
Endochitinase and Chitobiosidase Production by Marine Aeromonas caviae CHZ306: Establishment of Nitrogen Supplementation


Abstract
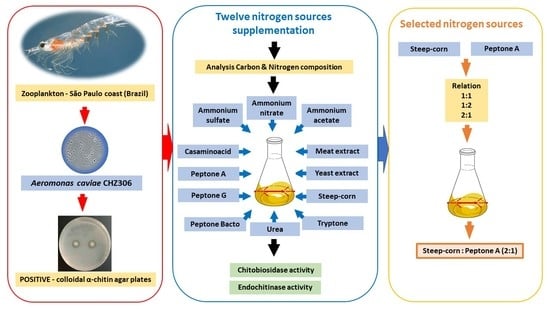
:
1. Introduction
2. Materials and Methods
2.1. Microorganisms and Growth Conditions
2.2. Evaluation of Nitrogen Supplementation in Bacterial Growth and Chitinase Production
2.3. Improvement of Endochitinase and Chitobiosidase Production by Nitrogen Source Mixture
2.4. Analytical Methods
2.4.1. Elemental Composition
2.4.2. Bacterial Growth
2.4.3. Chitinase Activity
2.5. Statistical Analysis
3. Results
3.1. Analysis of the Elemental Composition of Chitin and Nitrogenous Sources
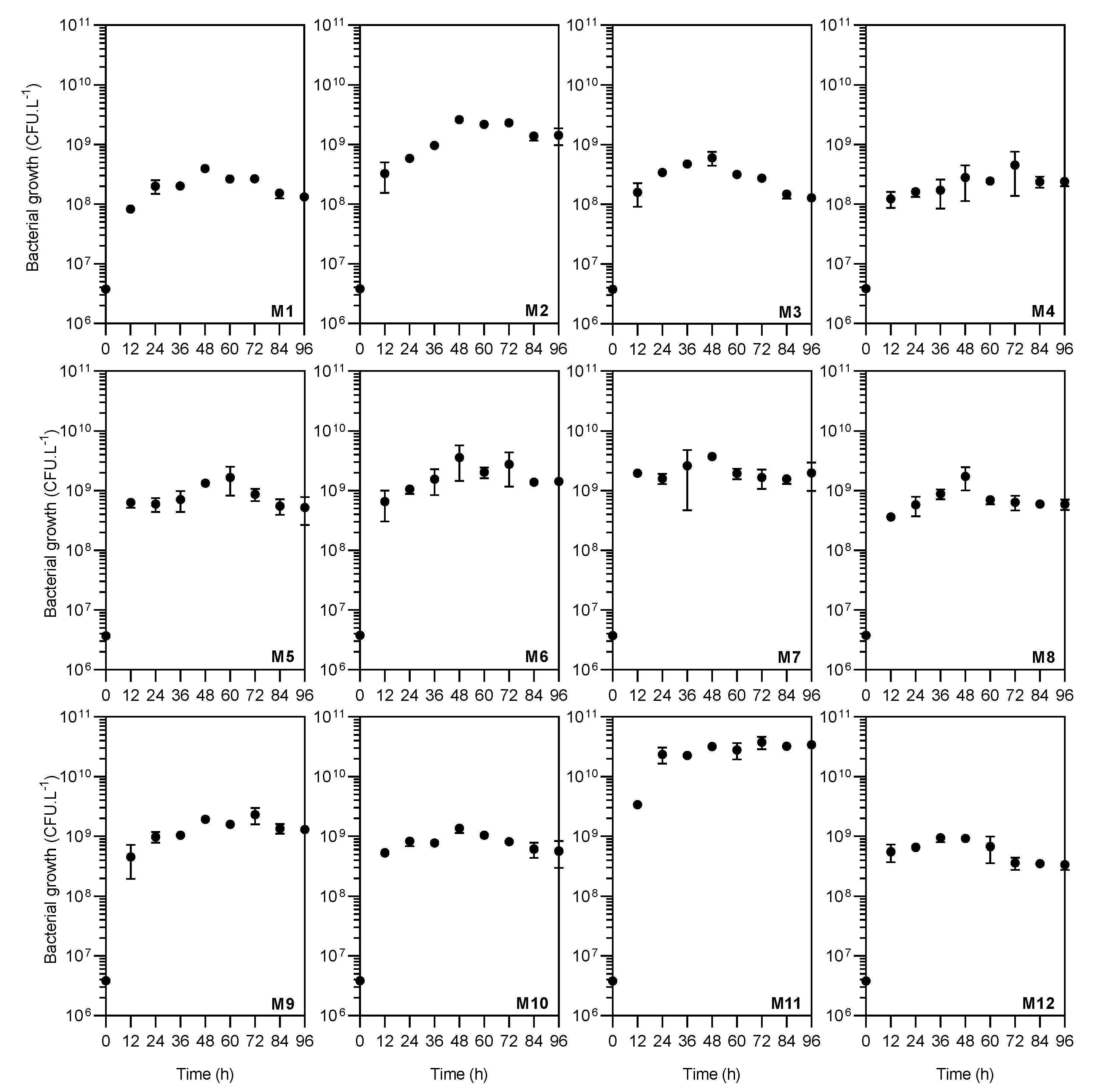
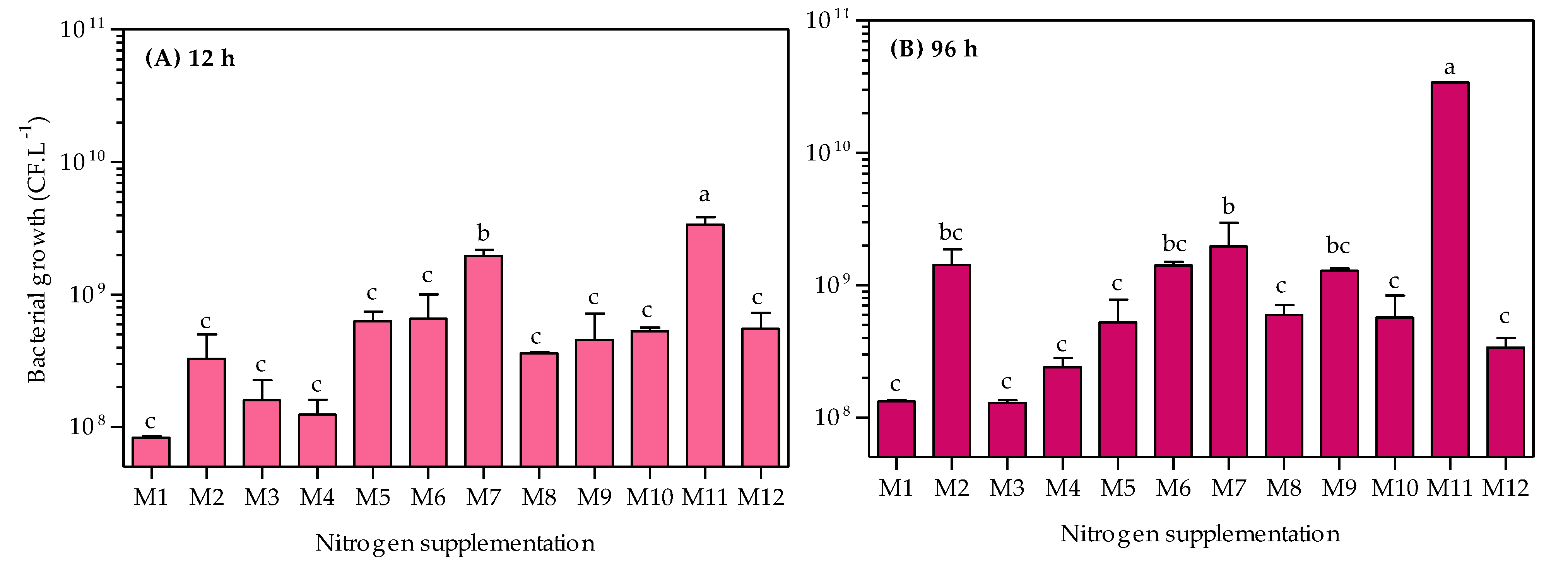
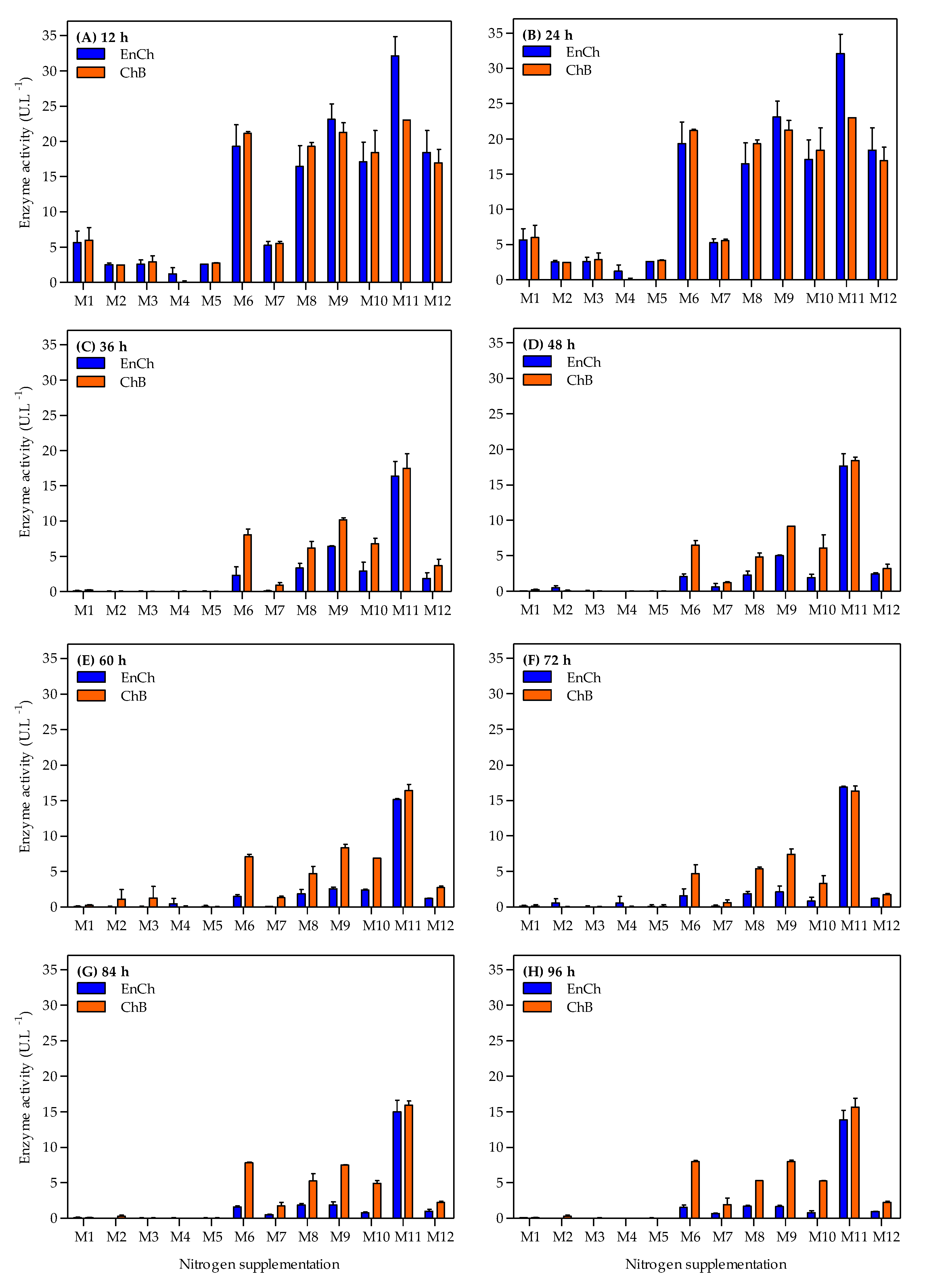
3.2. Evaluation of Nitrogen Supplementation in Bacterial Growth and Chitinase Production
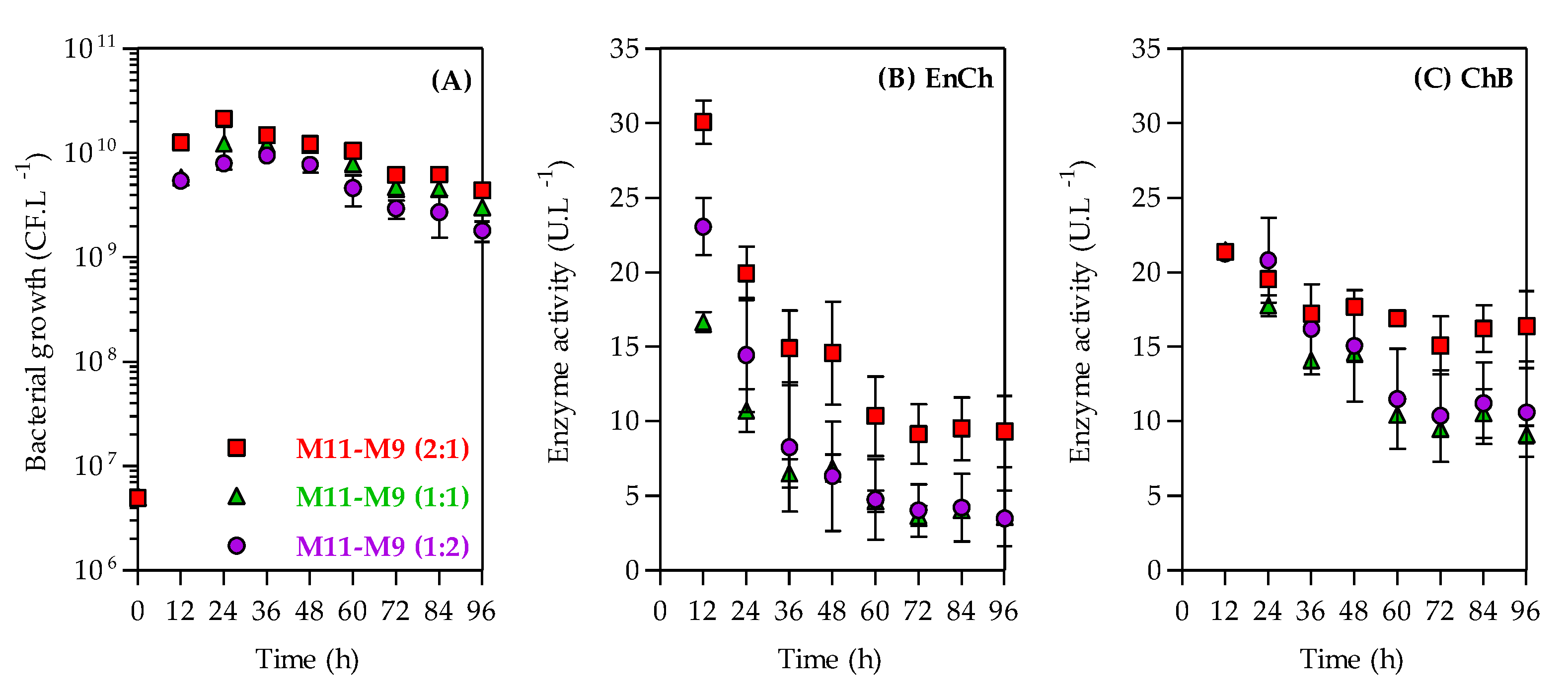
3.3. Improvement of Selected Nitrogen Sources in Endochitinase and Chitobiosidase Production
4. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Carbon Source (g.L−1) | Nitrogen Source (g.L−1) | Time Culture (h) | Chitinase Activity | Ref | |
|---|---|---|---|---|---|---|
| (U.L−1) | (U.mg−1) | |||||
| Aeromonas caviae CHZ306 | Colloidal chitin (10.0) | Corn-steep solids (1.77) Peptone A (0.51) | 12 | 30 (EnCh) 21.4 (ChB) | nr | This study |
| Aeromonas hydrophila H-2330 | Colloidal chitin (5.0) | Polypepton (5.0) Yeast extract (3.0) | 24 | 140 | 1.7 | [58] |
| Aeromonas schubertii | Colloidal chitin (10.0) | Tryptone (1.0) Yeast extract (1.0) | 96 | 155 | 0.47 | [59] |
| Aeromonas sp. GJ-18 | Swollen chitin (10.0) | Tryptone (10.0) | 120 | 1,440 | 14.4 | [19] |
| Aeromonas sp. JK1 | Colloidal chitin (7.5) | Ammonium sulfate (1.5) | 48 | 9000 * | nr | [31] |
| Aeromonas sp. ZD_05 | Colloidal chitin (10.0) | Peptone (7.0) | 72 | 10,000 * | nr | [53] |
| Aeromonas punctata HS6 | Colloidal chitin (10.0) Starch (10) | Yeast extract (10.0) | 48 | 82,640 | nr | [3] |
| Aeromonas hydrophila HS4 | Colloidal chitin (10.0) Starch (10) | Malt extract (10.0) | 24–48 | 86,010 | nr | [3] |
