
Epicatechin Inhibited Lipid Oxidation and Protein Lipoxidation in a Fish Oil-Fortified Dairy Mimicking System

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Emulsion Preparation
2.3. Analyses of Lipid Oxidation
2.4. Analyses of Protein Carbonylation
2.5. Total Sulfhydryl Groups
2.6. Intrinsic Tryptophan Fluorescence
2.7. Surface Hydrophobicity
2.8. Measurement of Protein Hydrolysis
2.9. SDS-PAGE Analysis
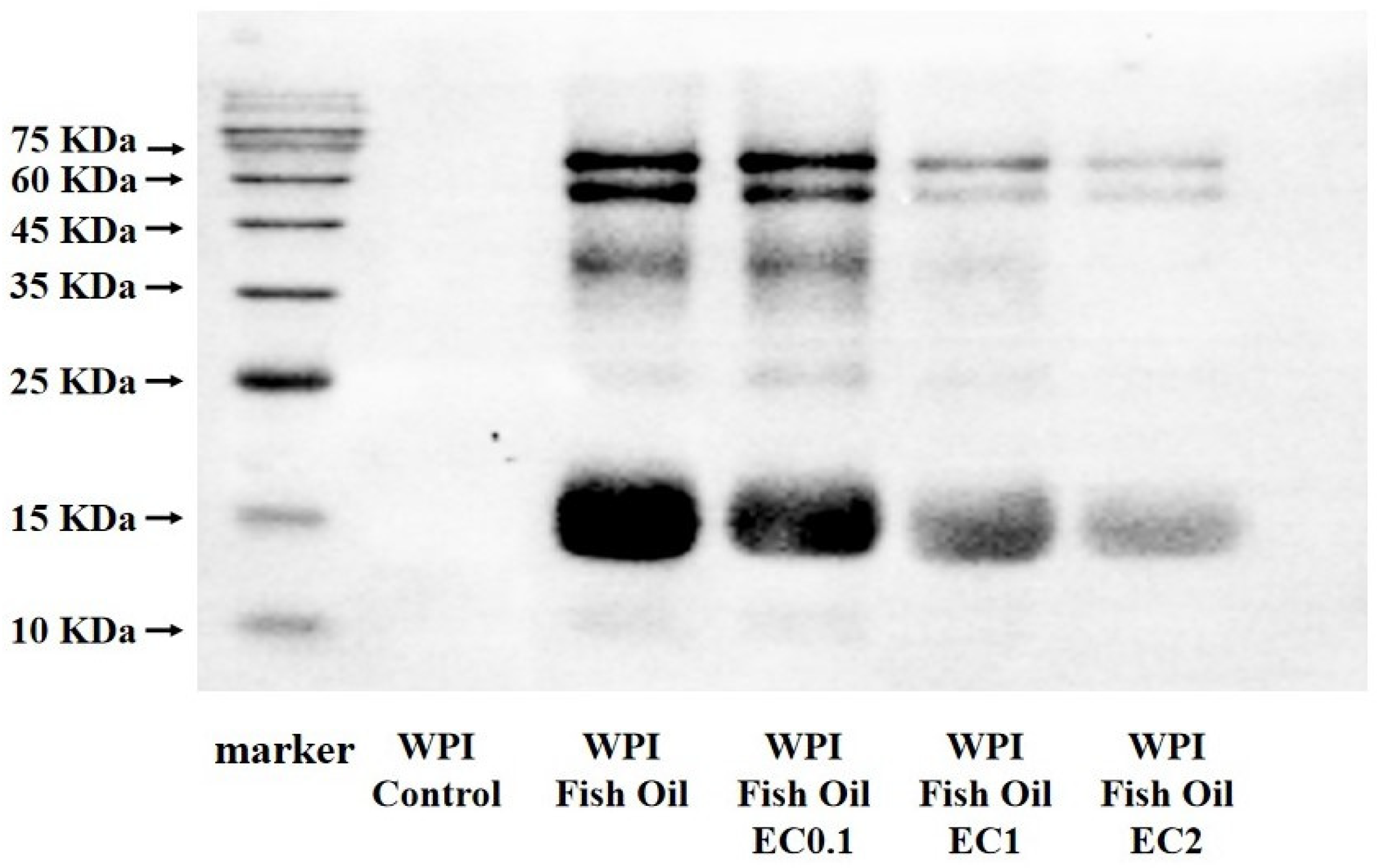
2.10. Western Blot Analysis
2.11. Scanning Electron Microscope (SEM) Observation
2.12. Data Analyses
3. Results and Analysis
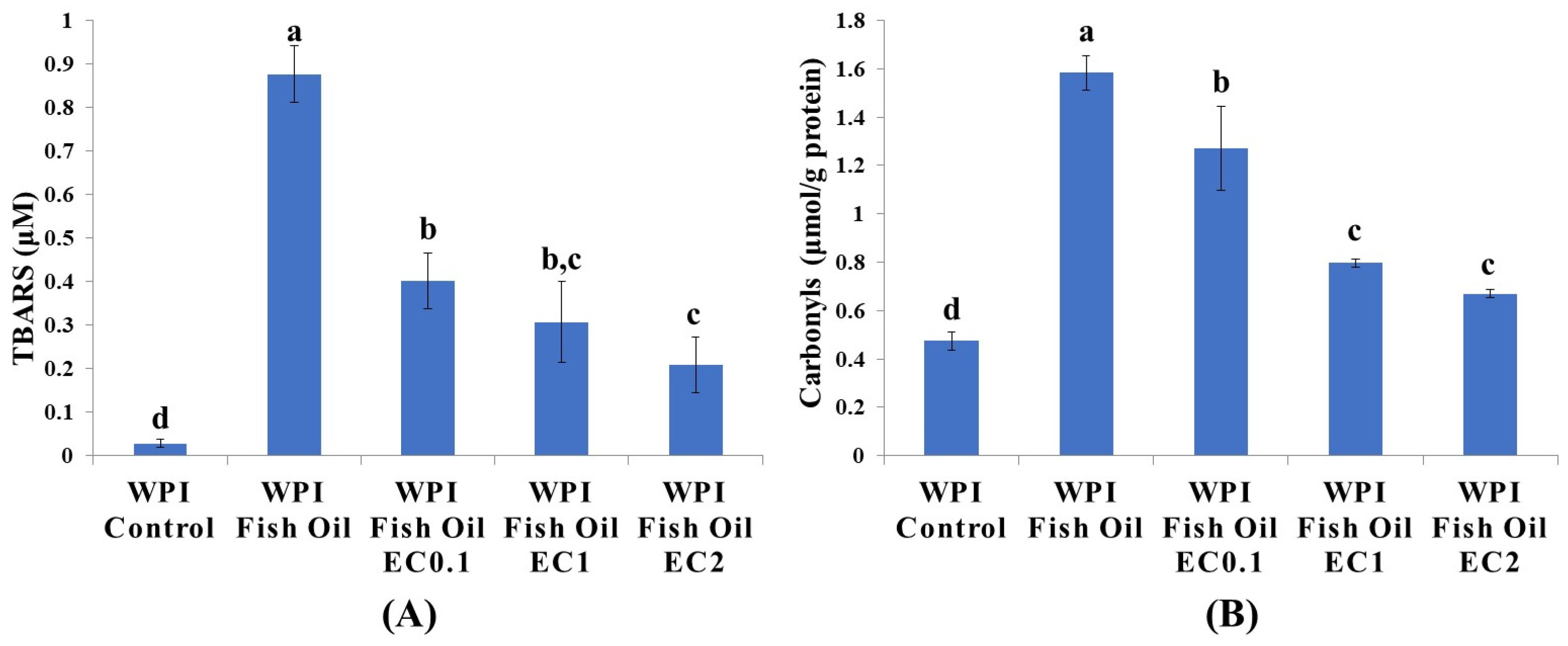
3.1. Lipid Oxidation
3.2. Protein Carbonylation
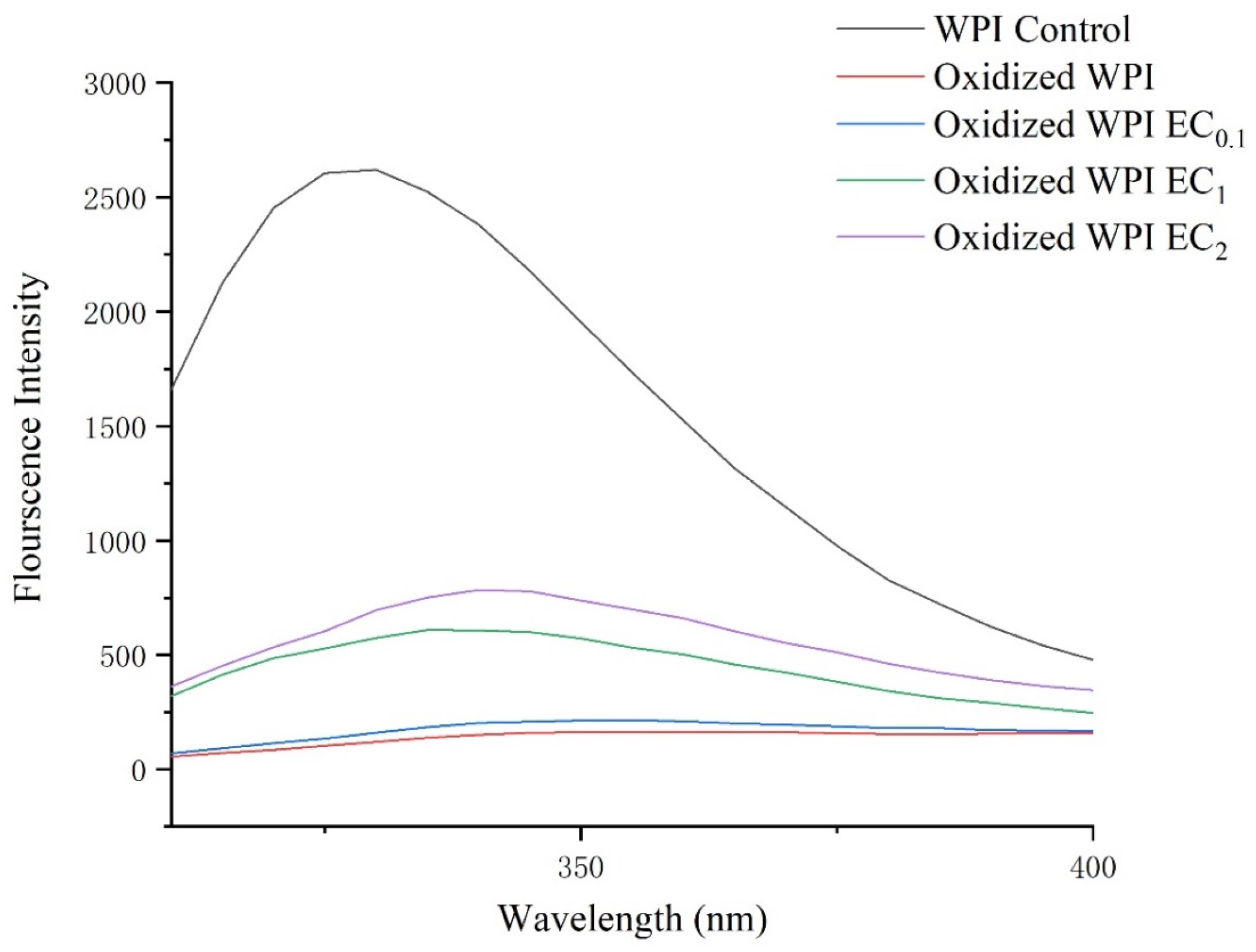
3.3. Intrinsic Fluorescence Changes
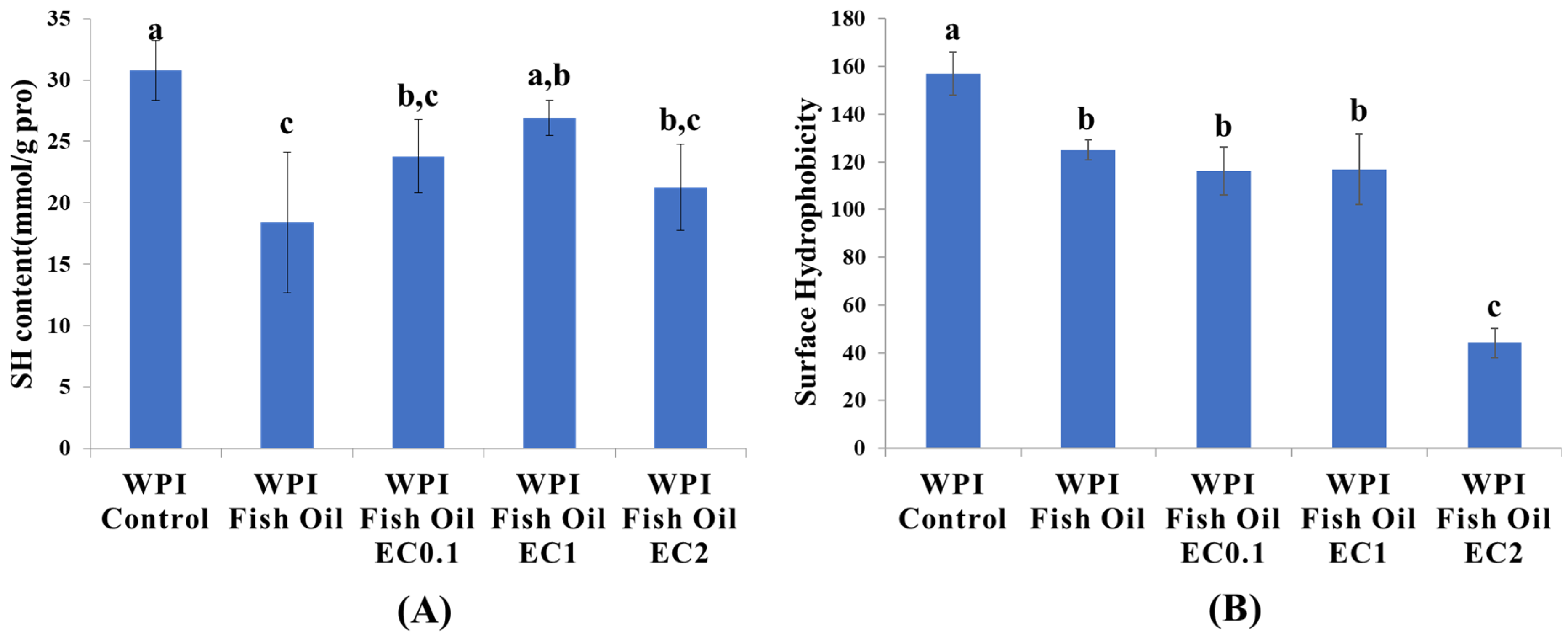
3.4. Protein Sulfhydryl Content
3.5. Protein Surface Hydrophobicity
3.6. Protein Hydrolysis
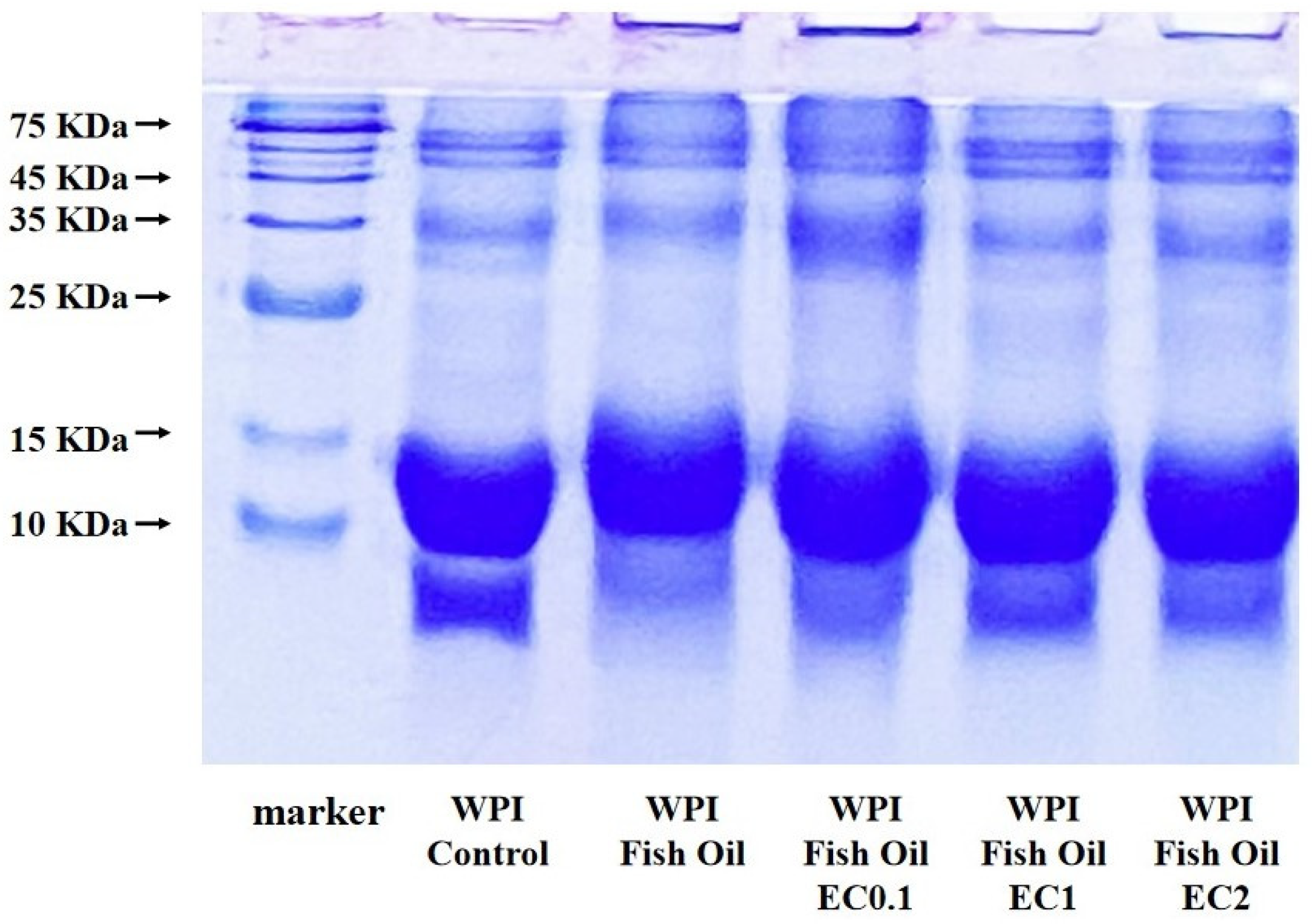
3.7. SDS-PAGE Analysis
3.8. Western Blot Analysis
3.9. Morphological Examination
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tapiero, H.; Ba, G.N.; Couvreur, P.; Tew, K.D. Polyunsaturated fatty acids (PUFA) and eicosanoids in human health and pathologies. Biomed. Pharmacother. 2002, 56, 215–222. [Google Scholar] [CrossRef]
- Chen, Y.; Sun, Y.; Ding, Y.; Ding, Y.; Liu, S.; Zhou, X.; Wu, H.; Xiao, J.; Lu, B. Recent progress in fish oil-based emulsions by various food-grade stabilizers: Fabrication strategy, interfacial stability mechanism and potential application. Crit. Rev. Food Sci. Nutr. 2022, 5, 1–24. [Google Scholar]
