
Evaluating the Role of Mashing in the Amino Acid Profiles of Worts Produced from Gluten-Free Malts



Department of Food Science, College of Agricultural Sciences, The Pennsylvania State University, University Park, PA 16802, USA
*
Author to whom correspondence should be addressed.
Beverages 2023, 9(1), 10; https://doi.org/10.3390/beverages9010010
Submission received: 28 October 2022
/
Revised: 28 December 2022
/
Accepted: 18 January 2023
/
Published: 28 January 2023
(This article belongs to the Special Issue Brewing Technology – Innovations in Raw Materials, Processing and Products)
Abstract
:A successful wort fermentation depends on both the sugar and the free amino nitrogen (FAN) content of a wort. The primary goal of the mashing step is to generate fermentable sugars, as FAN is regarded as being primarily determined by malt quality; however, the role of mashing in modifying FAN has not been extensively studied, especially with respect to non-barley brewing materials. In this study, the FAN content of gluten-free (GF) worts varied greatly from barley (73–490 mg/L vs. 201 mg/L, respectively) and yielded different amino acid profiles, including lower proline and higher γ-aminobutyric acid concentrations. While most of the amino acids were present in the malt or generated in a brief window early in the mashing, significant increases in amino acid concentrations could be generated by mashing at temperatures below 55 °C. Overall, GF malts are promising brewing ingredients that can produce quality worts if appropriate mashing conditions are implemented.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
The primary purpose of the mashing step in brewing is to generate small yeast-assimilable sugars and nitrogen sources into the wort to support healthy fermentation. During mashing, the brewer is generally most concerned with maximizing the endogenous malt amylase activities to break down starch into high concentrations of fermentable sugars (FS), as in the wort, the concentration of FS (~90–100 g/L) will be higher than the total concentration of amino acids (~1–2 g/L) [1,2]. Values for free amino nitrogen (FAN) in worts ranging approximately from 150 to 200 mg/L are considered sufficient, although the optimum amount of FAN in any wort will be dependent on the yeast being utilized, wort sugar concentrations, and other process variables [3,4,5]. The standard method for measuring FAN, the ninhydrin method, measures the concentration of nitrogen derived from amines in the wort (primarily α-amino acids), but does not provide information about the concentration of individual amino acids. For a brewer utilizing standard malts, the relative abundance of each amino acid is assumed to be stable such that differences in the FAN across samples would relate to differences in the total amino nitrogen content rather than differences in the underlying amino acid distribution. However, this assumption may not hold if a brewer utilizes different brewing ingredients and brewing procedures [6]. The concentrations of the individual amino acids are important because the carbon skeletons of some amino acids can be converted into fusel alcohols and acetates via the Ehrlich pathway, directly impacting the flavor profiles of the beer [3,4,7,8,9,10]. Furthermore, valine concentrations can affect diacetyl production, and the dynamic yeast metabolic pathways can change fermentation and flavor characteristics if the nutrients in the wort are changed [5,11]. Therefore, it is important to understand both the total amino nitrogen content (as measured by FAN) and the underlying amino acid concentrations in a wort.
While “gluten-free” beers are often produced from malted barley worts using enzymatic treatments with Aspergillus niger prolyl-endopeptidase, these treatments do not actually remove all risk of gluten peptides persisting in the beer, nor assess the potential immunogenicity of all the peptide hydrolysates formed by its action, nor the limitations in the most common ELISA-based assays used for gluten detection, and thus in the USA, beers produced in this manner cannot be labelled gluten-free [12,13,14,15,16,17]. Brewing a true gluten-free beer then necessitates the utilization of alternative brewing ingredients that do not contain gluten and alternative processes to better utilize these ingredients. Previous research in GF brewing has shown that GF worts can be successfully prepared from GF malts with comparable amounts of FAN to a typical barley wort with differing individual amino acid concentrations, but with procedures that produce sugar profiles different from barley [18,19]. We have previously demonstrated a modified brewing framework (ExGM decoction) that can successfully utilize various GF grains to produce high concentrations of FS with the desired maltose-dominated sugar profile [20]. However, the amino acid and FAN contents of those worts were not investigated, and so the impact of the ExGM framework on these parameters was unknown.
In general, there is no consensus on the amount of wort nitrogen that can be generated during the mash as it is process-dependent, but it is known that much of the extracted amino acids are derived from proteolytic activity during malting and not a direct result from mashing [9,21,22,23,24,25]. A step mash procedure generally includes a “protein rest” step at 45–55 °C for typically ~30 min to improve protein degradation, as the proteolytic enzymes are generally regarded as thermolabile at the temperatures typically used to maximize amylase activity for FS production (≥62 °C), while an infusion mash at ~65 °C bypasses a protein rest altogether [19,22,25,26]. If brewing with a GF malt, modified procedures are already required to improve sugar generation, and the ExGM process we described previously utilized mashing temperatures (55 °C) that are closer to the range of temperatures used for a protein rest; therefore, it was hypothesized that an extended time mash at this lower temperature could be an avenue to improve the amino acid content of a wort, if increasing the amino acid content of a wort is necessary, without requiring a brewer to utilize exogenous sources of nitrogen as a supplementary ingredient.
The purpose of this work was to understand the concentration of amino acids that could be generated from several GF malts using the ExGM decoction, and how the mash could be manipulated to increase the amino acid content of a millet wort. A previous analysis of the fermentable sugar concentrations of the various GF malt and millet malt worts discussed here contained sugar concentrations in the range expected for malted barley worts (~90–100 g/L) [20,27]. Therefore, given sufficient fermentable sugar generation in these GF mashes, the amino acid content of these resulting worts in relation to benchmarks targeted in traditional brewing and the role of mashing in amino acid production was investigated. The generation of amino acids from millet across mashes with different mashing temperatures, grind regimes, or exogenous enzymes was investigated, to develop a strategy to improve the amino acid content of the wort solely during the mash. The purpose of this work was to better understand the role of the mashing process in the generation of amino acids from GF malts, so that a brewer can exert more control over both total FAN levels and amino acid profiles to produce worts with the desired characteristics.
2. Materials and Methods
2.1. Materials
Mixed amino acid standard solutions, supplementary amino acid standards, 0.4 N Borate Buffer (pH 10.2), concentrated o-pthalaldehyde (OPA) reagent, and concentrated 9-fluorenylmethylchloroformate (FMOC) reagent were purchased from Agilent Technologies (Santa Clara, CA, USA). Bovine serum albumin (BSA), ethylenediaminetetraacetic acid (EDTA), maleic acid, malic acid, p-nitrophenol, glycine > 99%, ninhydrin reagent, γ-aminobutyric acid (GABA) ≥ 99%, potassium iodate > 99%, sodium tetraborate decahydrate ≥ 99.5%, sodium phosphate dibasic ≥ 99%, potassium phosphate monobasic, and HPLC-grade acetonitrile were purchased from Sigma-Aldrich (St. Louis, MO, USA). HPLC-grade methanol, calcium chloride dihydrate, and tris(hydroxymethyl)aminomethane were purchased from Fisher Scientific (Hampton, NH, USA). Fructose was purchased from Alfa Aesar (Heysham, England), 88% lactic acid was purchased from LD Carlson Co. (Kent, OH, USA), and 200 proof ethanol was purchased from VWR International (Radnor, PA, USA).
2.2. Malt Preparation
GF malts and barley malts were produced as previously described [20] or purchased from their respective suppliers and were ground to pass a 200 µm screen using a Retsch SR200 Hammer Mill (Retsch GmbH, Haan, Germany). Two lots of millet malts (Grouse Malt House, Wellington, CO, USA) were purchased commercially and either ground to pass a 200 µm screen (fine grind) using a Retsch SR200 Hammer Mill or ground to pass a 381 µm roller gap (coarse grind) using a Monster Mills MM-2Pro Gluten-Free Malt Edition 2-roller mill (Monster Brewing Hardware, Fayetteville, GA, USA) for use in the mashing experiments described below. These millet malts were the same commercial malt product from the sample supplier purchased at different times, to serve as an investigation into effects that may primarily be due to variation from malting. Ceremix® Flex and Ondea® Pro were purchased from Novozymes (Bagsværd, Denmark).
2.3. GF Wort and Barley Wort Sample Generation
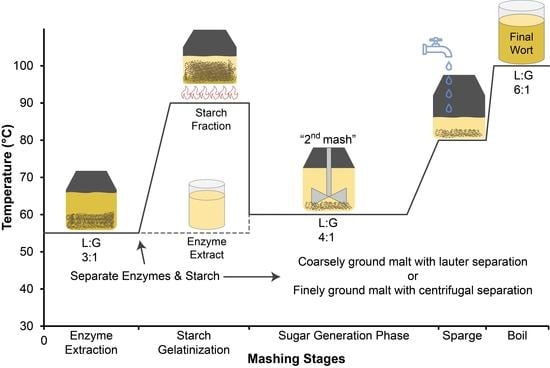
In this work, the mashing procedures generally followed the ExGM protocol (Extract, Gelatinize, Mash) that was developed previously for gluten-free brewing that allows for the production of high levels of FS from a variety of gluten-free grains [20,27]. In general, the ExGM protocol consists of a “first mash” at a 3:1 liquor to grist ratio (L:G) that serves as an enzyme extraction step where the milled malt is held at a temperature where the endogenous amylolytic enzymes are stable (typically 55 °C for 30 min, but conditions varied in some experiments below). In the case of finely ground malts, the extracted enzymes are then separated from the starch and other insoluble material by centrifugation. For the coarsely ground malts, this separation is achieved by lautering. An additional equivalent of water is then added to the starch fraction, and it is brought to a high enough temperature to gelatinize the starch (typically 90 °C). This is cooled and then recombined with the enzyme extract, and the ”second mash” is performed at a now 4:1 L:G and a temperature that maintains the activity of all of the endogenous amylolytic enzymes (typically 55 °C for 2 h, but conditions varied in some experiments below). This is followed by a high temperature mash out and then the wort being separated from the grain bed by centrifugation in the case of the finely milled malts, and by lautering in the case of the coarsely ground malts. Several variations on this procedure were used and described below. Note that the first and second mashes refer to the sequential manipulations to the same material, not two unique mashes.
