
Protective Role of Kelulut Honey against Toxicity Effects of Polystyrene Microplastics on Morphology, Hormones, and Sex Steroid Receptor Expression in the Uterus of Rats
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Kelulut Honey (Agromas, Malaysia)
2.2. Animal Model and Experimental Design
- (1)
- Normal control group (NC), treated with vehicle (0.2 mL deionized water);
- (2)
- Microplastic control group (M), treated with polystyrene microplastic (PS-MPs) at 2.5 mg/kg body weight;
- (3)
- Kelulut honey dose + MPs group (DM), pretreated with 1200 mg/kg Kelulut honey for 30 minutes before administrated of PS-MPs at 2.5 mg/kg body weight;
- (4)
- Kelulut honey control group (DC), treated with 1200 mg/kg body weight of Kelulut honey only.
2.3. Histopathological and Histomorphometric Analysis of the Uterus
2.4. Assessment of Serum Estrogen, Progesterone, Follicle-Stimulating Hormones (FSHs), and Luteinizing Hormones (LHs) Levels
2.5. mRNA Expression of ER-α and ER-β
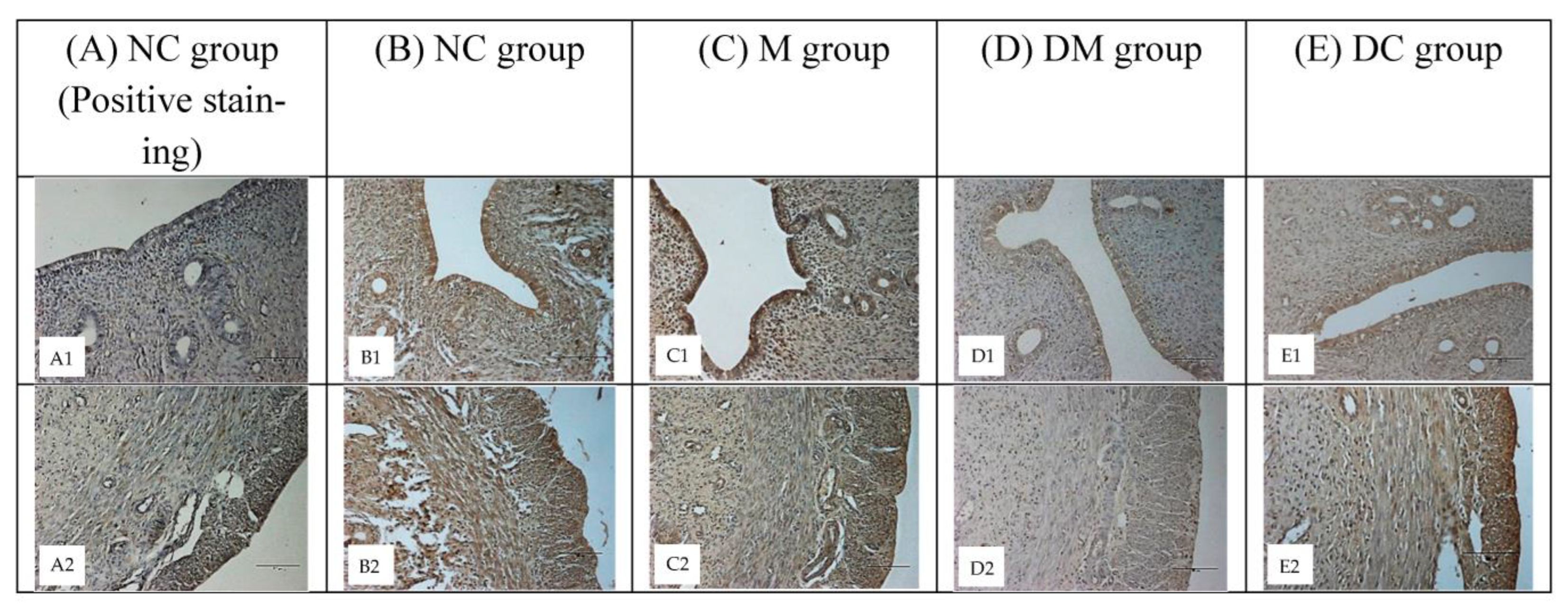
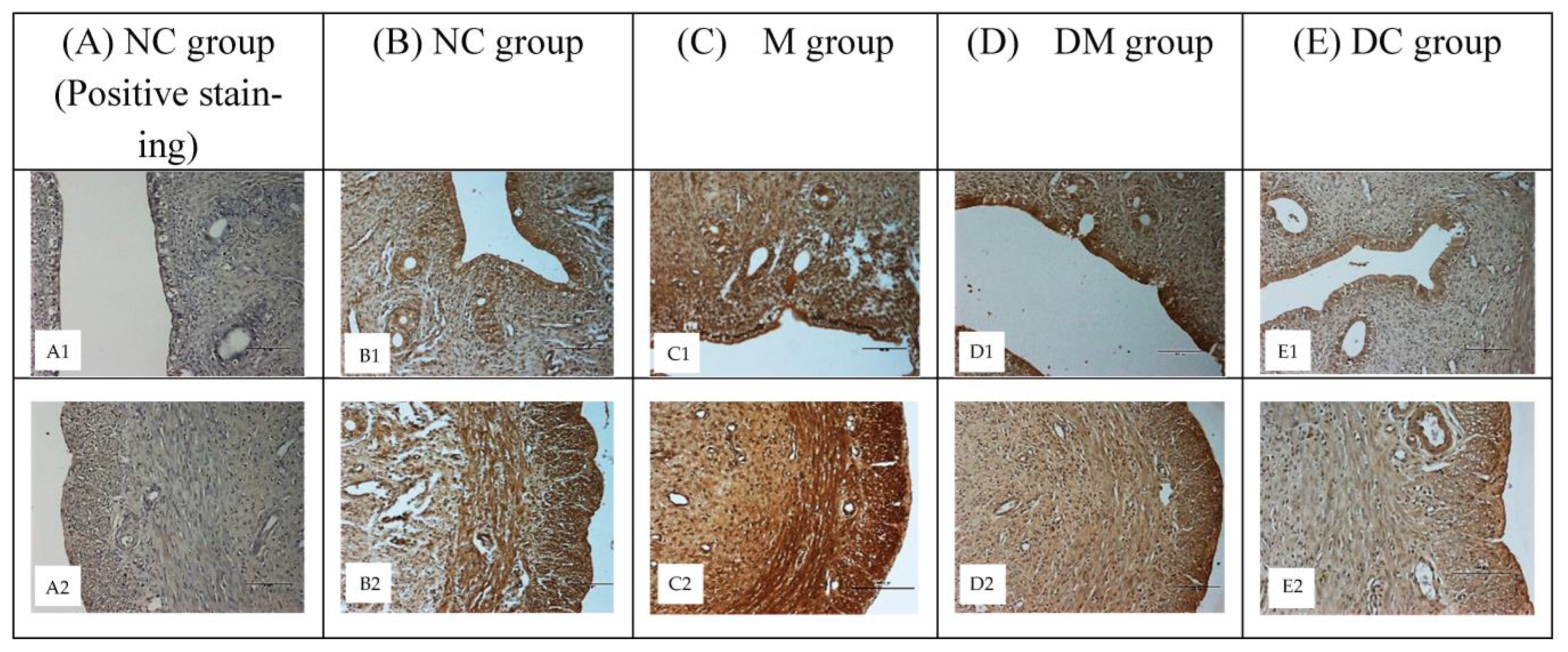
2.6. Immunohistochemistry
2.7. Statistical Analysis
3. Results
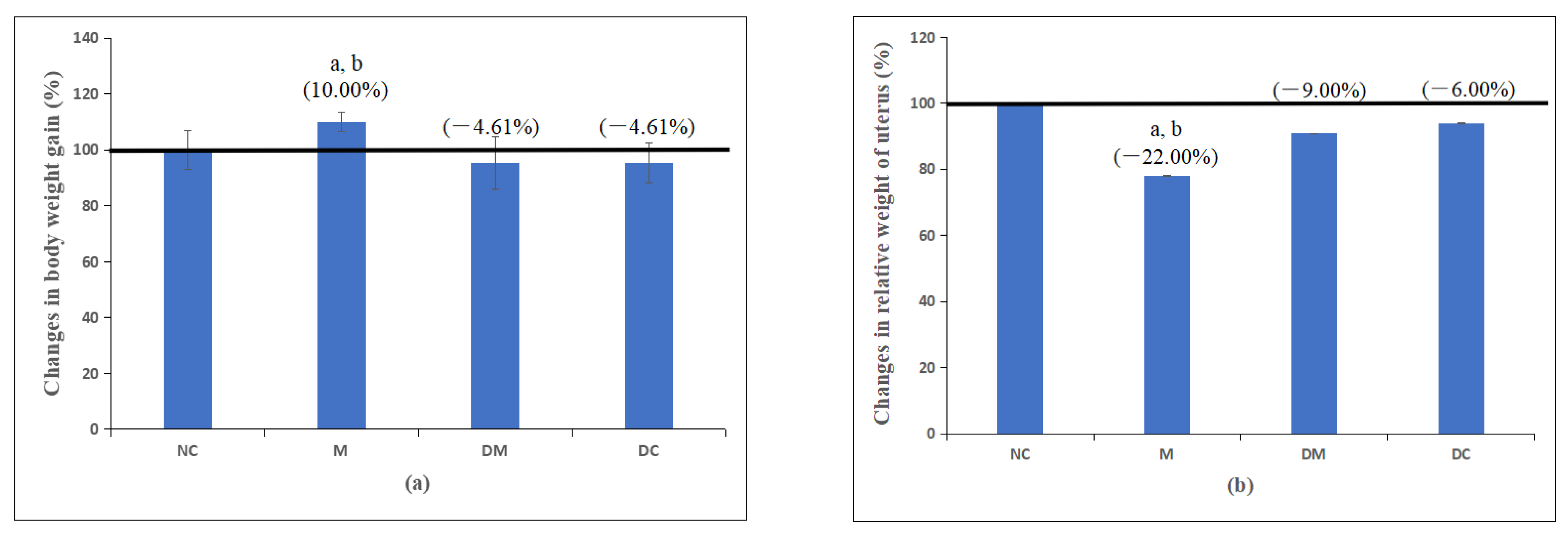
3.1. Body Weight and Uterine Weight
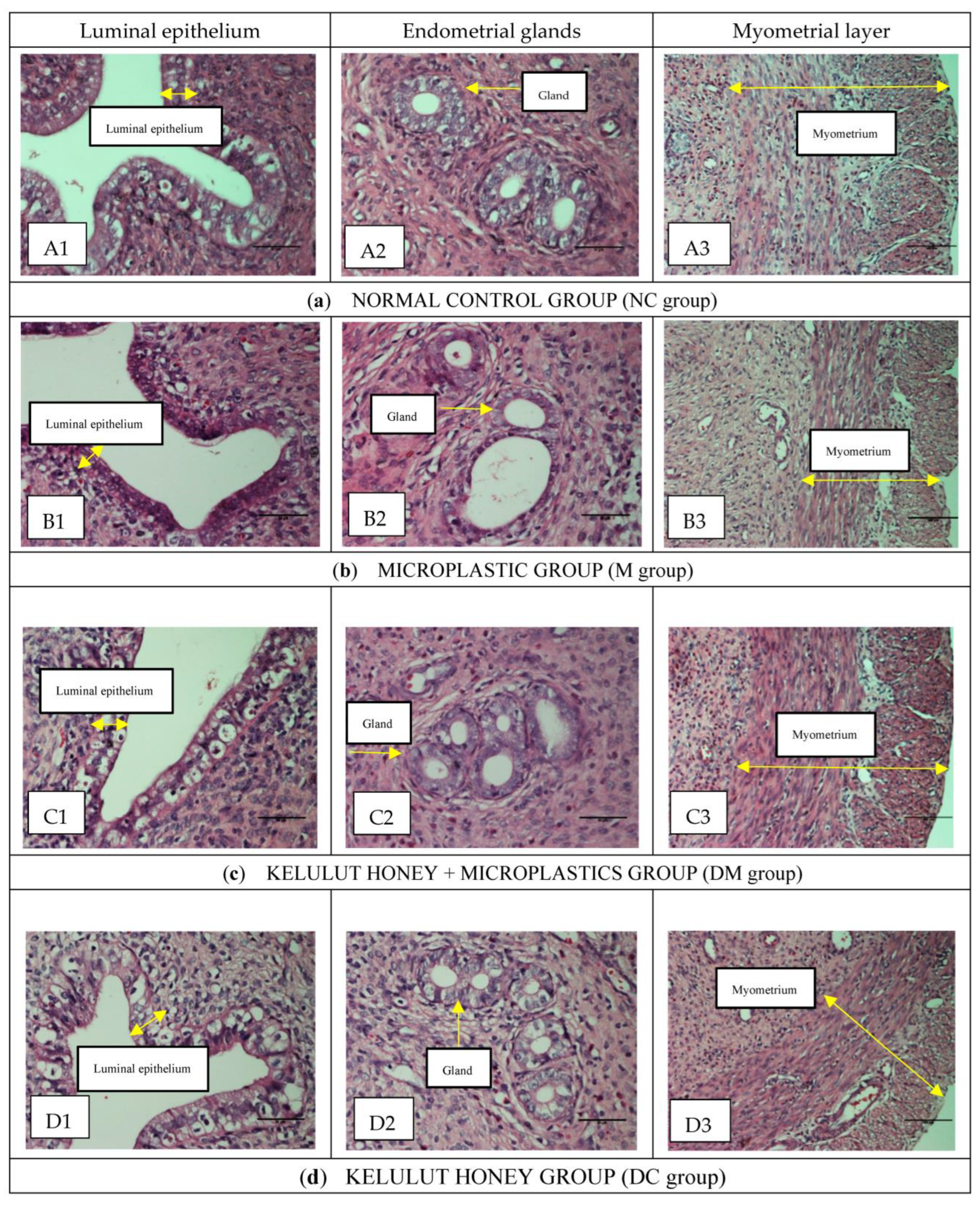
3.2. Histopathological Examination and Histomorphometry Analysis
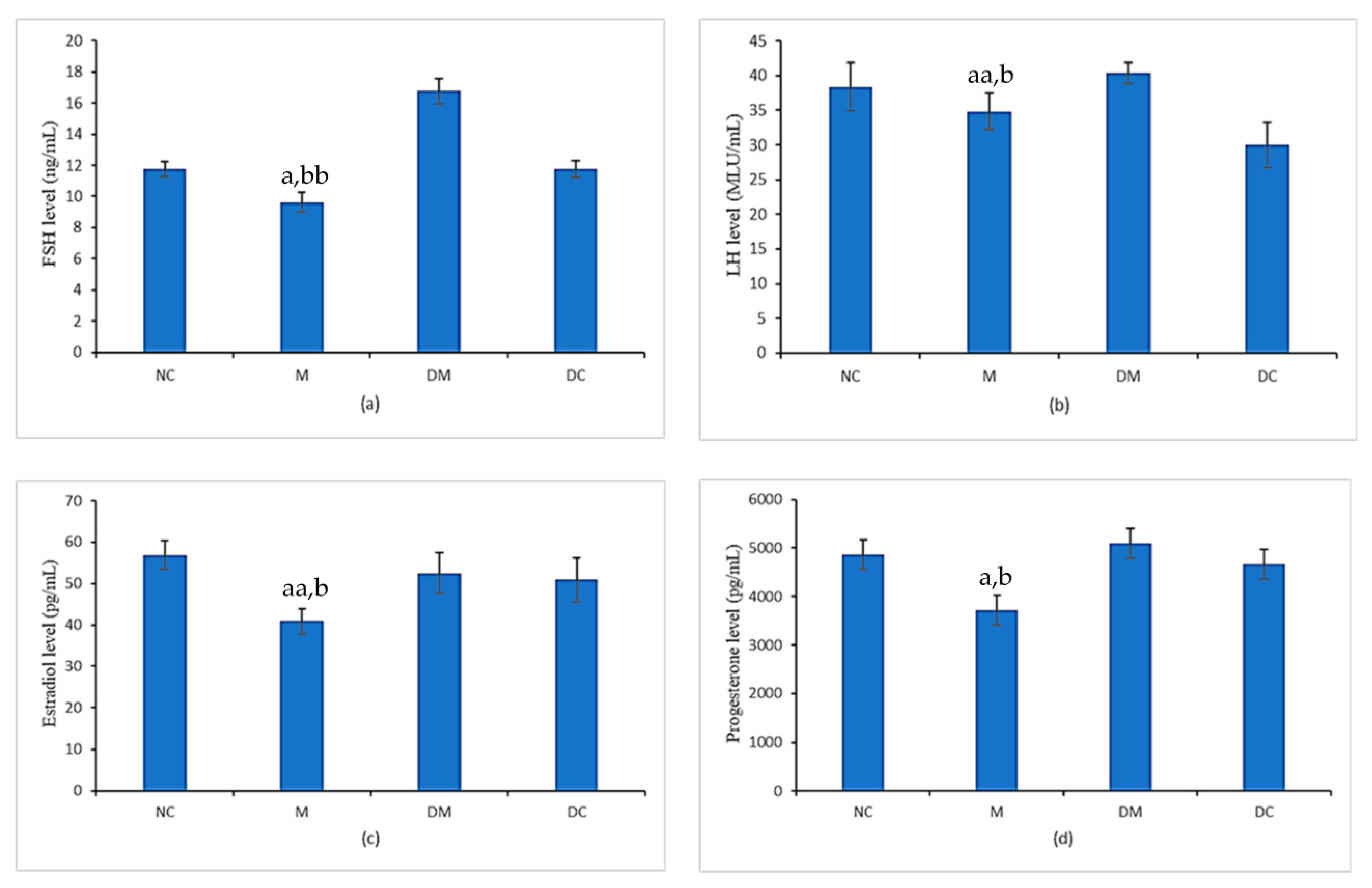
3.3. Gonadotropins Hormones (FSH and LH) and Sex Steroid Hormones (17β-Estradiol and Progesterone)
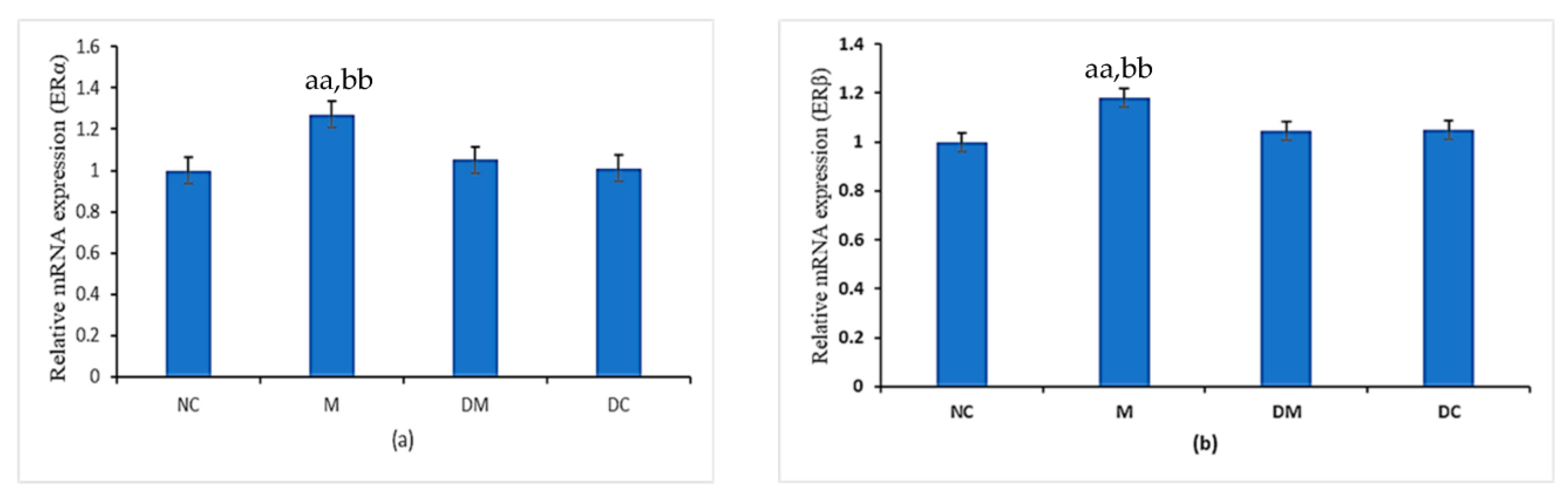
3.4. ERα and ERβ Expression
3.5. ERα and ERβ Protein Distribution
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amaral-Zettler, L.A.; Zettler, E.R.; Slikas, B.; Boyd, G.D.; Melvin, D.W.; Morrall, C.E.; Proskurowski, G.; Mincer, T.J. The biogeography of the Plastisphere: Implications for policy. Front. Ecol. Environ. 2015, 13, 541–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avio, C.G.; Gorbi, S.; Milan, M.; Benedetti, M.; Fattorini, D.; d'Errico, G.; Pauletto, M.; Bargelloni, L.; Regoli, F. Pollutants bioavailability and toxicological risk from microplastics to marine mussels. Environ. Pollut. 2015, 198, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Van Cauwenberghe, L.; Janssen, C.R. Microplastics in bivalves cultured for human consumption. Environ. Pollut. 2014, 193, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Huerta, E.; Mendoza Vega, J.; Quej, V.K.; Chi, J.d.l.A.; Sanchez del Cid, L.; Quijano, C.; Escalona-Segura, G.; Gertsen, H.; Salánki, T.; van der Ploeg, M.; et al. Bioaccumulation of microplastics in the terrestrial food chain: An example from home gardens in SE Mexico. In Proceedings of the EGU General Assembly Conference Abstracts, Vienna, Austria, 23–28 April 2017; p. 15847. [Google Scholar]
