
Risk Assessment of Fluxametamide Resistance and Fitness Costs in Fall Armyworm (Spodoptera frugiperda)
 , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Populations and Rearing
2.2. Insecticides
2.3. Bioassays
2.4. Selection Using Fluxametamide
2.5. Estimation of Realized Heritability (h2)
2.6. Detoxification Enzyme Activities
2.7. Evaluation of Fitness Costs
2.8. Data Analysis
3. Results
3.1. Assortment for Fluxametamide Resistance
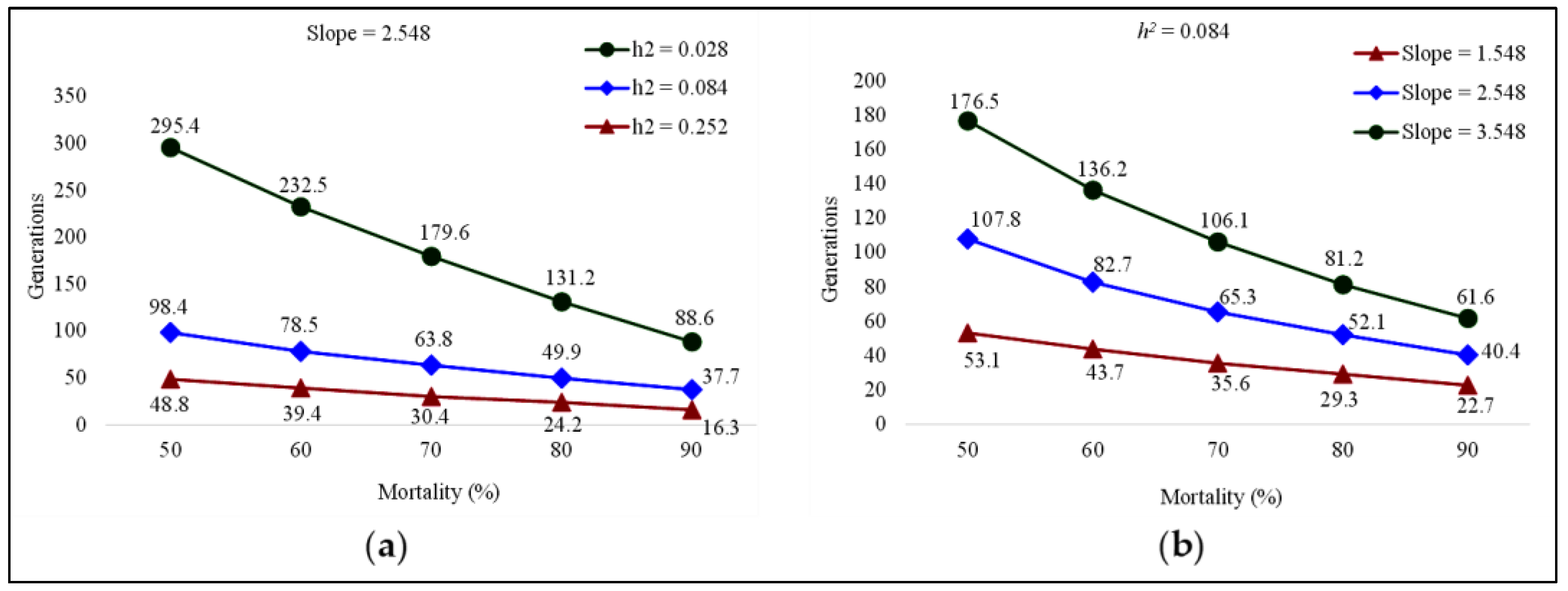
3.2. Estimation of Fluxametamide Resistance Development
3.3. Cross-Resistance Pattern
3.4. Activities of Detoxification Enzymes
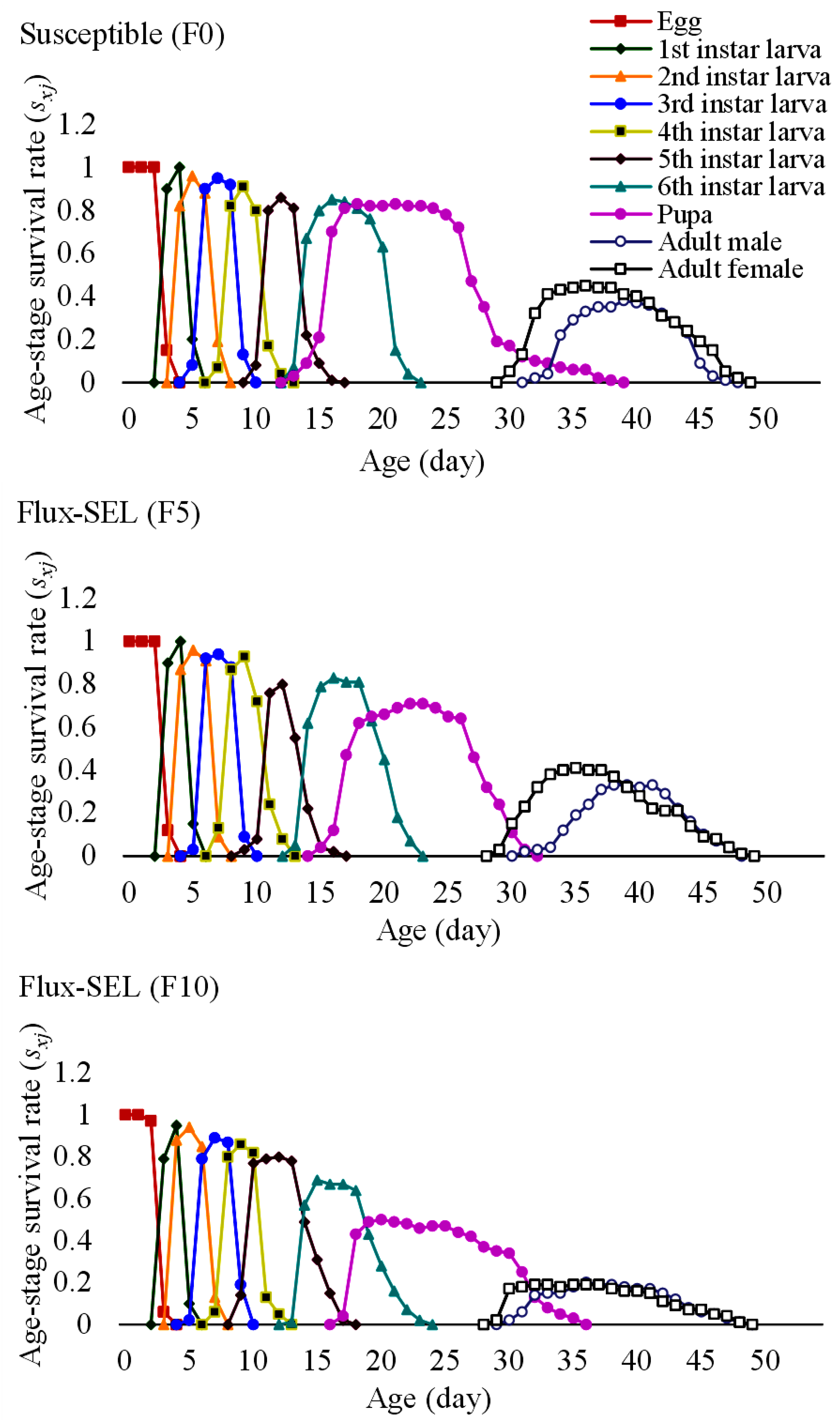
3.5. Effects of Fluxametamide Selection on the Growth and Reproductive Parameters of FAW
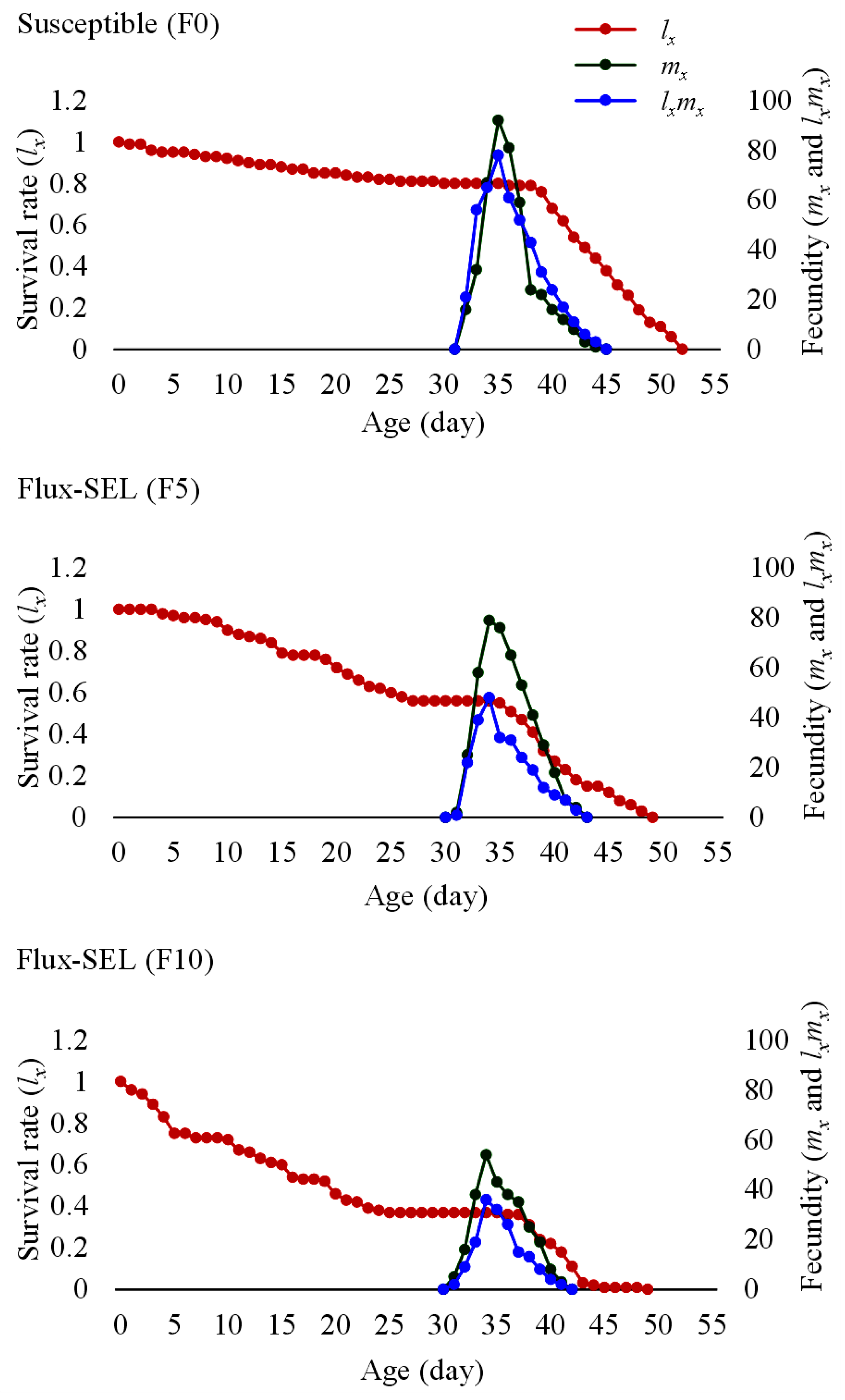
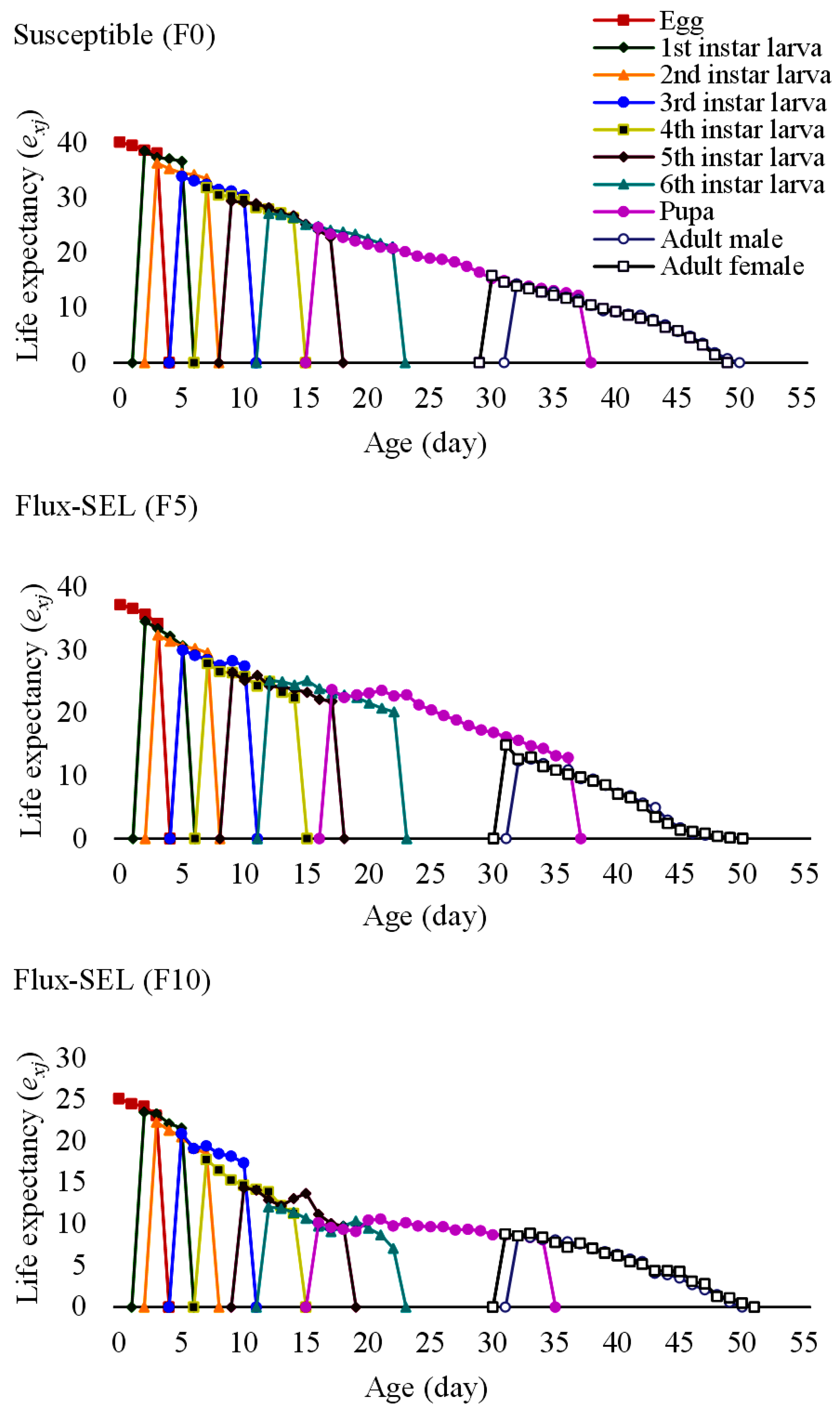
3.6. Effects of Fluxametamide Selection on the Fitness Costs and Life Table Parameters of FAW
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (J. E. Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in west and central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [Green Version]
- Ganiger, P.C.; Yeshwanth, H.M.; Muralimohan, K.; Vinay, N.; Kumar, A.R.V.; Chandrashekara, K. Occurrence of the new invasive pest, fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae), in the maize fields of Karnataka, India. Curr. Sci. 2018, 115, 621. [Google Scholar] [CrossRef]
- Early, R.; González-Moreno, P.; Murphy, S.T.; Day, R. Forecasting the global extent of invasion of the cereal pest Spodoptera frugiperda, the fall armyworm. NeoBiota 2018, 40, 25–50. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Qi, G.; Chen, H.; Ma, J.; Liu, J.; Jiang, Y.; Lee, G.; Otuka, A.; Hu, G. Overseas immigration of fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), invading Korea and Japan in 2019. Insect Sci. 2021, 29, 505–520. [Google Scholar] [CrossRef]
- Kuate, A.F.; Hanna, R.; Fotio, A.R.P.D.; Abang, A.F.; Nanga, S.N.; Ngatat, S.; Tindo, M.; Masso, C.; Ndemah, R.; Suh, C.; et al. Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) in Cameroon: Case study on its distribution, damage, pesticide use, genetic differentiation and host plants. PLoS ONE 2019, 14, e0215749. [Google Scholar] [CrossRef] [Green Version]
- Sharanabasappa, D.; Kalleshwaraswamy, C.M.; Asokan, R.; Swamy, H.M.; Maruthi, M.S.; Pavithra, H.B.; Hegde, K.; Navi, S.; Prabhu, S.T.; Goergen, G. First report of the fall armyworm, Spodoptera frugiperda (J E Smith) (Lepidoptera: Noctuidae), an alien invasive pest on maize in India. Pest Manag. Hort. Ecosyst. 2018, 24, 23–29. [Google Scholar]
- Sun, X.X.; Hu, C.X.; Jia, H.R.; Wu, Q.L.; Shen, X.J.; Zhao, S.Y.; Jiang, Y.Y.; Wu, K.M. Case study on the first immigration of fall armyworm, Spodoptera frugiperda invading into China. J. Integr. Agric. 2021, 20, 664–672. [Google Scholar] [CrossRef]
- Zhang, D.D.; Xiao, Y.T.; Xu, P.J.; Yang, X.M.; Wu, Q.L.; Wu, K.M. Insecticide resistance monitoring for the invasive populations of fall armyworm, Spodoptera frugiperda in China. J. Integr. Agric. 2021, 20, 783–791. [Google Scholar] [CrossRef]
- Kulye, M.; Mehlhorn, S.; Boaventura, D.; Godley, N.; Venkatesh, S.K.; Rudrappa, T.; Charan, T.; Rathi, D.; Nauen, R. Baseline susceptibility of Spodoptera frugiperda populations collected in India towards different chemical classes of insecticides. Insects 2021, 12, 758. [Google Scholar] [CrossRef]
- Wu, H.M.; Feng, H.L.; Wang, G.D.; Zhang, L.L.; Zulu, L.; Liu, Y.H.; Zheng, Y.L.; Rao, Q. Sublethal effects of three insecticides on development and reproduction of Spodoptera frugiperda (Lepidoptera: Noctuidae). Agronomy 2022, 12, 1334. [Google Scholar] [CrossRef]