| Aeromonas sp. PTCC 1691 | Colloidal chitin (7.5) | Ammonium sulfate (1.5) | 48 | 92,000 | nr | [35] |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Halder, S.K.; Maity, C.; Jana, A.; Ghosh, K.; Das, A.; Paul, T.; Das Mohapatra, P.K.; Pati, B.R.; Mondal, K.C. Chitinases biosynthesis by immobilized Aeromonas hydrophila SBK1 by prawn shells valorization and application of enzyme cocktail for fungal protoplast preparation. J. Biosci. Bioeng. 2014, 117, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, M.A.; Gai, Y.; Zhuang, Q.; Wang, F.; Xiao, X.; Wang, F. Aeromonas caviae CB101 Contains Four chitinases encoded by a single gene chi1. Mol. Biotechnol. 2010, 44, 213–220. [Google Scholar] [CrossRef]
- Saima; Kuddus, M.; Roohi; Ahmad, I. Isolation of novel chitinolytic bacteria and production optimization of extracellular chitinase. J. Genet. Eng. Biotechnol. 2013, 11, 39–46. [Google Scholar] [CrossRef]
- Lan, X.; Zhang, X.; Hu, J.; Shimosaka, M. Cloning, expression, and characterization of a chitinase from the chitinolytic bacterium Aeromonas hydrophila strain SUWA-9. Biosci. Biotechnol. Biochem. 2006, 70, 2437–2442. [Google Scholar] [CrossRef] [PubMed]
- Stumpf, A.K.; Vortmann, M.; E Dirks-Hofmeister, M.; Moerschbacher, B.M.; Philipp, B. Identification of a novel chitinase from Aeromonas hydrophila AH-1N for the degradation of chitin within fungal mycelium. FEMS Microbiol. Lett. 2019, 366, fny294. [Google Scholar] [CrossRef]
- Pentekhina, I.; Hattori, T.; Tran, D.M.; Shima, M.; Watanabe, T.; Sugimoto, H.; Suzuki, K. Chitinase system of Aeromonas salmonicida, and characterization of enzymes involved in chitin degradation. Biosci. Biotechnol. Biochem. 2020, 84, 1936–1947. [Google Scholar] [CrossRef]
- Jeong, H.C.; Ju, W.-T.; Jo, K.-H.; Park, R.D. Purification and characterization of a 34-kDa chitobiosidase from Aeromonas sp. GJ-18. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 7–12. [Google Scholar] [CrossRef]
- Dahiya, N.; Tewari, R.; Hoondal, G.S. Biotechnological aspects of chitinolytic enzymes: A review. Appl. Microbiol. Biotechnol. 2006, 71, 773–782. [Google Scholar] [CrossRef]
- Cardozo, F.A.; Gonzalez, J.M.; Feitosa, V.A.; Pessoa, A.; Rivera, I.N.G. Bioconversion of α-chitin into N-acetyl-glucosamine using chitinases produced by marine-derived Aeromonas caviae isolates. World J. Microbiol. Biotechnol. 2017, 33, 201. [Google Scholar] [CrossRef]