- Karami, A.; Golieskardi, A.; Ho, Y.B.; Larat, V.; Salamatinia, B. Microplastics in eviscerated flesh and excised organs of dried fish. Sci. Rep. 2017, 7, 5473. [Google Scholar] [CrossRef] [Green Version]
- Pivokonsky, M.; Cermakova, L.; Novotna, K.; Peer, P.; Cajthaml, T.; Janda, V. Occurrence of microplastics in raw and treated drinking water. Sci. Total Environ. 2018, 643, 1644–1651. [Google Scholar] [CrossRef]
- Murphy, F.; Quinn, B. The effects of microplastic on freshwater Hydra attenuata feeding, morphology & reproduction. Environ. Pollut 2018, 234, 487–494. [Google Scholar] [CrossRef] [Green Version]
- Assas, M.; Qiu, X.; Chen, K.; Ogawa, H.; Xu, H.; Shimasaki, Y.; Oshima, Y. Bioaccumulation and reproductive effects of fluorescent microplastics in medaka fish. Mar. Pollut. Bull. 2020, 158, 111446. [Google Scholar] [CrossRef]
- Sussarellu, R.; Suquet, M.; Thomas, Y.; Lambert, C.; Fabioux, C.; Pernet, M.E.J.; Le Goïc, N.; Quillien, V.; Mingant, C.; Epelboin, Y.; et al. Oyster reproduction is affected by exposure to polystyrene microplastics. Proc. Natl. Acad. Sci. USA 2016, 113, 2430–2435. [Google Scholar] [CrossRef] [Green Version]
- Hou, B.; Wang, F.; Liu, T.; Wang, Z. Reproductive toxicity of polystyrene microplastics: In vivo experimental study on testicular toxicity in mice. J. Hazard. Mater. 2021, 405, 124028. [Google Scholar] [CrossRef]
- Xie, X.; Deng, T.; Duan, J.; Xie, J.; Yuan, J.; Chen, M. Exposure to polystyrene microplastics causes reproductive toxicity through oxidative stress and activation of the p38 MAPK signaling pathway. Ecotoxicol. Environ. Saf. 2020, 190, 110133. [Google Scholar] [CrossRef]
- Amereh, F.; Babaei, M.; Eslami, A.; Fazelipour, S.; Rafiee, M. The emerging risk of exposure to nano(micro)plastics on endocrine disturbance and reproductive toxicity: From a hypothetical scenario to a global public health challenge. Environ. Pollut. 2020, 261, 114158. [Google Scholar] [CrossRef]
- Zaid, S.S.; Sulaiman, S.A.; Sirajudeen, K.N.; Othman, N.H. The effects of Tualang honey on female reproductive organs, tibia bone and hormonal profile in ovariectomised rats--animal model for menopause. BMC Complement. Altern. Med. 2010, 10, 82. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Wang, Y.; Wang, S.; Xie, J.; Han, Q.; Chen, M. Comparing the effects of polystyrene microplastics exposure on reproduction and fertility in male and female mice. Toxicology 2022, 465, 153059. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, H.A.; Santambrosio, N.; Santamaría, C.G.; Muñoz-de-Toro, M.; Luque, E.H. Neonatal exposure to bisphenol A reduces the pool of primordial follicles in the rat ovary. Reprod. Toxicol. 2010, 30, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Lei, Z.; Cui, L.; Hou, Y.; Yang, L.; An, R.; Wang, Q.; Li, S.; Zhang, H.; Zhang, L. Polystyrene microplastics lead to pyroptosis and apoptosis of ovarian granulosa cells via NLRP3/Caspase-1 signaling pathway in rats. Ecotoxicol. Environ. Saf. 2021, 212, 112012. [Google Scholar] [CrossRef] [PubMed]