- Kanner, J. Dietary advanced lipid oxidation endproducts are risk factors to human health. Mol. Nutr. Food Res. 2007, 51, 1094–1101. [Google Scholar] [CrossRef] [PubMed]
- Aldini, G.; Domingues, M.R.; Spickett, C.M.; Domingues, P.; Altomare, A.; Sánchez-Gómez, F.J.; Oeste, C.L.; Pérez-Sala, D. Protein lipoxidation: Detection strategies and challenges. Redox Biol. 2015, 5, 253–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, M.; Losada-Barreiro, S.; Paiva-Martins, F.; Bravo-Diaz, C. Polyphenolic antioxidants in lipid emulsions: Partitioning effects and interfacial phenomena. Foods 2021, 10, 539. [Google Scholar] [CrossRef]
- Gutiérrez-Del-Río, I.; López-Ibáñez, S.; Magadán-Corpas, P.; Fernández-Calleja, L.; Pérez-Valero, A.; Tuñón-Granda, M.; Miguélez, E.; Villar, C.; Lombó, F. Terpenoids and polyphenols as natural antioxidant agents in food preservation. Antioxidants 2021, 10, 1264. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Wang, Y.; Pan, D.; Li, Y.; Li, R.; Xu, B.; Zhou, C. Individual effects of rosemary extract and green tea polyphenols on the physicochemical properties of soybean oil–myosin emulsion with L-arginine or L-lysine. Food Chem. 2022, 395, 133583. [Google Scholar] [CrossRef]
