

The Middle Ear Microbiota in Healthy Dogs Is Similar to That of the External Ear Canal
 , , , , and
, , , , and 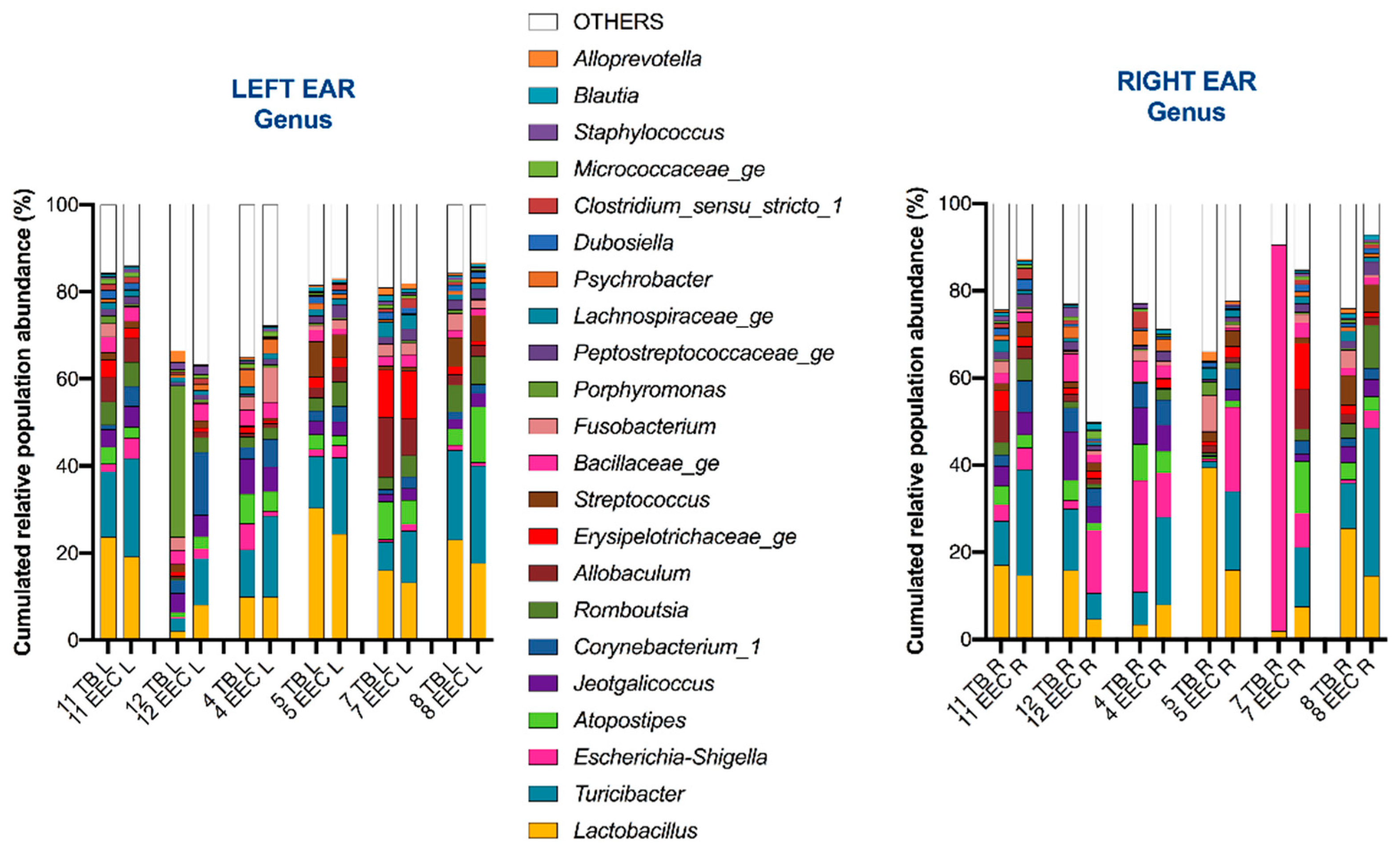{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Sample Collection
2.3. 16S rDNA Extraction and High Throughput Sequencing
2.4. Data Analysis
3. Results
3.1. Study Population
3.2. Most Common Taxa
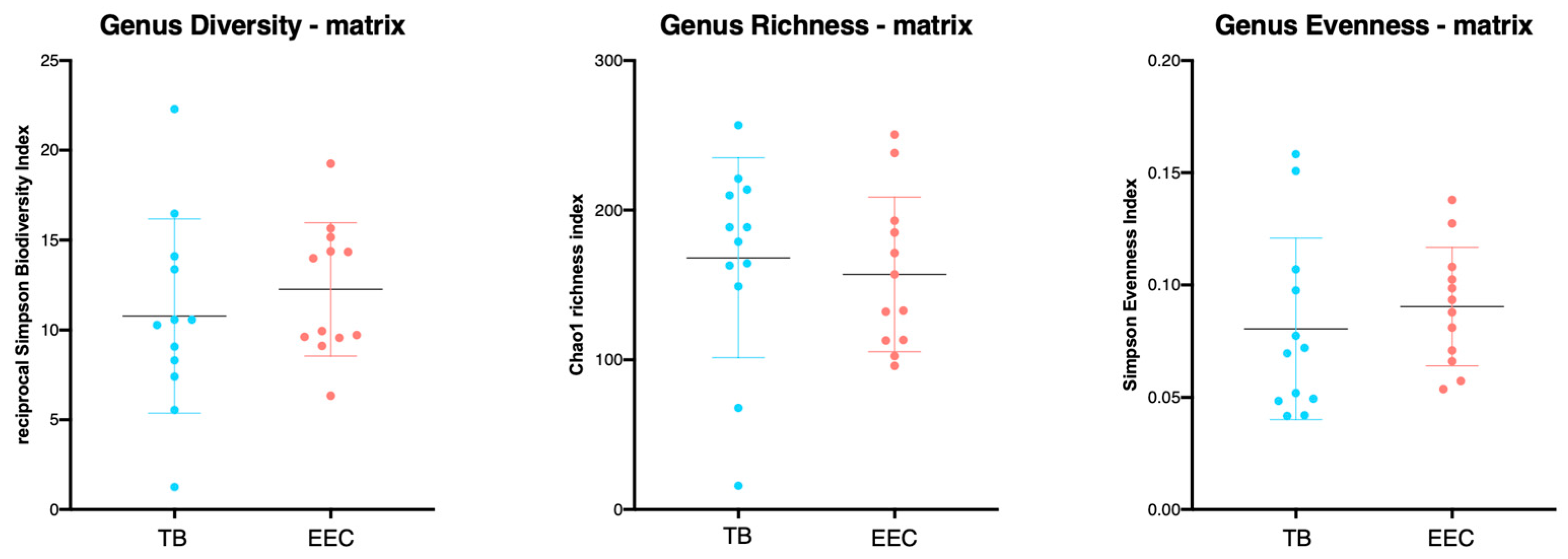
3.3. Richness, Evenness and α-Diversity
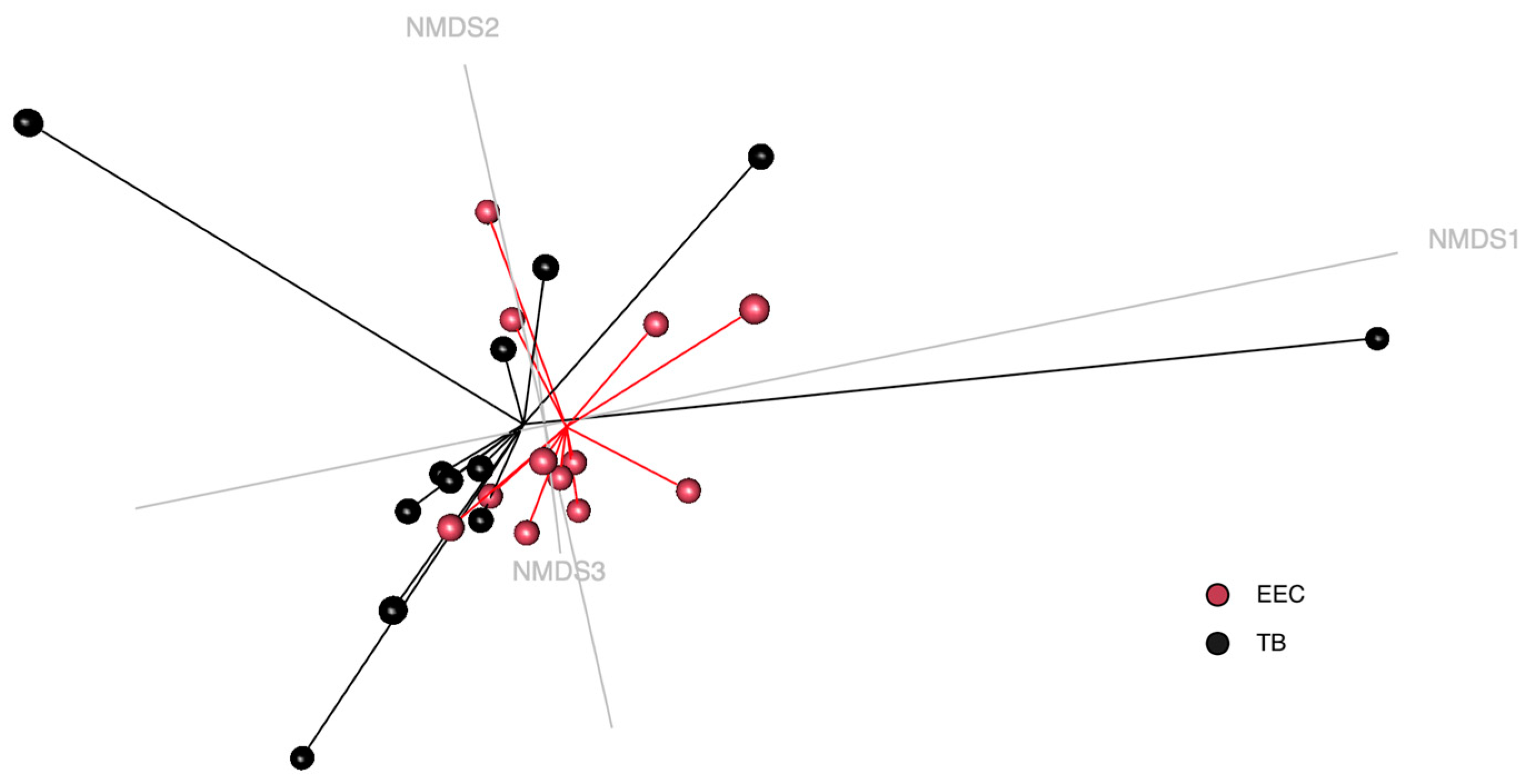
3.4. Beta-Diversity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Spreull, J.S.A. Treatment of Otitis Media in the Dog. J. Small Anim. Pract. 1964, 5, 107–122. [Google Scholar] [CrossRef]
- Gotthelf, L.N. Diagnosis and treatment of otitis media in dogs and cats. Vet. Clin. Small Anim. Pract. 2004, 34, 469–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, L.K.; Kwochka, K.W.; Kowalski, J.J.; Hillier, A. Microbial flora and antimicrobial susceptibility patterns of isolated pathogens from the horizontal ear canal and middle ear in dogs with otitis media. J. Am. Vet. Med. Assoc. 1998, 212, 534–538. [Google Scholar]
- Morris, D.O. Medical therapy of otitis externa and otitis media. Vet. Clin. Small Anim. Pract. 2004, 34, 541–555. [Google Scholar] [CrossRef]
- Bourély, C.; Cazeau, G.; Jarrige, N.; Leblond, A.; Madec, J.; Haenni, M.; Gay, E. Antimicrobial resistance patterns of bacteria isolated from dogs with otitis. Epidemiol. Infect. 2019, 147, e121. [Google Scholar] [CrossRef] [Green Version]
- Smeak, D.D.; Kerpsack, S.J. Total ear canal ablation and lateral bulla osteotomy for management of end-stage otitis. Semin. Vet. Med. Surg. Small Anim. 1993, 8, 30–41. [Google Scholar]