Samples of previously generated and frozen worts from barley, millet, sorghum, ivory teff, brown teff, rice, corn, or buckwheat [20] were analyzed for their FAN and amino acid contents. The malted barley control used for these experiments was a recipe of 80% 2-row base malt and 20% caramel-150 malt and was mashed using a simple 1 h infusion mash at 65 °C to model a commercial recipe. The millet, sorghum, ivory teff, brown teff, GFB1 (a recipe of 60% millet, 25% rice, 15% buckwheat), rice, corn, and buckwheat samples were all produced from malts of those GF grains by following the ExGM procedure described above for finely ground malts. The XT samples were produced from millet malt without a gelatinization step, and an isothermal infusion mash at 55 °C for 2 h and 45 min (approximately the same total process time as the ExGM procedure). The IE samples, also from millet malt, were initially mashed at 65 °C, immediately placed in a boiling water bath to increase the mash temperature to 99 °C and rapidly denature the endogenous enzyme activity to estimate the freely extractable FS and amino acid concentrations from millet. All mashes were performed in duplicate.
2.4. Millet Time-Course Mashing Experiments
To evaluate the effect of different malt grinding regimes and different temperatures during the second mash on the generation of FS and amino acids, experimental mashes were produced using two generalized mashing procedures: mashes prepared from finely ground millet and centrifugal separation (FF), and mashes prepared from coarsely ground millet malt and lauter separation (CL). All mashes were performed in duplicate with 20 g or 30 g of malt. The mashes all employed the same mashing water (0.7 mM calcium chloride in 18.2 MΩ cm resistivity water), were pH-adjusted to ~5.3 with 0.5 M lactic acid, mashed at an initial liquor-to-grist ratio (L:G) of 3 mL water: 1 g malt (3:1). The first mash was held at 55 °C for 30 min, after which the enzyme extract was separated, and the separated enzyme extract was held at 55 °C awaiting the start of the second mash. An additional equivalent of water was added to the separated starch to ensure enough water for gelatinization and then brought to 90 °C on a hot plate. The gelatinized starch was cooled in an ice bath to the desired second mash temperature (55, 60, 65, or 70 °C), and the enzyme extract was reintroduced to begin the second mash (second mash L:G 4:1). At the end of the second mash, the mashes were brought to 85 °C, the first wort was separated, and then the sparge was conducted in 2 equal batches (for a constant final L:G of 6:1) held at 80 °C: the first for 10 min and the second for 5 min. All wort fractions were combined after sparging and boiled for 90 s, then the wort was transferred to 50 mL sterile centrifuge tubes, cooled in an ice bath, and stored in a −80 °C freezer prior to analysis.
The coarse grind and lauter approach samples (CL and CL-SR) both utilized a 60 °C second mash temperature. A variant of the CL base procedure, coarse and lauter with starch recovery (CL-SR) utilized a centrifugation step on the lautered enzyme extract to recover the starch retained in the enzyme extract and return that starch to the gelatinization step; otherwise, the retained starch would not be gelatinized, thus limiting the FS generation. Similarly, another variant of the CL protocol utilized exogenous enzyme additions (CL-Exo) of Ondea® Pro (8 µL during the enzyme extraction) and of Ceremix® Flex (8 µL during the enzyme extraction, and an additional 8 µL to the lautered enzyme extract) with a modified step mash procedure for the second mash of 1 h at 60 °C, a ramp to 80 °C, and held for 1 h prior to the final temperature ramp to end the mash and begin the sparge. The exogenous enzyme doses were based on the dosing rate recommendations of the manufacturer adjusted to scale.
To assess the change in amino acid concentrations across the second mash, aliquots of the mash were taken at various timepoints during the second mash, dispensed into 1.5 mL microcentrifuge tubes, and placed in a 99 °C block heater for 5 min to denature any remaining enzyme activity. After the incubation, aliquots were cooled in an ice bath and stored at −80 °C prior to the analysis. A more detailed and thorough discussion of the rationale and procedure for the generation of these samples can be found in our work describing their fermentable sugar concentrations [27]. Samples were labelled regarding the base mashing approach (FF or CL, as described above) with any additional modifications. For example, FF-65 °C refers to the millet samples produced using finely ground malt, centrifugal separation, and a second mash at 65 °C, and CL-Exo refers to samples produced using coarsely ground malt, lauter separation, and a step mash with added exogenous enzymes.
2.5. Lower Temperature Enzyme Extractions
To assess the potential of increasing the amino acid generation during the first mash, additional experiments were performed with the CL approach where the first mash was modified to 120 min at 40 °C either with only the malt (CL-40 °C) or supplemented with Ondea® Pro (CL-Exo 40 °C). Samples of the enzyme extractions were taken at 30-, 60-, 90-, and 120 min; placed in 1.5 mL microcentrifuge tubes; and incubated for 5 min at 99 °C to denature the enzyme activity.
Additional samples from CL-40 °C were taken at 30-, 60-, 90-, and 120 min to assess the effect of the reduced temperature and increased time (compared to the first mash at 55 °C for 30 min described above) on the extraction of the amylolytic enzymes β-amylase, α-amylase, and limit dextrinase, the main starch degrading enzymes in malt responsible for the generation of FS. Measurements of the extracted enzyme activity were taken using the modified Betamyl-3, Ceralpha, and PullG6 methods for β-amylase, α-amylase, and limit dextrinase, respectively, as previously described [20,28,29]. Aliquots from the mash were taken and centrifuged at 5000× g (Microfuge 16, Beckman Coulter, Brea, CA) for 5 min to clarify the extract, then 100 µL of the mash extract was diluted with 400 µL of a pH 5.5 100 mM maleic acid solution with 1 mg/mL added BSA (LD reaction buffer), mixed, then added to a 500 µL 10 kDa molecular weight cut-off centrifugal filter and centrifuged for 10 min at 14,000× g. The filtrate was discarded, and an additional 400 µL of the LD reaction buffer was added to the filter and centrifuged again for 10 min at 14,000× g. The process was repeated twice more to wash the mash enzyme extract of natural substrates that would otherwise interfere with the digestion of the synthetic substrates in the enzymatic assays. After the washes, the retentate was diluted in the appropriate LD reaction buffer, β-amylase reaction buffer (50 mM malic acid, 50 mM maleic acid, 1 mg/mL BSA, and 1mM EDTA pH 5.4), or α-amylase reaction buffer (25 mM malic acid, 25 mM maleic acid, and 2 mM calcium chloride pH 5.4) and then the enzymatic analyses were performed in triplicate, and these lower temperature mashes were performed in duplicate. The units of enzyme activity were described as in the assay kit instructions from the specified substrate for β-amylase (BU), α-amylase (CU), or limit dextrinase (PU).
2.6. FAN and Soluble Nitrogen Analysis
For the free amino nitrogen analysis, samples of retained and frozen worts were thawed, vortexed to re-homogenize, then centrifuged for 10 min at 14,000× g to standardize the clarity of the wort, as the different grind and separation approaches led to the samples with differing clarities and different amounts of insoluble or suspended materials. The FAN concentrations for each wort were then analyzed using the ninhydrin method in the ASBC official methods of analysis using glycine as the nitrogen standard [30]. The FAN assays of each mash were performed in triplicate.
For the soluble nitrogen analysis, samples were combusted using a LECO FP-528 Nitrogen Analyzer (LECO Corporation, St. Joseph, MI, USA) and the manufacturer’s recommended method for analyzing nitrogen in wort and beer using glycine as the nitrogen standard [31]. Prior to combustion, samples were centrifuged for 10 min at 3220× g (Centrifuge 5810 R, A-4-62 Rotor, Eppendorf, Hamburg, Germany) to initially clarify the wort, and were then filtered through 0.45 µm nylon syringe filters. As the lautered samples contained more suspended insoluble material than the samples that had been centrifuged throughout the mashing process, all samples were clarified by centrifugation and filtration to limit the possibility of wort clarity being a confounding factor in the soluble nitrogen (SN) analysis. Samples from each mash were analyzed in triplicate and expressed as % nitrogen by mass.
2.7. Amino Acid Analysis
Samples or frozen wort for the amino acid analysis were thawed, vortexed, and centrifuged for 10 min at 14,000× g. An aliquot of the clarified wort (100 µL) was then mixed with 100 µL of a solution of the internal standards norvaline and sarcosine into a 1.5 mL microcentrifuge tube. These mixed worts were then deproteinized by adding 600 µL of HPLC-grade methanol, vortexing, and resting in an ice bath for 30 min. The samples were then centrifuged for 10 min at 14,000× g, and aliquots were further diluted with ultrapure water and filtered through 0.45 µm nylon syringe filters.
Amino acids were analyzed via HPLC separation and fluorescence detection of OPA- and FMOC-derivatized amino acids using an Agilent 1100 Series HPLC system with a quaternary pump, autosampler, and fluorescence detector (FLD) (Agilent Technologies, Santa Clara, CA, USA). Amino acids were derivatized in the injection loop and separated using InfinityLab Poroshell HPH-C18 guard (2.7 µm, 4.6 × 5 mm) and analytical (2.7 µm, 4.6 × 100 mm) columns held at 40 °C (Agilent Technologies, Santa Clara, CA, USA). The injection program, FLD settings, and gradient conditions were based on the method literature from the manufacturer [32,33,34], and were slightly modified to adapt newer column technology with enhanced resistance to the basic conditions of the separation with a lower maximum operating pressure HPLC system. The mobile phases consisted of 10 mM Na2HPO4 and 10 mM Na2B4O7 pH 8.2 ± 0.02 (A) (0.45 µm regenerated cellulose filtered) and 45:45:10 (v:v:v) acetonitrile:methanol:water (B). The gradient program to separate the derivatized amino acids at a flow rate of 1.5 mL/min was as follows: 0–0.35 min, 2% B; 0.35–16.40 min, 2% to 57% B; 16.40–16.50 min, 57% to 100% B; 16.5–22.7 min 100% B; 22.70 min-22.80 min 100% to 2% B; 22.80–22.90 min, 2% B. Detection and quantification of the OPA- and FMOC-derivatized amino acids were achieved using an FLD set for a 340 nm excitation and 450 nm emission (OPA amino acids), with a programmed switch at 12.15 min to an excitation of 260 nm and an emission of 325 nm (FMOC amino acids). Amino acids were quantified based on a standard curve of a mixed solution of the amino acid standards containing 25 µM each of the internal standards norvaline and sarcosine. A standard curve of GABA containing the internal standards was prepared separately. Amino acids were analyzed with duplicate injections of the duplicate mashes.