- Ahmad, L.; Habib Kanth, R.; Parvaze, S.; Sheraz Mahdi, S. Agro-climatic and agro-ecological zones of India. In Experimental Agrometeorology: A Practical Manual; Ahmad, L., Habib Kanth, R., Parvaze, S., Sheraz Mahdi, S., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 99–118. [Google Scholar] [CrossRef]
- Firake, D.M.; Behere, G.T. Natural mortality of invasive fall armyworm, Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) in maize agroecosystems of northeast India. Biol. Control 2020, 148, 104303. [Google Scholar] [CrossRef]
- Varshney, R.; Poornesha, B.; Raghavendra, A.; Lalitha, Y.; Apoorva, V.; Ramanujam, B.; Rangeshwaran, R.; Subaharan, K.; Shylesha, A.N.; Bakthavatsalam, N.; et al. Biocontrol-based management of fall armyworm, Spodoptera frugiperda (J E Smith) (Lepidoptera: Noctuidae) on Indian Maize. J. Pl. Dis. Prot. 2021, 128, 87–95. [Google Scholar] [CrossRef]
- Shylesha, A.N.; Jalali, S.K.; Gupta, A.; Varshney, R.; Venkatesan, T.; Shetty, P.; Ojha, R.; Ganiger, P.C.; Navik, O.; Subaharan, K.; et al. Studies on new invasive pest Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) and its natural enemies. J. Biol. Control 2018, 32, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Roy, D.; Biswas, S.; Mondal, D.; Majumder, S.; Sarkar, P.K. Efficacy and safety-evaluation of insecticidal modules against Spodoptera frugiperda (Lepidoptera: Noctuidae) and the residues of the most effective schedule in maize. Int. J. Trop. Insect Sci. 2021, 41, 3155–3166. [Google Scholar] [CrossRef]
- Carvalho, R.A.; Omoto, C.; Field, L.M.; Williamson, M.S.; Bass, C. Investigating the molecular mechanisms of organophosphate and pyrethroid resistance in the fall armyworm Spodoptera frugiperda. PLoS ONE 2013, 8, e62268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolzan, A.; Padovez, F.E.; Nascimento, A.R.; Kaiser, I.S.; Lira, E.C.; Amaral, F.S.; Kanno, R.H.; Malaquias, J.B.; Omoto, C. Selection and characterization of the inheritance of resistance of Spodoptera frugiperda (Lepidoptera: Noctuidae) to chlorantraniliprole and cross-resistance to other diamide insecticides. Pest Manag. Sci. 2019, 75, 2682–2689. [Google Scholar] [CrossRef]
- do Nascimento, A.R.B.; Farias, J.R.; Bernardi, D.; Horikoshi, R.J.; Omoto, C. Genetic basis of Spodoptera frugiperda (Lepidoptera: Noctuidae) resistance to the chitin synthesis inhibitor lufenuron. Pest Manag. Sci. 2016, 72, 810–815. [Google Scholar] [CrossRef]
- Okuma, D.M.; Bernardi, D.; Horikoshi, R.J.; Bernardi, O.; Silva, A.P.; Omoto, C. Inheritance and fitness costs of Spodoptera frugiperda (Lepidoptera: Noctuidae) resistance to spinosad in Brazil. Pest Manag. Sci. 2018, 74, 1441–1448. [Google Scholar] [CrossRef]
- Diez-Rodríguez, G.I.; Omoto, C. Herança da resistência de Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) a lambda-cialotrina. Neotrop. Entomol. 2001, 30, 311–316. [Google Scholar] [CrossRef] [Green Version]
- Farias, J.R.; Andow, D.A.; Horikoshi, R.J.; Sorgatto, R.J.; Fresia, P.; dos Santos, A.C.; Omoto, C. Field-evolved resistance to Cry1F maize by Spodoptera frugiperda (Lepidoptera: Noctuidae) in Brazil. Crop Prot. 2014, 64, 150–158. [Google Scholar] [CrossRef]
- Horikoshi, R.J.; Bernardi, D.; Bernardi, O.; Malaquias, J.B.; Okuma, D.M.; Miraldo, L.L.; Amaral, F.S.; Omoto, C. Effective dominance of resistance of Spodoptera frugiperda to Bt maize and cotton varieties: Implications for resistance management. Sci. Rep. 2016, 6, 34864. [Google Scholar] [CrossRef] [Green Version]