- Kek, S.P.; Chin, N.L.; Yusof, Y.A.; Tan, S.W.; Chua, L.S. Total Phenolic Contents and Colour Intensity of Malaysian Honeys from the Apis spp. and Trigona spp. Bees. Agric. Agric. Sci. Procedia 2014, 2, 150–155. [Google Scholar] [CrossRef] [Green Version]
- Zainol, M.I.; Mohd Yusoff, K.; Mohd Yusof, M.Y. Antibacterial activity of selected Malaysian honey. BMC Complement. Altern. Med. 2013, 13, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordin, A.; Omar, N.; Sainik, N.Q.A.V.; Chowdhury, S.R.; Omar, E.; Bin Saim, A.; Bt Hj Idrus, R. Low dose stingless bee honey increases viability of human dermal fibroblasts that could potentially promote wound healing. Wound Med. 2018, 23, 22–27. [Google Scholar] [CrossRef]
- Saiful Yazan, L.; Muhamad Zali, M.F.; Mohd Ali, R.; Zainal, N.A.; Esa, N.; Sapuan, S.; Ong, Y.S.; Tor, Y.S.; Gopalsamy, B.; Voon, F.L.; et al. Chemopreventive Properties and Toxicity of Kelulut Honey in Sprague Dawley Rats Induced with Azoxymethane. Biomed Res. Int. 2016, 2016, 4036926. [Google Scholar] [CrossRef] [Green Version]
- Yazan, L.S.; Zainal, N.A.; Ali, R.M.; Zali, M.F.S.; Yong Sze, O.; Sim, T.Y.; Gopalsamy, B.; Voon, F.L.; Sapuan, S.; Esa, N.; et al. Antiulcer properties of kelulut honey against ethanol-induced gastric ulcer. Pertanika J. Sci. Technol. 2018, 26, 121–132. [Google Scholar]
- Zaid, S.S.; Othman, S.; Kassim, N.M. Potential protective effect of Tualang honey on BPA-induced ovarian toxicity in prepubertal rat. BMC Complement. Altern. Med. 2014, 14, 509. [Google Scholar] [CrossRef] [Green Version]
- Mohamad Zaid, S.S.; Kassim, N.M.; Othman, S. Tualang Honey Protects against BPA-Induced Morphological Abnormalities and Disruption of ERα, ERβ, and C3 mRNA and Protein Expressions in the Uterus of Rats. Evid. Based. Complement. Altern. Med. 2015, 2015, 202874. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, M.; Sulaiman, S.A.; Jaafar, H.; Sirajudeen, K.N. Antioxidant protective effect of honey in cigarette smoke-induced testicular damage in rats. Int. J. Mol. Sci. 2011, 12, 5508–5521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul-Ghani, A.S.; Dabdoub, N.; Muhammad, R.; Abdul-Ghani, R.; Qazzaz, M. Effect of Palestinian honey on spermatogenesis in rats. J. Med. Food 2008, 11, 799–802. [Google Scholar] [CrossRef] [PubMed]
- Muncke, J. Endocrine disrupting chemicals and other substances of concern in food contact materials: An updated review of exposure, effect and risk assessment. J. Steroid Biochem. Mol. Biol. 2011, 127, 118–127. [Google Scholar] [CrossRef]