2.8. Statistical Analysis
Statistical analyses were performed using Minitab 21 (Minitab LLC, State College, PA, USA), and p-values < 0.05 were considered statistically significant. Analyses of the GF worts, the 40 °C enzyme extractions, the amino acid profiles of millet worts compared to barley, and the individual time courses were performed using a one-way ANOVA, with Tukey’s honestly significant difference post-hoc test where appropriate. Analyses of the millet extracts, and endpoint FAN, soluble nitrogen, and amino acid groups were performed using a general linear model ANOVA with the mashing variables of interest as nested terms.
3. Results and Discussion
3.1. FAN and Amino Acid Profiles of GF Worts
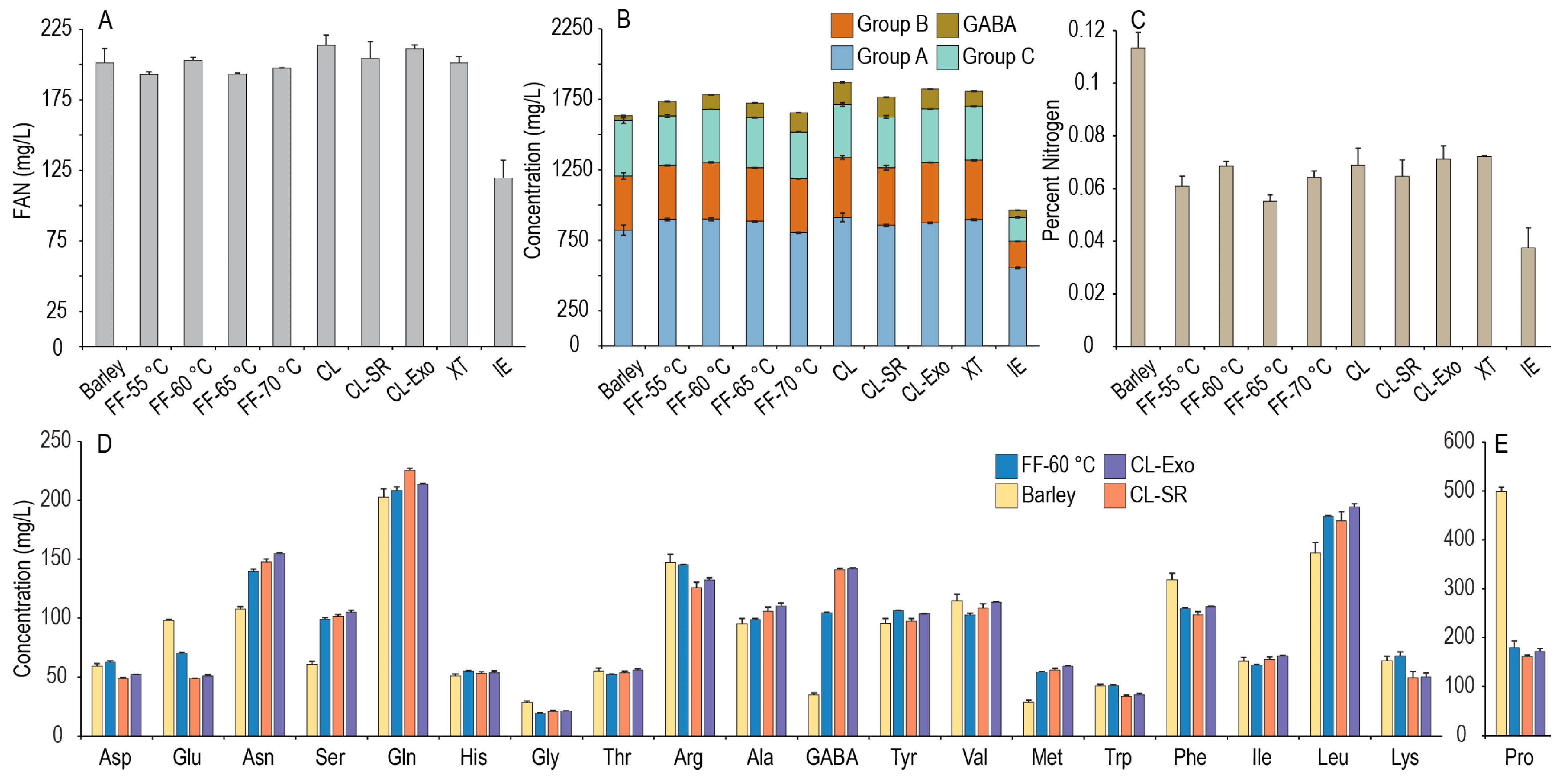
Nitrogen metabolism by yeast during primary fermentation is complex and dependent on the other constituents in the wort (e.g., FS), the type of yeast utilized, and fermentation conditions, but in terms of amino acids, it is dependent on both the total concentration of assimilable nitrogen (FAN) and the concentrations of individual amino acids [4,8,9]. The uptake of amino acids from the wort by brewing yeasts tends to follow a specific order, and the amino acids are classified into 4 groups based on the uptake preference Group A (most rapidly assimilated) to Group D, solely proline (little to no utilization). The amino acids were segmented into their respective groups using the initial classifications proposed by Jones and Pierce in 1964, except that methionine was included in Group A, as proposed by Lekkas, Hill, and Stewart in 2014 [1,25]. The sums of these amino acid groups and the relative abundances of each amino acid within its group are shown for both barley and each GF wort in Figure 1. An unidentified peak in the chromatograms varied significantly across the malts, suggesting the presence of an amino-containing compound not included in the amino acid standards was of a similar concentration to the α-amino acids in the worts. Injection and derivatization of a γ-aminobutyric acid (GABA) standard indicated that this unknown peak was indeed GABA, and therefore was identified and quantified as such in each of the worts. GABA is a non-proteogenic γ-amino acid found in the wort that can react with the ninhydrin reagent [30], thus included in the FAN measurement, and has been shown to be utilized by yeasts [11,35,36,37], suggesting it can also be a source of nitrogen for yeast in beer. However, investigations into its role in beer fermentations are limited, and it is not classified into one of the utilization groups. We then defined total amino acid content as the sum of all quantified amino acids, and total usable amino acids as the sum of all the α-amino acids and GABA concentrations in the wort, excluding proline, which requires an aerobic environment for utilization and thus is not utilized in a typical beer (i.e., anaerobic) fermentation [1].
As wort fermentations require both sufficient concentrations of yeast assimilable sugar and nitrogen sources, the FAN and amino acid profiles of GF worts made by the ExGM mashing procedure [20] were investigated. While several of these worts from GF malts (millet, sorghum, ivory teff, brown teff, and GFB1) contained similar or even higher concentrations of FS compared to a barley-derived wort prepared using a common isothermal infusion mash, deficiencies in FAN would require supplementation, and changes in the relative abundance of the amino acid profiles could lead to differences in fermentation characteristics. As shown in Figure 1A, there were large differences in the FAN content across the tested worts (the individual amino acid concentrations are provided in the supplementary data). Using the typical goal of producing a wort with 150–200 mg/L FAN, the GFB1 and sorghum malt worts could be considered FAN deficient (143 mg/L and 129 mg/L, respectively), indicating a need for further optimization. The corn, buckwheat, and rice malt worts were all deficient in FAN and deficient in FS using this mashing process, suggesting they lacked the required level of enzyme activity and would not currently be suitable for brewing using this approach. In contrast, the ivory and brown teff malt samples were high in FAN (445 mg/L and 490 mg/L) and produced high concentrations of FS (~90 g/L [20]), so while these malts were highly successful in producing FAN and FS, excess FAN could have some negative impacts on beer flavor or microbial stability [4,38]. The barley malt wort (201 mg/L FAN) and the millet malt wort (169 mg/L FAN) both had FAN contents within the suggested range. Overall, these results indicate that GF malts can be utilized successfully to produce worts with sufficient concentrations of FS and FAN. While some of the worts shown in Figure 1A were either FAN-sufficient or -abundant, differences in the amino acid profiles could lead to differences in the fermentation and flavor profiles.
As shown in Figure 1B,C, there were significant differences in the total amino acid contents for the various worts, with usable amino acids generally following the same trend as FAN measurements. On average, the measured FAN content is ~1/8 the sum of the usable amino acid concentrations because the ninhydrin reaction measures the nitrogen component of the amino group in each amino acid (nitrogen accounts for ~1/8 of the average mass of the amino acids). As is also shown in Figure 1C, there were significant differences in the GABA and proline contents of the worts. Proline was the most abundant free amino acid in the barley malt wort, as reported previously [1], and the barley malt contained more proline than any GF wort tested. Although proline is not typically utilized by yeast, it is an osmolyte that can serve as a cryoprotectant for yeast [39], and the proline-specific yeast permease can also uptake GABA [40], so differences between barley and GF malts in proline and GABA could lead to differences in fermentation and fermentation metabolites. Additionally, the proteins in each malt are likely different; for instance, barley is gluten-containing, and the GF malts are not. One of the hallmarks of immunogenic gluten peptides is the presence of proline-rich regions [41], and so the high proline content in a barley wort may be due to some gluten proteolysis during malting and mashing, but gluten peptides survive the brewing and fermentation processes. In contrast, GF malts may contain less proline overall compared to barley [41,42], so the lower proline content observed in these GF worts may be due in part to the lack of gluten proteins. The proline content alone does not make a gluten protein, as they are a complex suite of proteins found in barley, wheat, and rye, and differing individual sequences of the gluten proteins from each grain can produce differing effects in the immunogenicity, but the proline differences observed here between barley and GF malts could be some initial rationale for further investigation as to why the proteins of GF malts tend not to trigger gluten-sensitive individuals.
The underlying distribution of amino acids in the wort is important, as many amino acids serve as metabolic precursors to flavor-active compounds such as fusel alcohols and acetate esters via the Ehrlich pathway, diacetyl or other vicinal diketones, or sulfur-containing compounds via yeast sulfur assimilation and sulfur amino acid pathways [4,5,8,9,43,44]. As shown in Figure 1D–F, there are differences in the relative abundances of most amino acids within each utilization group between the produced worts. Millet, ivory teff, and brown teff malt worts all had higher relative abundances of methionine and lower phenylalanine than the barley wort, but overall, the millet malt wort had a similar profile to the barley malt wort (except proline). While the differences in relative abundances across these worts could lead to differences in the generation of flavor-active compounds, the relative profiles still need to be considered in the context of the absolute amino acid content as well. The effects of high amino acid contents on the fermentation of a teff wort are of particular interest as the FAN content was more than double that of barley, and it is unknown what impact this would have on the fermentation and flavor profiles. As the millet was close in FAN content to barley, could produce adequate FS, and was commercially available, we decided to utilize millet as a model GF malt to investigate if the FAN and amino acid content of a wort could be improved by modifying the mashing procedures. The remainder of this study now focuses on various manipulations to mashing with millet malt to investigate the role of mashing in the generation of amino acids to increase the wort amino acid content.