- Leopoldini, M.; Russo, N.; Toscano, M. The molecular basis of working mechanism of natural polyphenolic antioxidants. Food Chem. 2011, 125, 288–306. [Google Scholar] [CrossRef]
- Lund, M.N.; Heinonen, M.; Baron, C.P.; Estévez, M. Protein oxidation in muscle foods: A review. Mol. Nutr. Food Res. 2011, 55, 83–95. [Google Scholar] [CrossRef]
- Tian, L.; Zhang, S.; Yi, J.; Zhu, Z.; Cui, L.; Decker, E.A.; McClements, D.J. Factors impacting the antioxidant/prooxidant activity of tea polyphenols on lipids and proteins in oil-in-water emulsions. LWT 2022, 156, 113024. [Google Scholar] [CrossRef]
- Paddon-Jones, D.; Short, K.R.; Campbell, W.W.; Volpi, E.; Wolfe, R.R. Role of dietary protein in the sarcopenia of aging. Am. J. Clin. Nutr. 2008, 87, 1562S–1566S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obando, M.; Papastergiadis, A.; Li, S.; De Meulenaer, B. Impact of lipid and protein co-oxidation on digestibility of dairy proteins in oil-in-water (O/W) emulsions. J. Agric. Food Chem. 2015, 63, 9820–9830. [Google Scholar] [CrossRef] [PubMed]
- Salih, A.M.; Smith, D.M.; Price, J.F.; Dawson, L.E. Modified extraction 2-thiobarbituric acid method for measuring lipid oxidation in poultry. Poult. Sci. 1987, 66, 1483–1488. [Google Scholar] [CrossRef] [PubMed]
