
Monitoring of Paenibacillus larvae in Lower Austria through DNA-Based Detection without De-Sporulation: 2018 to 2022

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
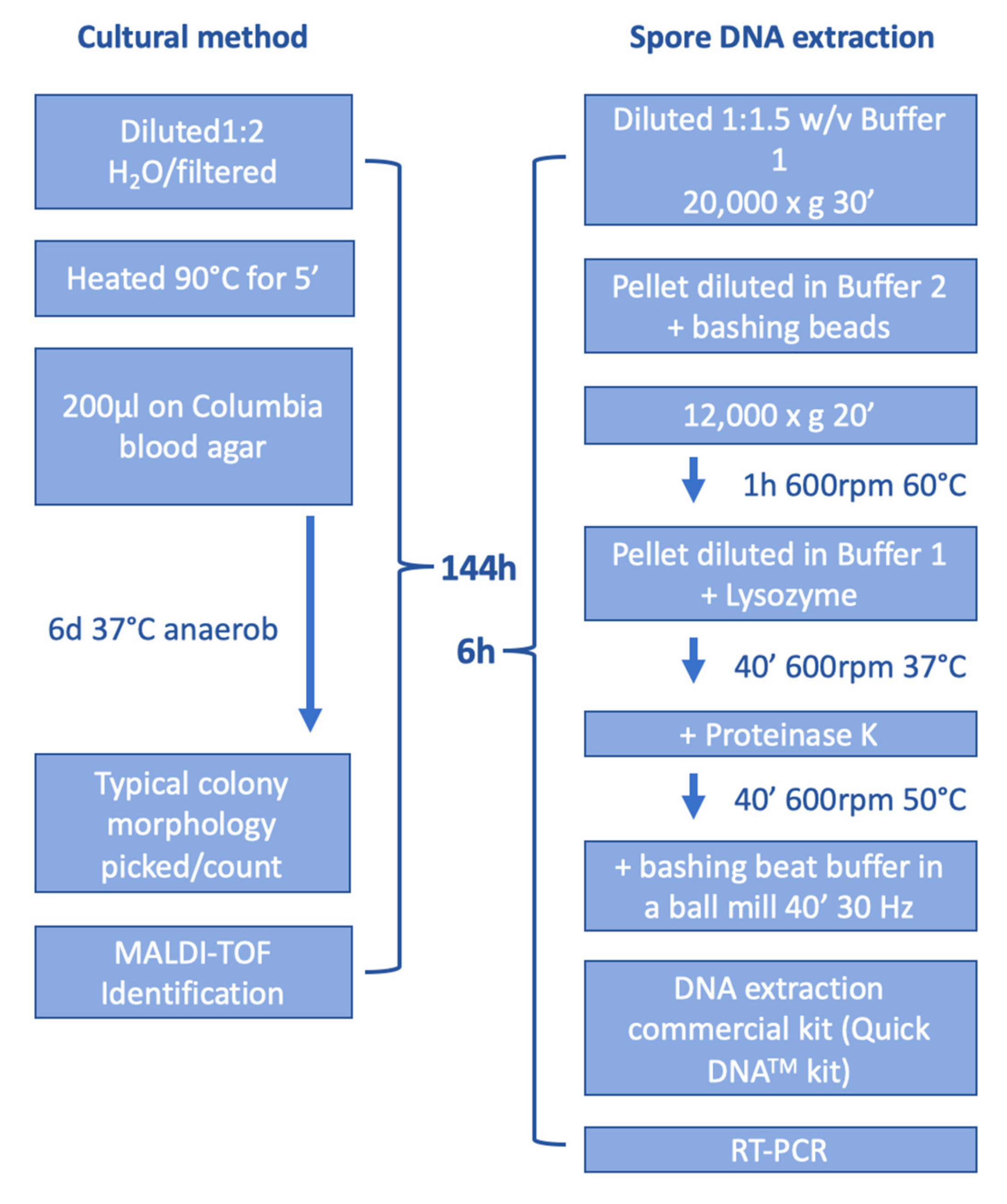
2. Materials and Methods
2.1. Beekeepers and Sampling under the Voluntary Monitoring Program
2.2. Cultural Detection of P. larvae
2.3. DNA Extraction
2.4. Real-Time-PCR (RT-PCR)
2.5. Classification of Results
2.6. Statistical Calculation of a CT-Value Using RT-PCR and Colony Counts and Sensitivity and Specificity
2.7. Prevalence of P. larvae
3. Results
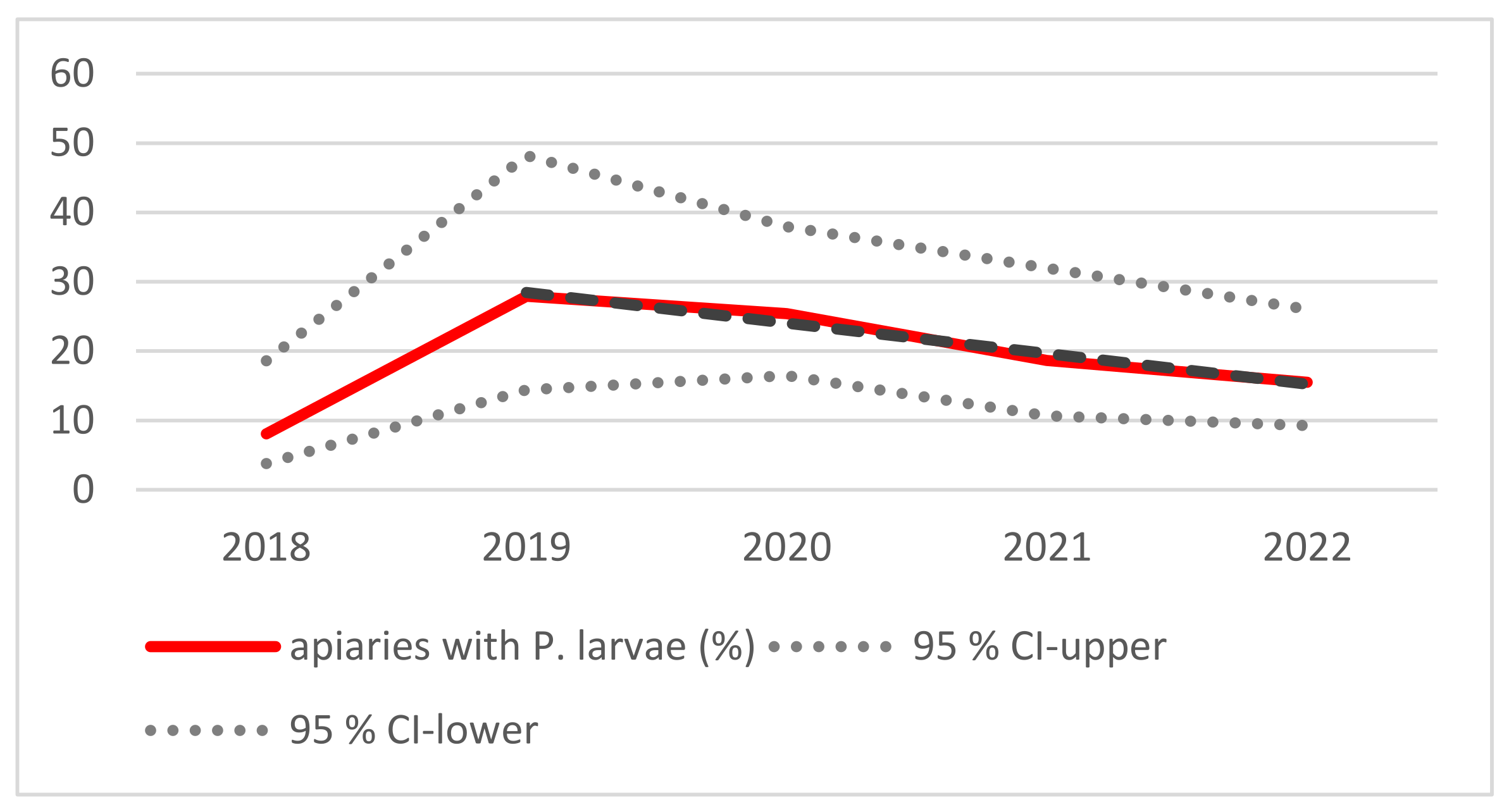
3.1. Monitoring
3.2. Sensitivity and Specificity of the RT-PCR Detection Method
3.3. Detection Limit with and without De-Coating
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ebeling, J.; Knispel, H.; Hertlein, G.; Fünfhaus, A.; Genersch, E. Biology of Paenibacillus larvae, a deadly pathogen of honey bee larvae. Appl. Microbiol. Biotechnol. 2016, 100, 7387–7395. [Google Scholar] [CrossRef]
- Poppinga, L.; Fünfhaus, A.; Aupperle, H.; Genersch, E. American foulbrood. In Diagnostic Colour Atlas of Bee Pathology, 1st ed.; Aupperle, H., Genersch, E., Eds.; Laboklin: Bad Kissingen, Germany, 2016; Volume 3, pp. 121–131. [Google Scholar]
- Riessberger-Gallé, U.; von der Ohe, W.; Crailsheim, K. Adult honeybee’s resistance against Paenibacillus larvae larvae, the causative agent of the American foulbrood. J. Invertebr. Pathol. 2001, 77, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, R.M.; Perry, J.H.; Houten, A.T. The effect of drifting honey bees on the spread of American foulbrood infections. J. Apic. Res. 1994, 33, 209–212. [Google Scholar] [CrossRef]