3.2. Amino Acid Content in the Mash from Millet Malt during the Enzyme Extraction Phase
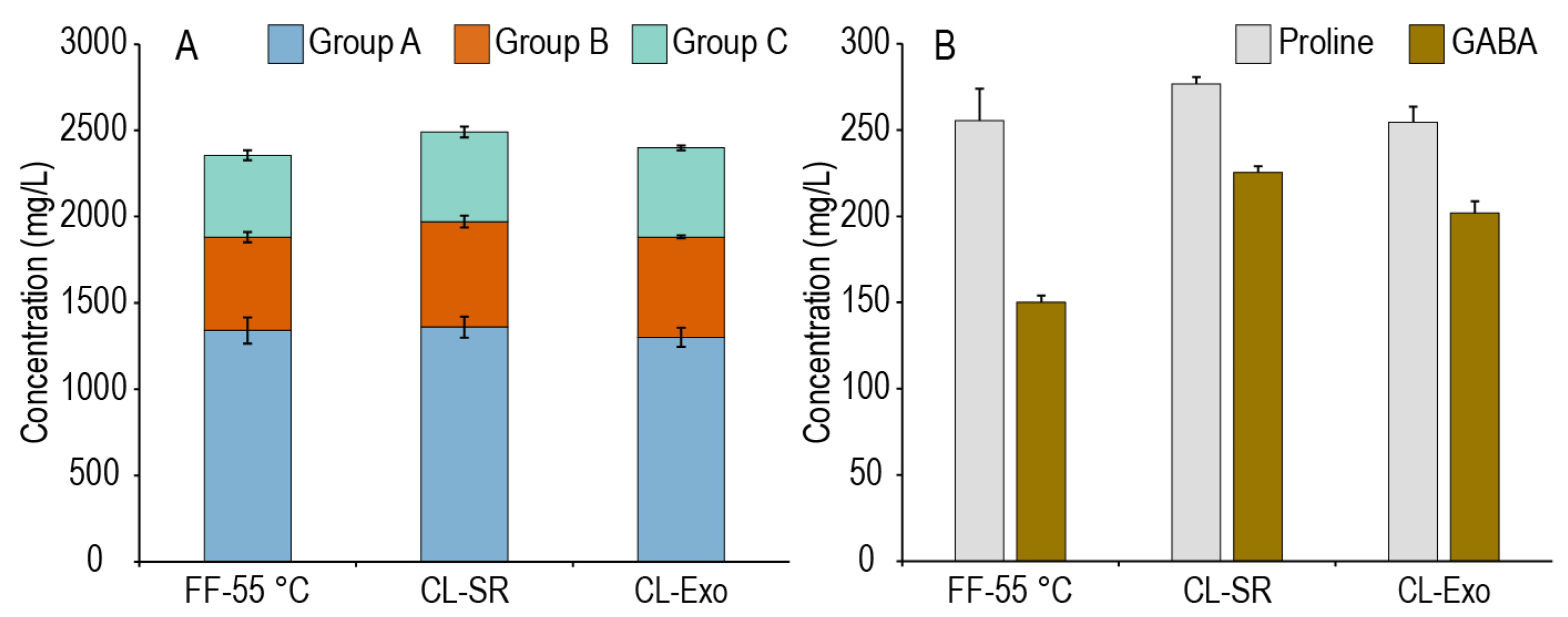
To investigate the effect of processing conditions such as grind size or exogenous enzyme supplementation on the generation of amino acids during the enzyme extraction from a millet malt mash, samples were taken from the enzyme extract at the conclusion of the enzyme extraction phase. In all these experiments, the enzyme extraction phase was held constant at 55 °C for 30 min at an L:G of 3:1, and the malt grind size (fine or coarse), extract separation technique (centrifuge or lauter), or enzyme system (endogenous or exogenous) was varied. As this step at 55 °C is comparable to the time and temperature regimes utilized for a protein rest in conventional barley brewing, it was expected that the different mashing modifications might lead to different amino acid contents in these extracts. In our previous study that analyzed the fermentable sugar concentrations of these samples, freely extracted sugar concentrations were higher with more finely ground samples, and the second batch had higher freely extracted fermentable sugars [27], so we would have expected a similar effect here. However, as shown in Figure 2A, there was largely no effect of our treatments on the overall amino acid content of this first wort (an analysis of all the worts is provided in Supplementary Figure S1). A fine or coarse grind yielded similar amino acid concentrations and profiles, and the addition of exogenous enzymes containing a neutral protease did not produce a large effect on the amino acid content. Rather, as shown in Figure 2B, the differences in the worts were largely due to which batch of millet was utilized, as CL-SR and CL-Exo were generated from the same batch of millet, and this batch tended to have higher GABA concentrations compared to the one utilized for FF-55 °C. The lack of effect of the grind size on amino acid extraction is interesting as it suggests that proteolysis at this stage may only be on small, easily extractable peptides that were likely generated during malting, and not larger protein bodies that might be rendered more accessible due to milling. The addition of an exogenous protease also had no significant effect on amino acid generation, suggesting that either the activity of the endogenous proteases was not limiting or, we suspect more likely, the added protease was not active under these conditions. To maintain consistency across these experiments, conditions deviated slightly from the protease manufacturer’s recommendations in terms of pH (acidification not recommended) and temperature (recommended maximum of 53 °C). Overall, these modifications to the initial phase of mashing had a minimal impact on amino acid production, leading us to further investigate whether there were any differences in amino acid generation between these protocols throughout the subsequent second mash.
3.3. Amino Acid Generation across a Mash
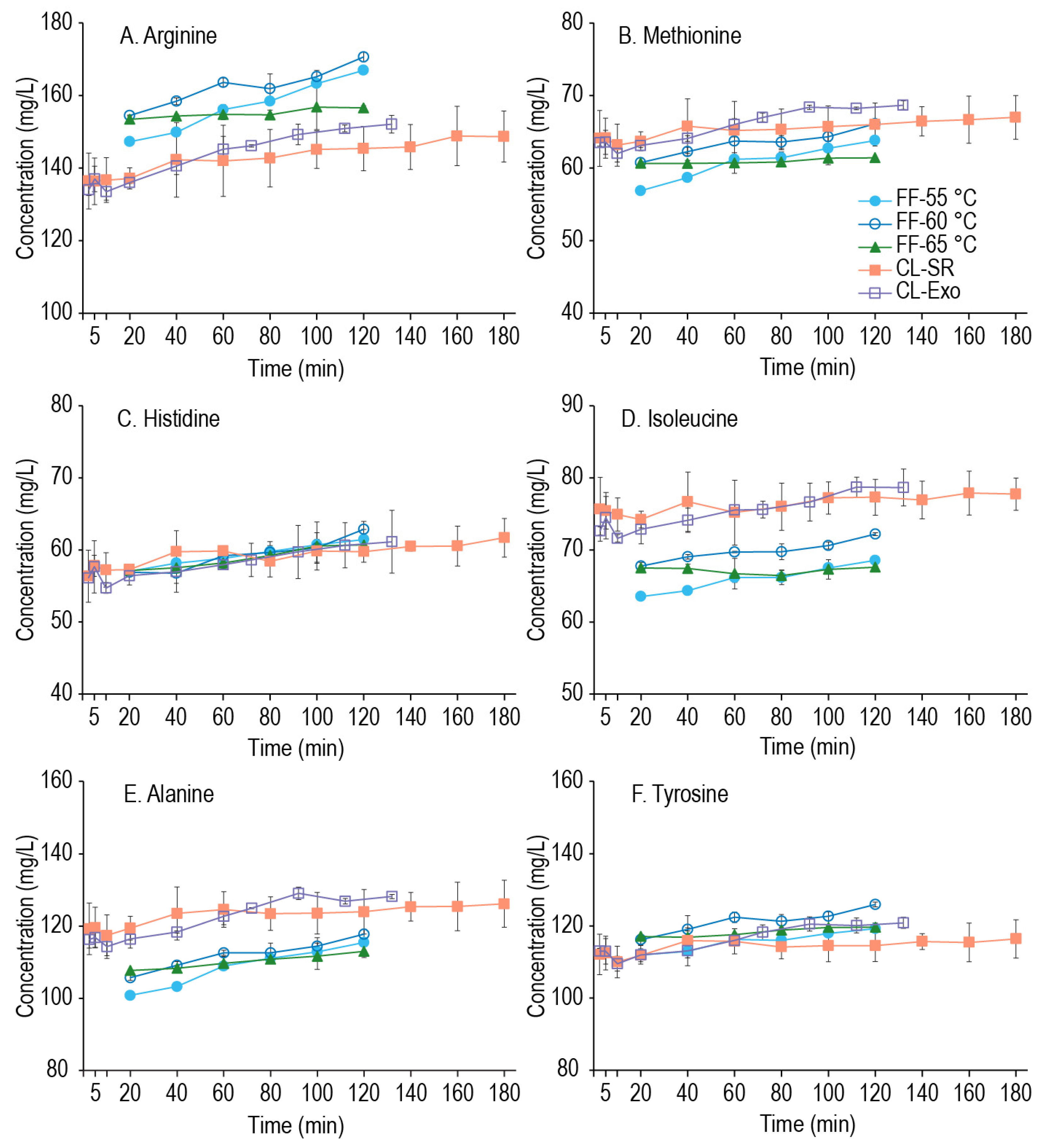
There is no consensus on the percentage of amino acids in a wort attributed to the result of mashing versus what is already present in the malt [21,23], but many barley malt step mashes utilize a protein rest of ~30 min at 55 °C or lower, and some commercial brewers will skip a protein rest all together, instead subjecting the mash to temperatures ≥ 62 °C. Given that good FS production was shown at 55 °C [20,27], we hypothesized that extended mashes at this temperature near that of a protein rest may be a simple avenue for improving the FAN content of a GF wort. The results shown in Figure 3 demonstrate the time-dependent concentration of 2 amino acids from each utilization group across the FF, CL-SR, and CL-Exo experiments, and graphs for all amino acids across time are provided as Supplemental Figures S2–S5, and values for the individual amino acid concentrations are provided in the supplementary data. As shown in all the panels of Figure 3, the concentration of amino acids that can be generated during the second mash was minimal. While Figure 3A,D,E show some change in those amino acids over time, arginine increased the most of any amino acid, and yet only increased 13% over the 2 h mash. As seen with the extracts, the batch of millet malt affected the amino acid content, while the grind size and addition of the exogenous enzymes had a minimal effect on amino acid generation. However, looking more closely at the FF samples across the three mashing temperatures (55–65 °C), there was some change in arginine, isoleucine, methionine, alanine, and tyrosine between 55 and 60 °C (all p ≤ 0.024), but no change at 65 °C (all p ≥ 0.162). This suggests that although the overall effects are minimal, some amino acid generation is possible within the relatively lower temperature range of 55–60 °C. Interestingly, the above-mentioned amino acids have hydrophobic side chains (except arginine), suggesting that exopeptidase(s) generating these amino acids may prefer substrates with terminal hydrophobic residues. Carboxypeptidases with similar product specificities (carboxypeptidase III) have been reported in triticale, wheat, and barley [45], and a similar enzyme in millet could be responsible for the formation of these amino acids. As some limited amino acid generation is possible during the mash, what might be limiting more significant amino acid generation in this system was the next investigation.
