
Molecular Docking Revealed the Potential Anti-Oxidative Stress Mechanism of the Walnut Polypeptide on HT22 Cells
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Walnut Protein Hydrolysis Products
2.3. Ultrafiltration
2.4. ABTS•+ Radical Scavenging Activity Assay
2.5. DPPH Radical Scavenging Activity Assay
2.6. Hydroxyl Radical (•OH) Scavenging Activity
2.7. Cell Culture and Cytotoxicity Assay
2.7.1. Cell Culture
2.7.2. Cytotoxicity Assay
2.8. Determination of ROS in HT22 Cells
2.9. Determination of SOD, CAT, and GSH-Px in HT22 Cells
2.10. Identification of Peptide Sequences by LC-MS/MS
2.11. Computer Analysis of Identified Peptides
2.12. Molecular Docking
2.13. Statistics Analysis
3. Results
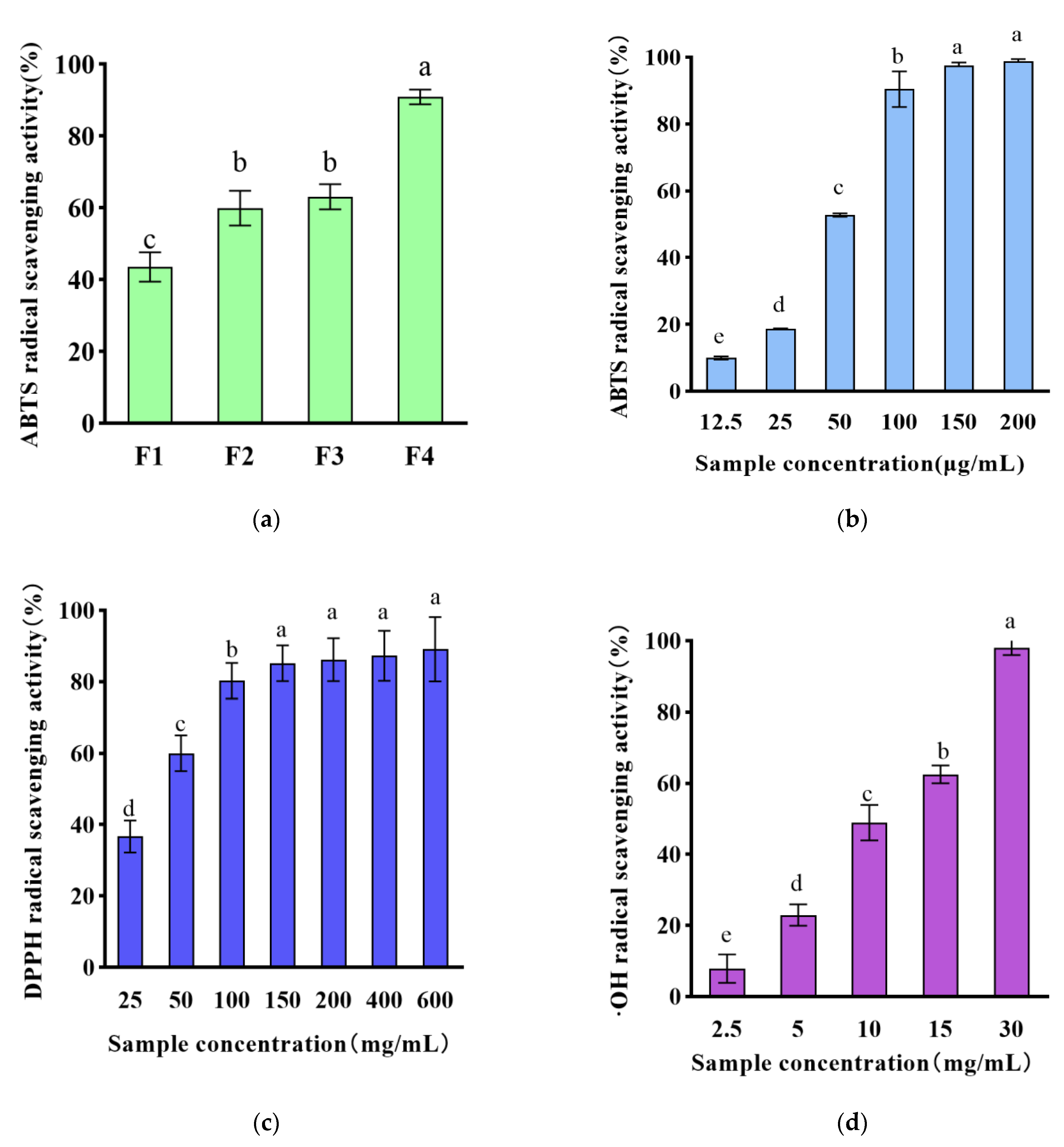
3.1. Chemical Antioxidant Activity of WPH Ultrafiltration Fractions
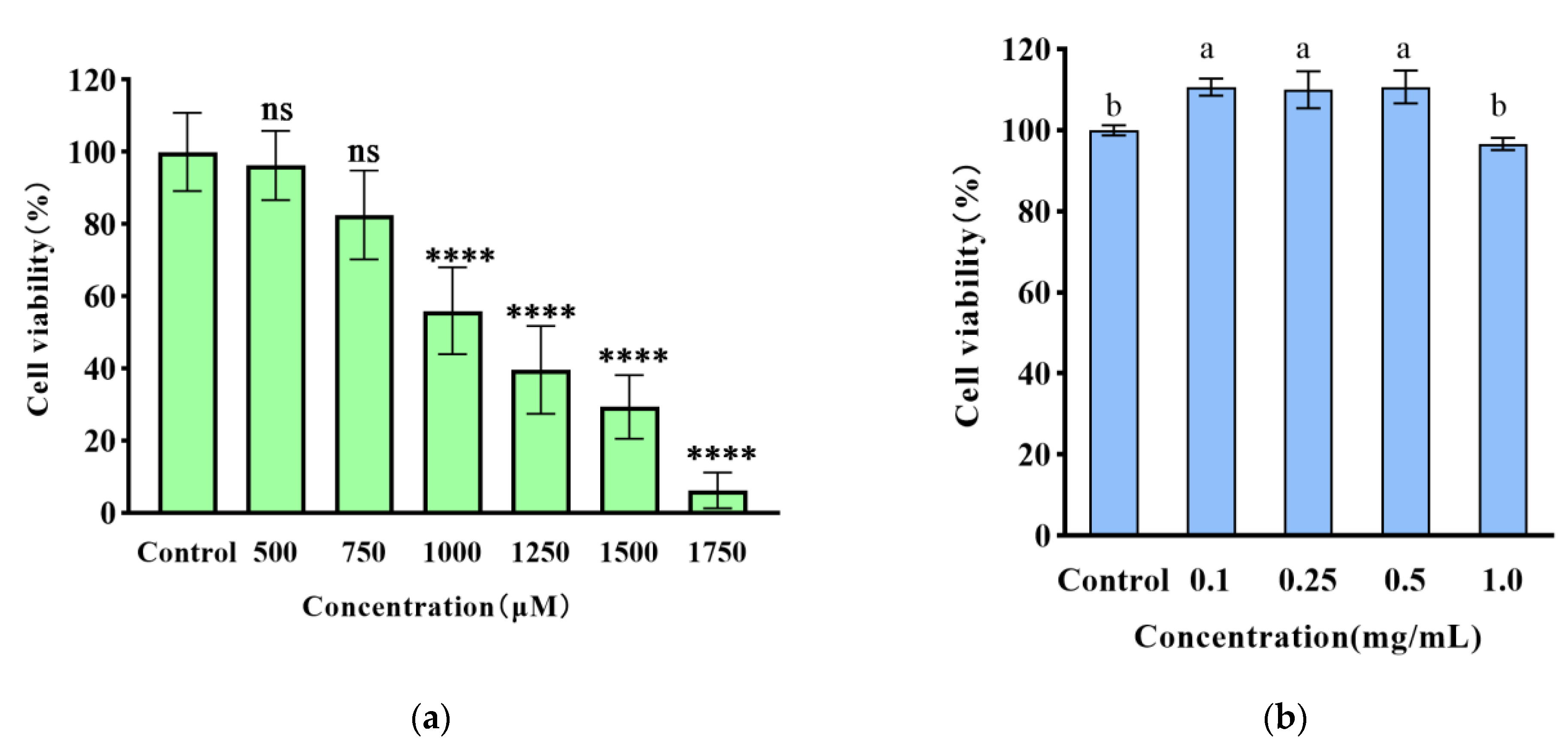
3.2. The Cytoprotective Effect of WPH on H2O2-Induced Damage in HT22 Cells
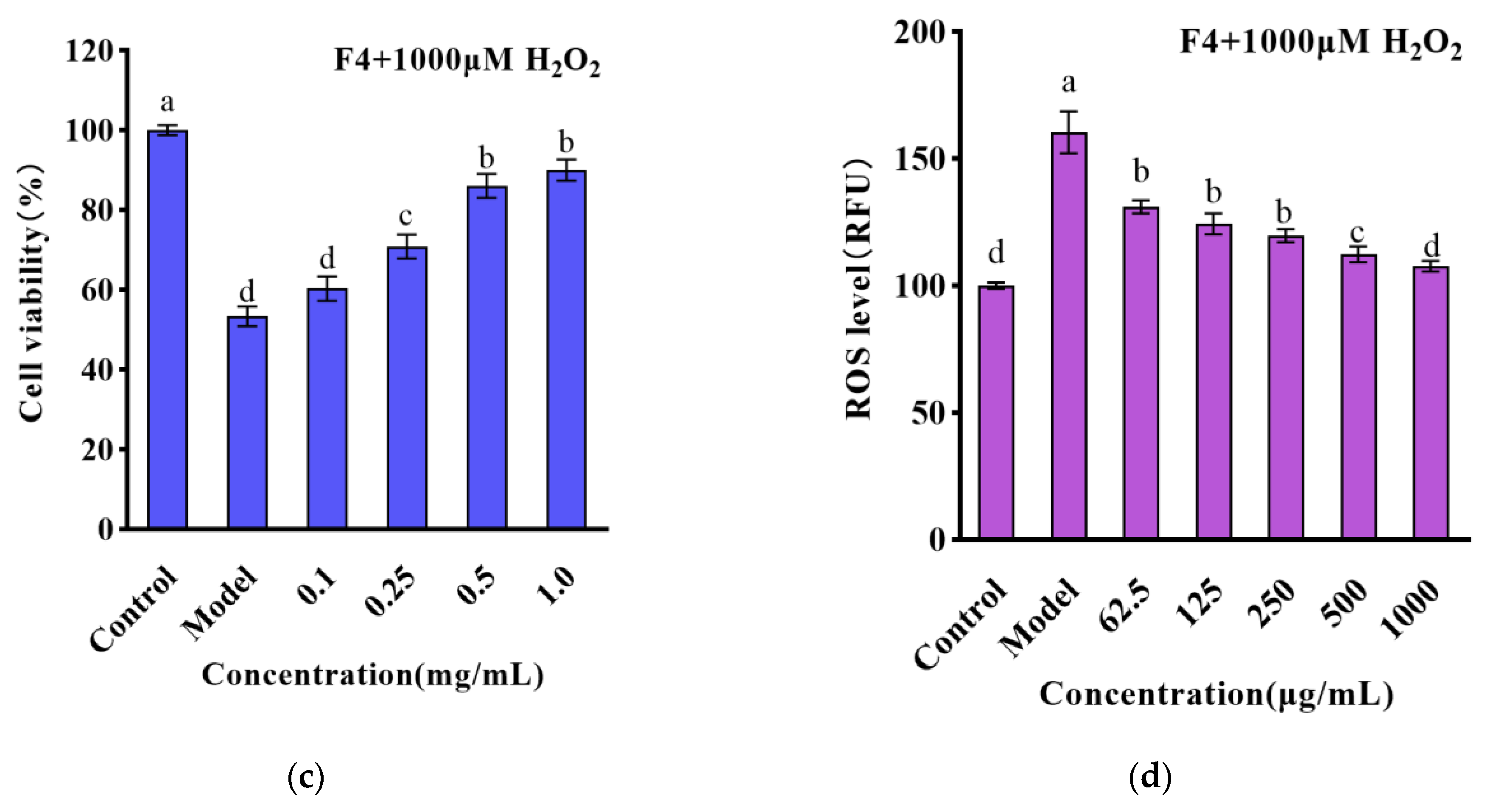
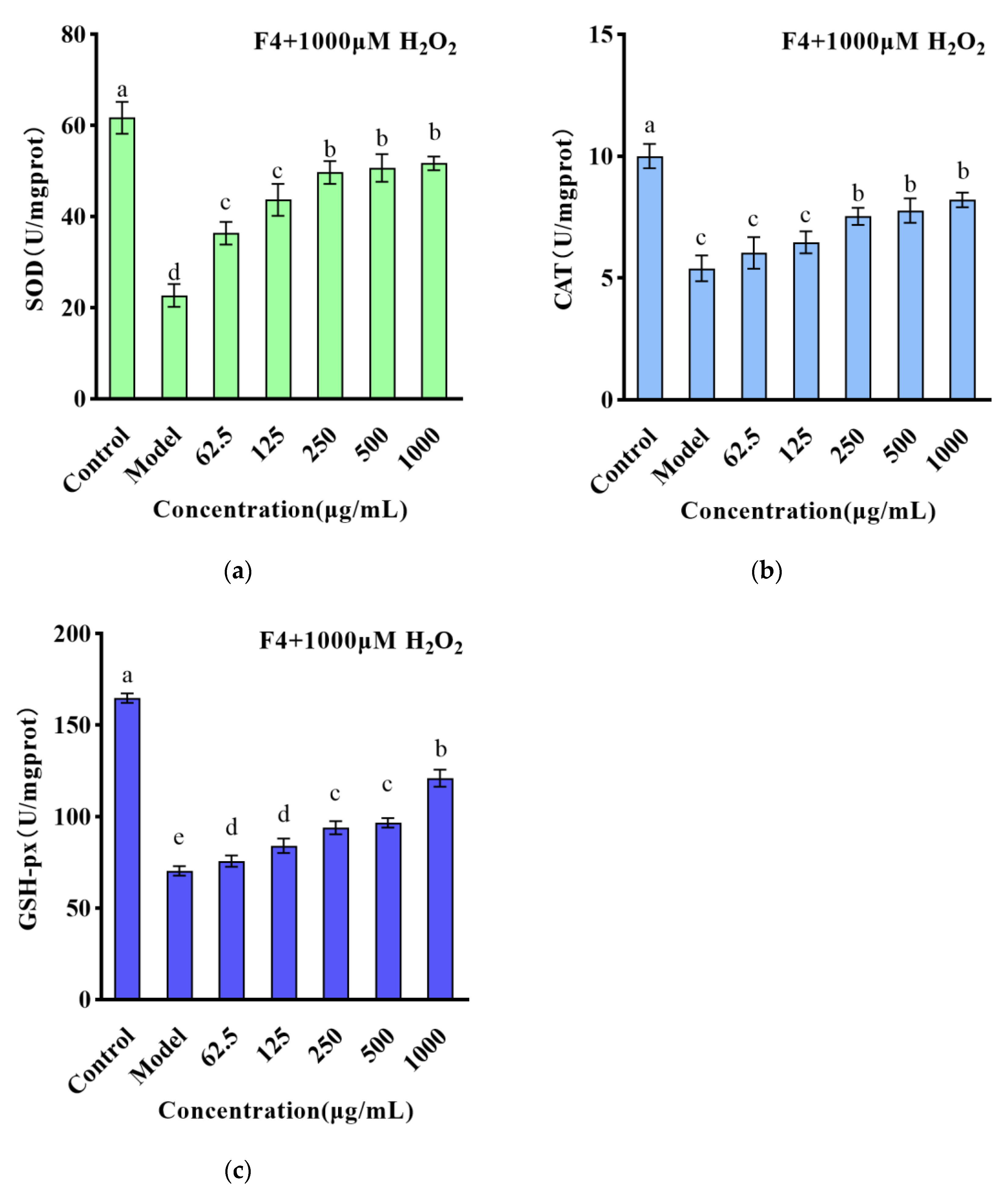
3.3. The Effect of Small Molecule WPH Fractions on Intracellular ROS and Antioxidant Enzymes in HT22 Cells
3.4. Identification of Antioxidant Peptides
3.5. A Molecular Docking Analysis of Keap1 and Peptides Derived from Walnut Proteins
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Changwen, L.; Lihua, Z.; Wei, Z. Progress on preparation and functional activity of walnut peptides. Food Ind. 2020, 41, 257–259. [Google Scholar]
- Martinez, M.; Labuckas, D.; Lamarque, A.; Maestri, D. Walnut (Juglans regia L.): Genetic resources, chemistry, by-products. J. Sci. Food Agric. 2010, 90, 1959–1967. [Google Scholar] [CrossRef] [PubMed]
- Yuntao, H.; Na, Z.; Yong, L. Research progress on preparation technology of walnut peptides. Food Ind. 2020, 41, 253–257. [Google Scholar]
- Zhou, X.; Peng, X.; Pei, H.; Chen, Y.; Meng, H.; Yuan, J.; Xing, H.; Wu, Y. An overview of walnuts application as a plant-based. Front. Endocrinol. 2022, 13, 1083707. [Google Scholar] [CrossRef]