- Kumar, M.; Rajput, M.; Soni, T.; Vivekanand, V.; Pareek, N. Chemoenzymatic Production and Engineering of Chitooligosaccharides and N-acetyl Glucosamine for Refining Biological Activities. Front. Chem. 2020, 8, 469. [Google Scholar] [CrossRef]
- Ren, X.-B.; Dang, Y.-R.; Liu, S.-S.; Huang, K.-X.; Qin, Q.-L.; Chen, X.-L.; Zhang, Y.-Z.; Wang, Y.-J.; Li, P.-Y. Identification and Characterization of Three Chitinases with Potential in Direct Conversion of Crystalline Chitin into N,N′-diacetylchitobiose. Mar. Drugs 2022, 20, 165. [Google Scholar] [CrossRef]
- Osada, M.; Kikuta, K.; Yoshida, K.; Totani, K.; Ogata, M.; Usui, T. Non-catalytic dehydration of N,N′-diacetylchitobiose in high-temperature water. RSC Adv. 2014, 4, 33651–33657. [Google Scholar] [CrossRef] [Green Version]
- Brzezinska, M.S.; Jankiewicz, U.M.; Burkowska-But, A.; Walczak, M. Chitinolytic microorganisms and their possible application in environmental protection. Curr. Microbiol. 2014, 68, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Bravo, A.; Figueras, M.J. An update on the genus Aeromonas: Taxonomy, epidemiology, and pathogenicity. Microorganisms 2020, 8, 129. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, M.; Mishra, A.; Prasad, A.; Azim, A.; Singh, R.; Baronia, A.; Prasad, K.; Dwivedi, U. Aeromonas caviae septicemia in immunocompetent gastrointestinal carriers. Braz. J. Infect. Dis. 2008, 12, 47–548. [Google Scholar] [CrossRef] [Green Version]
- Janda, J.M.; Abbott, S.L. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canellas, A.L.B.; Laport, M.S. The biotechnological potential of Aeromonas: A bird’s eye view. Crit. Rev. Biotechnol. 2022, 1–13. [Google Scholar] [CrossRef]
- Cardozo, F.A.; Facchinatto, W.M.; Colnago, L.A.; Campana-Filho, S.P.; Pessoa, A. Bioproduction of N-acetyl-glucosamine from colloidal α-chitin using an enzyme cocktail produced by Aeromonas caviae CHZ306. World J. Microbiol. Biotechnol. 2019, 35, 114. [Google Scholar] [CrossRef]
