
High Genetic Diversity and Virulence Potential in Bacillus cereus sensu lato Isolated from Milk and Cheeses in Apulia Region, Southern Italy
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Detection and Isolation of B. cereus (s.l.)
2.3. Multi-Locus Sequence Typing (MLST)
2.4. Detection of Enterotoxin and Emetic Genes
2.5. Statistical Analysis
3. Results
3.1. Detection Rate of B. cereus s.l.
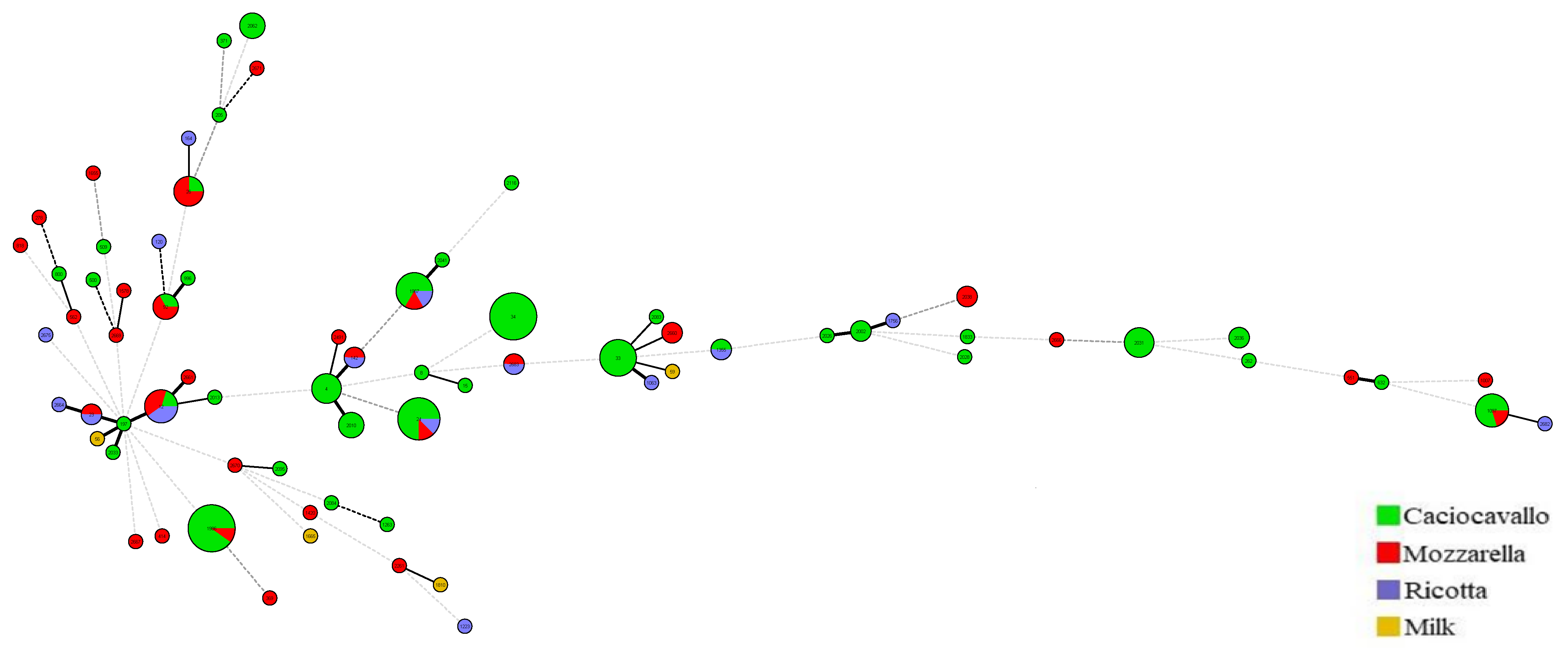
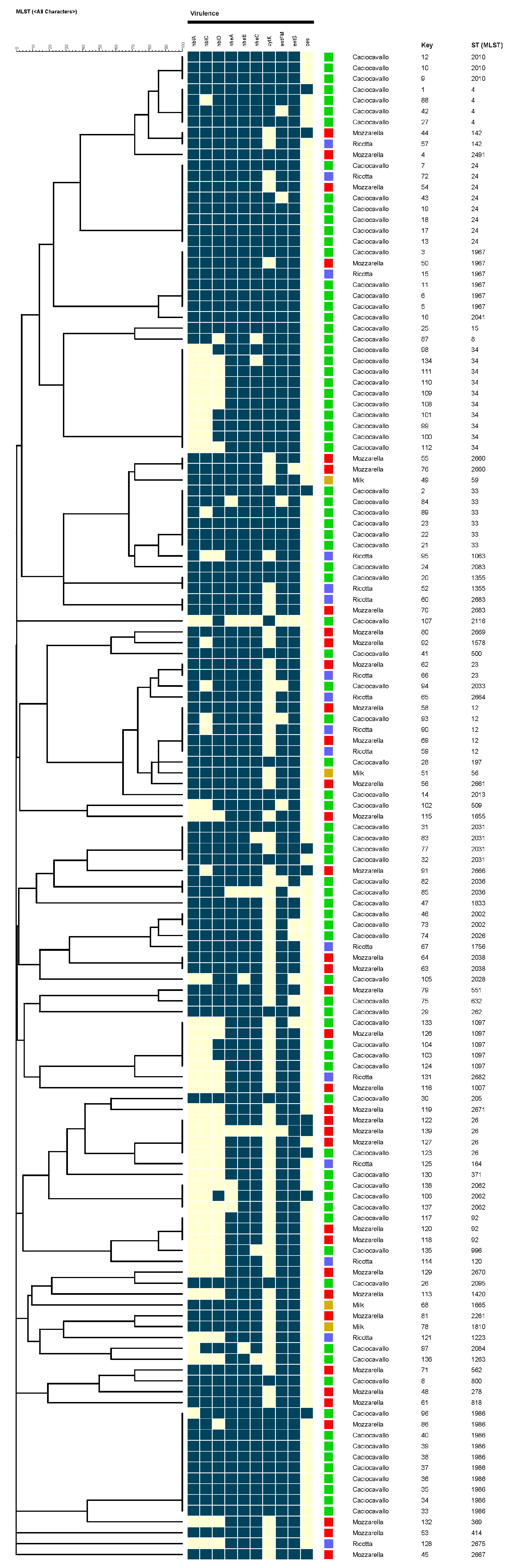
3.2. Strains Characterization by MLST
3.3. Distribution of Enterotoxin Genes
3.4. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Carroll, L.M.; Wiedmann, M.; Kovac, J. Proposal of a taxonomic nomenclature for the Bacillus cereus group which reconciles genomic definitions of bacterial species with clinical and industrial phenotypes. MBio 2020, 11, e00034-20. [Google Scholar] [CrossRef] [Green Version]
- Gao, T.; Ding, Y.; Wu, Q.; Wang, J.; Zhang, J.; Yu, S.; Yu, P.; Liu, C.; Kong, L.; Feng, Z.; et al. Prevalence, virulence genes, antimicrobial susceptibility, and genetic diversity of Bacillus cereus isolated from pasteurized milk in China. Front. Microbiol. 2018, 9, 533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owusu-Kwarteng, J.; Wuni, A.; Akabanda, F.; Tano-Debrah, K.; Jespersen, L. Prevalence, virulence factor genes and antibiotic resistance of Bacillus cereus sensu lato isolated from dairy farms and traditional dairy products. BMC Microbiol. 2017, 17, 65. [Google Scholar] [CrossRef] [Green Version]
- Rather, M.; Aulakh, R.S.; Gill, J.P.S.; Verma, R.; Rao, T.S. Enterotoxigenic profile of Bacillus cereus strains isolated from raw and pasteurized milk. Indian J. Anim. Sci. 2011, 81, 16–20. [Google Scholar]
- Dréan, P.; McAuley, C.M.; Moore, S.C.; Fegan, N.; Fox, E.M. Characterization of the spore-forming Bacillus cereus sensu lato group and Clostridium perfringens bacteria isolated from the Australian dairy farm environment. BMC Microbiol. 2015, 15, 38. [Google Scholar] [CrossRef] [Green Version]