- Hornitzky, M.A.Z. The spread of Paenibacillus larvae subsp. larvae in an apiary. J. Apic. Res. 1998, 37, 261–265. [Google Scholar] [CrossRef]
- Lindström, A.; Korpela, S.; Fries, I. The distribution of Paenibacillus larvae spores in adult bees and honey and larval mortality, following the addition of American foulbrood diseased brood or spore-contaminated honey in honey bee (Apis mellifera) colonies. J. Invertebr. Pathol. 2008, 99, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Haseman, L. How long can spores of American foulbrood live? Am. Bee J. 1961, 101, 298–299. [Google Scholar]
- Fries, I.; Camazine, S. Implications of horizontal and vertical pathogen transmission for honey bee epidemiology. Apidologie 2001, 32, 199–214. [Google Scholar] [CrossRef] [Green Version]
- Gillard, M.; Charriere, J.D.; Belloy, L. Distribution of Paenibacillus larvae spores inside honey bee colonies and its relevance for diagnosis. J. Invertebr. Pathol. 2008, 99, 92–95. [Google Scholar] [CrossRef]
- De Graaf, D.C.; Alippi, A.M.; Brown, M.; Evans, J.D.; Feldlaufer, M.; Gregorc, A.; Hornitzky, M.; Pernal, S.F.; Schuch, D.M.; Titera, D.; et al. Diagnosis of American foulbrood in honey bees: A synthesis and proposed analytical protocols. Lett. Appl. Microbiol. 2006, 43, 583–590. [Google Scholar] [CrossRef] [Green Version]
