
Silver Carp (Hypophthalmichthys molitrix) Scales Collagen Peptides (SCPs): Preparation, Whitening Activity Screening and Characterization

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Fish Scale Pre-Treatment and Principal Component Determination
2.3. Optimization of Enzymatic Hydrolysis Process
2.3.1. Screening of Proteases
2.3.2. Single-Factor Experiments and Response Surface Optimization of Enzymatic Hydrolysis
2.4. Separation and Purification of SCPs
2.5. Determination of Peptide Yield
2.6. Amino Acid Composition of SCPs
2.7. Comprehensive Evaluation of SCP Whitening Activity
2.7.1. Determination of DPPH Radical Scavenging Ability
2.7.2. Determination of Hydroxyl Radical Scavenging Rate
2.7.3. Determination of Tyrosinase-Inhibiting Activity
2.7.4. Determination of Moisture Absorption Rate and Moisture Retention Rate
2.8. SCPs1 Structural Characterization and Molecular Weight Analysis
2.9. Identification of the SCPs1 Peptide Sequence
2.10. Data Analysis
3. Results and Discussion
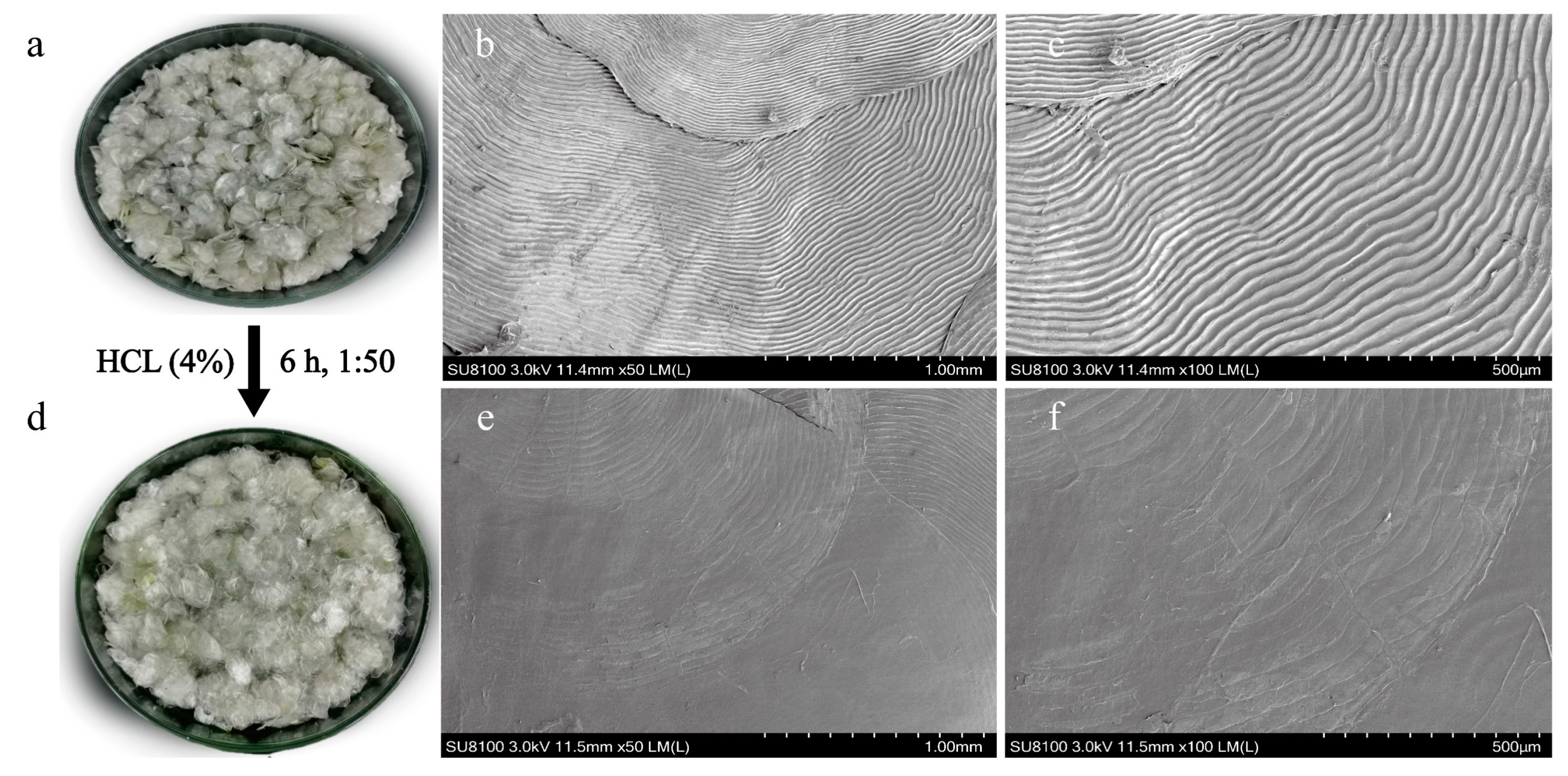
3.1. Basic Composition and Surface Microstructure of Fish Scales
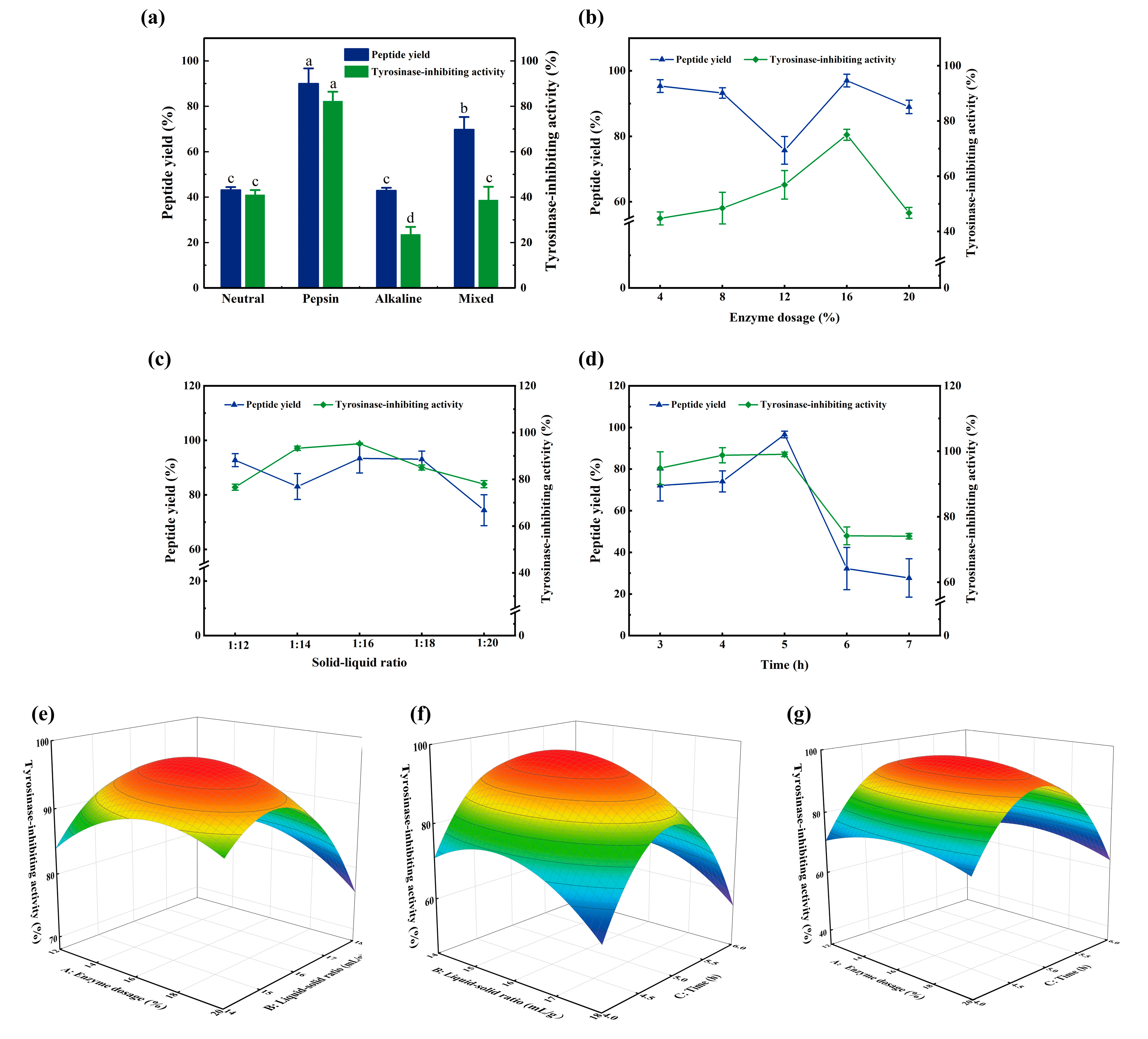
3.2. Optimization of the Enzymatic Hydrolysis Process
3.3. Amino Acid Composition of SCPs
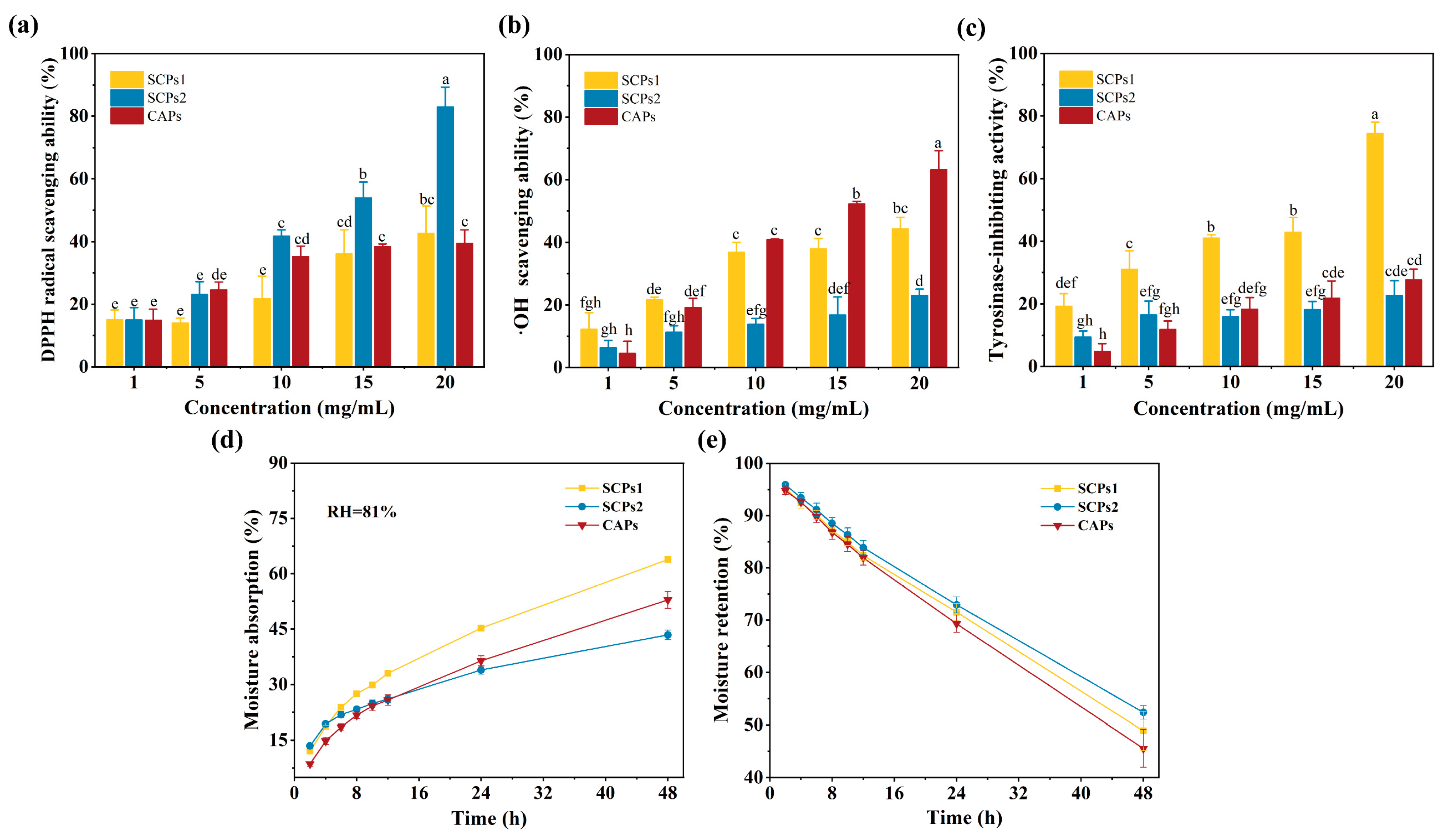
3.4. Whitening Activity of SCPs
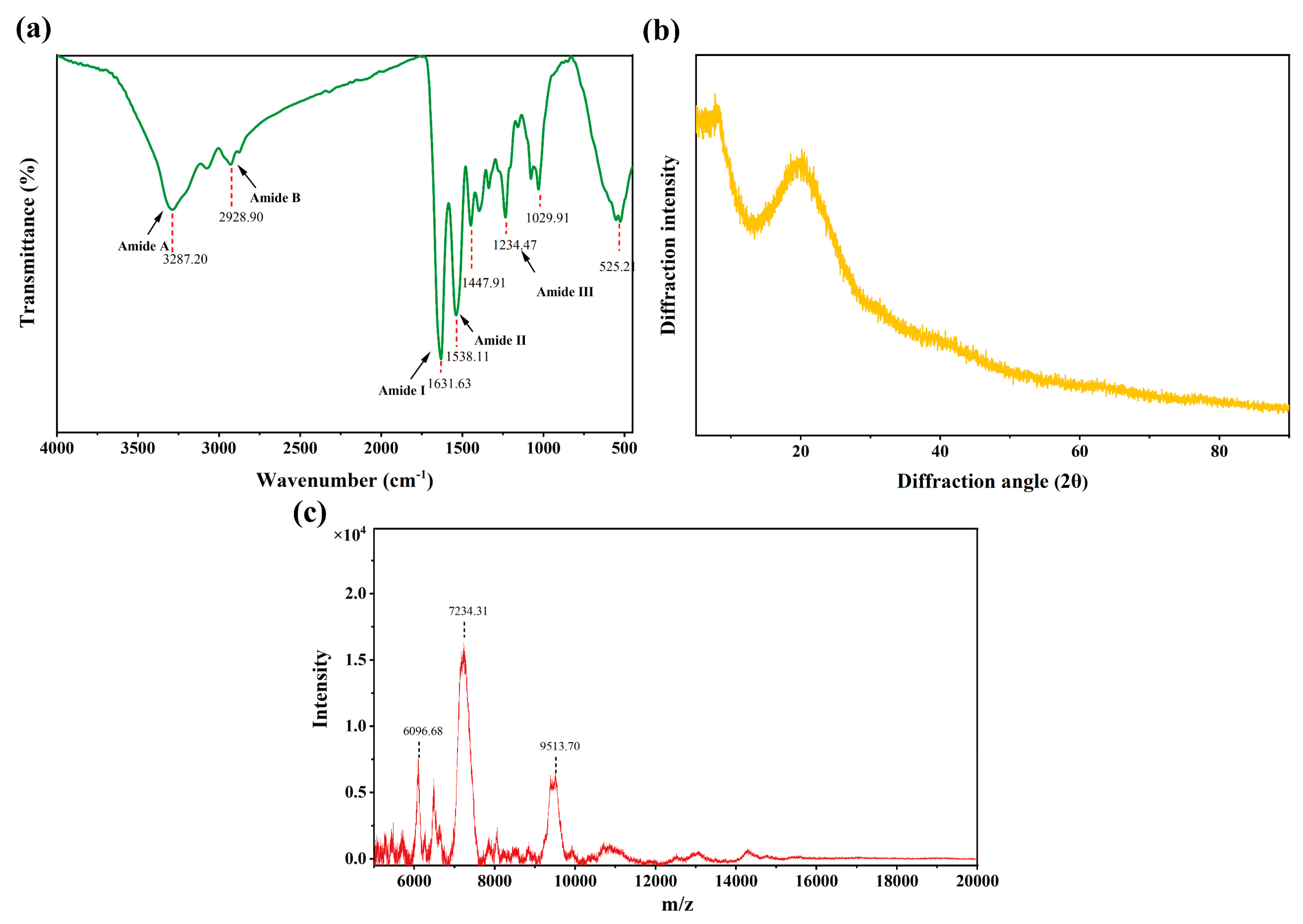
3.5. Structure and Molecular Weight Range of SCPs1
3.6. Active Peptide Sequence of SCPs1
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hong, H.; Zheng, Y.; Song, S.; Zhang, Y.; Zhang, C.; Liu, J.; Luo, Y. Identification and characterization of DPP-IV inhibitory peptides from silver carp swim bladder hydrolysates. Food Biosci. 2020, 38, 100748. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Q.; Hong, H.; Luo, Y. Prevention of protein oxidation and enhancement of gel properties of silver carp (Hypophthalmichthys molitrix) surimi by addition of protein hydrolysates derived from surimi processing by-products. Food Chem. 2020, 316, 126343. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Shan, Y.; Hong, H.; Luo, Y.; Hong, X.; Ye, W. Prevention of protein and lipid oxidation in freeze-thawed bighead carp (Hypophthalmichthys nobilis) fillets using silver carp (Hypophthalmichthys molitrix) fin hydrolysates. LWT-Food Sci. Technol. 