- Tapia, M.I.; Sánchez-Morgado, J.R.; García-Parra, J.; Ramírez, R.; Hernández, T.; González-Gómez, D. Comparative study of the nutritional and bioactive compounds content of four walnut (Juglans regia L.) cultivars. J. Food Compos. Anal. 2013, 31, 232–237. [Google Scholar] [CrossRef]
- Jiang, L.; Xu, H.-D.; Chen, J.-H. Functional of walnut polypeptide. Food Sci. Technol. 2013, 38, 228–231. [Google Scholar]
- Hayes, D.; Angove, M.J.; Tucci, J.; Dennis, C. Walnuts (Juglans regia) Chemical Composition and Research in Human Health. Crit. Rev. Food Sci. Nutr. 2016, 56, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Gharibzahedi, S.M.T.; Mousavi, S.M.; Hamedi, M.; Khodaiyan, F. Determination and characterization of kernel biochemical composition and functional compounds of Persian walnut oil. J. Food Sci. Technol. 2014, 51, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Su, G.; Zhang, Q.; Zhao, T.; Liu, Y.; Zheng, L.; Zhao, M. Walnut (Juglans regia) Peptides Reverse Sleep Deprivation-Induced Memory Impairment in Rat via Alleviating Oxidative Stress. J. Agric. Food Chem. 2018, 66, 10617–10627. [Google Scholar] [CrossRef]
- Zhao, F.; Wang, J.; Lu, H.; Fang, L.; Qin, H.; Liu, C.; Min, W. Neuroprotection by Walnut-Derived Peptides through Autophagy Promotion via Akt/mTOR Signaling Pathway against Oxidative Stress in PC12 Cells. J. Agric. Food Chem. 2020, 68, 3638–3648. [Google Scholar] [CrossRef]
- Azzi, A. Oxidative Stress: What Is It? Can It Be Measured? Where Is It Located? Can It Be Good or Bad? Can It Be Prevented? Can It Be Cured? Antioxidants 2022, 11, 1431. [Google Scholar] [CrossRef]
- Burgos-Morón, E.; Abad-Jiménez, Z.; de Marañón, A.M.; Iannantuoni, F.; López, E.; López-Domènech, S.; Salom, C.; Jover, A.; Mora, V.; Roldan, I.; et al. Relationship between Oxidative Stress, ER Stress, and Inflammation in Type 2 Diabetes: The Battle Continues. J. Clin. Med. 2019, 8, 1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teleanu, D.M.; Niculescu, A.-G.; Lungu, I.I.; Radu, C.I.; Vladâcenco, O.; Roza, E.; Costăchescu, B.; Grumezescu, A.M.; Teleanu, R.I. An Overview of Oxidative Stress, Neuroinflammation, and Neurodegenerative Diseases. Int. J. Mol. Sci. 2022, 23, 5938. [Google Scholar] [CrossRef]