- Locke, B.; Low, M.; Forsgren, E. An integrated management strategy to prevent outbreaks and eliminate infection pressure of American foulbrood disease in a commercial beekeeping operation. Prev. Vet. Med. 2019, 167, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Von der Ohe, W.; Dustmann, J.H. Efficient prophylactic measures against American Foulbrood by bacteriological analysis for spore contamination. Am. Bee J. 1997, 137, 603–606. [Google Scholar]
- Kušar, D.; Papić, B.; Zajc, U.; Zdovc, I.; Golob, M.; Žvokelj, L.; Knific, T.; Avberšek, J.; Ocepek, M.; Pislak Ocepek, M. Novel TaqMan PCR Assay for the Quantification of Paenibacillus larvae Spores in Bee-Related Samples. Insects 2021, 12, 1034. [Google Scholar] [CrossRef] [PubMed]
- Beims, H.; Janke, M.; von der Ohe, W.; Steinert, M. Rapid identification and genotyping of the honeybee pathogen Paenibacillus larvae by combining culturing and multiplex quantitative PCR. Open Vet. J. 2020, 10, 53–58. [Google Scholar] [CrossRef]
- D’Alessandro, B.; Antúnez, K.; Piccini, C.; Zumino, P. DNA extraction and PCR detection of Paenibacillus larvae spores from naturally contaminated honey and bees using spore-decoating and freeze-thawing techniques. World J. Microbiol. Biotechnol. 2007, 23, 593–597. [Google Scholar] [CrossRef]
- Cramer, J.S. The Origins of Logistic Regression, Tinbergen Institute Discussion Paper. 2002. TI 2002-119/4. Available online: https://doi.org/10.2139/ssrn.360300 (accessed on 8 March 2023).