2020, 123, 109050. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, T.; Zhang, Y.; Yang, L.; Nie, Y.; Tao, N.; Wang, X.; Zhong, J. Silver carp scale gelatins for the stabilization of fish oil-loaded emulsions. Int. J. Biol. Macromol. 2021, 186, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, X.; Yang, F.; Wu, J.; Huang, D.; Huang, J.; Wang, S. Effects and mechanism of antifreeze peptides from silver carp scales on the freeze-thaw stability of frozen surimi. Food Chem. 2022, 396, 133717. [Google Scholar] [CrossRef]
- Luo, X.; Liu, W.; Zhao, M.; Liu, T.; Xiong, F.; Lei, L.; Jia, F.; Feng, F. A novel Atlantic salmon (Salmo salar) bone collagen peptide delays osteoarthritis development by inhibiting cartilage matrix degradation and anti-inflammatory. Food Res. Int. 2022, 162, 112148. [Google Scholar] [CrossRef]
- Li, Y.; Fan, Y.; Liu, J.; Meng, Z.; Huang, A.; Xu, F.; Wang, X. Identification, characterization and in vitro activity of hypoglycemic peptides in whey hydrolysates from rubing cheese by-product. Food Res. Int. 2023, 164, 112382. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, X.; Liu, D. Collagen peptides and the related synthetic peptides: A review on improving skin health. J. Funct. Foods 2021, 86, 104680. [Google Scholar] [CrossRef]
- Jin, K.-S.; Lee, J.Y.; Hyun, S.K.; Kim, B.W.; Kwon, H.J. Juniperus chinensis and the functional compounds, cedrol and widdrol, ameliorate α-melanocyte stimulating hormone-induced melanin formation in B16F10 cells. Food Sci. Biotechnol. 2015, 24, 611–618. [Google Scholar] [CrossRef]
- Sierra, L.; Fan, H.; Zapata, J.; Wu, J. Antioxidant peptides derived from hydrolysates of red tilapia (Oreochromis sp.) scale. LWT-Food Sci. Technol. 2021, 146, 111631. [Google Scholar] [CrossRef]