- Stanzione, F.; Giangreco, I.; Cole, J.C. Use of molecular docking computational tools in drug discovery. Prog. Med. Chem. 2021, 60, 273–343. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.J.; Della Corte, D. Using molecular docking and molecular dynamics to investigate protein-ligand interactions. Mod. Phys. Lett. B 2021, 35, 2130002. [Google Scholar] [CrossRef]
- Gao, J.; Li, T.; Chen, D.; Gu, H.; Mao, X. Identification and molecular docking of antioxidant peptides from hemp seed protein hydrolysates. LWT 2021, 147, 111453. [Google Scholar] [CrossRef]
- Hu, X.; Ren, J.; Zhao, M.; Cui, C.; He, P. Emulsifying properties of the transglutaminase-treated crosslinked product between peanut protein and fish (Decapterus maruadsi) protein hydrolysates. J. Sci. Food Agric. 2011, 91, 578–585. [Google Scholar] [CrossRef]
- Wen, L.; Jiang, Y.; Zhou, X.; Bi, H.; Yang, B. Structure identification of soybean peptides and their immunomodulatory activity. Food Chem. 2021, 359, 129970. [Google Scholar] [CrossRef]
- Corsetto, P.A.; Montorfano, G.; Zava, S.; Colombo, I.; Ingadottir, B.; Jonsdottir, R.; Sveinsdottir, K.; Rizzo, A.M. Characterization of Antioxidant Potential of Seaweed Extracts for Enrichment of Convenience Food. Antioxidants 2020, 9, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, X.; Xiong, Y.L.; Kong, B. Antioxidant activity of peptide fractions from whey protein hydrolysates as measured by electron spin resonance. Food Chem. 2009, 113, 196–201. [Google Scholar] [CrossRef]
- Qin, T.; Ren, Z.; Liu, X.; Luo, Y.; Long, Y.; Peng, S.; Chen, S.; Zhang, J.; Ma, Y.; Li, J.; et al. Study of the selenizing Codonopsis pilosula polysaccharides protects RAW264.7 cells from hydrogen peroxide-induced injury. Int. J. Biol. Macromol. 2019, 125, 534–543. [Google Scholar] [CrossRef] [PubMed]
- Iram, D.; Sansi, M.S.; Zanab, S.; Vij, S.; Ashutosh; Meena, S. In silico identification of antidiabetic and hypotensive potential bioactive peptides from the sheep milk proteins—A molecular docking study. J. Food Biochem. 2022, 46, e14137. [Google Scholar] [CrossRef]