- Kuk, J.H.; Jung, W.J.; Jo, G.H.; Ahn, J.S.; Kim, K.Y.; Park, R.D. Selective preparation of N-acetyl-D-glucosamine and N,N′- diacetylchitobiose from chitin using a crude enzyme preparation from Aeromonas sp. Biotechnol. Lett. 2005, 27, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Pillaca-Pullo, O.S.; Lopes, A.M.; Rodriguez-Portilla, L.M.; Estela-Escalante, W. Optimizing medium composition with wastewater from Coffea arabica processing to produce single-cell protein using Candida sorboxylosa. J. Chem. Technol. Biotechnol. 2023, 98, 106–116. [Google Scholar] [CrossRef]
- Yang, X.; Yang, Y.; Huang, J.; Man, D.; Guo, M. Comparisons of urea or ammonium on growth and fermentative metabolism of Saccharomyces cerevisiae in ethanol fermentation. World J. Microbiol. Biotechnol. 2021, 37, 71. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, P.; Chen, Y.; Cao, Q.; Liu, X.; Li, D. The production of single cell protein from biogas slurry with high ammonia-nitrogen content by screened Nectaromyces rattus. Poult. Sci. 2021, 100, 101334. [Google Scholar] [CrossRef]
- Marius, K.S.; Mahamadi, N.; Ibrahim, K.; Iliassou, M.; Sonagnon, H.S.K.; Yerobessor, D.; Wahauwouele, H.C.; Essodolom, T.; Alfred, S. Production of single cell protein (SCP) and essentials amino acids from Candida utilis FMJ12 by solid state fermentation using mango waste supplemented with nitrogen sources. Afr. J. Biotechnol. 2018, 17, 716–723. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Seguinot, P.; Sanchez, I.; Ortiz-Julien, A.; Heras, J.M.; Querol, A.; Camarasa, C.; Guillamón, J.M. Nitrogen sources preferences of non-Saccharomyces yeasts to sustain growth and fermentation under winemaking conditions. Food Microbiol. 2020, 85, 103287. [Google Scholar] [CrossRef]
- Roca-Mesa, H.; Sendra, S.; Mas, A.; Beltran, G.; Torija, M.-J. Nitrogen preferences during alcoholic fermentation of different non-saccharomyces yeasts of oenological interest. Microorganisms 2020, 8, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broach, J.R. Nutritional control of growth and development in yeast. Genetics 2012, 192, 73–105. [Google Scholar] [CrossRef] [Green Version]
- Linder, T. Nitrogen assimilation pathways in budding yeasts. In Non-Conventional Yeasts: From Basic Research to Application; Springer: Cham, Switzerland, 2019; pp. 197–236. [Google Scholar]