3.4. FAN and Amino Acid Profiles Produced from Millet under Different Mashing Conditions
As there were little observed differences in the amino acid concentrations in the extracts over the course of the mash, we decided to analyze two additional millet malt mashing treatments: the XT (55 °C for 2 h 45 min, no gelatinization step) and IE (temperature abused to denature enzyme activity) from our previous study [20] to determine if the gelatinization step in our experiments might be inhibiting protein degradation, or if the millet malt used could only produce small concentrations of amino acids overall during mashing. Figure 4A,B show the FAN and usable amino acid concentrations at the endpoints of the various mashing experiments. For FAN, no significant differences were observed between barley or any of the millet mashes, except for the IE samples. These results show that a millet wort can match the FAN content of a barley wort (~200 mg/L). Furthermore, given that the IE mash was intentionally temperature-abused to assess the endogenous background content of FS and amino acids in the malt, these results suggest that some proteolysis occurred during the 30 min enzyme extraction at 55 °C leading to the production of the additional observed amino acids. However, the XT samples held at a constant temperature of 55 °C did not differ in usable amino acid content from any other millet mash (except the IE samples), suggesting that the gelatinization step following enzyme extraction is not limiting with respect to additional proteolytic reactions, but that nearly all the additional amino acids observed occurred within 30 min at 55 °C. These results demonstrate that amino acids can be generated during the initial enzyme extraction phase of the mash, but that some mechanism is limiting further protein degradation in this system.
The two main classes of protein-degrading enzymes (endopeptidases and exopeptidases) are both required to efficiently produce small peptides and free amino acids in malts. Despite the different enzymatic properties of the various peptidase classes (pH, temperature, substrate, etc.) [22,23,24,45], in mashing, the major variable that a brewer can change to modify proteolytic activity is temperature (mash pH can be changed, but modification to <5 where many peptidases have high activity is probably not reasonable), so a brewer should utilize temperatures where the peptidases are thermostable if a goal is to maximize proteolysis. As our results show, some proteolysis occurs during the enzyme extraction at 55 °C, which indicates that some endo- and exoproteolytic activities remain in the malt; however, as this proteolysis seems limited to a short timeframe at 55 °C, they likely are not thermostable at that temperature.
To ascertain whether endo- or exopeptidases are limiting amino acid generation beyond the enzyme extraction, the soluble nitrogen (SN) contents of the worts were evaluated. As each wort had similar amino acid concentrations regardless of modifying the mashing temperature or adding exogenous enzymes, differences in soluble nitrogen content could suggest endopeptidases may still be active in the wort and capable of producing soluble peptides. As the combustion analysis measures all nitrogenous species in the wort, and FAN measures the nitrogen primarily from amino acids and small peptides, assuming the total nitrogen content of a wort is dominated by proteins (rather than other nitrogenous compounds) one can estimate the percentages of SN contributed by FAN and soluble proteins. The SN analysis in Figure 4C shows only that the IE sample was significantly different from the other millet worts in terms of total soluble nitrogen due to thermally induced enzyme denaturation, and that in the remaining millet worts, FAN accounted for between 27 and 35% of the SN. Since FAN does not account for most of the measured soluble nitrogen, there are likely soluble peptides remaining in the wort that were not measured by the FAN analysis, suggesting that incomplete peptide degradation by exopeptidases (possibly due to inactivation) occurred. Furthermore, that the SN values are not different between millet samples mashed at different temperatures suggests that endopeptidases do not remain active to any significant degree beyond the enzyme extraction step, and both enzyme classes may be limiting. Furthermore, all the millet malt worts had significantly less soluble nitrogen content than the barley malt wort, despite having similar amino acid contents (total amino acids in barley wort: 2133 mg/L vs. average of all millet worts: 1938 mg/L). This suggests there may be more soluble peptides present in the barley malt wort, as FAN only accounted for 17% of the SN in the barley wort, and differences in soluble peptides could lead to attributes such as good foam formation and stability that millet-based worts may be lacking. This may be due to the less peptidase activity in a millet malt that survives kilning and could be utilized in the enzyme extraction or less overall proteolytic degradation during malting. Although previous studies have demonstrated that barley’s endopeptidases can survive kilning and can affect the SN and FAN content of a wort, the proteolytic activities of malts remain relatively understudied [22,24].
The amino acid profiles of three different final millet worts are shown in Figure 4D, and, when compared to the barley malt wort, show that although there are differences in amino acids such as proline, glutamine, serine, GABA, phenylalanine, and leucine, the overall amino acid profile of a millet malt wort can be similar to a barley malt wort. While these differences could still lead to changes in concentrations of higher alcohols in finished beers, there is no obvious amino acid deficiency in the millet malt-based worts, suggesting their promise as a base malt for GF beer production. Returning to our overall hypothesis that the amino acid content of a wort could be modified by maintaining milder temperatures (55–60 °C) across the mash, a small quantity of amino acids can be generated, but the only step where significant protein degradation can likely occur is during the enzyme extraction.
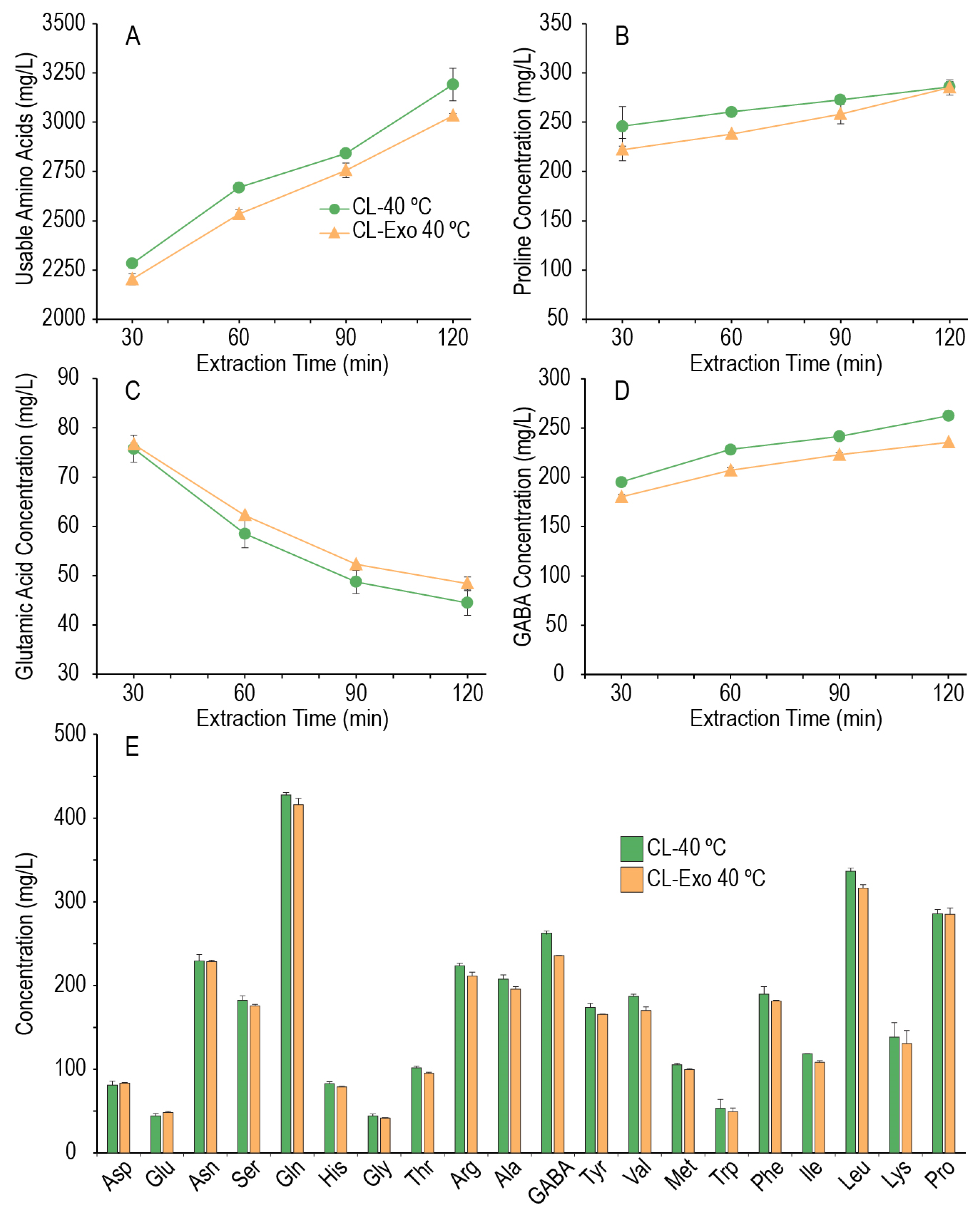
3.5. Effect of Mild Temperature Mashing on Amino Acid Generation
As it appeared that some amino acids could be generated in the enzyme extraction phase, but the window of protein degradation was limited, we hypothesized that the peptidases present in the system may not be thermostable at 55 °C; therefore, additional mashes were performed where only the enzyme extraction phase was performed and was held at 40 °C for 2 h, with aliquots taken every 30 min to assess the change in amino acid content over time. As the exogenous enzymes did not appear to have a major effect at temperatures ≥ 55 °C, the effect of the exogenous enzymes at this lower temperature extraction phase was also tested. Figure 5 shows that at 40 °C, amino acid concentrations could increase over the course of 2 h (individual amino acid concentrations are provided in the supplementary data). For the endogenous millet system, 30 min at 40 °C extracted slightly less usable amino acids into the wort than the average of the extracts at 55 °C (2284 mg/L vs. 2577 mg/L), but more will be produced at 40 °C if the time is increased. After 2 h at 40 °C, the endogenous millet system increased usable amino acids (CL-40 °C 3191 mg/L, 40% increase; CL-Exo 40 °C 3033 mg/L, 38% increase, respectively over 2 h) and increased 614 mg/L compared to an extraction for 30 min at 55 °C. As these experiments did not undergo the entire mash process, we did not measure what the amino acid content would be in the final wort, but across all the other experiments where the amino acid concentrations varied little beyond the enzyme extraction step, the total (including proline) amino acid content in the final wort was 68.11 ± 2.64% of the total amino acid content of the extract (despite a change in volume ratio from L:G 3:1 to 6:1, the drop is not by half as the sparge process rinses the grain). Using that modified dilution factor, it was estimated what the total wort total amino acid concentrations could have been for CL-40 °C and CL-Exo 40 °C if the process was continued with a second mash temperature of ≥55 °C (estimated final wort total amino acids: 2364 mg/L and 2260 mg/L, respectively), with an estimated increase of 418 mg/L usable amino acids in the final wort for CL-40 °C. Similar estimations for FAN and soluble nitrogen (2 h: 512 mg/L, 0.14% nitrogen; estimated final wort: 348 mg/L, 0.095% nitrogen) would suggest that an extended mash at a mild temperature will improve proteolytic degradation in the millet malt mash, producing a final wort with higher amino acid content, FAN, and soluble nitrogen.