- Mishra, K.; Ojha, H.; Chaudhury, N.K. Estimation of antiradical properties of antioxidants using DPPH assay: A critical review and results. Food Chem. 2012, 130, 1036–1043. [Google Scholar] [CrossRef]
- Hu, X.; Pan, C.; Cai, M.; Li, L.; Yang, X.; Xiang, H.; Chen, S. Novel Antioxidant Peptides from Grateloupia livida Hydrolysates: Purification and Identification. Foods 2022, 11, 1498. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhang, D.; Liu, M.; Li, Y.; Lv, R.; Li, X.; Wang, Q.; Ren, D.; Wu, L.; Zhou, H. Identification of Antioxidant Peptides Derived from Tilapia (Oreochromis niloticus) Skin and Their Mechanism of Action by Molecular Docking. Foods 2022, 11, 2576. [Google Scholar] [CrossRef]
- Devita, L.; Lioe, H.N.; Nurilmala, M.; Suhartono, M.T. The Bioactivity Prediction of Peptides from Tuna Skin Collagen Using Integrated Method Combining In Vitro and In Silico. Foods 2021, 10, 2739. [Google Scholar] [CrossRef]
- Nwachukwu, I.D.; Aluko, R.E. Structural and functional properties of food protein-derived antioxidant peptides. J. Food Biochem. 2019, 43, e12761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granato, D.; Shahidi, F.; Wrolstad, R.; Kilmartin, P.; Melton, L.D.; Hidalgo, F.J.; Miyashita, K.; van Camp, J.; Alasalvar, C.; Ismail, A.B.; et al. Antioxidant activity, total phenolics and flavonoids contents: Should we ban in vitro screening methods? Food Chem. 2018, 264, 471–475. [Google Scholar] [CrossRef]
- Wen, C.; Zhang, J.; Feng, Y.; Duan, Y.; Ma, H.; Zhang, H. Purification and identification of novel antioxidant peptides from watermelon seed protein hydrolysates and their cytoprotective effects on H2O2-induced oxidative stress. Food Chem. 2020, 327, 127059. [Google Scholar] [CrossRef]
- Wong, F.-C.; Xiao, J.; Wang, S.; Ee, K.-Y.; Chai, T.-T. Advances on the antioxidant peptides from edible plant sources. Trends Food Sci. Technol. 2020, 99, 44–57. [Google Scholar] [CrossRef]