- Ngo, J.; Taminiau, B.; Fall, P.A.; Daube, G.; Fontaine, J. Ear canal microbiota—A comparison between healthy dogs and atopic dogs without clinical signs of otitis externa. Vet. Dermatol. 2018, 29, 425-e140. [Google Scholar] [CrossRef] [PubMed]
- Korbelik, J.; Singh, A.; Rousseau, J.; Weese, J.S. Characterization of the otic bacterial microbiota in dogs with otitis externa compared to healthy individuals. Vet. Dermatol. 2019, 30, 228-e70. [Google Scholar] [CrossRef]
- Borriello, G.; Paradiso, R.; Catozzi, C.; Brunetti, R.; Roccabianca, P.; Riccardi, M.G.; Cecere, B.; Lecchi, C.; Fusco, G.; Ceciliani, F.; et al. Cerumen microbial community shifts between healthy and otitis affected dogs. PLoS ONE 2020, 15, e0241447. [Google Scholar] [CrossRef] [PubMed]
- Puigdemont, A.; D’Andreano, S.; Ramió-Lluch, L.; Cuscó, A.; Francino, O.; Brazis, P. Effect of an anti-inflammatory pomegranate otic treatment on the clinical evolution and microbiota profile of dogs with otitis externa. Vet. Dermatol. 2021, 32, 158-e37. [Google Scholar] [CrossRef]
- Bradley, C.W.; Lee, F.F.; Rankin, S.C.; Kalan, L.R.; Horwinski, J.; Morris, D.O.; Grice, E.A.; Cain, C.L. The otic microbiota and mycobiota in a referral population of dogs in eastern USA with otitis externa. Vet. Dermatol. 2020, 31, 225-e49. [Google Scholar] [CrossRef] [PubMed]
- Kasai, T.; Fukui, Y.; Aoki, K.; Ishii, Y.; Tateda, K. Changes in the ear canal microbiota of dogs with otitis externa. J. Appl. Microbiol. 2021, 130, 1084–1091. [Google Scholar] [CrossRef] [PubMed]
- Léonard, C.; Taminiau, B.; Ngo, J.; Fantini, O.; Daube, G.; Fontaine, J. Preventive use of a topical anti-inflammatory glucocorticoid in atopic dogs without clinical sign of otitis does not affect ear canal microbiota and mycobiota. Vet. Dermatol. 2021, 32, 355-e98. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Tojo, M.; Fukui, K.; Imori, T.; Baba, E. The aerobic bacterial flora of the middle and external ears in normal dogs. J. Small Anim. Pract. 1984, 25, 269–274. [Google Scholar] [CrossRef]
