1329. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, Q.; Fan, R.; Zhong, N.; Liu, J.; Pan, X.; Yao, H.; Ma, J. Host PTX3 Protein and Bacterial Capsule Coordinately Regulate the Inflammatory Response during Streptococcus suis Infection. Vet. Sci. 2023, 10, 239. https://doi.org/10.3390/vetsci10030239
Bai Q, Fan R, Zhong N, Liu J, Pan X, Yao H, Ma J. Host PTX3 Protein and Bacterial Capsule Coordinately Regulate the Inflammatory Response during Streptococcus suis Infection. Veterinary Sciences. 2023; 10(3):239. https://doi.org/10.3390/vetsci10030239
Chicago/Turabian StyleBai, Qiankun, Ruhui Fan, Ningyuan Zhong, Jianan Liu, Xinming Pan, Huochun Yao, and Jiale Ma. 2023. "Host PTX3 Protein and Bacterial Capsule Coordinately Regulate the Inflammatory Response during Streptococcus suis Infection" Veterinary Sciences 10, no. 3: 239. https://doi.org/10.3390/vetsci10030239