- Zaid, S.S.M.; Othman, S.; Kassim, N.M. Protective role of Mas Cotek (Ficus deltoidea) against the toxic effects of bisphenol A on morphology and sex steroid receptor expression in the rat uterus. Biomed. Pharmacother. 2021, 140, 111757. [Google Scholar] [CrossRef]
- Kuiper, G.G.; Enmark, E.; Pelto-Huikko, M.; Nilsson, S.; Gustafsson, J.A. Cloning of a novel receptor expressed in rat prostate and ovary. Proc. Natl. Acad. Sci. USA 1996, 93, 5925–5930. [Google Scholar] [CrossRef] [Green Version]
- Spreafico, E.; Bettini, E.; Pollio, G.; Maggi, A. Nucleotide sequence of estrogen receptor cDNA from Sprague-Dawley rat. Eur. J. Pharmacol. Mol. Pharmacol. 1992, 227, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Allard, P.; Colaiácovo, M.P. Chapter 50-Bisphenol A. In Reproductive and Developmental Toxicology; Gupta, R.C., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 673–686. [Google Scholar]
- Chen, Q.; Allgeier, A.; Yin, D.; Hollert, H. Leaching of endocrine disrupting chemicals from marine microplastics and mesoplastics under common life stress conditions. Environ. Int. 2019, 130, 104938. [Google Scholar] [CrossRef]
- Liu, Z.; Zhuan, Q.; Zhang, L.; Meng, L.; Fu, X.; Hou, Y. Polystyrene microplastics induced female reproductive toxicity in mice. J. Hazard. Mater. 2022, 424, 127629. [Google Scholar] [CrossRef]
- Wolfsegger, M.J.; Jaki, T.; Dietrich, B.; Kunzler, J.A.; Barker, K. A note on statistical analysis of organ weights in non-clinical toxicological studies. Toxicol. Appl. Pharm. 2009, 240, 117–122. [Google Scholar] [CrossRef]
- Kannan, K.; Vimalkumar, K. A Review of Human Exposure to Microplastics and Insights Into Microplastics as Obesogens. Front. Endocrinol. 2021, 12, 724989. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.; Stedman, D.B.; Elswick, B.A.; Cattley, R.C.; Welsch, F. Pubertal development and reproductive functions of Crl:CD BR Sprague-Dawley rats exposed to bisphenol A during prenatal and postnatal development. Toxicol. Sci. 2000, 55, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Ashby, J.; Tinwell, H.; Lefevre, P.A.; Joiner, R.; Haseman, J. The effect on sperm production in adult Sprague-Dawley rats exposed by gavage to bisphenol A between postnatal days 91–97. Toxicol. Sci. 2003, 74, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Salleh, N.; Ahmad, V.N. In-Vitro effect of Ficus deltoidea on the contraction of isolated rat’s uteri is mediated via multiple receptors binding and is dependent on extracellular calcium. BMC Complement. Altern. Med. 2013, 13, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigezzi, L.; Bosquiazzo, V.L.; Kass, L.; Ramos, J.G.; Muñoz-de-Toro, M.; Luque, E.H. Developmental exposure to bisphenol A alters the differentiation and functional response of the adult rat uterus to estrogen treatment. Reprod. Toxicol. 