- Cole, L.K.; Rajala-Schultz, P.J.; Lorch, G.; Daniels, J.B. Bacteriology and cytology of otic exudates in 41 cavalier King Charles spaniels with primary secretory otitis media. Vet. Dermatol. 2019, 30, 151-e44. [Google Scholar] [CrossRef]
- Hettlich, B.F.; Boothe, H.W.; Simpson, R.B.; DuBose, K.A.; Boothe, D.M.; Carpenter, M. Effect of tympanic cavity evacuation and flushing on microbial isolates during total ear canal ablation with lateral bulla osteotomy in dogs. J. Am. Vet. Med. Assoc. 2005, 227, 748–755. [Google Scholar] [CrossRef]
- Vogel, P.L.; Komtebedde, J.; Hirsh, D.C.; Kass, P.H. Wound contamination and antimicrobial susceptibility of bacteria cultured during total ear canal ablation and lateral bulla osteotomy in dogs. J. Am. Vet. Med. Assoc. 1999, 214, 1641–1643. [Google Scholar]
- Palmeiro, B.S.; Morris, D.O.; Wiemelt, S.P.; Shofer, F.S. Evaluation of outcome of otitis media after lavage of the tympanic bulla and long-term antimicrobial drug treatment in dogs: 44 cases (1998–2002). J. Am. Vet. Med. Assoc. 2004, 225, 548–553. [Google Scholar] [CrossRef]
- Milne, E.; Nuttall, T.; Marioni-Henry, K.; Piccinelli, C.; Schwarz, T.; Azar, A.; Harris, J.; Duncan, J.; Cheeseman, M. Cytological and microbiological characteristics of middle ear effusions in brachycephalic dogs. J. Vet. Intern. Med. 2020, 34, 1454–1463. [Google Scholar] [CrossRef]
- Reinbacher, E.; Kneissl, S.; Hirt, R.; Spergser, J.; Panakova, L. Myringotomy in dogs: Contamination rate from the external ear canal—A pilot study. Vet. Anim. Sci. 2020, 10, 100125. [Google Scholar] [CrossRef]
