
Dataset for Spectroscopic, Structural and Dynamic Analysis of Human Fe(II)/2OG-Dependent Dioxygenase ALKBH3
 , and
, and
Abstract
:1. Summary
2. Data Description
2.1. Visualization of the ALKBH3–DNA Complex
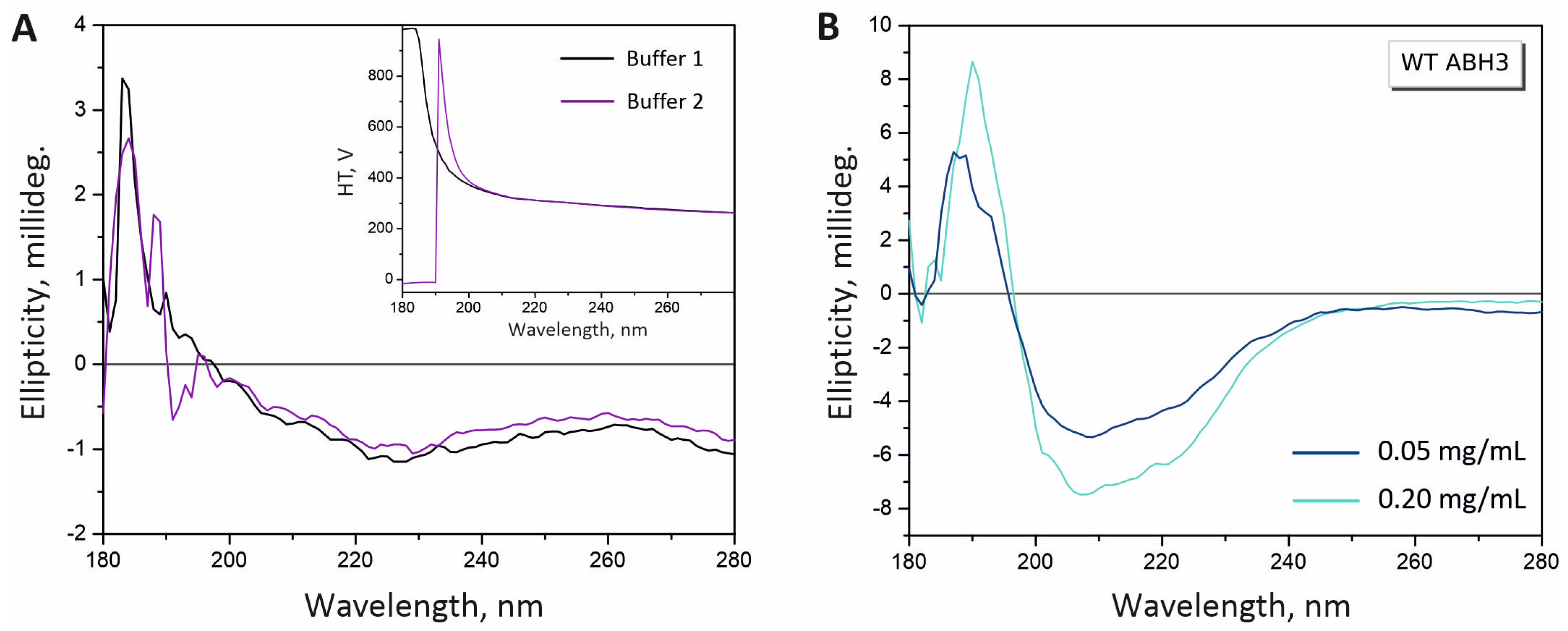
2.2. Optimization of Experimental Conditions for CD Measurements
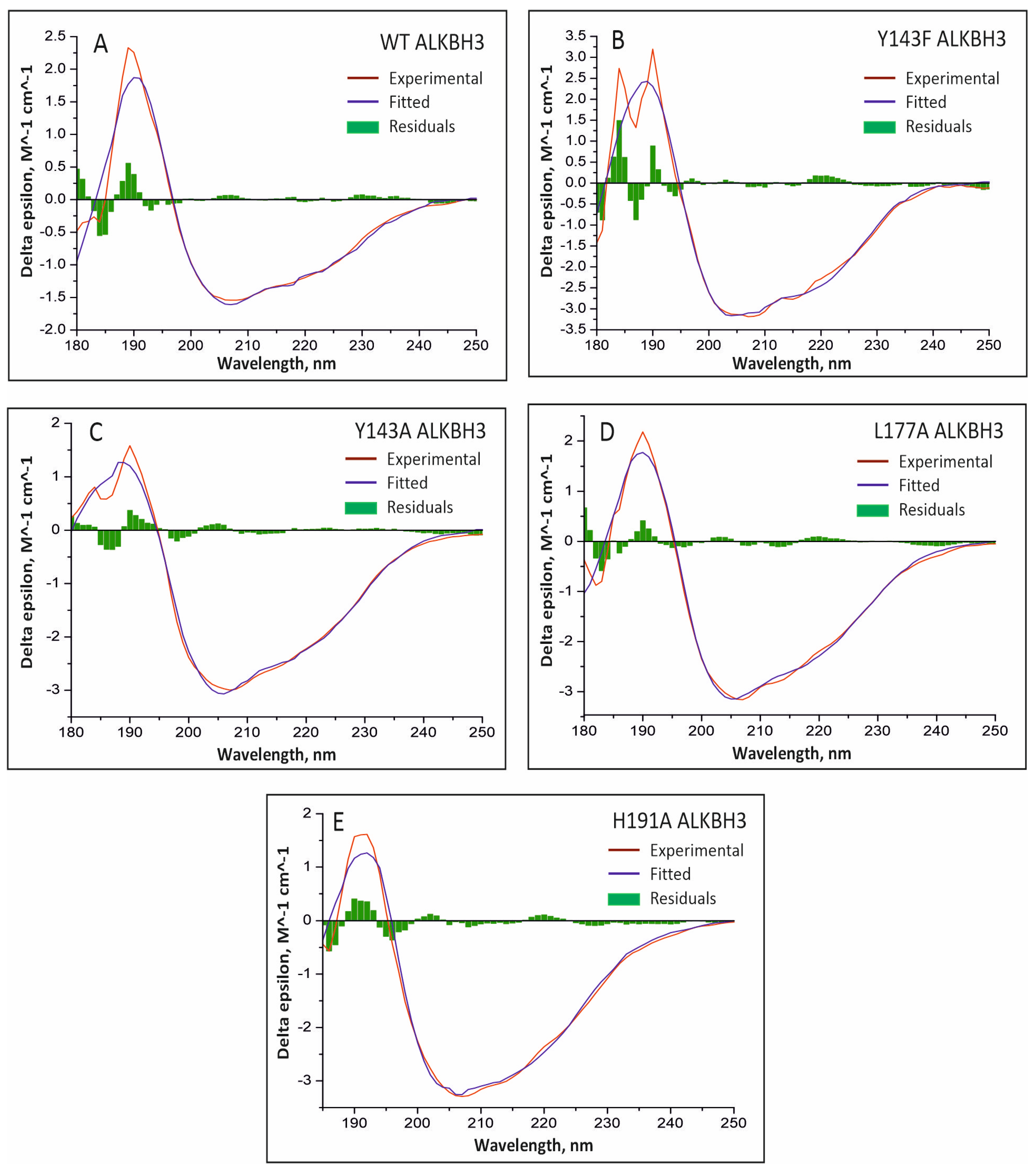
2.3. Deconvolution of the Individual CD Spectrum
2.4. Measuring the Melting of the ALKBH3 Protein Globule by CD Spectroscopy
3. Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fedeles, B.I.; Singh, V.; Delaney, J.C.; Li, D.; Essigmann, J.M. The AlkB Family of Fe(II)/alpha-Ketoglutarate-dependent Dioxygenases: Repairing Nucleic Acid Alkylation Damage and Beyond. J. Biol. Chem. 2015, 290, 20734–20742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aas, P.A.; Otterlei, M.; Falnes, P.O.; Vagbo, C.B.; Skorpen, F.; Akbari, M.; Sundheim, O.; Bjoras, M.; Slupphaug, G.; Seeberg, E.; et al. Human and bacterial oxidative demethylases repair alkylation damage in both RNA and DNA. Nature 2003, 421, 859–863. [Google Scholar] [CrossRef] [PubMed]
- Tasaki, M.; Shimada, K.; Kimura, H.; Tsujikawa, K.; Konishi, N. ALKBH3, a human AlkB homologue, contributes to cell survival in human non-small-cell lung cancer. Br. J. Cancer 2011, 104, 700–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimada, K.; Fujii, T.; Tsujikawa, K.; Anai, S.; Fujimoto, K.; Konishi, N. ALKBH3 contributes to survival and angiogenesis of human urothelial carcinoma cells through NADPH oxidase and tweak/Fn14/VEGF signals. Clin. Cancer Res. 2012, 18, 5247–5255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, N.R.; McDonough, M.A.; King, O.N.; Kawamura, A.; Schofield, C.J. Inhibition of 2-oxoglutarate dependent oxygenases. Chem. Soc. Rev. 2011, 40, 4364–4397. [Google Scholar] [CrossRef] [PubMed]