- Santos-Amaya, O.F.; Rodrigues, J.V.C.; Souza, T.C.; Tavares, C.S.; Campos, S.O.; Guedes, R.N.C.; Pereira, E.J.G. Resistance to dual-gene Bt maize in Spodoptera frugiperda: Selection, inheritance and cross-resistance to other transgenic events. Sci. Rep. 2015, 5, 18243. [Google Scholar] [CrossRef] [Green Version]
- Boaventura, D.; Ulrich, J.; Lueke, B.; Bolzan, A.; Okuma, D.; Gutbrod, O.; Geibel, S.; Zeng, Q.; Dourado, P.M.; Martinelli, S.; et al. Molecular characterization of Cry1F resistance in fall armyworm, Spodoptera frugiperda from Brazil. Insect Biochem. Mol. Biol. 2020, 116, 103280. [Google Scholar] [CrossRef] [PubMed]
- Sparks, T.C.; Storer, N.; Porter, A.; Slater, R.; Nauen, R. Insecticide resistance management and industry: The origins and evolution of the Insecticide Resistance Action Committee (IRAC) and the mode of action classification scheme. Pest Manag. Sci. 2021, 77, 2609–2619. [Google Scholar] [CrossRef] [PubMed]
- Asahi, M.; Kobayashi, M.; Kagami, T.; Nakahira, K.; Furukawa, Y.; Ozoe, Y. Fluxametamide: A novel isoxazoline insecticide that acts via distinctive antagonism of insect ligand-gated chloride channels. Pestic. Biochem. Physiol. 2018, 151, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Umetsu, N.; Shirai, Y. Development of the novel pesticides in the 21st century. J. Pestic. Sci. 2020, 45, 54–74. [Google Scholar] [CrossRef] [Green Version]
- Kagami, T.; Hori, M.; Haruyama, H. Studies on a novel insecticide, fluxametamide (Part 2) biological activity. Abstr. Annu. Meeting Pestic. Sci. Soc. Jpn. 2017, 35, 83. [Google Scholar]
- Mita, T.; Furukawa, Y.; Iwasa, M.; Kikuchi, T.; Komoda, M. Studies on a novel insecticide, fluxametamide. In Recent Highlights in the Discovery and Optimization of Crop Protection Products; Maienfisch, P., Mangelinckx, S., Eds.; Academic Press Publishers: Cambridge, MA, USA, 2021; pp. 157–163. [Google Scholar] [CrossRef]
- Jeschke, P. Status and outlook for acaricide and insecticide discovery. Pest Manag. Sci. 2021, 77, 64–76. [Google Scholar] [CrossRef]
- Gope, A.; Chakraborty, G.; Ghosh, S.M.; Sau, S.; Mondal, K.; Biswas, A.; Sarkar, S.; Sarkar, P.K.; Roy, D. Toxicity and sublethal effects of fluxametamide on the key biological parameters and life history traits of diamondback moth Plutella xylostella (L.). Agronomy 2022, 12, 1656. [Google Scholar] [CrossRef]
- Roy, D.; Sau, S.; Adhikary, S.; Biswas, A.; Biswas, S.; Chakraborty, G.; Sarkar, P.K. Resistance risk assessment in diamondback moth, Plutella xylostella (L.) to fluxametamide. Crop Protec. 2022, 163, 106101. [Google Scholar] [CrossRef]
- Sayyed, A.H.; Omar, D.; Wright, D.J. Genetics of spinosad resistance in a multi-resistant field-selected population of Plutella xylostella. Pest Manag. Sci. 2004, 60, 827–832. [Google Scholar] [CrossRef]
- Lai, T.; Su, J. Assessment of resistance risk in Spodoptera exigua (Hubner) (Lepidoptera: Noctuidae) to chlorantraniliprole. Pest Manag. Sci. 2011, 67, 1468–1472. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Qian, C.; Tang, T.; Shen, N.; Wu, W.; Wang, J.; Han, Z.; Zhao, C. Lethal and sublethal effects of fluxametamide on rice-boring pest, rice stem borer Chilo suppressalis. Agronomy 2022, 12, 2429. [Google Scholar] [CrossRef]
- Kumar, N.T.D.; Mohan, K.M. Variations in the susceptibility of Indian populations of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) to selected insecticides. Int. J. Trop. Insect Sci. 2022, 42, 1707–1712. [Google Scholar] [CrossRef]