- Hajfathalian, M.; Ghelichi, S.; García-Moreno, P.J.; Sørensen, A.-D.M.; Jacobsen, C. Peptides: Production, bioactivity, functionality, and applications. Crit. Rev. Food Sci. Nutr. 2018, 58, 3097–3129. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Zhang, C.; Chen, Y.; Zheng, B. Purification and Characterization of Antioxidant Peptides of Pseudosciaena crocea Protein Hydrolysates. Molecules 2017, 22, 57. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Tong, X.; Li, Y.; Wang, H.; Wang, Z.; Qi, B.; Sui, X.; Jiang, L. Purification and Characterization of Antioxidant Peptides from Alcalase-Hydrolyzed Soybean (Glycine max L.) Hydrolysate and Their Cytoprotective Effects in Human Intestinal Caco-2 Cells. J. Agric. Food Chem. 2019, 67, 5772–5781. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Zhang, J.; Zhang, H.; Duan, Y.; Ma, H. Study on the structure–activity relationship of watermelon seed antioxidant peptides by using molecular simulations. Food Chem. 2021, 364, 130432. [Google Scholar] [CrossRef]
- Jeong, W.-S.; Jun, M.; Kong, A.-N.T. Nrf2: A Potential Molecular Target for Cancer Chemoprevention by Natural Compounds. Antioxid. Redox Signal. 2006, 8, 99–106. [Google Scholar] [CrossRef]
- Adams, J.; Kelso, R.; Cooley, L. The kelch repeat superfamily of proteins: Propellers of cell function. Trends Cell Biol. 2000, 10, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Liang, F.; Cao, W.; Huang, Y.; Fang, Y.; Cheng, Y.; Pan, S.; Xu, X. Isoflavone biochanin A, a novel nuclear factor erythroid 2-related factor 2 (Nrf2)-antioxidant response element activator, protects against oxidative damage in HepG2 cells. Biofactors 2019, 45, 563–574. [Google Scholar] [CrossRef]
- Wakabayashi, N.; Dinkova-Kostova, A.T.; Holtzclaw, W.D.; Kang, M.-I.; Kobayashi, A.; Yamamoto, M.; Kensler, T.W.; Talalay, P. Protection against electrophile and oxidant stress by induction of the phase 2 response: Fate of cysteines of the Keap1 sensor modified by inducers. Proc. Natl. Acad. Sci. USA 2004, 101, 2040–2045. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Ma, M.; Yu, Z.; Du, S.-K. Preparation and identification of antioxidant peptides from cottonseed proteins. Food Chem. 2021, 352, 129399. [Google Scholar] [CrossRef]