- Djellouli, M.; Martínez-Álvarez, O.; Arancibia, M.Y.; Florez-Cuadrado, D.; Ugarte-Ruíz, M.; Domínguez, L.; Zadi-Karam, H.; Karam, N.; Roudj, S.; López-Caballero, M.E. Effect of seafood peptones on biomass and metabolic activity by Enterococcus faecalis DM19. LWT Food Sci. Technol. 2017, 81, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Martí, E.; Del Olmo, M. Addition of ammonia or amino acids to a nitrogen-depleted medium affects gene expression patterns in yeast cells during alcoholic fermentation. FEMS Yeast Res. 2008, 8, 245–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, K.G. Microbiology. In Bioprocess Engineering: An Introductory Engineering and Life Science Approach; Woodhead Publishing: Cambridge, UK, 2013; pp. 7–24. [Google Scholar]
- Al-Ahmadi, K.J.; Yazdi, M.T.; Najafi, M.F.; Shahverdi, A.R.; Faramarzi, M.A.; Zarrini, G.; Behravan, J. Optimization of medium and cultivation conditions for chitinase production by newly isolated: Aeromonas sp. Biotechnology 2008, 7, 266–272. [Google Scholar]
- Souza, C.P.; Burbano-Rosero, E.M.; Almeida, B.C.; Martins, G.G.; Albertini, L.S.; Rivera, I.N.G. Culture medium for isolating chitinolytic bacteria from seawater and plankton. World J. Microbiol. Biotechnol. 2009, 25, 2079–2082. [Google Scholar] [CrossRef]
- Cardozo, F.A.; Zimpel, C.K.; Guimaraes, A.; Pessoa, A.; Rivera, I.N.G. Draft genome sequence of marine-derived Aeromonas caviae CHZ306, a potential chitinase producer strain. Genome Announc. 2016, 4, e01293-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, H.; Rodrigues, M.; Campos, C.; Chaves, M.; Nunes, I.; Juliano, Y.; Novo, N. Counting of viable cluster-forming and non cluster-forming bacteria: A comparison between the drop and the spread methods. J. Microbiol. Methods 1995, 22, 39–50. [Google Scholar] [CrossRef]
- Jamialahma, K.; Behravan, J.; Najafi, M.F.; Yazdi, M.T.; Shahverdi, A.; Faramarzi, M.A. Enzymatic production of N-acetyl-D-glucosamine from chitin using crude enzyme preparation of Aeromonas sp. PTCC1691. Biotechnology 2011, 10, 292–297. [Google Scholar] [CrossRef] [Green Version]