- Tobias, K.M.; Johnston, S.A. Pinna and external ear canal. In Veterinary Surgery: Small Animal; Elsevier Saunders: St Louis, MO, USA, 2017; pp. 2310–2328. [Google Scholar]
- Wimmer-Scherr, C.; Taminiau, B.; Renaud, B.; van Loon, G.; Palmers, K.; Votion, D.; Amory, H.; Daube, G.; Cesarini, C. Comparison of Fecal Microbiota of Horses Suffering from Atypical Myopathy and Healthy Co-Grazers. Animals 2021, 11, 506. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misic, A.M.; Davis, M.F.; Tyldsley, A.S.; Hodkinson, B.P.; Tolomeo, P.; Hu, B.; Nachamkin, I.; Lautenbach, E.; Morris, D.O.; Grice, E. The shared microbiota of humans and companion animals as evaluated from Staphylococcus carriage sites. Microbiome 2015, 3, 2. [Google Scholar] [CrossRef] [Green Version]
- Parks, D.H.; Beiko, R.G. Identifying biologically relevant differences between metagenomic communities. Bioinformatics 2010, 26, 715–721. [Google Scholar] [CrossRef] [Green Version]
- Jervis-Bardy, J.; Leong, L.E.X.; Papanicolas, L.E.; Ivey, K.; Chawla, S.; Woods, C.M.; Frauenfelder, C.; Ooi, E.H.; Rogers, G. Examining the Evidence for an Adult Healthy Middle Ear Microbiome. Msphere 2019, 4, e00456-19. [Google Scholar] [CrossRef] [Green Version]
- Brugger, S.D.; Kraemer, J.G.; Qi, W.; Bomar, L.; Oppliger, A.; Hilty, M. Age-Dependent Dissimilarity of the Nasopharyngeal and Middle Ear Microbiota in Children with Acute Otitis Media. Front. Genet. 2019, 10, 555. Available online: https://www.frontiersin.org/articles/10.3389/fgene.2019.00555 (accessed on 19 September 2022). [CrossRef] [PubMed]