- Zamorano-Apodaca, J.C.; Garcia-Sifuentes, C.O.; Carvajal-Millan, E.; Vallejo-Galland, B.; Scheuren-Acevedo, S.M.; Lugo-Sanchez, M.E. Biological and functional properties of peptide fractions obtained from collagen hydrolysate derived from mixed by-products of different fish species. Food Chem. 2020, 331, 127350. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Jiang, Y.; Wang, X.; Zhou, J.; Cui, H.; Xu, W.; He, Y.; Ma, H.; Gao, R. Effect of oral administration of collagen hydrolysates from Nile tilapia on the chronologically aged skin. J. Funct. Foods 2018, 44, 112–117. [Google Scholar] [CrossRef]
- Xu, D.; Zhao, M. Photoaging induced by long-term exposure to UV irradiation and amelioration by Skipjack tuna skin hydrolysates: Targeting inhibition of MAPK and NF-κB signaling hyperactivation. J. Funct. Foods 2022, 98, 105281. [Google Scholar] [CrossRef]
- Shi, H.; Zhang, M.; Yang, C. Effect of low-temperature vacuum frying assisted by microwave on the property of fish fillets (Aristichthys nobilis). J. Food Process Eng. 2019, 42, e13050. [Google Scholar] [CrossRef]
- Ahn, C.-B.; Je, J.-Y. Anti-adipogenic peptides from ark shell protein hydrolysate: Purification, identification and anti-adipogenic effect. Process Biochem. 2021, 109, 143–147. [Google Scholar] [CrossRef]
- Huang, J.-j.; Li, H.-l.; Xiong, G.-q.; Cai, J.; Liao, T.; Zu, X.-y. Extraction, identification and anti-photoaging activity evaluation of collagen peptides from silver carp (Hypophthalmichthys molitrix) skin. LWT-Food Sci. Technol. 2023, 173, 114384. [Google Scholar] [CrossRef]
- Yang, J.; Huang, J.; Zhu, Z.; Huang, M. Investigation of optimal conditions for production of antioxidant peptides from duck blood plasma: Response surface methodology. Poult. Sci. 2020, 99, 7159–7168. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, Y.-S.; Ahn, C.-B.; Je, J.-Y. Partial purification and identification of three antioxidant peptides with hepatoprotective effects from blue mussel (Mytilus edulis) hydrolysate by peptic hydrolysis. J. Funct. Foods 2016, 20, 88–95. [Google Scholar] [CrossRef]
- Chen, M.L.; Ning, P.; Jiao, Y.; Xu, Z.; Cheng, Y.H. Extraction of antioxidant peptides from rice dreg protein hydrolysate via an angling method. Food Chem. 2021, 337, 128069. [Google Scholar] [CrossRef]