- Pinto, J.R.L.; Torres, A.F.; Truzi, C.C.; Vieira, N.F.; Vacari, A.M.; De Bortoli, S.A. Artificial corn-based diet for rearing Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Insect Sci. 2019, 19, 2. [Google Scholar] [CrossRef] [PubMed]
- Gergs, A.; Baden, C.U. A dynamic energy budget approach for the prediction of development times and variability in Spodoptera frugiperda rearing. Insects 2021, 12, 300. [Google Scholar] [CrossRef] [PubMed]
- Test Methods|Insecticide Resistance Action Committee (IRAC). Available online: https://irac-online.org/methods/ (accessed on 19 January 2022).
- Tabashnik, B.E. Resistance risk assessment: Realized heritability of resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera: Plutellidae), tobacco budworm (Lepidoptera: Noctuidae), and Colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1992, 85, 1551–1559. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Mcgaughey, W.H. Resistance risk assessment for single and multiple insecticides: Responses of Indian meal moth (Lepidoptera: Pyralidae) to Bacillus thuringiensis. J. Econ. Entomol. 1994, 87, 834–841. [Google Scholar] [CrossRef]
- Chen, C.; Han, P.; Yan, W.; Wang, S.; Shi, X.; Zhou, X.; Desneux, N.; Gao, X. Uptake of quercetin reduces larval sensitivity to lambda-cyhalothrin in Helicoverpa armigera. J. Pest Sci. 2018, 91, 919–926. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Nauen, R.; Stumpf, N. Fluorometric microplate assay to measure glutathione S-transferase activity in insects and mites using monochlorobimane. Anal. Biochem. 2002, 303, 194–198. [Google Scholar] [CrossRef]
- Grant, D.F.; Bender, D.M.; Hammock, B.D. Quantitative kinetic assays for glutathione S-transferase and general esterase in individual mosquitoes using an EIA reader. Insect Biochem. 1989, 19, 741–751. [Google Scholar] [CrossRef]
- Abbott, S.W. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Robertson, J.L.; Russell, R.M.; Preisler, H.K.; Savin, N.E. Pesticide Bioassays with Arthropods, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar] [CrossRef]
- Chi, H.; Liu, H.I.S. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age Stage, Two-Sex Life Table Analysis. 2018. Available online: http://140.120.197.173/Ecology/Download/Twosex-MSChart-exe-B200000.rar (accessed on 22 December 2022).
- Moreno, R.G.; Sanchez, D.M.; Blanco, C.A.; Whalon, M.E.; Santofimio, H.T.; Maciel, J.C.R.; DiFonzo, C. Field-evolved resistance of the fall armyworm (Lepidoptera: Noctuidae) to synthetic insecticides in Puerto Rico and Mexico. J. Econ. Entomol. 2019, 112, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Sparks, T.C.; Crossthwaite, A.J.; Nauen, R.; Banba, S.; Cordova, D.; Earley, F.; Ebbinghaus-Kintscher, U.; Fujioka, S.; Hirao, A.; Karmon, D.; et al. Insecticides, biologics and nematicides: Updates to IRAC’s mode of action classification—A tool for resistance management. Pestic. Biochem. Physiol. 2020, 167, 104587. [Google Scholar] [CrossRef] [PubMed]
- Yainna, S.; Negre, N.; Silvie, P.J.; Brevault, T.; Tay, W.T.; Gordon, K.; dAlencon, E.; Walsh, T.; Nam, K. Geographic monitoring of insecticide resistance mutations in native and invasive populations of the fall armyworm. Insects 2021, 12, 468. [Google Scholar] [CrossRef]
- Jia, Z.Q.; Zhan, E.L.; Zhang, S.G.; Jones, A.K.; Zhu, L.; Wang, Y.N.; Huang, Q.T.; Han, Z.J.; Zhao, C.Q. Sublethal doses of broflanilide prevents molting in the fall armyworm, Spodoptera frugiperda via altering molting hormone biosynthesis. Pestic. Biochem. Physiol. 2022, 181, 105017. [Google Scholar] [CrossRef]