- Levine, R.L.; Garland, D.; Oliver, C.N.; Amici, A.; Climent, I.; Lenz, A.G.; Ahn, B.W.; Shaltiel, S.; Stadtman, E.R. Determination of carbonyl content in oxidatively modified proteins. Methods Enzymol. 1990, 186, 464–478. [Google Scholar]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Wu, W.; Zhang, C.; Hua, Y. Structural modification of soy protein by the lipid peroxidation product malondialdehyde. J. Sci. Food Agric. 2009, 89, 1416–1423. [Google Scholar] [CrossRef]
- Li, W.; Gao, H.; Mu, H.; Chen, H.; Fang, X.; Zhou, Y.; Tao, F. Three different active aldehydes induce the production of advanced lipoxidation end products upon incubation with bovine serum albumin. Eur. J. Lipid Sci. Technol. 2015, 117, 1432–1443. [Google Scholar] [CrossRef]
- Church, F.C.; Swaisgood, H.E.; Porter, D.H.; Catignani, G.L. Spectrophotometric assay using o-phthaldialdehyde for determination of proteolysis in milk and isolated milk proteins. J. Dairy Sci. 1983, 66, 1219–1227. [Google Scholar] [CrossRef]
- Yi, J.; Qiu, M.; Liu, N.; Tian, L.; Zhu, X.; Decker, E.A.; McClements, D.J. Inhibition of lipid and protein oxidation in whey-protein-stabilized emulsions using a natural antioxidant: Black rice anthocyanins. J. Agric. Food Chem. 2020, 68, 10149–10156. [Google Scholar] [CrossRef]
- Tian, L.; Kejing, Y.; Zhang, S.; Yi, J.; Zhu, Z.; Decker, E.A.; McClements, D.J. Impact of tea polyphenols on the stability of oil-in-water emulsions coated by whey proteins. Food Chem. 2021, 343, 128448. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, L.; Liu, Y.; Li, J. Effects of antioxidants, proteins, and their combination on emulsion oxidation. Crit. Rev. Food Sci. Nutr. 2022, 62, 8137–8160. [Google Scholar] [CrossRef]