2015, 52, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Markey, C.M.; Wadia, P.R.; Rubin, B.S.; Sonnenschein, C.; Soto, A.M. Long-term effects of fetal exposure to low doses of the xenoestrogen bisphenol-A in the female mouse genital tract. Biol. Reprod. 2005, 72, 1344–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosquiazzo, V.L.; Vigezzi, L.; Muñoz-de-Toro, M.; Luque, E.H. Perinatal exposure to diethylstilbestrol alters the functional differentiation of the adult rat uterus. J. Steroid. Biochem. Mol. Biol. 2013, 138, 1–9. [Google Scholar] [CrossRef]
- Westwood, F.R. The female rat reproductive cycle: A practical histological guide to staging. Toxicol. Pathol. 2008, 36, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Khalyfa, A.; Klinge, C.M.; Hall, W.C.; Zhao, X.; Miller, M.M.; Wang, E. Transcription profiling of estrogen target genes in young and old mouse uterus. Exp. Gerontol. 2003, 38, 1087–1099. [Google Scholar] [CrossRef]
- Jewett, E.; Arnott, G.; Connolly, L.; Vasudevan, N.; Kevei, E. Microplastics and Their Impact on Reproduction-Can we Learn From the C. elegans Model? Front. Toxicol. 2022, 4, 748912. [Google Scholar] [CrossRef]
- Huang, C.; Ge, Y.; Yue, S.; Zhao, L.; Qiao, Y. Microplastics aggravate the joint toxicity to earthworm Eisenia fetida with cadmium by altering its availability. Sci. Total. Environ. 2021, 753, 142042. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Wu, X.; Liu, S.; Wang, Z.; Chen, L. Size-dependent effects of polystyrene microplastics on cytotoxicity and efflux pump inhibition in human Caco-2 cells. Chemosphere 2019, 221, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Edmonds, J.S.; Nomachi, M.; Terasaki, M.; Morita, M.; Skelton, B.W.; White, A.H. The reaction of bisphenol A 3,4-quinone with DNA. Biochem. Biophys. Res. Commun. 2004, 319, 556–561. [Google Scholar] [CrossRef]
- Atkinson, A.; Roy, D. In vivo DNA adduct formation by bisphenol A. Environ. Mol. Mutagen. 1995, 26, 60–66. [Google Scholar] [CrossRef]
- Hazirah, H.; Yasmin, A.M.; Norwahidah, A. Antioxidant properties of stingless bee honey and its effect on the viability of lymphoblastoid cell line. Med. Health 2019, 14, 91–105. [Google Scholar]
- Shamsudin, S.; Selamat, J.; Abdul Shomad, M.; Ab Aziz, M.F.; Haque Akanda, M.J. Antioxidant Properties and Characterization of Heterotrigona itama Honey from Various Botanical Origins according to Their Polyphenol Compounds. J. Food Qual. 2022, 2022, 2893401. [Google Scholar] [CrossRef]
- Adenan, M.N.H.; Yazan, L.S.; Christianus, A.; Md Hashim, N.F.; Mohd Noor, S.; Shamsudin, S.; Ahmad Bahri, F.J.; Abdul Rahim, K. Radioprotective Effects of Kelulut Honey in Zebrafish Model. Molecules 2021, 26, 1557. [Google Scholar] [CrossRef]