- Ijaz, M.; Afzal, M.B.S.; Shad, S.A. Resistance risk analysis to acetamiprid and other insecticides in acetamiprid-selected population of Phenacoccus solenopsis. Phytoparasitica 2016, 44, 177–186. [Google Scholar] [CrossRef]
- Ismail, M.; Ejaz, M.; Abbas, N.; Shad, S.A.; Afzal, M.B.S. Resistance risk assessment to chlorpyrifos and cross-resistance to other insecticides in a field strain of Phenacoccus solenopsis Tinsley. Crop Protect. 2017, 94, 38–43. [Google Scholar] [CrossRef]
- Abbas, N.; Shad, S.A. Assessment of resistance risk to lambda-cyhalothrin and cross-resistance to four other insecticides in the house fly, Musca domestica L. (Diptera: Muscidae). Parasitol. Res. 2015, 114, 2629–2637. [Google Scholar] [CrossRef] [PubMed]
- Mousseau, T.A.; Roff, D.A. Natural selection and heritability of fitness components. Heredity 1987, 59, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Diez, J.D.R.; Benjumea, C.I.S. Susceptibility of Spodoptera frugiperda (Lepidoptera: Noctuidae) strains from central Colombia to two insecticides, methomyl and lambda cyhalothrin: A study of the genetic basis of resistance. J. Econ. Entomol. 2011, 104, 1698–1705. [Google Scholar] [CrossRef]
- Gorman, K.; Slater, R.; Blande, J.D.; Clarke, A.; Wren, J.; McCaffery, A.; Denholm, I. Cross-resistance relationships between neonicotinoids and pymetrozine in Bemisia tabaci (Hemiptera: Aleyrodidae). Pest Manag. Sci. 2010, 66, 1186–1190. [Google Scholar] [CrossRef] [PubMed]
- Elbert, A.; Nauen, R. Resistance of Bemisia tabaci (Homoptera: Aleyrodidae) to insecticides in southern Spain with special reference to neonicotinoids. Pest Manag. Sci. 2000, 56, 60–64. [Google Scholar] [CrossRef]
- Mahalanobish, D.; Dutta, S.; Roy, D.; Biswas, A.; Sarkar, S.; Mondal, D.; Gaber, A.; Hossain, A.; Sarkar, P.K. Field-evolved resistance and mechanisms in Bemisia tabaci Asia I to a novel pyropene insecticide, afidopyropen, in India. Crop Protect. 2022, 162, 106078. [Google Scholar] [CrossRef]
- Bernardi, D.; Salmeron, E.; Horikoshi, R.J.; Bernardi, O.; Dourado, P.M.; Carvalho, R.A.; Martinelli, S.; Head, G.P.; Omoto, C. Cross-resistance between Cry1 proteins in fall armyworm (Spodoptera frugiperda) may affect the durability of current pyramided Bt maize hybrids in Brazil. PLoS ONE 2015, 10, e0140130. [Google Scholar] [CrossRef] [Green Version]
- Lira, E.C.; Bolzan, A.; Nascimento, A.R.B.; Amaral, F.S.A.; Kanno, R.H.; Kaiser, I.S.; Omoto, C. Resistance of Spodoptera frugiperda (Lepidoptera: Noctuidae) to spinetoram: Inheritance and cross-resistance to spinosad. Pest Manag. Sci. 2020, 76, 2674–2680. [Google Scholar] [CrossRef] [PubMed]
- Hafeez, M.; Li, X.; Ullah, F.; Zhang, Z.; Zhang, J.; Huang, J.; Chen, L.; Siddiqui, J.A.; Ren, X.; Zhou, S.; et al. Characterization of indoxacarb resistance in the fall armyworm: Selection, inheritance, cross-resistance, possible biochemical mechanisms, and fitness costs. Biology 2022, 11, 1718. [Google Scholar] [CrossRef]
- Sun, X.I.; Wei, R.; Li, L.; Zhu, B.; Liang, P.; Gao, X. Resistance and fitness costs in diamondback moths after selection using broflanilide, a novel meta-diamide insecticide. Insect Sci. 2021, 29, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Roy, D.; Samanta, A.; Biswas, A.; Chakraborty, G.; Sarkar, P.K. Insecticide resistance status of Hyposidra talaca (Lepidoptera: Geometridae) in major tea growing zone of India. Phytoparasitica 2021, 49, 983–1002. [Google Scholar] [CrossRef]
- Giraudo, M.; Hilliou, F.; Fricaux, T.; Audant, P.; Feyereisen, R.; LeGoff, G. Cytochrome P450s from the fall armyworm (Spodoptera frugiperda): Responses to plant allelochemicals and pesticides. Insect Mol. Biol. 2015, 24, 115–128. [Google Scholar] [CrossRef]