- Hellwig, M. The chemistry of protein oxidation in food. Angew. Chem. Int. Ed. 2019, 58, 16742–16763. [Google Scholar] [CrossRef]
- Fenaille, F.; Parisod, V.; Tabet, J.-C.; Guy, P.A. Carbonylation of milk powder proteins as a consequence of processing conditions. Proteomics 2005, 5, 3097–3104. [Google Scholar] [CrossRef]
- Ronsein, G.E.; de Oliveira, M.C.B.; de Medeiros, M.H.G.; Di Mascio, P. Mechanism of dioxindolylalanine formation by singlet molecular oxygen-mediated oxidation of tryptophan residues. Photochem. Photobiol. Sci. 2011, 10, 1727–1730. [Google Scholar] [CrossRef] [PubMed]
- Krämer, A.C.; Thulstrup, P.W.; Lund, M.N.; Davies, M.J. Key role of cysteine residues and sulfenic acids in thermal- and H2O2-mediated modification of beta-lactoglobulin. Free Radic. Biol. Med. 2016, 97, 544–555. [Google Scholar] [CrossRef]
- Niu, X.; Wang, X.; Han, Y.; Lu, C.; Chen, X.; Wang, T.; Xu, M.; Zhu, Q. Influence of malondialdehyde-induced modifications on physicochemical and digestibility characteristics of whey protein isolate. J. Food Biochem. 2019, 43, e13041. [Google Scholar] [CrossRef] [PubMed]
- Burstein, E.A.; Vedenkina, N.S.; Ivkova, M.N. Fluorescence and the location of tryptophan residues in protein molecules. Photochem. Photobiol. 1973, 18, 263–279. [Google Scholar] [CrossRef]