- Man, W.H.; van Dongen, T.M.; Venekamp, R.P.; Pluimakers, V.G.; Chu, M.L.J.; van Houten, M.A.; Sanders, E.A.; Schilder, A.G.M.; Bogaert, D. Respiratory Microbiota Predicts Clinical Disease Course of Acute Otorrhea in Children with Tympanostomy Tubes. Pediatr. Infect. Dis. J. 2019, 38, e116. [Google Scholar] [CrossRef] [Green Version]
- Bhutta, M.F.; Thornton, R.B.; Kirkham, L.-A.S.; Kerschner, J.E.; Cheeseman, M.T. Understanding the aetiology and resolution of chronic otitis media from animal and human studies. Dis. Model. Mech. 2017, 10, 1289–1300. [Google Scholar] [CrossRef] [Green Version]
- Monasta, L.; Ronfani, L.; Marchetti, F.; Montico, M.; Brumatti, L.V.; Bavcar, A.; Grasso, D.; Barbiero, C.; Tamburlini, G. Burden of Disease Caused by Otitis Media: Systematic Review and Global Estimates. PLoS ONE 2012, 7, e36226. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, H.; Wallis, S.; Coatesworth, A.P. Acute otitis media. Postgrad. Med. 2015, 127, 386–390. [Google Scholar] [CrossRef]
- Cuscó, A.; Belanger, J.M.; Gershony, L.; Islas-Trejo, A.; Levy, K.; Medrano, J.F.; Sánchez, A.; Oberbauer, A.M.; Francino, O. Individual signatures and environmental factors shape skin microbiota in healthy dogs. Microbiome 2017, 5, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehtimäki, J.; Sinkko, H.; Hielm-Björkman, A.; Salmela, E.; Tiira, K.; Laatikainen, T.; Mäkeläinen, S.; Kaukonen, M.; Uusitalo, L.; Hanski, I.; et al. Skin microbiota and allergic symptoms associate with exposure to environmental microbes. Proc. Natl. Acad. Sci. USA 2018, 115, 4897–4902. [Google Scholar] [CrossRef] [Green Version]
- Javan, G.T.; Finley, S.J.; Can, I.; Wilkinson, J.E.; Hanson, J.D.; Tarone, A.M. Human Thanatomicrobiome Succession and Time Since Death. Sci. Rep. 2016, 6, 29598. [Google Scholar] [CrossRef] [Green Version]