- Pu, X.; Yang, Y.; Wu, S.; Wu, Y. Characterization of abamectin resistance in a field-evolved multiresistant population of Plutella xylostella. Pest Manag. Sci. 2010, 66, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.J.; Wang, Z.H.; Shi, B.C.; Kang, Z.J.; Zhu, L.; Jin, G.H.; Wei, S.J. Correlation between pesticide resistance and enzyme activity in the diamondback moth, Plutella xylostella. J. Insect Sci. 2013, 13, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Punzo, F. Detoxification enzymes and the effect of temperature on the toxicity of pyrethroids to the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Comp. Biochem. Physiol. 1993, 105, 155–158. [Google Scholar] [CrossRef]
- Saingamsook, J.; Yanola, J.; Lumjuan, N.; Walton, C.; Somboon, P. Investigation of relative development and reproductivity fitness cost in three insecticide-resistant strains of Aedes aegypti from Thailand. Insects 2019, 10, 265. [Google Scholar] [CrossRef] [Green Version]
- Kliot, A.; Ghanim, M. Fitness costs associated with insecticide resistance. Pest Manag. Sci. 2012, 68, 1431–1437. [Google Scholar] [CrossRef]
- Barbosa, M.G.; Andre, T.P.P.; Pontes, A.D.S.; Souza, S.A.; Oliveira, N.R.X.; Pastori, P.L. Insecticide rotation and adaptive fitness cost underlying insecticide resistance management for Spodoptera frugiperda (Lepidoptera: Noctuidae). Neotrop. Entomol. 2020, 49, 882–892. [Google Scholar] [CrossRef]
- Santos-Amaya, O.F.; Tavares, C.S.; Rodrigues, J.V.C.; Oliveira, E.E.; Guedes, R.N.C.; Pereira, E.J.G. Strong fitness costs of fall armyworm resistance to dual-gene Bt maize are magnified on less-suitable host-crop cultivars. Agronomy 2022, 12, 682. [Google Scholar] [CrossRef]
- Hannig, G.T.; Ziegler, M.; Marcon, P.G. Feeding cessation effects of chlorantraniliprole, a new anthranilic diamide insecticide, in comparison with several insecticides in distinct chemical classes and mode-of-action groups. Pest Manag. Sci. 2009, 65, 969–974. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Jia, Z.Q.; Peng, Y.C.; Sheng, C.W.; Tang, T.; Xu, L.; Han, Z.J.; Zhao, C.Q. Toxicity and sublethal effects of fluralaner on Spodoptera litura Fabricius (Lepidoptera: Noctuidae). Pestic. Biochem. Physiol. 2018, 152, 8–16. [Google Scholar] [CrossRef]
- Weseloh, R. Effects of the feeding inhibitor plictran and low Bacillus thuringiensis Berliner doses on Lymantria dispar (L.) (Lepidoptera: Lymantriidae): Implications for Cotesia melanoscelus (Ratzeburg) (Hymenoptera: Braconidae). Environ. Entomol. 1984, 13, 1371–1376. [Google Scholar] [CrossRef]
- Erb, S.L.; Bourchier, R.S.; van Frankenhuyzen, K.; Smith, S.M. Sublethal effects of Bacillus thuringiensis Berliner subsp. kurstaki on Lymantria dispar (Lepidoptera: Lymantriidae) and the tachinid parasitoid Compsilura concinnata (Diptera: Tachinidae). Environ. Entomol. 2001, 30, 1174–1181. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Sun, H.; Yasoob, H.; Tian, Z.; Li, Y.; Li, R.; Zheng, S.; Liu, J.; Zhang, Y. Biogenetic cantharidin is a promising leading compound to manage insecticide resistance of Mythimna separata (Lepidoptera: Noctuidae). Pestic. Biochem. Physiol. 2021, 172, 104769. [Google Scholar] [CrossRef]
- Shahout, H.A.; Xu, J.X.; Yao, X.M.; Jia, Q.D. Influence and mechanism of different host plants on the growth, development, and fecundity of reproductive system of common cut worm Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae). Asian J. Agric. Sci. 2011, 3, 291–300. [Google Scholar]