- Song, Y.; Li, J.; Tian, H.; Xiang, H.; Chen, S.; Li, L.; Hu, X. Copper chelating peptides derived from tilapia (Oreochromis niloticus) skin as tyrosinase inhibitor: Biological evaluation, in silico investigation and in vivo effects. Food Res. Int. 2023, 163, 112307. [Google Scholar] [CrossRef]
- Wang, Z.; Cheng, S.; Wu, D.; Xu, Z.; Xu, S.; Chen, H.; Du, M. Hydrophobic peptides from oyster protein hydrolysates show better zinc-chelating ability. Food Biosci. 2021, 41, 110985. [Google Scholar] [CrossRef]
- Qu, W.; Feng, Y.; Xiong, T.; Li, Y.; Wahia, H.; Ma, H. Preparation of corn ACE inhibitory peptide-ferrous chelate by dual-frequency ultrasound and its structure and stability analyses. Ultrason Sonochem. 2022, 83, 105937. [Google Scholar] [CrossRef] [PubMed]
- Cotabarren, J.; Rosso, A.M.; Tellechea, M.; Garcia-Pardo, J.; Rivera, J.L.; Obregon, W.D.; Parisi, M.G. Adding value to the chia (Salvia hispanica L.) expeller: Production of bioactive peptides with antioxidant properties by enzymatic hydrolysis with Papain. Food Chem. 2019, 274, 848–856. [Google Scholar] [CrossRef] [PubMed]
- Mohd Pu’ad, N.A.S.; Abdul Haq, R.H.; Mohd Noh, H.; Abdullah, H.Z.; Idris, M.I.; Lee, T.C. Nano-size hydroxyapatite extracted from tilapia scale using alkaline heat treatment method. Mater. Today Proc. 2020, 29, 218–222. [Google Scholar] [CrossRef]
- Zhu, S.; Yuan, Q.; Yang, M.; You, J.; Yin, T.; Gu, Z.; Hu, Y.; Xiong, S. A quantitative comparable study on multi-hierarchy conformation of acid and pepsin-solubilized collagens from the skin of grass carp (Ctenopharyngodon idella). Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 96, 446–457. [Google Scholar] [CrossRef]
- Mohammadi, R.; Mohammadifar, M.A.; Mortazavian, A.M.; Rouhi, M.; Ghasemi, J.B.; Delshadian, Z. Extraction optimization of pepsin-soluble collagen from eggshell membrane by response surface methodology (RSM). Food Chem. 2016, 190, 186–193. [Google Scholar] [CrossRef]
- Sierra-Lopera, L.M.; Zapata-Montoya, J.E. Optimization of enzymatic hydrolysis of red tilapia scales (Oreochromis sp.) to obtain bioactive peptides. Biotechnol. Rep. 2021, 30, e00611. [Google Scholar] [CrossRef]
- Chen, X.; Jin, W.; Chen, D.; Dong, M.; Xin, X.; Li, C.; Xu, Z. Collagens made from giant salamander (Andrias davidianus) skin and their odorants. Food Chem. 2021, 361, 130061. [Google Scholar] [CrossRef]