- Sachs, L. Angewandte Statistik, 7th ed.; Springer: Berlin/Heidelberg, Germany, 1982; pp. 347–350, 433, 618–621. [Google Scholar]
- Zabrodski, M.W.; DeBruyne, J.E.; Wilson, G.; Moshynskyy, I.; Sharafi, M.; Wood, S.C.; Kozii, I.V.; Thebeau, J.; Klein, C.D.; Medici de Mattos, I.; et al. Comparison of individual hive and apiary-level sample types for spores of Paenibacillus larvae in Saskatchewan honey bee operations. PLoS ONE 2022, 17, e0263602. [Google Scholar] [CrossRef] [PubMed]
- Steinkraus, K.H.; Morse, R.A. American foulbrood incidence in some US and Canadian honeys. Apidiologie 1992, 23, 497–501. [Google Scholar] [CrossRef] [Green Version]
- Ebeling, J.; Reinecke, A.; Sibum, N.; Fünfhaus, A.; Aumeier, P.; Otten, C.; Genersch, E. A comparison of different matrices for the laboratory diagnosis of the epizootic American foulbrood of honey bees. Vet. Sci. 2023, 10, 103. [Google Scholar] [CrossRef]
- Bassi, S.; Galletti, G.; Carpana, E.; Palminteri, S.; Bosi, F.; Loglio, G.; Carra, E. Powdered Sugar Examination as a Tool for the Assessment of Paenibacillus larvae Infection Levels in Honey Bee Colonies. Front. Vet. Sci. 2022, 9, 853707. [Google Scholar] [CrossRef]
- Forsgren, E.; Laugen, A.T. Prognostic value of using bee and hive debris samples for the detection of American foulbrood disease in honey bee colonies. Apidologie 2014, 45, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Pohorecka, K.; Skubida, M.; Bober, A.; Zdańska, D. Screening of Paenibacillus larvae spores in apiaries from Eastern Poland. Nationwide survey. Part, I. Bull. Vet. Inst. Pulawy 2012, 56, 539–545. [Google Scholar] [CrossRef] [Green Version]
- Ribani, A.; Utzeri, V.J.; Taurisano, V.; Fontanesi, L. Honey as a Source of Environmental DNA for the Detection and Monitoring of Honey Bee Pathogens and Parasites. Vet. Sci. 2020, 7, 113. [Google Scholar] [CrossRef] [PubMed]
- Setlow, P. Observations on research with spores of Bacillales and Clostridiales species. J. Appl. Microbiol. 2019, 126, 348–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusenova, N.; Parvanova, P.; Stanilova, S. Detection of Paenibacillus larvae spores in honey by conventional PCR and its potential for American Foulbrood control. Bulg. J. Vet. Med. 2019, 22, 163–170. [Google Scholar] [CrossRef]
- Rossi, F.; Amadoro, C.; Ruberto, A.; Ricchiuti, L. Evaluation of Quantitative PCR (qPCR) Paenibacillus larvae Targeted Assays and Definition of Optimal Conditions for Its Detection/Quantification in Honey and Hive Debris. Insects 2018, 9, 165. [Google Scholar] [CrossRef] [Green Version]
- Bassi, S.; Carpana, E.; Bergomi, P.; Galetti, G. Detection and quantification of Paenibacillus larvae spores in samples of bees, honey and hive debris as a tool for American foulbrood risk assessment. Bull. Insectol. 2018, 71, 235–241. [Google Scholar]