As in the extractions at 55 °C, it appears the addition of exogenous enzymes did not markedly improve amino acid production at 40 °C (Figure 5A,B,E), suggesting that the thermostability of the exogenous protease was not limiting. While the possibility that acidifying the mash may have limited the proteolytic activity of the neutral protease in the exogenous enzyme cocktail cannot be ruled out, under the conditions tested here, the addition of the exogenous enzymes appears to have slightly hindered the proteolytic activity of the mash. While an underlying mechanism for this result was not explored, one hypothesis could be that the exogenous protease, if active, may be producing peptides that are not ideal substrates for the millet’s exopeptidases.
In this mild (40 °C) temperature system, it was observed that every proteogenic amino acid increased in concentration (to differing extents) across the 2 h step, except for glutamic acid, which decreased. GABA also increased in these mashes, and although non-proteogenic, it can be enzymatically produced by the decarboxylation of glutamic acid by the enzyme glutamate decarboxylase found in plants, animals, and yeasts [35,37,43]. The increase in GABA and decrease in glutamate suggest that there may be some glutamate decarboxylase activity in these millet malts.
These results provide some evidence to support our initial hypothesis that mashing can be utilized to modify the amino acid content of the wort, but that it must occur during a rest in the mashing scheduling at a temperature where the proteolytic enzymes are thermostable. In our system, this translated to a maximum of 55 °C for some amino acid generation, but a lower temperature for a longer time if a larger amount of amino acid generation was required.
3.6. Effect of Mild Temperature Mashing on Enzyme Extraction
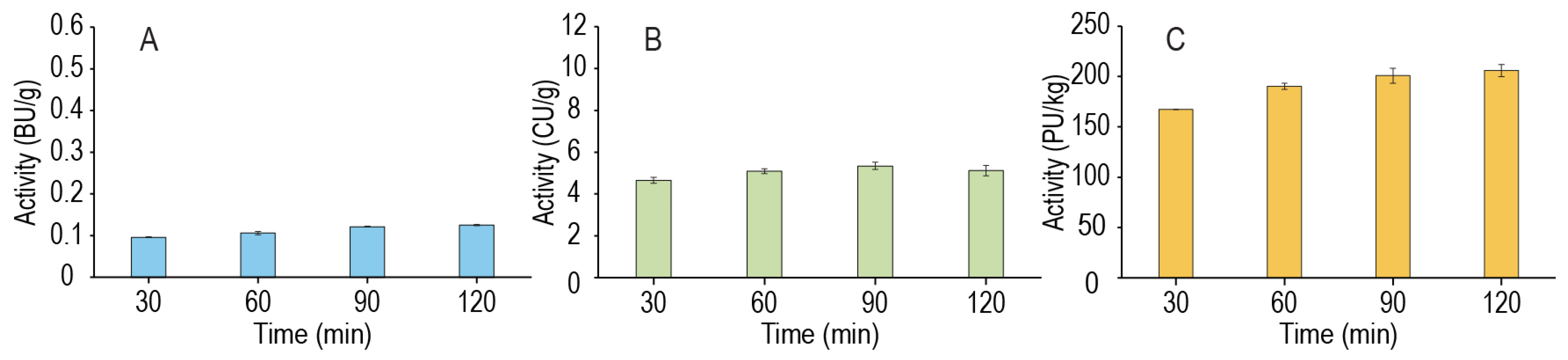
As the primary goal of the mash is still the production of FS, procedures that could improve the amylolytic enzyme extraction to potentially increase the rate of sugar production were also of interest. In particular, as previous studies have shown the amylolytic enzymes β-amylase and limit dextrinase have proteinaceous inhibitors [29], we sought to investigate if this proteolytically active low temperature enzyme extraction could also improve amylolytic enzyme extraction from a coarsely ground malt by liberating these bound enzymes. Figure 6 shows that both β-amylase and limit dextrinase significantly increased (p = 0.001 and p = 0.005) across a 2 h extraction. While statistically significant, the magnitude of the increase was small enough that we believe the effect on sugar generation would be minor, although further investigation may be warranted. As there was no significant change in the α-amylase extracted (p = 0.072), one hypothesis is that a physical factor such as the cell wall structure or enzyme compartmentalization in the millet grain may be a limiting factor in the extraction of the amylolytic enzymes. Thus, improvements in the proteolytic activity at 40 °C would have a minimal effect on the release of the enzymes from their inhibitors if they remained physically inaccessible to the aqueous mash phase.
3.7. The Potential Impact of GF Worts on Fermentation
Overall, all the results shown here demonstrate that GF worts can vary both in terms of FAN and amino acid profile, influenced by the malt used (Section 3.1), or in the millet malt experiments by the times and temperatures of the initial enzyme extraction phase and/or protein rest (Section 3.5). Questions remain as to what effect differences in the amino acid profiles and FS may have on the subsequent fermentations. Higher amino acid contents would likely lead to additional products of Maillard browning during the boil, and the various flavor-active compounds produced by Strecker degradation of those intermediates [8]. Differences in leucine, isoleucine, and phenylalanine concentrations could lead to differences in the formation of isoamyl alcohol, amyl alcohol, and phenylethanol, and flavor-active esters derived from the condensation of alcohols with acetyl-CoA or longer fatty acids (isoamyl acetate, phenylethyl acetate, ethyl hexanoate, etc.) directly impacting the beer flavor. Prior research has shown that the fermentation of worts with higher concentrations of monosaccharides tends to produce beers with higher concentrations of higher alcohols and esters [10,46]. As one of the results observed from these GF worts is they tended to have higher glucose and fructose concentrations compared to barley [20], those differences in both sugar distribution and the amino acid results shown here could also lead to differences in the production of those important flavor-active metabolites.
Different GABA concentrations in the GF worts compared to barley were also observed, but studies tracking the utilization of GABA in wort and its effects on beer fermentation are limited. Some studies in wine show that GABA can be utilized to a significant degree, and that its primary metabolic product is succinate [36,37]. However, the extent to which GABA from a GF wort will be utilized in relation to the other amino acids is unknown, as is its effect on fermentation or flavor profile. Yeast generally has two primary mechanisms of amino acid uptake, a general amino acid permease system that dominates in nitrogen-poor environments, or more specific amino acid transporters in nitrogen-rich environments [47], and GABA can been assimilated by yeast through the general system, its own specific permeases, or through proline permeases [40]. The degree to which GABA can be utilized in a wort would likely be dependent on its concentration, the total amino acid concentration, potentially proline concentration, and any other variable that could affect yeast fermentation. Therefore, further research is needed to elucidate the role of increased GABA concentrations present in these GF worts on finished beer characteristics.
While the simplest method to improve the FAN content of a wort may be for a brewer to supplement with exogenous sources of nitrogen, there has been relatively little research investigating avenues to modify the amino acid content and profile of a wort, even when using more traditional barley malts [22]. Similarly, while yeast nutrient supplements can provide some benefits to yeast nutrition, the supplements can be variable and are often under characterized, and their utilization (especially at higher gravities) without careful consideration could have unintended consequences on the resulting fermentation [48]. This means further research on various wort components and manners in which brewers could influence those components could aid in identifying more targeted approaches to supplement potential specific wort nutrient deficiencies, such as amino acids. However, beyond a way to support a healthy fermentation, amino acids will serve as flavor precursors, and recent research has sought to better manipulate the amino acid profile of a wort to drive the specific formation of fusel alcohols and acetates [49]. In that work, the authors sought to improve the release of specific amino acids (valine, phenylalanine, leucine, isoleucine) through the use of exogenous endo- and exoproteases to drive the increased production of specific fusel alcohols and acetates. While this particular study was performed with exogenous enzymes, the results do suggest that better understanding and manipulating the proteolytic mechanisms in a mash could be a potential avenue to purposefully drive beer flavor differences during brewing. Our results here show that the amino acid content of a millet malt wort can be improved by relying on the endogenous proteolytic system with a simple extension of a mild temperature step in the mashing schedule (Figure 5), potentially providing new opportunities to drive flavor differences without requiring exogenous ingredients. While improving amino acid generation is not strictly necessary in most cases of traditional barley malt brewing or here with the millet malt samples, this study presents evidence that the endogenous proteolytic system could provide more of a direct role in the formation of amino acids during mashing, and that a better understanding of these systems could give brewers new opportunities to influence beer flavor and quality.