- Lund, M.N. Reactions of plant polyphenols in foods: Impact of molecular structure. Trends Food Sci. Technol. 2021, 112, 241–251. [Google Scholar] [CrossRef]
- Xiong, Y.L.; Guo, A. Animal and plant protein oxidation: Chemical and functional property significance. Foods 2020, 10, 40. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Zhang, S.; Yi, J.; Zhu, Z.; Li, M.; Decker, E.A.; McClements, D.J. Formation of antioxidant multilayered coatings for the prevention of lipid and protein oxidation in oil-in-water emulsions: Lycium barbarum polysaccharides and whey proteins. J. Agric. Food Chem. 2021, 69, 15691–15698. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Gao, X.; Hao, M.; Tang, L. Comparison of binding interaction between β-lactoglobulin and three common polyphenols using multi-spectroscopy and modeling methods. Food Chem. 2017, 228, 143–151. [Google Scholar] [CrossRef]
- Cai, S.; Huang, J.; Wang, L.-L.; Dong, X.-R.; Gong, Y.-S.; Li, J.; Li, Q.; Liu, Z.-H.; Luo, G.-A. Inhibiting effects of epigallocatechin gallate (EGCG) on the formation of age pigment in vitro and in vivo. J. Med. Plants Res. 2011, 5, 5470–5478. [Google Scholar]
- Nivala, O.; Nordlund, E.; Kruus, K.; Ercili-Cura, D. The effect of heat and transglutaminase treatment on emulsifying and gelling properties of faba bean protein isolate. LWT 2021, 139, 110517. [Google Scholar] [CrossRef]