- Heimesaat, M.M.; Boelke, S.; Fischer, A.; Haag, L.-M.; Loddenkemper, C.; Kühl, A.A.; Göbel, U.B.; Bereswill, S. Comprehensive Postmortem Analyses of Intestinal Microbiota Changes and Bacterial Translocation in Human Flora Associated Mice. PLoS ONE 2012, 7, e40758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaller, M.A.; Mujeeb, I.; Hollis, R.J.; Jones, R.N.; Doern, G.V. Evaluation of the discriminatory powers of the Dienes test and ribotyping as typing methods for Proteus mirabilis. J. Clin. Microbiol. 2000, 38, 1077–1080. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, K.E.; Lam, K.; Morgun, A.; Shulzhenko, N.; Löhr, C.V. Effect of temperature and time on the thanatomicrobiome of the cecum, ileum, kidney, and lung of domestic rabbits. J. Vet. Diagn. Investig. 2019, 31, 155–163. [Google Scholar] [CrossRef] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leonard, C.; Picavet, P.P.; Fontaine, J.; Clercx, C.; Taminiau, B.; Daube, G.; Claeys, S. The Middle Ear Microbiota in Healthy Dogs Is Similar to That of the External Ear Canal. Vet. Sci. 2023, 10, 216. https://doi.org/10.3390/vetsci10030216
Leonard C, Picavet PP, Fontaine J, Clercx C, Taminiau B, Daube G, Claeys S. The Middle Ear Microbiota in Healthy Dogs Is Similar to That of the External Ear Canal. Veterinary Sciences. 2023; 10(3):216. https://doi.org/10.3390/vetsci10030216
Chicago/Turabian StyleLeonard, Caroline, Pierre P. Picavet, Jacques Fontaine, Cécile Clercx, Bernard Taminiau, Georges Daube, and Stéphanie Claeys. 2023. "The Middle Ear Microbiota in Healthy Dogs Is Similar to That of the External Ear Canal" Veterinary Sciences 10, no. 3: 216. https://doi.org/10.3390/vetsci10030216