4. Conclusions
The purpose of this work was to evaluate the FAN and amino acid content of a variety of GF worts, and to assess if such worts were deficient in FAN and/or if their amino acid profiles differed significantly from those made from barley. It was determined that depending on the GF malt used, FAN contents either within, exceeding, or below the range suggested for barley could be produced, and that the profiles differed across malts, potentially leading to differences in fermentation. A goal of this work was also to determine if altered mash conditions could be utilized to increase amino acid generation so that brewers could manipulate their process to produce a wort with the desired amino acid contents or to produce a general increase if using FAN-deficient malts. Our results showed that extending a protein rest or enzyme extraction at the relatively low temperature of 40 °C could serve as a simple and effective way of improving amino acid generation when using millet malt. Furthermore, the endogenous proteolytic enzymes present in these malted millet worts do not appear to be thermally stable for extended periods of time at temperatures ≥ 55 °C. While some amino acids were generated over the course of a mash at 55 or 60 °C, the effect was vanishingly small, and it is unlikely that amino acids can be generated in high concentrations at these higher temperatures where sugar generation is optimal. Overall, a simple method (longer time at 40 °C) for modifying the amino acid content of a millet malt wort was presented, that could potentially be adaptable to other GF malts. It was also demonstrated that GF malts can be valuable brewing ingredients, previously, in terms of fermentable sugars [20,27], and here in terms of amino acids.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/beverages9010010/s1, Figure S1: Sum of usable amino acids by utilization Group (A) and proline and GABA concentrations (B) of the samples collected at the end of the enzyme extraction phase of each respective mashing experiment. FF-55 °C through FF-65 °C were samples from the first batch of millet malt, while the remainder were from the second batch of millet malt. Figure S2: Time-dependent amino acid concentration of each Group A amino acid. Amino acid concentrations were analyzed as described in Section 2.7. Figure S3: Time-dependent amino acid concentration of each Group B amino acid. Amino acid concentrations were analyzed as described in Section 2.7. Figure S4: Time-dependent amino acid concentration of each Group C amino acid. Amino acid concentrations were analyzed as described in Section 2.7. Figure S5: Time-dependent amino acid concentration of proline (Group D) and γ-aminobutyric acid (GABA). Amino acid concentrations were analyzed as described in Section 2.7. Supplementary data: Individual amino acid, FAN, and nitrogen analysis values for the GF worts, millet time-course, and CL-40 °C experiments
Author Contributions
Conceptualization, A.J.L., R.J.E. and D.W.C.; methodology, A.J.L., R.J.E. and D.W.C.; formal analysis, A.J.L.; investigation, A.J.L.; resources, R.J.E. and D.W.C.; writing—original draft preparation, A.J.L.; writing—review and editing, A.J.L., R.J.E. and D.W.C.; visualization, A.J.L.; supervision, R.J.E. and D.W.C.; project administration, R.J.E. and D.W.C.; funding acquisition, R.J.E. and D.W.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded, in part, through technical assistance grants to D. Cockburn and R. Elias (PO 63020683 and PO63019744) by the Pennsylvania Department of Agriculture through the Pennsylvania Malt and Brewed Beverage Industry Promotion Board. This work was also supported by the USDA National Institute of Food and Agriculture Federal Appropriations to D. Cockburn (project PEN04650 accession no. 1015962) and R. Elias (project PEN04522, accession no. 0233376).
Data Availability Statement
Available data pertinent to the results of this study are reported in the manuscript and the Supplementary Materials.
Acknowledgments
The authors would like to thank Laura Rolon for her feedback on figure layout and design.
Conflicts of Interest
The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Jones, M.; Pierce, J.S. Absorption of amino acids from wort by yeasts. J. Inst. Brew. 1964, 70, 307–315. [Google Scholar] [CrossRef]
- Patel, G.B.; Ingledew, W.M. Trends in Wort Carbohydrate Utilization. Appl. Microbiol. 1973, 26, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Fontana, M.; Buiatti, S. 25—Amino Acids in Beer. In Beer in Health and Disease Prevention; Preedy, V.R., Ed.; Academic Press: San Diego, CA, USA, 2009; pp. 273–284. [Google Scholar]
- Hill, A.E.; Stewart, G.G. Free amino nitrogen in brewing. Fermentation 2019, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Krogerus, K.; Gibson, B.R. 125th Anniversary Review: Diacetyl and its control during brewery fermentation. J. Inst. Brew. 2013, 119, 86–97. [Google Scholar] [CrossRef]
- Lie, S. The EBC-ninhydrin method for determination of free alpha amino nitrogen. J. Inst. Brew. 1973, 79, 37–41. [Google Scholar] [CrossRef]
- Engan, S. Wort composition and beer flavour. I. The influence of some amino acids on the formation of higher aliphatic alcohols and esters. J. Inst. Brew. 1970, 76, 254–261. [Google Scholar] [CrossRef]
- Ferreira, I.M.; Guido, L.F. Impact of Wort Amino Acids on Beer Flavour: A Review. Fermentation 2018, 4, 23. [Google Scholar] [CrossRef] [Green Version]
- Stewart, G.G.; Hill, A.; Lekkas, C. Wort FAN—Its Characteristics and Importance during Fermentation. J. Am. Soc. Brew. Chem. 2013, 71, 179–185. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Derdelinckx, G.; Dufour, J.-P.; Winderickx, J.; Thevelein, J.M.; Pretorius, I.S.; Delvaux, F.R. Flavor-active esters: Adding fruitiness to beer. J. Biosci. Bioeng. 2003, 96, 110–118. [Google Scholar] [CrossRef]
- Ljungdahl, P.O.; Daignan-Fornier, B. Regulation of amino acid, nucleotide, and phosphate metabolism in Saccharomyces cerevisiae. Genetics 2012, 190, 885–929. [Google Scholar] [CrossRef]
- FDA. 85 FR 49240—Food Labeling; Gluten-Free Labeling of Fermented or Hydrolyzed Foods; FDA: Silver Spring, MD, USA, 2020.
- Allred, L.K.; Lesko, K.; McKiernan, D.; Kupper, C.; Guandalini, S. The Celiac Patient Antibody Response to Conventional and Gluten-Removed Beer. J. AOAC Int. 2019, 100, 485–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanner, G.J.; Colgrave, M.L.; Howitt, C.A. Gluten, Celiac Disease, and Gluten Intolerance and the Impact of Gluten Minimization Treatments with Prolylendopeptidase on the Measurement of Gluten in Beer. J. Am. Soc. Brew. Chem. 2014, 72, 36–50. [Google Scholar] [CrossRef]
- Fiedler, K.L.; Panda, R.; Croley, T.R. Analysis of Gluten in a Wheat-Gluten-Incurred Sorghum Beer Brewed in the Presence of Proline Endopeptidase by LC/MS/MS. Anal. Chem. 2018, 90, 2111–2118. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, K.L.; Cao, W.; Zhang, L.; Naziemiec, M.; Bedford, B.; Yin, L.; Smith, N.; Arbuckle, M.; Lopez-Hernandez, A.; Jackson, L.S. Detection of gluten in a pilot-scale barley-based beer produced with and without a prolyl endopeptidase enzyme. Food Addit. Contam. Part A 2019, 36, 1151–1162. [Google Scholar] [CrossRef]
- Watson, H.G.; Decloedt, A.I.; Hemeryck, L.Y.; Van Landschoot, A.; Prenni, J. Peptidomics of an industrial gluten-free barley malt beer and its non-gluten-free counterpart: Characterisation and immunogenicity. Food Chem. 2021, 355, 129597. [Google Scholar] [CrossRef]
- Ceccaroni, D.; Marconi, O.; Sileoni, V.; Wray, E.; Perretti, G. Rice malting optimization for the production of top-fermented gluten-free beer. J. Sci. Food Agric. 2019, 99, 2726–2734. [Google Scholar] [CrossRef]
- Di Ghionno, L.; Sileoni, V.; Marconi, O.; De Francesco, G.; Perretti, G. Comparative study on quality attributes of gluten-free beer from malted and unmalted teff [Eragrostis tef (zucc.) trotter]. LWT 2017, 84, 746–752. [Google Scholar] [CrossRef]
- Ledley, A.J.; Elias, R.J.; Hopfer, H.; Cockburn, D.W. A Modified Brewing Procedure Informed by the Enzymatic Profiles of Gluten-Free Malts Significantly Improves Fermentable Sugar Generation in Gluten-Free Brewing. Beverages 2021, 7, 53. [Google Scholar] [CrossRef]
- Barrett, J.; Kirsop, B.H. The relative contributions to wort nitrogen of nitrogenous substances solubilized during malting and mashing. J. Inst. Brew. 1971, 77, 39–42. [Google Scholar] [CrossRef]
- Jones, B.L.; Budde, A.D. How various malt endoproteinase classes affect wort soluble protein levels. J. Cereal Sci. 2005, 41, 95–106. [Google Scholar] [CrossRef]
- Jones, B.L.; Marinac, L. The Effect of Mashing on Malt Endoproteolytic Activities. J. Agric. Food Chem. 2002, 50, 858–864. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.L.; Marinac, L.A.; Fontanini, D. Quantitative Study of the Formation of Endoproteolytic Activities during Malting and Their Stabilities to Kilning. J. Agric. Food Chem. 2000, 48, 3898–3905. [Google Scholar] [CrossRef] [PubMed]
- Lekkas, C.; Hill, A.E.; Stewart, G.G. Extraction of FAN from Malting Barley during Malting and Mashing. J. Am. Soc. Brew. Chem. 2014, 72, 6–11. [Google Scholar] [CrossRef]
- Evans, D.E.; Fox, G.P. Comparison of Diastatic Power Enzyme Release and Persistence during Modified Institute of Brewing 65°C and Congress Programmed Mashes. J. Am. Soc. Brew. Chem. 2017, 75, 302–311. [Google Scholar] [CrossRef]
- Ledley, A.J.; Elias, R.J.; Cockburn, D.W. Impact of Mashing Protocol on the Formation of Fermentable Sugars from Millet in Gluten-Free Brewing. Food Chem. 2022; accepted for publication. [Google Scholar]
- Cornaggia, C.; Evans, D.E.; Draga, A.; Mangan, D.; McCleary, B.V. Prediction of potential malt extract and beer filterability using conventional and novel malt assays. J. Inst. Brew. 2019, 125, 294–309. [Google Scholar] [CrossRef]