- Cruz-Lopez, H.; Rodriguez-Morales, S.; Enriquez-Paredes, L.M.; Villarreal-Gomez, L.J.; Olivera-Castillo, L.; Cortes-Santiago, Y.; Lopez, L.M. Comparison of collagen characteristic from the skin and swim bladder of Gulf corvina (Cynoscion othonopterus). Tissue Cell 2021, 72, 101593. [Google Scholar] [CrossRef]
- He, L.; Gao, Y.; Wang, X.; Han, L.; Yu, Q.; Shi, H.; Song, R. Ultrasonication promotes extraction of antioxidant peptides from oxhide gelatin by modifying collagen molecule structure. Ultrason Sonochem. 2021, 78, 105738. [Google Scholar] [CrossRef]
- Hu, Z.-Z.; Ma, T.-X.; Sha, X.-M.; Zhang, L.; Tu, Z.-C. Improving tyrosinase inhibitory activity of grass carp fish scale gelatin hydrolysate by gastrointestinal digestion: Purification, identification and action mechanism. LWT-Food Sci. Technol. 2022, 159, 113205. [Google Scholar] [CrossRef]
- Sarikurkcu, C.; Sahinler, S.S.; Ceylan, O.; Tepe, B. Onosma pulchra: Phytochemical composition, antioxidant, skin-whitening and anti-diabetic activity. Ind. Crop. Prod. 2020, 154, 112632. [Google Scholar] [CrossRef]
- Lin, Y.-S.; Peng, H.-Y.; Zhu, Y.-X.; Meng, Y.-X.; Xiao, H.-H.; Chen, G.-Q. Evaluation of the extract of traditional Chinese medicine formula Si Ben Cao for skin whitening. Pharmacol. Res. 2022, 5, 100202. [Google Scholar] [CrossRef]
- Chi, C.F.; Hu, F.Y.; Wang, B.; Li, Z.R.; Luo, H.Y. Influence of Amino Acid Compositions and Peptide Profiles on Antioxidant Capacities of Two Protein Hydrolysates from Skipjack Tuna (Katsuwonus pelamis) Dark Muscle. Mar. Drugs 2015, 13, 2580–2601. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, A.; Tanaka, S.; Imai, Y.; Yoshida, H.; Kanaoka, T.; Tanaka, T.; Taniguchi, M. New tyrosinase inhibitory decapeptide: Molecular insights into the role of tyrosine residues. J. Biosci. Bioeng. 2016, 121, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Feissner, R.F.; Skalska, J.; Gaum, W.E.; Sheu, S.S. Crosstalk signaling between mitochondrial Ca2+ and ROS. Front. Biosci. 