- Stark, J.D.; Banks, J.E. Population-level effects of pesticides and other toxicants on arthropods. Annu. Rev. Entomol. 2003, 48, 505–519. [Google Scholar] [CrossRef]
- Hamedi, N.; Fathipour, Y.; Saber, M. Sublethal effects of fenpyroximate on life table parameters of the predatory mite Phytoseius plumifer. BioControl 2010, 55, 271–278. [Google Scholar] [CrossRef]
- Yu, N.; Tian, J.; Zhang, Y.; Li, Z.; Liu, Z. Imidacloprid-susceptible Nilaparvata lugens individuals exceeded resistant individuals in a mixture population with density pressure. Pest Manag. Sci. 2018, 74, 234–239. [Google Scholar] [CrossRef]
- Wang, R.; Qu, C.; Wang, Z.; Yang, G.F. Cross-resistance, biochemical mechanism and fitness costs of laboratory-selected resistance to pyridalyl in diamondback moth, Plutella xylostella. Pestic. Biochem. Physiol. 2020, 163, 8–13. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Generation | Concentration for Selection (mg L−1) | N | LC50 (95% CI) (mg L−1) | Slope ± SE | ꭓ2 (df) | RF (95% CI) |
|---|---|---|---|---|---|---|
| Susceptible (F0) | - | 280 | 0.024 (0.015–0.032) | 1.56 ± 0.19 | 1.23 (5) | - |
| Flux-SEL (F1) | 0.035 | 280 | 0.029 (0.021–0.036) | 2.74 ± 0.32 | 1.85 (5) | 1.21 (1.12–1.28) |
| Flux-SEL (F2) | 0.043 | 280 | 0.025 (0.018–0.034) | 2.49 ± 0.26 | 2.32 (5) | 1.04 (0.93–1.13) |
| Flux-SEL (F3) | 0.032 | 280 | 0.027 (0.021–0.034) | 3.28 ± 0.18 | 3.51 (5) | 1.13 (0.98–1.22) |
| Flux-SEL (F4) | 0.037 | 280 | 0.032 (0.025–0.042) | 2.67 ± 0.22 | 2.94 (5) | 1.33 (1.19–1.44) |
| Flux-SEL (F5) | 0.046 | 280 | 0.036 (0.024–0.049) | 2.82 ± 0.29 | 1.77 (5) | 1.50 (1.42–1.59) |
| Flux-SEL (F6) | 0.051 | 280 | 0.045 (0.033–0.057) | 3.08 ± 0.34 | 2.26 (5) | 1.88 (1.76–1.99) |
| Flux-SEL (F7) | 0.063 | 280 | 0.042 (0.029–0.053) | 1.93 ± 0.21 | 2.47 (5) | 1.75 (1.63–1.90) |
| Flux-SEL (F8) | 0.058 | 280 | 0.049 (0.038–0.064) | 2.51 ± 0.29 | 1.39 (5) | 2.04 (1.89–2.17) |
| Flux-SEL (F9) | 0.074 | 280 | 0.055 (0.043–0.069) | 2.19 ± 0.25 | 3.15 (5) | 2.29 (2.20–2.36) |
| Flux-SEL (F10) | 0.096 | 280 | 0.063 (0.051–0.074) | 2.75 ± 0.31 | 2.45 (5) | 2.63 (2.52–2.74) |
| Selected Generations | Response to Selection | Selection Differential | h2 | |||||
|---|---|---|---|---|---|---|---|---|
| Initial LC50 (95% CI) | Final LC50 (95% CI) | R | Average Slope | σp | i | S | ||
| 10 (F0–F10) | 0.024 (0.013–0.032) | 0.063 (0.051–0.074) | 0.042 | 2.548 | 0.393 | 1.273 | 0.501 | 0.084 |
| Insecticide | Generation | LC50 (95% CI) (mg L−1) | Slope ± SE | ꭓ2 (df) | RF (95% CI) |
|---|---|---|---|---|---|
| Broflanilide | Susceptible (F0) | 0.025 (0.016–0.034) | 1.62 ± 0.14 | 2.04 (5) | - |
| Flux-SEL (F10) | 0.029 (0.022–0.037) | 1.49 ± 0.21 | 2.16 (5) | 1.16 (0.88–1.45) | |
| Chlorantraniliprole | Susceptible (F0) | 2.268 (2.079–2.350) | 2.31 ± 0.18 | 1.42 (5) | - |
| Flux-SEL (F10) | 3.231 (2.946–3.512) | 1.79 ± 0.22 | 1.35 (5) | 1.43 (0.97–1.75) | |
| Emamectin benzoate | Susceptible (F0) | 1.537 (1.414–1.628) | 2.57 ± 0.29 | 1.98 (5) | - |
| Flux-SEL (F10) | 3.197 (2.993–3.345) | 1.90 ± 0.15 | 2.12 (5) | 2.08 (1.69–2.43) | |
| Fipronil | Susceptible (F0) | 5.193 (4.827–5.376) | 1.49 ± 0.25 | 2.47 (5) | - |
| Flux-SEL (F10) | 6.803 (6.534–7.028) | 1.88 ± 0.18 | 1.52 (5) | 1.31 (0.92–1.67) | |
| Indoxacarb | Susceptible (F0) | 2.590 (2.380–2.764) | 3.07 ± 0.20 | 1.70 (5) | - |
| Flux-SEL (F10) | 3.056 (2.892–3.174) | 1.64 ± 0.19 | 1.32 (5) | 1.18 (0.86–1.40) | |
| Lambda cyhalothrin | Susceptible (F0) | 3.946 (3.751–4.188) | 1.81 ± 0.25 | 2.34 (5) | - |
| Flux-SEL (F10) | 4.893 (4.812–4.995) | 2.39 ± 0.16 | 1.49 (5) | 1.24 (0.95–1.52) | |