- Obando, M.; Soto, E.; De Meulenaer, B. Influence of oxidized oils on digestibility of caseins in o/w emulsions. Eur. J. Lipid Sci. Technol. 2018, 120, 1700331. [Google Scholar] [CrossRef] [Green Version]
- Vandemoortele, A.; De Meulenaer, B. Behavior of malondialdehyde in oil-in-water emulsions. J. Agric. Food Chem. 2015, 63, 5694–5701. [Google Scholar] [CrossRef] [PubMed]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Li, S.; Huang, J.; Wei, C.-C.; Wan, X.; Sang, S.; Ho, C.-T. Importance of the nucleophilic property of tea polyphenols. J. Agric. Food Chem. 2019, 67, 5379–5383. [Google Scholar] [CrossRef]
- Tian, L.; Zhang, S.; Yi, J.; Zhu, Z.; Cui, L.; Decker, E.A.; McClements, D.J. Antioxidant and prooxidant activities of tea polyphenols in oil-in-water emulsions depend on the level used and the location of proteins. Food Chem. 2022, 375, 131672. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, Y.; Ohland, C.; Jobin, C.; Sang, S. Microbiota facilitates the formation of the aminated metabolite of green tea polyphenol (-)-epigallocatechin-3-gallate which trap deleterious reactive endogenous metabolites. Free Radic. Biol. Med. 2019, 131, 332–344. [Google Scholar] [CrossRef]
- Sun, W.; Zhou, F.; Zhao, M.; Yang, B.; Cui, C. Physicochemical changes of myofibrillar proteins during processing of Cantonese sausage in relation to their aggregation behaviour and in vitro digestibility. Food Chem. 2011, 129, 472–478. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Time (min) | |||