- Lo, S.-C.; Li, X.; Henzl, M.T.; Beamer, L.J.; Hannink, M. Structure of the Keap1:Nrf2 interface provides mechanistic insight into Nrf2 signaling. EMBO J. 2006, 25, 3605–3617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Lu, A.; Sun, Y.; Liu, B.; Zhang, J.; Zhang, L.; Huang, P.; Yang, A.; Li, Z.; Cao, Y.; et al. Purification and identification of antioxidant and angiotensin converting enzyme-inhibitory peptides from Guangdong glutinous rice wine. LWT 2022, 169, 113953. [Google Scholar] [CrossRef]
- Ren, L.-K.; Yang, Y.; Ma, C.-M.; Fan, J.; Bian, X.; Liu, B.-X.; Wang, D.-F.; Zhu, P.-Y.; Fu, Y.; Zhang, N. Identification and in silico analysis of novel antioxidant peptides in broken rice protein hydrolysate and its cytoprotective effect against H2O2-induced 2BS cell model. Food Res. Int. 2022, 162, 112108. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
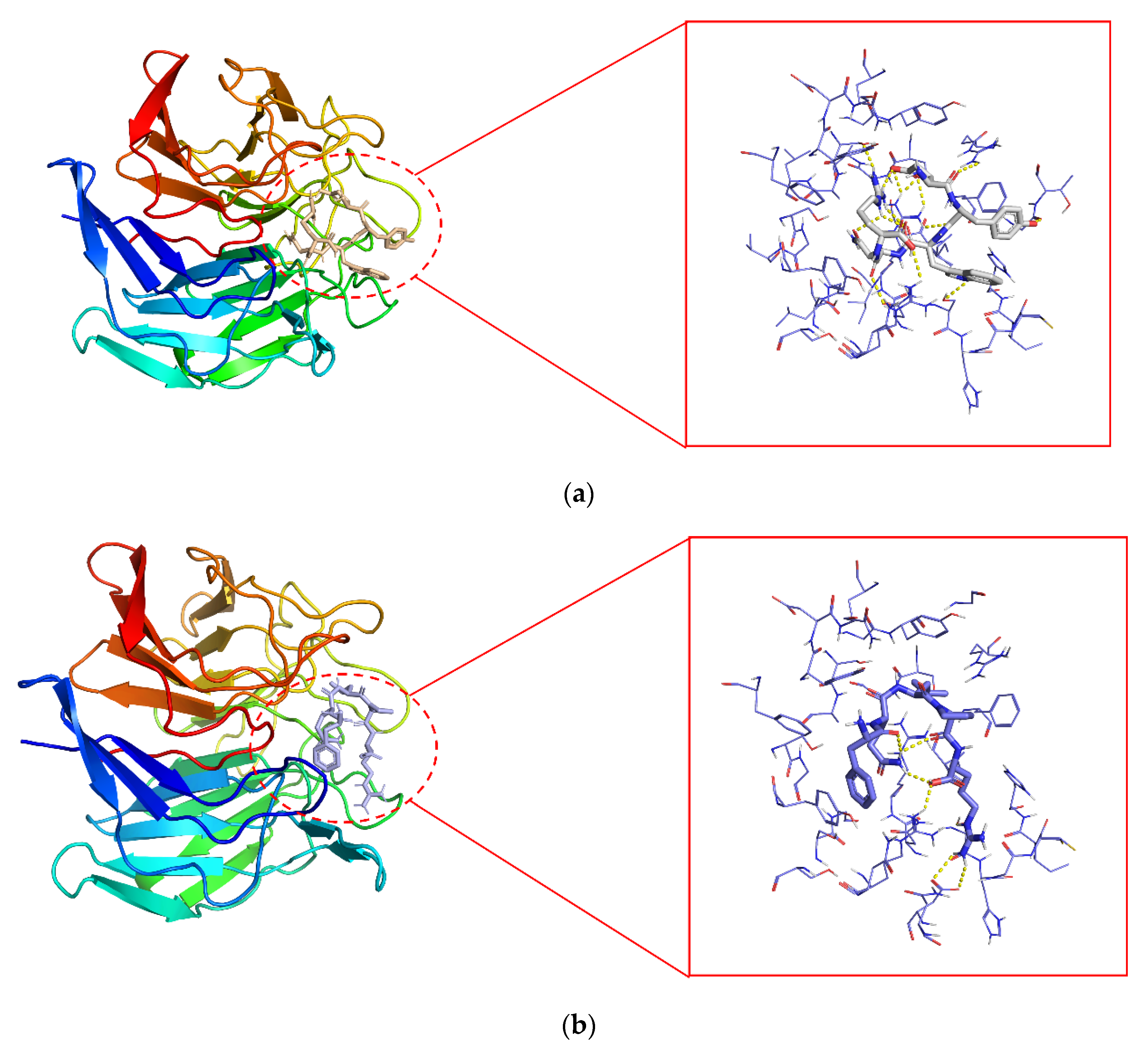{kind=link}
{kind=link}
| Sequence | Length | Isoelectric Point | Net Charge | GRAVY |
|---|---|---|---|---|
| VFDDELR | 7 | 4.03 | −2 | −0.6 |
| RVVDSEGGR | 9 | 6.07 | 0 | −1.022 |
| TELLR | 5 | 5.66 | 0 | −0.22 |
| LLSSSR | 6 | 9.75 | 1 | 0.117 |
| VDHLR | 5 | 6.71 | 0 | −0.64 |
| PDTLAR | 6 | 6.27 | 0 | −0.783 |
| YVTLK | 5 | 8.59 | 1 | 0.42 |
| FGNVLTK | 7 | 8.75 | 1 | 0.329 |
| TVEDELR | 7 | 4.14 | −2 | −1.1 |
| ESWPGSR | 7 | 6.1 | 0 | −1.786 |
| VLRPR | 5 | 12 | 2 | −0.52 |
| EYWNR | 5 | 6.1 | 0 | −2.74 |
| LAGNPHQQ | 8 | 6.74 | 0 | −1.262 |
| VYDDELR | 7 | 4.03 | −2 | −1.186 |
| SDFVSR | 6 | 5.55 | 0 | −0.433 |
| AGVDLVR | 7 | 5.88 | 0 | 0.8 |
| LLPEER | 6 | 4.53 | −1 | −0.917 |
| VVAPEPR | 7 | 5.97 | 0 | −0.143 |
| SSTDRLR | 7 | 9.31 | −1 | −1.571 |
| LPDTLAR | 7 | 5.84 | 0 | −0.129 |
| VPGLLEKEP | 9 | 4.53 | −1 | −0.3 |
| LLSSSR | 6 | 9.75 | 1 | 0.117 |
| RTLEPTR | 7 | 9.6 | 1 | −1.671 |
| VFDGELR | 7 | 4.37 | −1 | −0.157 |
| SPDQSYLR | 8 | 5.55 | 0 | −1.525 |
| YAELKR | 6 | 8.59 | 1 | −1.267 |
| VVLLR | 5 | 9.72 | 1 | 2.3 |
| SNHANQLDR | 9 | 6.46 | 0 | −1.878 |
| KLPLLR | 6 | 11 | 2 | 0.233 |
| GVSEDKLQR | 9 | 6.07 | 0 | −1.344 |
| KVVVPK | 6 | 10 | 2 | 0.533 |
| LDVLR | 5 | 5.84 | 0 | 0.76 |