2009, 14, 1197–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Zhao, F.; Li, R.; Wu, Y.; Liu, S.; Liang, Q. Purification, characterization, antioxidant and moisture-preserving activities of polysaccharides from Rosa rugosa petals. Int. J. Biol. Macromol. 2019, 124, 938–945. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Yao, S.; Qiao, X.; Wang, Z.; Zhao, X.; Huang, Z.; Gu, Q.; Wang, N.; Peng, C. Evaluations of Marine Collagen Peptides from tilapia skin on experimental oral ulcer model of mice. Mater. Today Commun. 2021, 26, 101893. [Google Scholar] [CrossRef]
- Matmaroh, K.; Benjakul, S.; Prodpran, T.; Encarnacion, A.B.; Kishimura, H. Characteristics of acid soluble collagen and pepsin soluble collagen from scale of spotted golden goatfish (Parupeneus heptacanthus). Food Chem. 2011, 129, 1179–1186. [Google Scholar] [CrossRef]
- Liang, Q.; Wang, L.; Sun, W.; Wang, Z.; Xu, J.; Ma, H. Isolation and characterization of collagen from the cartilage of Amur sturgeon (Acipenser schrenckii). Process Biochem. 2014, 49, 318–323. [Google Scholar] [CrossRef]
- Tamilmozhi, S.; Veeruraj, A.; Arumugam, M. Isolation and characterization of acid and pepsin-solubilized collagen from the skin of sailfish (Istiophorus platypterus). Food Res. Int. 2013, 54, 1499–1505. [Google Scholar] [CrossRef]
- Guzzi Plepis, A.M.D.; Goissis, G.; Das-Gupta, D.K. Dielectric and pyroelectric characterization of anionic and native collagen. Polym. Eng. Sci. 1996, 36, 2932–2938. [Google Scholar] [CrossRef]
- Zhu, L.; Li, J.; Wang, Y.; Sun, X.; Li, B.; Poungchawanwong, S.; Hou, H. Structural feature and self-assembly properties of type II collagens from the cartilages of skate and sturgeon. Food Chem. 2020, 331, 127340. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, L.; Wu, S.; Chen, J.; Lin, H. Structural, functional, rheological, and biological properties of the swim bladder collagen extracted from grass carp (Ctenopharyngodon idella). LWT-Food Sci. Technol. 2022, 153, 112518. [Google Scholar] [CrossRef]
- Gao, X.; He, J.; Chen, J.; Zheng, Y.; Li, Y.; Ye, T. Double-spotted pufferfish (Takifugu bimaculatus) skin collagen: Preparation, structure, cytocompatibility, rheological, and functional properties. Arab. J. Chem. 2023, 16, 104402. [Google Scholar] [CrossRef]