|---|---|---|---|---|
| Gastric Tract | Intestinal Tract | |||
| 30 | 60 | 90 | 180 | |
| WPI Control | 4.20 ± 0.3 a | 4.51 ± 0.3 a | 6.76 ± 1.5 a | 13.92 ± 0.4 b |
| Oxidized WPI | 6.52 ± 0.3 a | 8.07 ± 0.8 b | 9.85 ± 0.3 a,b | 8.22 ± 2.7 a |
| Oxidized WPI EC0.1 | 6.32 ± 0.5 a | 8.01 ± 1.7 b | 7.41 ± 0.8 a | 11.40 ± 0.6 a |
| Oxidized WPI EC1 | 10.82 ± 2.1 b | 11.11 ± 2.3 b,c | 14.20 ± 2.0 c | 12.04 ± 0.7 a |
| Oxidized WPI EC2 | 10.03 ± 1.1 b | 12.02 ± 1.1 c | 13.13 ± 2.4 b,c | 14.95 ± 0.7 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lian, Z.; Han, J.; Cao, Y.; Yao, W.; Niu, X.; Xu, M.; Xu, J.; Zhu, Q. Epicatechin Inhibited Lipid Oxidation and Protein Lipoxidation in a Fish Oil-Fortified Dairy Mimicking System. Foods 2023, 12, 1559. https://doi.org/10.3390/foods12071559
Lian Z, Han J, Cao Y, Yao W, Niu X, Xu M, Xu J, Zhu Q. Epicatechin Inhibited Lipid Oxidation and Protein Lipoxidation in a Fish Oil-Fortified Dairy Mimicking System. Foods. 2023; 12(7):1559. https://doi.org/10.3390/foods12071559
Chicago/Turabian StyleLian, Zhenghao, Jiahui Han, Yue Cao, Wenhua Yao, Xiaoying Niu, Mingfeng Xu, Jun Xu, and Qin Zhu. 2023. "Epicatechin Inhibited Lipid Oxidation and Protein Lipoxidation in a Fish Oil-Fortified Dairy Mimicking System" Foods 12, no. 7: 1559. https://doi.org/10.3390/foods12071559