- Dinarvand, M.; Rezaee, M.; Masomian, M.; Jazayeri, S.D.; Zareian, M.; Abbasi, S.; Ariff, A.B. Effect of C/N ratio and media optimization through response surface methodology on simultaneous productions of intra- and extracellular inulinase and invertase from aspergillus Niger ATCC 20611. Biomed Res. Int. 2013, 2013, 508968. [Google Scholar] [CrossRef] [Green Version]
- Moguel, I.S.; Yamakawa, C.K.; Brumano, L.P.; Pessoa, A.; Mussatto, S.I. Selection and Optimization of Medium Components for the Efficient Production of L-Asparaginase by Leucosporidium scottii L115—A Psychrotolerant Yeast. Fermentation 2022, 8, 398. [Google Scholar] [CrossRef]
- Da Cruz, S.H.; Cilli, E.M.; Ernandes, J.R. Structural complexity of the nitrogen source and influence on yeast growth and fermentation. J. Inst. Brew. 2002, 108, 54–61. [Google Scholar] [CrossRef]
- Raita, S.; Kusnere, Z.; Spalvins, K.; Blumberga, D. Optimization of Yeast Cultivation Factors for Improved SCP Production. Environ. Clim. Technol. 2022, 26, 848–861. [Google Scholar] [CrossRef]
- Prins, R.C.; Billerbeck, S. A buffered media system for yeast batch culture growth. BMC Microbiol. 2021, 21, 127. [Google Scholar] [CrossRef] [PubMed]
- Konermann, L. Addressing a Common Misconception: Ammonium Acetate as Neutral pH ‘Buffer’ for Native Electrospray Mass Spectrometry. J. Am. Soc. Mass Spectrom. 2017, 28, 1827–1835. [Google Scholar] [CrossRef] [PubMed]
- Hosseany, E.N.; Elhamid, S.M.A.; Khair, A.G.A.-E.; Azzaz, H.; Zahran, M.O. Utilization of hydrolyzed UF-permeate Supplemented with different nitrogen sources and vitamins for production of Baker’s yeast. Biotechnology 2019, 18, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Taabodi, M.; May, E.B.; Bryant, R.B.; Saporito, L.S.; Skeen, O.K.; Hashem, F.M.; Allen, A.L. Aeromonas hydrophila, Bacillus thuringiensis, Escherichia coli and Pseudomonas aeruginosa utilization of Ammonium-N, Nitrate-N and Urea-N in culture. Heliyon 2020, 6, e03711. [Google Scholar] [CrossRef] [PubMed]
- Becton, D.A.C. BD BionutrientsTM Technical Manual—Advanced Bioprocessing. Available online: http://static.bdbiosciences.com/eu/documents/bionutrients_tech_manual.pdf (accessed on 20 December 2022).