- Chai, W.M.; Lin, M.Z.; Wang, Y.X.; Xu, K.L.; Huang, W.Y.; Pan, D.D.; Zou, Z.R.; Peng, Y.Y. Inhibition of tyrosinase by cherimoya pericarp proanthocyanidins: Structural characterization, inhibitory activity and mechanism. Food Res. Int. 2017, 100, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Schurink, M.; van Berkel, W.J.; Wichers, H.J.; Boeriu, C.G. Novel peptides with tyrosinase inhibitory activity. Peptides 2007, 28, 485–495. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level | Factor | ||
|---|---|---|---|
| X1 Enzyme Dosage/% | X2 Solid-Liquid Ratio/g/mL | X3 time/h | |
| −1 | 12 | 1:14 | 4 |
| 0 | 16 | 1:16 | 5 |
| 1 | 20 | 1:18 | 6 |
| Source | Sum of Square | Df | Means Square | F Value | Pr > F | Significant |
|---|---|---|---|---|---|---|
| Model | 3562.80 | 9 | 395.87 | 23.16 | 0.0002 | * |
| X1 | 0.43 | 1 | 0.43 | 0.025 | 0.8784 | |
| X2 | 111.31 | 1 | 111.31 | 6.51 | 0.0380 | * |
| X3 | 108.51 | 1 | 18.51 | 6.35 | 0.0398 | * |
| X1×2 | 26.47 | 1 | 26.47 | 1.55 | 0.2534 | |
| X1X3 | 1.96 | 1 | 1.96 | 0.11 | 0.7451 | |
| X2X3 | 3.99 | 1 | 3.99 | 0.23 | 0.6437 | |
| X12 | 98.85 | 1 | 98.85 | 5.78 | 0.0471 | * |
| X22 | 396.14 | 1 | 396.14 | 23.18 | 0.0019 | * |
| X32 | 2604.57 | 1 | 2604.57 | 152.40 | <0.0001 | ** |
| Residual | 119.64 | 7 | 17.09 | 5.77 | 0.0617 | |
| Lack of Fit | 97.19 | 3 | 32.40 | |||
| Pure Error | 22.44 | 4 | 5.61 | |||
| Total | 3682.43 | 16 | R2 = 0.9675 | R2Adj = 0.9257 |
| Amino Acid | Residues/ mg·g−1 | % | Amino Acid | Residues/ mg·g−1 | % |
|---|---|---|---|---|---|
| Threonine (Thr) | 23.73 | 3.27 | Leucine (Leu) * | 22.44 | 3.09 |
| Serine (Ser) | 28.78 | 3.97 | Tyrosine (Tyr) | 10.79 | 1.49 |
| Glutamic acid (Glu) | 78.78 | 10.86 | Lysine (Lys) # | 24.77 | 3.41 |
| Proline (Pro) * | 75.19 | 10.37 | Hlstidine (His) # | 11.64 | 1.60 |
| Glycine (Gly) | 143.35 | 19.76 | Argnine (Arg) # | 58.74 | 8.11 |
| Alanine (Ala) * | 68.21 | 9.40 | Isoleucine (Ile) * | 14.17 | 1.95 |
| Valine (Val) * | 18.52 | 2.55 | Phenylalanine (Phe) * | 18.32 | 2.5 |
| Methionine (Met) * | 16.41 | 2.26 | Aspartic acid (Asp) | 45.43 | 6.3 |
| Hydroxyproline (HyP) | 66.11 | 9.11 | Hydrophobic amino acid | 233.26 | 32.15 |