- Rubin, B.S.; Murray, M.K.; Damassa, D.A.; King, J.C.; Soto, A.M. Perinatal exposure to low doses of bisphenol A affects body weight, patterns of estrous cyclicity, and plasma LH levels. Environ. Health Perspect 2001, 109, 675–680. [Google Scholar] [CrossRef]
- Jin, H.; Yan, M.; Pan, C.; Liu, Z.; Sha, X.; Jiang, C.; Li, L.; Pan, M.; Li, D.; Han, X.; et al. Chronic exposure to polystyrene microplastics induced male reproductive toxicity and decreased testosterone levels via the LH-mediated LHR/cAMP/PKA/StAR pathway. Part. Fibre Toxicol. 2022, 19, 13. [Google Scholar] [CrossRef]
- Dubey, I.; Khan, S.; Kushwaha, S. Developmental and reproductive toxic effects of exposure to microplastics: A review of associated signaling pathways. Front. Toxicol. 2022, 4, 901798. [Google Scholar] [CrossRef] [PubMed]
- Ijaz, M.U.; Shahzadi, S.; Samad, A.; Ehsan, N.; Ahmed, H.; Tahir, A.; Rehman, H.; Anwar, H. Dose-Dependent Effect of Polystyrene Microplastics on the Testicular Tissues of the Male Sprague Dawley Rats. Dose. Response. 2021, 19, 15593258211019882. [Google Scholar] [CrossRef] [PubMed]
- Kamal, D.A.M.; Ibrahim, S.F.; Ugusman, A.; Mokhtar, M.H. Kelulut Honey Ameliorates Oestrus Cycle, Hormonal Profiles, and Oxidative Stress in Letrozole-Induced Polycystic Ovary Syndrome Rats. Antioxidants 2022, 11, 1879. [Google Scholar] [CrossRef] [PubMed]
- Budin, S.; Jubaidi, F.; Azam, S.; Mohammed Yusof, N.; Taib, I.S.; Mohamed, J. Kelulut honey supplementation prevents sperm and testicular oxidative damage in streptozotocin-induced diabetic rats. J. Teknol. 2017, 79, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Kamaruzzaman, M.; Thanu, A.; Yusof, M.; Soelaiman, I.; Ramli, E. Kelulut honey ameliorates glucocorticoid induced osteoporosis via its antioxidant activity in rats. Asian Pac. J. Trop. Biomed. 2019, 9, 493–500. [Google Scholar] [CrossRef]
- Schönfelder, G.; Friedrich, K.; Paul, M.; Chahoud, I. Developmental effects of prenatal exposure to bisphenol a on the uterus of rat offspring. Neoplasia 2004, 6, 584–594. [Google Scholar] [CrossRef] [Green Version]
- Seidlová-Wuttke, D.; Jarry, H.; Wuttke, W. Pure estrogenic effect of benzophenone-2 (BP2) but not of bisphenol A (BPA) and dibutylphtalate (DBP) in uterus, vagina and bone. Toxicology 2004, 205, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Kabuto, H.; Hasuike, S.; Minagawa, N.; Shishibori, T. Effects of bisphenol A on the metabolisms of active oxygen species in mouse tissues. Environ. Res. 2003, 93, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Kauffman, A.E.; Li, L.; McFee, W.; Cai, B.; Weinstein, J.; Lead, J.R.; Chatterjee, S.; Scott, G.I.; Xiao, S. Health impacts of environmental contamination of micro- and nanoplastics: A review. Environ. Health Prev. Med. 2020, 25, 29. [Google Scholar] [CrossRef]