- Sundheim, O.; Vagbo, C.B.; Bjoras, M.; Sousa, M.M.; Talstad, V.; Aas, P.A.; Drablos, F.; Krokan, H.E.; Tainer, J.A.; Slupphaug, G. Human ABH3 structure and key residues for oxidative demethylation to reverse DNA/RNA damage. EMBO J. 2006, 25, 3389–3397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sreerama, N.; Woody, R.W. Estimation of protein secondary structure from circular dichroism spectra: Comparison of CONTIN, SELCON, and CDSSTR methods with an expanded reference set. Anal. Biochem. 2000, 287, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Micsonai, A.; Wien, F.; Kernya, L.; Lee, Y.H.; Goto, Y.; Refregiers, M.; Kardos, J. Accurate secondary structure prediction and fold recognition for circular dichroism spectroscopy. Proc. Natl. Acad. Sci. USA 2015, 112, E3095–E3103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bleijlevens, B.; Shivarattan, T.; van den Boom, K.S.; de Haan, A.; van der Zwan, G.; Simpson, P.J.; Matthews, S.J. Changes in protein dynamics of the DNA repair dioxygenase AlkB upon binding of Fe(2+) and 2-oxoglutarate. Biochemistry 2012, 51, 3334–3341. [Google Scholar] [CrossRef] [PubMed]
- Dunwell, J.M.; Purvis, A.; Khuri, S. Cupins: The most functionally diverse protein superfamily? Phytochemistry 2004, 65, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Kuwajima, K. The molten globule state as a clue for understanding the folding and cooperativity of globular-protein structure. Proteins 1989, 6, 87–103. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Protein | Wavelength Range, nm | Regular α-Helix | Distorted α-Helix | Antiparallel β-Strand | Parallel β-Strand | Turns | Other | NRMSD |
|---|---|---|---|---|---|---|---|---|
| WT ALKBH3 | 180–250 | 0.05 | 0.01 | 0.33 | 0 | 0.14 | 0.47 | 0.049 |
| 185–250 | 0.04 | 0 | 0.41 | 0 | 0.12 | 0.43 | 0.031 | |