| Total | 725.37 | 100 |
| No. | Sequence | Length | Mass (Da) | Score | Protein Source |
|---|---|---|---|---|---|
| 1 | GDKGETGEAGERGMKGHRGF | 20 | 2074.96 | 268.5 | Collagen type I alpha 1 |
| 2 | TGPRGPVGPAGARGDKGETGEAGE | 24 | 2222.07 | 258.02 | Collagen type I alpha 1 |
| 3 | AGPPGADGQTGQRGE | 15 | 1396.63 | 254.19 | Collagen type I alpha 2 |
| 4 | YRGLEGNAGRDGA | 13 | 1334.63 | 248.69 | Collagen type I alpha 2 |
| 5 | YRGLEGNAGR | 10 | 1091.55 | 223.36 | Collagen type I alpha 2 |
| 6 | GARGDKGETGEAGERGMKGHRGF | 23 | 2359.12 | 218.69 | Collagen type I alpha 1 |
| 7 | SGPAGIAGPAGPRGPAGPNGPPGKD | 25 | 2151.08 | 218.13 | Collagen type I alpha 2 |
| 8 | KGETGEAGERGMKGHRGF | 18 | 1902.91 | 217.68 | Collagen type I alpha 1 |
| 9 | KRGSTGEQGSTGPLGMRGPRGAA | 23 | 2227.12 | 215.32 | Collagen type I alpha 2 |
| 10 | VGLTGPRGPVGPAGARGD | 18 | 1632.87 | 213.73 | Collagen type I alpha 1 |
| No. | Sequence | Mass (Da) | Score | Bioactivity Prediction |
|---|---|---|---|---|
| 1 | AGPPGADGQTGQRGE | 1396.63 | 254.19 | antioxidative, antiamnestic, DPP-IV, ACE |
| 2 | SGPAGIAGPAGPRGPAGPNGPPGKD | 2151.08 | 218.13 | antioxidative, antithrombotic, antiamnestic, α-glucosidase inhibitor |
| 3 | KRGSTGEQGSTGPLGMRGPRGAA | 2227.12 | 215.32 | antioxidative, hypotensive, DPP-IV, ACE |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zu, X.-Y.; Li, M.-J.; Xiong, G.-Q.; Cai, J.; Liao, T.; Li, H.-L. Silver Carp (Hypophthalmichthys molitrix) Scales Collagen Peptides (SCPs): Preparation, Whitening Activity Screening and Characterization. Foods 2023, 12, 1552. https://doi.org/10.3390/foods12071552
Zu X-Y, Li M-J, Xiong G-Q, Cai J, Liao T, Li H-L. Silver Carp (Hypophthalmichthys molitrix) Scales Collagen Peptides (SCPs): Preparation, Whitening Activity Screening and Characterization. Foods. 2023; 12(7):1552. https://doi.org/10.3390/foods12071552
Chicago/Turabian StyleZu, Xiao-Yan, Mei-Jin Li, Guang-Quan Xiong, Jun Cai, Tao Liao, and Hai-Lan Li. 2023. "Silver Carp (Hypophthalmichthys molitrix) Scales Collagen Peptides (SCPs): Preparation, Whitening Activity Screening and Characterization" Foods 12, no. 7: 1552. https://doi.org/10.3390/foods12071552