- Llorca, M.; Farré, M. Current Insights into Potential Effects of Micro-Nanoplastics on Human Health by in-vitro Tests. Front. Toxicol. 2021, 3, 752140. [Google Scholar] [CrossRef]
- Kuiper, G.G.; Carlsson, B.; Grandien, K.; Enmark, E.; Häggblad, J.; Nilsson, S.; Gustafsson, J.A. Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta. Endocrinology 1997, 138, 863–870. [Google Scholar] [CrossRef]
- Paech, K.; Webb, P.; Kuiper, G.G.; Nilsson, S.; Gustafsson, J.; Kushner, P.J.; Scanlan, T.S. Differential ligand activation of estrogen receptors ERalpha and ERbeta at AP1 sites. Science 1997, 277, 1508–1510. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.M.; McDonnell, D.P. The estrogen receptor beta-isoform (ERbeta) of the human estrogen receptor modulates ERalpha transcriptional activity and is a key regulator of the cellular response to estrogens and antiestrogens. Endocrinology 1999, 140, 5566–5578. [Google Scholar] [CrossRef]
- Pettersson, K.; Grandien, K.; Kuiper, G.G.; Gustafsson, J.A. Mouse estrogen receptor beta forms estrogen response element-binding heterodimers with estrogen receptor alpha. Mol. Endocrinol. 1997, 11, 1486–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, J.R.; Chakraborty, S.; Chakraborty, T.R. Estrogen-like endocrine disrupting chemicals affecting puberty in humans—A review. Med. Sci. Monit. 2009, 15, Ra137-45. [Google Scholar] [PubMed]
- Wattel, A.; Kamel, S.; Mentaverri, R.; Lorget, F.; Prouillet, C.; Petit, J.P.; Fardelonne, P.; Brazier, M. Potent inhibitory effect of naturally occurring flavonoids quercetin and kaempferol on in vitro osteoclastic bone resorption. Biochem. Pharm. 2003, 65, 35–42. [Google Scholar] [CrossRef]
- Al-Hatamleh, M.A.I.; Boer, J.C.; Wilson, K.L.; Plebanski, M.; Mohamud, R.; Mustafa, M.Z. Antioxidant-Based Medicinal Properties of Stingless Bee Products: Recent Progress and Future Directions. Biomolecules 2020, 10, 923. [Google Scholar] [CrossRef] [PubMed]
- Zaid, S.S.; Sulaiman, S.A.; Othman, N.H.; Soelaiman, I.N.; Shuid, A.N.; Mohamad, N.; Muhamad, N. Protective effects of Tualang honey on bone structure in experimental postmenopausal rats. Clinics 2012, 67, 779–784. [Google Scholar] [CrossRef]
- Działo, M.; Mierziak, J.; Korzun, U.; Preisner, M.; Szopa, J.; Kulma, A. The Potential of Plant Phenolics in Prevention and Therapy of Skin Disorders. Int. J. Mol. Sci. 2016, 17, 160. [Google Scholar] [CrossRef] [Green Version]
- Dharmaraja, A.T. Role of Reactive Oxygen Species (ROS) in Therapeutics and Drug Resistance in Cancer and Bacteria. J. Med. Chem. 2017, 60, 3221–3240. [Google Scholar] [CrossRef]