- Evans, D.E. A More Cost- and Labor-Efficient Assay for the Combined Measurement of the Diastatic Power Enzymes β-Amylase, α-Amylase, and Limit Dextrinase. J. Am. Soc. Brew. Chem. 2008, 66, 215–222. [Google Scholar] [CrossRef]
- American Society of Brewing Chemists, ASBC Methods of Analysis, Online. Wort 12A. Free Amino Nitrogen; American Society of Brewing Chemists: St. Paul, MN, USA, 2010. [CrossRef]
- LECO. Nitrogen in Wort and Beer; Form Number 203-821-153; LECO Corporation: St. Joseph, MI, USA, 2008. [Google Scholar]
- Henderson, J.W.; Brooks, A. Improved Amino Acid Methods Using Agilent ZORBAX Eclipse Plus C18 Columns for a Variety of Agilent LC Instrumentation and Separation Goals; Publication Number 5990-4547EN; Agilent Technologies, Inc.: Santa Clara, CA, USA, 2010. [Google Scholar]
- Henderson, J.W.; Ricker, R.D.; Bidlingmeyer, B.A.; Woodward, C. Rapid, Accurate, Sensitive, and Reproducible HPLC Analysis of Amino Acids; Publication Number 5980-1193E; Agilent Technologies, Inc.: Santa Clara, CA, USA, 2000. [Google Scholar]
- Long, W. Automated Amino Acid Analysis Using an Agilent Poroshell HPH-C18 Column; Publication Number 5991-5571EN; Agilent Technologies, Inc.: Santa Clara, CA, USA, 2015. [Google Scholar]
- Bach, B.; Meudec, E.; Lepoutre, J.-P.; Rossignol, T.; Blondin, B.; Dequin, S.; Camarasa, C. New insights into γ-aminobutyric acid catabolism: Evidence for γ-hydroxybutyric acid and polyhydroxybutyrate synthesis in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2009, 75, 4231–4239. [Google Scholar] [CrossRef] [Green Version]
- Bach, B.; Sauvage, F.-X.; Dequin, S.; Camarasa, C. Role of γ-Aminobutyric Acid as a Source of Nitrogen and Succinate in Wine. Am. J. Enol. Vitic. 2009, 60, 508. [Google Scholar] [CrossRef]
- Gobert, A.; Tourdot-Maréchal, R.; Sparrow, C.; Morge, C.; Alexandre, H. Influence of nitrogen status in wine alcoholic fermentation. Food Microbiol. 2019, 83, 71–85. [Google Scholar] [CrossRef]
- Pierce, J.S. Horace brown memorial lecture the role of nitrogen in brewing. J. Inst. Brew. 1987, 93, 378–381. [Google Scholar] [CrossRef]
- Takagi, H. Proline as a stress protectant in yeast: Physiological functions, metabolic regulations, and biotechnological applications. Appl. Microbiol. Biotechnol. 2008, 81, 211–223. [Google Scholar] [CrossRef]
- André, B.; Hein, C.; Grenson, M.; Jauniaux, J.-C. Cloning and expression of the UGA4 gene coding for the inducible GABA-specific transport protein of Saccharomyces cerevisiae. Mol. Gen. Genet. MGG 1993, 237, 17–25. [Google Scholar] [CrossRef]
- Scherf, K.A.; Wieser, H.; Koehler, P. Improved Quantitation of Gluten in Wheat Starch for Celiac Disease Patients by Gel-Permeation High-Performance Liquid Chromatography with Fluorescence Detection (GP-HPLC-FLD). J. Agric. Food Chem. 2016, 64, 7622–7631. [Google Scholar] [CrossRef]
- Gebremariam, M.M.; Zarnkow, M.; Becker, T. Teff (Eragrostis tef) as a raw material for malting, brewing and manufacturing of gluten-free foods and beverages: A review. J. Food Sci. Technol. 2014, 51, 2881–2895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ardö, Y. Flavour formation by amino acid catabolism. Biotechnol. Adv. 2006, 24, 238–242. [Google Scholar] [CrossRef]
- Thomas, D.; Surdin-Kerjan, Y. Metabolism of sulfur amino acids in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 1997, 61, 503–532. [Google Scholar] [CrossRef]
- Drzymała, A.; Bielawski, W. Isolation and characterization of carboxypeptidase III from germinating triticale grains. Acta Biochim. Biophys. Sin. 2009, 41, 69–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Younis, O.S.; Stewart, G.G. Sugar uptake and subsequent ester and higher alcohol production by Saccharomyces cerevisiae. J. Inst. Brew. 1998, 104, 255–264. [Google Scholar] [CrossRef]
- Stewart, G.G. Yeast Nutrition. In Brewing and Distilling Yeasts; Stewart, G.G., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 109–146. [Google Scholar]
- Gibson, B.R. 125th Anniversary Review: Improvement of Higher Gravity Brewery Fermentation via Wort Enrichment and Supplementation. J. Inst. Brew. 2011, 117, 268–284. [Google Scholar] [CrossRef]
- Lin, C.L.; Petersen, M.A.; Mauch, A.; Gottlieb, A. Towards lager beer aroma improvement via selective amino acid release by proteases during mashing. J. Inst. Brew. 2022, 128, 15–21. [Google Scholar] [CrossRef]
Figure 1.
FAN measurements of GF worts compared to barley (A); sum of usable amino acids by utilization group (B); proline and GABA concentrations (C); and relative abundances of each amino acid within the utilization group, Group A (D), Group B (E), and Group C (F) of barley infusion mash samples and GF ExGM mash samples. All samples had a final liquor-to-grist ratio of 6:1.
Figure 1.
FAN measurements of GF worts compared to barley (A); sum of usable amino acids by utilization group (B); proline and GABA concentrations (C); and relative abundances of each amino acid within the utilization group, Group A (D), Group B (E), and Group C (F) of barley infusion mash samples and GF ExGM mash samples. All samples had a final liquor-to-grist ratio of 6:1.

Figure 2.
Sum of usable amino acids by utilization Group (A) and proline and GABA concentrations (B) of the samples collected at the end of the enzyme extraction phase of the FF-55 °C, CL-SR, and CL-Exo mashing experiments. FF-55 °C was sampled from the first batch of millet malt, while the CL samples were from the second batch of millet malt. Worts from this enzyme extraction phase are at a liquor-to-grist ratio of 3:1.
Figure 2.
Sum of usable amino acids by utilization Group (A) and proline and GABA concentrations (B) of the samples collected at the end of the enzyme extraction phase of the FF-55 °C, CL-SR, and CL-Exo mashing experiments. FF-55 °C was sampled from the first batch of millet malt, while the CL samples were from the second batch of millet malt. Worts from this enzyme extraction phase are at a liquor-to-grist ratio of 3:1.

Figure 3.
Time-dependent concentrations of two amino acids from each utilization group. Group A: (A) Arginine, (B) Methionine; Group B: (C) Histidine, (D) Isoleucine; and Group C: (E) Alanine, (F) Tyrosine from the malted millet worts described in Section 2.4. Time 0 min refers to the start of the second mash, upon recombination of the enzyme extract and the cooled gelatinized starch, and all samples are at a liquor-to-grist ratio of 4:1.
Figure 3.
Time-dependent concentrations of two amino acids from each utilization group. Group A: (A) Arginine, (B) Methionine; Group B: (C) Histidine, (D) Isoleucine; and Group C: (E) Alanine, (F) Tyrosine from the malted millet worts described in Section 2.4. Time 0 min refers to the start of the second mash, upon recombination of the enzyme extract and the cooled gelatinized starch, and all samples are at a liquor-to-grist ratio of 4:1.

Figure 4.
FAN measurements (A); sum of usable amino acids by utilization group and GABA (B); soluble nitrogen measurements (C); and select amino acid profiles (D) comparing the barley infusion mash samples and the endpoints of the time-course millet mash samples. XT and IE samples refer to the millet-XT and millet-IE samples described in Section 2.3, and all samples are from worts at a final liquor-to-grist ratio of 6:1 and (E) comparing proline levels from these worts.
Figure 4.
FAN measurements (A); sum of usable amino acids by utilization group and GABA (B); soluble nitrogen measurements (C); and select amino acid profiles (D) comparing the barley infusion mash samples and the endpoints of the time-course millet mash samples. XT and IE samples refer to the millet-XT and millet-IE samples described in Section 2.3, and all samples are from worts at a final liquor-to-grist ratio of 6:1 and (E) comparing proline levels from these worts.

Figure 5.
Sum of the usable amino acids, including GABA (A), proline concentration (B), glutamic acid concentration (C), and GABA concentration (D) across the extended enzyme extraction (L:G 3:1) at 40 °C. Amino acid profiles after 120 min at 40 °C (E). Glutamic acid was the only amino acid to decrease in concentration across this extended enzyme extraction.
Figure 5.
Sum of the usable amino acids, including GABA (A), proline concentration (B), glutamic acid concentration (C), and GABA concentration (D) across the extended enzyme extraction (L:G 3:1) at 40 °C. Amino acid profiles after 120 min at 40 °C (E). Glutamic acid was the only amino acid to decrease in concentration across this extended enzyme extraction.

Figure 6.
Enzyme activity extracted into the mash of the 40 °C enzyme extraction phase samples of β-amylase (A), α-amylase (B), and limit dextrinase (C). Activity was assayed as described in Section 2.5.
Figure 6.
Enzyme activity extracted into the mash of the 40 °C enzyme extraction phase samples of β-amylase (A), α-amylase (B), and limit dextrinase (C). Activity was assayed as described in Section 2.5.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ledley, A.J.; Elias, R.J.; Cockburn, D.W. Evaluating the Role of Mashing in the Amino Acid Profiles of Worts Produced from Gluten-Free Malts. Beverages 2023, 9, 10. https://doi.org/10.3390/beverages9010010
AMA Style
Ledley AJ, Elias RJ, Cockburn DW. Evaluating the Role of Mashing in the Amino Acid Profiles of Worts Produced from Gluten-Free Malts. Beverages. 2023; 9(1):10. https://doi.org/10.3390/beverages9010010
Chicago/Turabian StyleLedley, Andrew J., Ryan J. Elias, and Darrell W. Cockburn. 2023. "Evaluating the Role of Mashing in the Amino Acid Profiles of Worts Produced from Gluten-Free Malts" Beverages 9, no. 1: 10. https://doi.org/10.3390/beverages9010010
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.