| Y143F ALKBH3 | 180–250 | 0.08 | 0.03 | 0.31 | 0 | 0.12 | 0.46 | 0.048 |
| 185–250 | 0.12 | 0.04 | 0.27 | 0.11 | 0.14 | 0.42 | 0.029 | |
| Y143A ALKBH3 | 180–250 | 0.09 | 0.06 | 0.17 | 0 | 0.19 | 0.49 | 0.027 |
| 185–250 | 0.12 | 0.07 | 0.16 | 0.06 | 0.14 | 0.46 | 0.017 | |
| L177A ALKBH3 | 180–250 | 0.08 | 0.06 | 0.21 | 0 | 0.17 | 0.48 | 0.055 |
| 185–250 | 0.09 | 0.06 | 0.20 | 0.06 | 0.12 | 0.47 | 0.022 | |
| H191A ALKBH3 | 180–250 | 0.08 | 0.08 | 0.16 | 0.11 | 0.17 | 0.50 | 0.061 |
| 185–250 | 0.08 | 0.06 | 0.16 | 0.08 | 0.12 | 0.49 | 0.030 |
| Protein | Regular α-Helix | Distorted α-Helix | Regular β-Strand | Distorted β-Strand | Turns | Unordered | NRMSD |
|---|---|---|---|---|---|---|---|
| WT ALKBH3 | 0.10 | 0.4 | 0.22 | 0.14 | 0.24 | 0.35 | 0.065 |
| Y143F ALKBH3 | 0.4 | 0.4 | 0.19 | 0.15 | 0.21 | 0.38 | 0.027 |
| Y143A ALKBH3 | 0.9 | 0.10 | 0.14 | 0.11 | 0.24 | 0.33 | 0.052 |
| L177A ALKBH3 | 0.9 | 0.10 | 0.14 | 0.11 | 0.24 | 0.32 | 0.057 |
| H191A ALKBH3 | 0.11 | 0.11 | 0.11 | 0.10 | 0.26 | 0.32 | 0.038 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanazhevskaya, L.Y.; Gorbunov, A.A.; Zhdanova, P.V.; Koval, V.V. Dataset for Spectroscopic, Structural and Dynamic Analysis of Human Fe(II)/2OG-Dependent Dioxygenase ALKBH3. Data 2023, 8, 57. https://doi.org/10.3390/data8030057
Kanazhevskaya LY, Gorbunov AA, Zhdanova PV, Koval VV. Dataset for Spectroscopic, Structural and Dynamic Analysis of Human Fe(II)/2OG-Dependent Dioxygenase ALKBH3. Data. 2023; 8(3):57. https://doi.org/10.3390/data8030057
Chicago/Turabian StyleKanazhevskaya, Lyubov Yu., Alexey A. Gorbunov, Polina V. Zhdanova, and Vladimir V. Koval. 2023. "Dataset for Spectroscopic, Structural and Dynamic Analysis of Human Fe(II)/2OG-Dependent Dioxygenase ALKBH3" Data 8, no. 3: 57. https://doi.org/10.3390/data8030057