| VEDELR | 6 | 4.14 | −2 | −1.167 |
| KLPLLR | 6 | 11 | 2 | 0.233 |
| RFEEERQR | 8 | 6.23 | 0 | −3.087 |
| TESDVFR | 7 | 4.37 | −1 | −0.857 |
| SQRPDLQPR | 9 | 9.31 | 1 | −2.189 |
| NDSTHPF | 7 | 5.08 | −1 | −1.5 |
| LLPVLR | 6 | 9.75 | 1 | 1.583 |
| FQLPR | 5 | 9.75 | 1 | −0.6 |
| LTVEDELR | 8 | 4.14 | −2 | −0.487 |
| HLVGPDK | 7 | 6.74 | 0 | −0.657 |
| LPDTLAR | 7 | 5.84 | 0 | −0.129 |
| FDVGVK | 6 | 5.84 | 0 | 0.567 |
| YFHSQ | 5 | 6.74 | 0 | −1.2 |
| EVLRPR | 6 | 9.7 | 1 | −1.017 |
| VEDELRVL | 8 | 4.14 | −2 | 0.125 |
| LLAGGERP | 8 | 6 | 0 | −0.125 |
| VFDDELER | 8 | 3.92 | −3 | −0.963 |
| SHSVLYR | 7 | 8.49 | 1 | −0.371 |
| AVDNAYAR | 8 | 5.88 | 0 | −0.4 |
| YSNAPR | 6 | 8.75 | 1 | −1.65 |
| FADLTK | 6 | 5.84 | 0 | 0.05 |
| YVPTER | 6 | 6 | 0 | −1.233 |
| SAGQRPW | 7 | 9.47 | 1 | −1.414 |
| TFLAR | 5 | 9.41 | 1 | 0.64 |
| YVTLRK | 6 | 9.99 | 2 | −0.4 |
| HADLPGLK | 8 | 6.74 | 0 | −0.4 |
| VFDDDRLR | 8 | 4.43 | −1 | −1.087 |
| TLPVLR | 6 | 9.41 | 1 | 0.833 |
| LNTPR | 5 | 9.75 | 1 | −1.3 |
| SEDKLQR | 7 | 5.79 | 0 | −2.271 |
| LPLLR | 5 | 9.75 | 1 | 1.06 |
| VDLVR | 5 | 5.81 | 0 | 0.84 |
| SSGGPLSLR | 9 | 9.47 | 1 | −0.189 |
| EGVLLRK | 7 | 8.85 | 1 | −0.071 |
| TNSFQMSPR | 9 | 9.41 | 1 | −1.189 |
| VYLAR | 5 | 9.81 | 1 | 1.06 |
| SDVFSR | 6 | 5.55 | 0 | −0.433 |
| VSLPKP | 6 | 8.72 | 1 | 0.017 |
| YGNVLR | 6 | 8.75 | 1 | −0.283 |
| FDGELR | 6 | 4.37 | −1 | −0.883 |
| SNDLLR | 6 | 5.55 | 0 | −0.783 |
| TNRPQF | 6 | 9.41 | 1 | −1.833 |
| LDREERAR | 8 | 6.18 | 0 | −2.3 |
| LVHPQR | 6 | 9.76 | 1 | −0.8 |
| FLDLLK | 6 | 5.84 | 0 | 1.133 |
| MPLDVLR | 7 | 5.59 | 0 | 0.586 |
| YAELK | 5 | 6 | 0 | −0.62 |
| GVDLVKR | 7 | 8.75 | 1 | −0.014 |
| TESDVFSR | 8 | 4.37 | −1 | −0.85 |
| VDAHPLR | 7 | 6.71 | 0 | −0.429 |
| LTPVLR | 6 | 9.75 | 1 | 0.833 |
| SPWER | 5 | 5.72 | 0 | −2.26 |
| ASFQR | 5 | 9.79 | 1 | −0.84 |
| AFNFPAR | 7 | 9.79 | 1 | −0.057 |
| SLVRVE | 6 | 5.72 | 0 | 0.567 |
| VSPDLVR | 7 | 5.81 | 0 | 0.257 |
| TDEYGNVLR | 9 | 4.37 | −1 | −1.044 |
| TEQAGRLS | 8 | 5.66 | 0 | −0.975 |
| Fraction No. | Sequence | Area | Toxin | Estimated Solubility | PeptideRanker Score |