- Puhm, M.; Ainelo, H.; Kivisaar, M.; Teras, R. Tryptone in Growth Media Enhances Pseudomonas putida Biofilm. Microorganisms 2022, 10, 618. [Google Scholar] [CrossRef] [PubMed]
- Malairuang, K.; Krajang, M.; Sukna, J.; Rattanapradit, K.; Chamsart, S. High cell density cultivation of Saccharomyces cerevisiae with intensive multiple sequential batches together with a novel technique of fed-batch at cell level (FBC). Processes 2020, 8, 1321. [Google Scholar] [CrossRef]
- Tanguler, H.; Erten, H. Utilisation of spent brewer’s yeast for yeast extract production by autolysis: The effect of temperature. Food Bioprod. Process. 2008, 86, 317–321. [Google Scholar] [CrossRef]
- Hahn-Hägerdal, B.; Karhumaa, K.; Larsson, C.U.; Gorwa-Grauslund, M.; Görgens, J.; van Zyl, W.H. Role of cultivation media in the development of yeast strains for large scale industrial use. Microb. Cell Factories 2005, 4, 31. [Google Scholar] [CrossRef] [Green Version]
- Jan, D.C.-H.; Jones, S.J.; Emery, A.N.; Al-Rubeai, M. Peptone, a low-cost growth-promoting nutrient for intensive animal cell culture. Cytotechnology 1994, 16, 17–26. [Google Scholar] [CrossRef]
- Gray, V.; Müller, C.; Watkins, I.; Lloyd, D. Peptones from diverse sources: Pivotal determinants of bacterial growth dynamics. J. Appl. Microbiol. 2008, 104, 554–565. [Google Scholar] [CrossRef]
- Saxena, J.; Tanner, R.S. Optimization of a corn steep medium for production of ethanol from synthesis gas fermentation by Clostridium ragsdalei. World J. Microbiol. Biotechnol. 2012, 28, 1553–1561. [Google Scholar] [CrossRef]
- Wang, G.; Shi, B.; Zhang, P.; Zhao, T.; Yin, H.; Qiao, C. Effects of corn steep liquor on β-poly(l-malic acid) production in Aureobasidium melanogenum. AMB Express 2020, 10, 211. [Google Scholar] [CrossRef]
- Ghasemi, Y.; Dehdari, Z.; Mohkam, M.; Kargar, M. Isolation and optimization of cultivation conditions for production of chitinase by Aeromonas sp. ZD-05 from the Persian Gulf. J. Pure Appl. Microbiol. 2013, 7, 913–918. [Google Scholar]
- Kennedy, M.; Krouse, D. Strategies for improving fermentation medium performance: A review. J. Ind. Microbiol. Biotechnol. 1999, 23, 456–475. [Google Scholar] [CrossRef]
- Arous, F.; Azabou, S.; Jaouani, A.; Zouari-Mechichi, H.; Nasri, M.; Mechichi, T. Biosynthesis of single-cell biomass from olive mill wastewater by newly isolated yeasts. Environ. Sci. Pollut. Res. 2016, 23, 6783–6792. [Google Scholar] [CrossRef]
- Jung, W.-J.; Park, R.-D. Bioproduction of chitooligosaccharides: Present and perspectives. Mar. Drugs 2014, 12, 5328–5356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, C.; Antelo, L.T.; Franco-Uría, A.; Alonso, A.A.; Pérez-Martín, R. Chitin production from crustacean biomass: Sustainability assessment of chemical and enzymatic processes. J. Clean. Prod. 2018, 172, 4140–4151. [Google Scholar] [CrossRef] [Green Version]
- Hiraga, K.; Shou, L.; Kitazawa, M.; Takahashi, S.; Shimada, M.; Sato, R.; Oda, K. Isolation and characterization of chitinase from a flake-chitin degrading marine bacterium, Aeromonas hydrophila H-2330. Biosci. Biotechnol. Biochem. 1997, 61, 174–176. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.-H.; Chen, J.-K.; Lee, W.-C. Purification and characterization of extracellular chitinase from Aeromonas schubertii. Enzym. Microb. Technol. 2004, 35, 550–556. [Google Scholar] [CrossRef]