| Spinetoram | Susceptible (F0) | 0.039 (0.028–0.046) | 1.63 ± 0.22 | 1.25 (5) | - |
| Flux-SEL (F10) | 0.041 (0.033–0.052) | 1.56 ± 0.28 | 2.30 (5) | 1.05 (0.71–1.36) | |
| Tetraniliprole | Susceptible (F0) | 0.028 (0.019–0.040) | 1.45 ± 0.17 | 1.54 (5) | - |
| Flux-SEL (F10) | 0.039 (0.032–0.054) | 1.96 ± 0.21 | 2.80 (5) | 1.39 (0.98–1.75) |
| Generation | P450 (pmol min−1 mg pro−1) | Ratio | CarE (nmol min−1 mg pro−1) | Ratio | GST (μmol min−1 mg pro−1) | Ratio |
|---|---|---|---|---|---|---|
| Susceptible (F0) | 0.83 ± 0.05 a | - | 79.32 ± 1.21 a | - | 12.47 ± 0.82 a | - |
| Flux-SEL (F5) | 0.86 ± 0.08 a | 1.04 | 84.16 ± 1.54 a | 1.06 | 15.09 ± 1.04 ab | 1.21 |
| Flux-SEL (F10) | 0.94 ± 0.07 ab | 1.13 | 87.49 ± 1.67 a | 1.10 | 24.18 ± 0.75 b | 1.94 |
| Generation | Developmental Duration (day) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Egg | 1st Instar | 2nd Instar | 3rd Instar | 4th Instar | 5th Instar | 6th Instar | Pupa | Adult Longevity | ||
| Male | Female | |||||||||
| Susceptible (F0) | 3.19 ± 0.03 a | 2.09 ± 0.05 a | 2.89 ± 0.04 a | 3.07 ± 0.05 a | 2.97 ± 0.04 a | 3.16 ± 0.06 a | 5.96 ± 0.05 a | 12.35 ± 0.14 a | 11.81 ± 0.18 a | 12.42 ± 0.18 c |
| Flux-SEL (F5) | 3.06 ± 0.02 a | 2.13 ± 0.05 a | 2.96 ± 0.03 a | 2.99 ± 0.05 a | 3.03 ± 0.03 a | 3.19 ± 0.04 a | 6.24 ± 0.07 ab | 13.49 ± 0.09 b | 11.64 ± 0.23 a | 12.09 ± 0.21 b |
| Flux-SEL (F10) | 3.12 ± 0.03 a | 2.25 ± 0.06 b | 2.93 ± 0.04 a | 3.06 ± 0.03 a | 3.12 ± 0.05 a | 3.34 ± 0.06 b | 6.78 ± 0.10 b | 14.18 ± 0.12 b | 11.39 ± 0.15 a | 11.65 ± 0.13 a |
| Generation | Reproductive Traits | |||||
|---|---|---|---|---|---|---|
| Pre-Oviposition Period (day) | Oviposition Period (day) | Fecundity (Eggs Female−1) | Hatchability (%) | Pupation Rate (%) | Pupal Weight (g) | |
| Susceptible (F0) | 3.69 ± 0.18 a | 5.28 ± 0.14 a | 1296.17 ± 31.48 a | 97.82 ± 0.67 a | 82.14 ± 0.52 b | 0.34 ± 0.04 b |
| Flux-SEL (F5) | 4.05 ± 0.20 a | 5.91 ± 0.08 a | 1159.39 ± 47.26 a | 90.19 ± 1.09 a | 74.96 ± 0.78 ab | 0.29 ± 0.04 b |
| Flux-SEL (F10) | 3.87 ± 0.19 a | 7.06 ± 0.11 b | 1046.52 ± 29.07 a | 85.45 ± 1.32 a | 58.25 ± 1.15 a | 0.21 ± 0.06 a |
| Generation | Demographic Traits | ||||
|---|---|---|---|---|---|
| r (day−1) | λ (day−1) | R0 (Offspring/Individual) | T (day) | Rf | |
| Susceptible (F0) | 0.186 ± 0.01 a | 1.235 ± 0.01 a | 728.24 ± 54.31 b | 35.434 ± 0.26 b | - |
| Flux-SEL (F5) | 0.181 ± 0.01 a | 1.230 ± 0.01 a | 539.62 ± 76.25 b | 34.756 ± 0.21 ab | 0.741 |
| Flux-SEL (F10) | 0.175 ± 0.01 a | 1.207 ± 0.01 a | 257.14 ± 38.91 a | 31.712 ± 0.33 a | 0.353 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roy, D.; Biswas, S.; Sarkar, S.; Adhikary, S.; Chakraborty, G.; Sarkar, P.K.; Al-Shuraym, L.A.; Sayed, S.; Gaber, A.; Hossain, A. Risk Assessment of Fluxametamide Resistance and Fitness Costs in Fall Armyworm (Spodoptera frugiperda). Toxics 2023, 11, 307. https://doi.org/10.3390/toxics11040307
Roy D, Biswas S, Sarkar S, Adhikary S, Chakraborty G, Sarkar PK, Al-Shuraym LA, Sayed S, Gaber A, Hossain A. Risk Assessment of Fluxametamide Resistance and Fitness Costs in Fall Armyworm (Spodoptera frugiperda). Toxics. 2023; 11(4):307. https://doi.org/10.3390/toxics11040307
Chicago/Turabian StyleRoy, Debashis, Sujan Biswas, Sukamal Sarkar, Samrat Adhikary, Gautam Chakraborty, Pijush Kanti Sarkar, Laila A. Al-Shuraym, Samy Sayed, Ahmed Gaber, and Akbar Hossain. 2023. "Risk Assessment of Fluxametamide Resistance and Fitness Costs in Fall Armyworm (Spodoptera frugiperda)" Toxics 11, no. 4: 307. https://doi.org/10.3390/toxics11040307