|---|---|---|---|---|---|
| P1 | ESWPGSR | 1.37 × 108 | Non-Toxin | Good | 0.614027 |
| P2 | EYWNR | 6.17 × 107 | Non-Toxin | Good | 0.516512 |
| P3 | SPDQSYLR | 3.77 × 107 | Non-Toxin | Good | 0.609955 |
| P4 | KLPLLR | 3.08 × 107 | Non-Toxin | Good | 0.548661 |
| P5 | NDSTHPF | 2.42 × 107 | Non-Toxin | Good | 0.549226 |
| P6 | FQLPR | 2.21 × 107 | Non-Toxin | Good | 0.812155 |
| Intermolecular Forces | Index | Residue | AA |
|---|---|---|---|
| Hydrophobic interaction | 1 | 415X | ARG |
| 2 | 525X | TYR | |
| Hydrogen bonds | 1 | 363X | SER |
| 2 | 380X | ARG | |
| 3 | 380X | ARG | |
| 4 | 415X | ARG | |
| 5 | 415X | ARG | |
| 6 | 431X | SER | |
| 7 | 431X | SER | |
| 8 | 480X | GLY | |
| 9 | 483X | ARG | |
| 10 | 555X | SER | |
| 11 | 555X | SER | |
| 12 | 602X | SER | |
| π-stacking | 1 | 478X | PHE |
| Salt bridge | 1 | 415X | ARG |
| Intermolecular Forces | Index | Residue | AA |
|---|---|---|---|
| Hydrophobic interaction | 1 | 334X | TYR |
| 2 | 525X | TYR | |
| 3 | 556X | ALA | |
| Hydrogen bonds | 1 | 380X | ARG |
| 2 | 380X | ARG | |
| 3 | 414X | ASN | |
| 4 | 431X | SER | |
| 5 | 431X | SER | |
| 6 | 433X | GLY | |
| 7 | 433X | GLY | |
| 8 | 530X | GLN | |
| 9 | 555X | SER | |
| 10 | 555X | SER | |
| 11 | 555X | SER | |
| π-stacking | 1 | 334X | TYR |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Shang, Y.; Li, S.; Chen, Z.; Xia, J.; Tian, Y.; Jia, Y.; Ma, A. Molecular Docking Revealed the Potential Anti-Oxidative Stress Mechanism of the Walnut Polypeptide on HT22 Cells. Foods 2023, 12, 1554. https://doi.org/10.3390/foods12071554
Zhang Z, Shang Y, Li S, Chen Z, Xia J, Tian Y, Jia Y, Ma A. Molecular Docking Revealed the Potential Anti-Oxidative Stress Mechanism of the Walnut Polypeptide on HT22 Cells. Foods. 2023; 12(7):1554. https://doi.org/10.3390/foods12071554
Chicago/Turabian StyleZhang, Zijie, Yuting Shang, Siting Li, Zhou Chen, Junxia Xia, Yiling Tian, Yingmin Jia, and Aijin Ma. 2023. "Molecular Docking Revealed the Potential Anti-Oxidative Stress Mechanism of the Walnut Polypeptide on HT22 Cells" Foods 12, no. 7: 1554. https://doi.org/10.3390/foods12071554