- Crudele, S.; Ricchiuti, L.; Ruberto, A.; Rossi, F. Quantitative PCR (qPCR) vs culture-dependent detection to assess honey contamination by Paenibacillus larvae. J. Apic. Res. 2020, 59, 218–222. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| 2018 | 2019 | 2020 | 2021 | 2022 | |
|---|---|---|---|---|---|
| Sampling time | 21.03–12.04 * | 17.04–14.05 | 31.03–22.04 | 11.03–15.04 | 22.02–06.04 |
| Samples | 155 | 111 | 139 | 155 | 206 |
| Beekeepers | 62 | 43 | 63 | 59 | 71 |
| CT-Value Range | Number of Samples | Culture Positive % |
|---|---|---|
| (23.0–31.2) | 9 | 100 |
| (31.2–33.9) | 10 | 100 |
| (33.9–35.9) | 8 | 62.5 |
| (35.9–37.9) | 7 | 28.6 |
| (37.9–38.6) | 7 | 14.3 |
| (38.6–43.2) | 8 | 25.0 |
| (45) | 31 | 0.0 |
| Year | Number of Samples | ||
|---|---|---|---|
| No Spores | Low Spore Content | High Spore Content | |
| 2018 | 143 | 4 | 8 |
| 2019 | 98 | 13 | 0 |
| 2020 | 116 | 20 | 3 |
| 2021 | 146 | 18 | 1 |
| 2022 | 206 | 12 | 5 |
| CT-Value | Sensitivity | Specificity | ||||
|---|---|---|---|---|---|---|
| Estimated Value | Lower Limit | Upper Limit | Estimated Value | Lower Limit | Upper Limit | |
| 35 | 0.793 | 0.655 | 0.931 | 1.000 | 1.000 | 1.000 |
| 36 | 0.828 | 0.690 | 0.966 | 0.941 | 0.863 | 1.000 |
| 37 | 0.862 | 0.724 | 0.966 | 0.902 | 0.823 | 0.980 |
| 38 | 0.897 | 0.759 | 1.000 | 0.824 | 0.706 | 0.922 |
| 39 | 0.931 | 0.828 | 1.000 | 0.725 | 0.588 | 0.843 |
| Sample | Colony Count | CT-Value Using De-Coating | CT-Value without De-Coating |
|---|---|---|---|
| 3795/1 | 10 | 31.2 | 29.8 |
| 3795/2 | 2 | 33.9 | 34.2 |
| 4/22 | 0 | 35.3 | 35.3 |
| 32/22 | 53 | 26.3 | 26.5 |
| 57/22 | 0 | 37.9 | 39.2 |
| 58/22 | 2 | 34.0 | 33.0 |
| 60/22 | 14 | 30.6 | 32.3 |
| 185/22 | 1 | 33.9 | 32.8 |
| 3796/2 | 2 | 33.8 | 32.1 |
| RSK-16-07-21 | 26 | 31.0 | 40.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilhelm, E.; Korschineck, I.; Sigmund, M.; Paulsen, P.; Hilbert, F.; Rossmanith, W. Monitoring of Paenibacillus larvae in Lower Austria through DNA-Based Detection without De-Sporulation: 2018 to 2022. Vet. Sci. 2023, 10, 213. https://doi.org/10.3390/vetsci10030213
Wilhelm E, Korschineck I, Sigmund M, Paulsen P, Hilbert F, Rossmanith W. Monitoring of Paenibacillus larvae in Lower Austria through DNA-Based Detection without De-Sporulation: 2018 to 2022. Veterinary Sciences. 2023; 10(3):213. https://doi.org/10.3390/vetsci10030213
Chicago/Turabian StyleWilhelm, Elfriede, Irina Korschineck, Michael Sigmund, Peter Paulsen, Friederike Hilbert, and Wigbert Rossmanith. 2023. "Monitoring of Paenibacillus larvae in Lower Austria through DNA-Based Detection without De-Sporulation: 2018 to 2022" Veterinary Sciences 10, no. 3: 213. https://doi.org/10.3390/vetsci10030213