- Güney, M.; Oral, B.; Demirin, H.; Ozgüner, M.; Take, G.; Mungan, T.; Altuntas, I. Evaluation of caspase-dependent apoptosis during methyl parathion-induced endometrial damage in rats: Ameliorating effect of Vitamins E and C. Environ. Toxicol Pharm. 2007, 23, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Aslan, R.; Sekeroğlu, M.R.; Tarakçioğlu, M.; Köylü, H. Investigation of malondialdehyde formation and antioxidant enzyme activity in stored blood. Haematologia 1997, 28, 233–237. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Reverse Primer/Forward Primer | References (Accession Number) |
|---|---|---|
| Β-actin | 5′-GCCGGGACCTGACTGACTAC-3′ 5′-TTCTCCTTAATGTCACGCACGAT-3′ | Kuipter et al., 1996 [28] (U57439) |
| ERα | 5′-GCAGGTCATAGAGAGGCACGA-3′ 5′-AAGCTGGCCTGACTCTGCAG-3′ | Spreafico et al., 1992 [29] (X61098) |
| ERβ | 5′-GGAGATACCACTCTTCGCAATC-3′ 5′-CTCTGTGTGAAGGCCATGAT-3′ | Kuipter et al., 1996 [28] (U57439) |
| Group | Body Weight Gain (g) | Changes in Body Weight Gain (%) | Uterine Wet Weight (mg) | The Relative Weight of the Uterus (Wet Weight/Body Weight) |
|---|---|---|---|---|
| NC | 144.54.13 | 60.3511.37 | 33312.02 | 1.840.11 |
| M | 151.53.29 a,b | 62.7017.74 a,b | 2602.00 a,b | 1.460.07 a,b |
| DM | 141.75.59 | 59.1818.20 | 3033.09 | 1.590.04 |
| DC | 141.58.33 | 59.1715.56 | 31414.59 | 1.720.10 |
| Group | Height of Luminal Epithelial Cell (µm) | The Thickness of the Endometrium (µm) | The Thickness of Myometrium (µm) | Height of Epithelial Endometrial Glands (µm) | Diameter of Endometrial Glands (µm) |
|---|---|---|---|---|---|
| NC | 29.12 2.38 | 643.55 15.83 | 343.4 17.10 | 19.36 1.29 | 71.11 4.58 |
| M | 23.71 1.013 a,b | 562.65 12.51 a,b | 290.96 15.93 a,b | 15.66 0.22 a,b | 68.43 9.28 a,b |
| DM | 35.21 2.351 | 581.84 24.69 | 378.89 17.43 | 17.25 1.60 | 65.79 2.35 |
| DC | 31.90 1.462 | 572.48 23.58 | 322.83 16.06 | 20.72 1.40 | 76.26 2.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amran, N.H.; Zaid, S.S.M.; Meng, G.Y.; Salleh, A.; Mokhtar, M.H. Protective Role of Kelulut Honey against Toxicity Effects of Polystyrene Microplastics on Morphology, Hormones, and Sex Steroid Receptor Expression in the Uterus of Rats. Toxics 2023, 11, 324. https://doi.org/10.3390/toxics11040324
Amran NH, Zaid SSM, Meng GY, Salleh A, Mokhtar MH. Protective Role of Kelulut Honey against Toxicity Effects of Polystyrene Microplastics on Morphology, Hormones, and Sex Steroid Receptor Expression in the Uterus of Rats. Toxics. 2023; 11(4):324. https://doi.org/10.3390/toxics11040324
Chicago/Turabian StyleAmran, Nur Hanisah, Siti Sarah Mohamad Zaid, Goh Yong Meng, Annas Salleh, and Mohd Helmy Mokhtar. 2023. "Protective Role of Kelulut Honey against Toxicity Effects of Polystyrene Microplastics on Morphology, Hormones, and Sex Steroid Receptor Expression in the Uterus of Rats" Toxics 11, no. 4: 324. https://doi.org/10.3390/toxics11040324