| Medium | Nutrient | Manufacturer | Chemical Formula | C (%) | N (%) | Concentration | Final C:N Ratio ** | ||
|---|---|---|---|---|---|---|---|---|---|
| (g.L−1) * | C (g.L−1) | N (g.L−1) | |||||||
| M1–M12 | Colloid α-chitin | Sigma-Aldrich, Saint Louis, MO, USA | (C8H13O5N)n | 44.0 | 0.60 | 10.0 | 4.400 | 0.060 | nc |
| M1 | Ammonium sulfate | Sigma-Aldrich, Saint Louis, MO, USA | (NH4)2SO4 | 0.0 | 21.2 | 1.00 | 0.000 | 0.212 | 16.2 |
| M2 | Ammonium acetate | Labsynth, Diadema, SP, Brazil | NH4CH3CO2 | 15.6 | 18.2 | 1.17 | 0.183 | 0.213 | 16.8 |
| M3 | Ammonium nitrate | Labsynth, Diadema, SP, Brazil | NH4NO3 | 0.0 | 35.0 | 0.61 | 0.000 | 0.214 | 16.1 |
| M4 | Urea | Labsynth, Diadema, SP, Brazil | CH4N2O | 20.0 | 46.6 | 0.45 | 0.090 | 0.210 | 16.6 |
| M5 | Casaminoacid | BD Biosciences, San Jose, CA, USA | nd | 33.3 | 10.7 | 1.97 | 0.656 | 0.211 | 18.7 |
| M6 | Meat extract | Acumedia, Lansing, MI, USA | nd | 41.9 | 12.5 | 1.70 | 0.712 | 0.213 | 18.7 |
| M7 | Yeast extract | BD Biosciences, San Jose, CA, USA | nd | 38.9 | 10.9 | 1.94 | 0.755 | 0.211 | 19.0 |
| M8 | Peptone Bacto | BD Biosciences, San Jose, CA, USA | nd | 43.3 | 14.9 | 1.42 | 0.615 | 0.212 | 18.4 |
| M9 | Peptone A | Acumedia, Lansing, MI, USA | nd | 41.8 | 13.2 | 1.61 | 0.673 | 0.213 | 18.6 |
| M10 | Peptone G | Acumedia, Lansing, MI, USA | nd | 44.3 | 16.0 | 1.32 | 0.585 | 0.211 | 18.4 |
| M11 | Corn-steep solids | Sigma-Aldrich, Saint Louis, MO, USA | nd | 37.6 | 7.6 | 2.81 | 1.057 | 0.214 | 19.9 |
| M12 | Tryptone | BD Biosciences, San Jose, CA, USA | nd | 43.5 | 12.7 | 1.67 | 0.726 | 0.212 | 18.8 |
| Medium (N Ratio) * | Corn-Steep Solids (M11) | Peptone A (M9) | Final C:N Ratio ** | ||||
|---|---|---|---|---|---|---|---|
| (g.L−1) | (C, g.L−1) | (N, g.L−1) | (g.L−1) | (C, g.L−1) | (N, g.L−1) | ||
| M11–M9 (1:1) | 1.33 | 0.500 | 0.101 | 0.76 | 0.318 | 0.100 | 20.0 |
| M11–M9 (1:2) | 0.94 | 0.353 | 0.071 | 1.01 | 0.422 | 0.133 | 19.6 |
| M11–M9 (2:1) | 1.77 | 0.665 | 0.135 | 0.51 | 0.213 | 0.067 | 26.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardozo, F.; Feitosa, V.; Pillaca-Pullo, O.; Pessoa, A. Endochitinase and Chitobiosidase Production by Marine Aeromonas caviae CHZ306: Establishment of Nitrogen Supplementation. Bioengineering 2023, 10, 431. https://doi.org/10.3390/bioengineering10040431
Cardozo F, Feitosa V, Pillaca-Pullo O, Pessoa A. Endochitinase and Chitobiosidase Production by Marine Aeromonas caviae CHZ306: Establishment of Nitrogen Supplementation. Bioengineering. 2023; 10(4):431. https://doi.org/10.3390/bioengineering10040431
Chicago/Turabian StyleCardozo, Flavio, Valker Feitosa, Omar Pillaca-Pullo, and Adalberto Pessoa. 2023. "Endochitinase and Chitobiosidase Production by Marine Aeromonas caviae CHZ306: Establishment of Nitrogen Supplementation" Bioengineering 10, no. 4: 431. https://doi.org/10.3390/bioengineering10040431