- Saleh-Lakha, S.; Leon-Velarde, C.; Chen, S.; Lee, S.; Shannon, K.; Fabri, M.; Downing, G.; Keown, B. A study to assess the numbers and prevalence of Bacillus cereus and its toxins in pasteurized fluid milk. J. Food Prot. 2017, 80, 1085–1089. [Google Scholar] [CrossRef] [PubMed]
- Hammad, A.M.; Eltahan, A.; Khalifa, E.; Abbas, N.H.; Shimamoto, T. Toxigenic potential of Bacillus cereus strains isolated from retail dairy products in Egypt. Foodborne Pathog. Dis. 2021, 20, 20. [Google Scholar] [CrossRef]
- Fraccalvieri, R.; Bianco, A.; Difato, L.M.; Capozzi, L.; Del Sambro, L.; Simone, D.; Catanzariti, R.; Caruso, M.; Galante, D.; Normanno, G.; et al. Toxigenic genes, pathogenic potential and antimicrobial resistance of Bacillus cereus group isolated from ice cream and characterized by whole genome sequencing. Foods 2022, 11, 2480. [Google Scholar] [CrossRef]
- Flores-Urbán, K.A.; Natividad-Bonifacio, I.; Vázquez-Quiñones, C.R.; Vázquez-Salinas, C.; Quiñones-Ramírez, E.I. Detection of toxigenic Bacillus cereus strains isolated from vegetables in Mexico City. J. Food Prot. 2014, 77, 2144–2147. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Kim, H.S.; Kim, K.Y.; Chon, J.W.; Kim, D.H.; Seo, K.H. High occurrence rate and contamination level of Bacillus cereus in organic vegetables on sale in retail markets. Foodborne Pathog. Dis. 2016, 13, 656–660. [Google Scholar] [CrossRef]
- Yu, P.; Yu, S.; Wang, J.; Guo, H.; Zhang, Y.; Liao, X.; Zhang, J.; Wu, S.; Gu, Q.; Xue, L.; et al. Bacillus cereus isolated from vegetables in China: Incidence, genetic diversity, virulence genes, and antimicrobial resistance. Front. Microbiol. 2019, 10, 948. [Google Scholar] [CrossRef]
- Smith, D.P.; Berrang, M.E.; Feldner, P.W.; Phillips, R.W.; Meinersmann, R.J. Detection of Bacillus cereus on selected retail chicken products. J. Food Prot. 2004, 67, 1770–1773. [Google Scholar] [CrossRef] [PubMed]
- Choo, E.; Jang, S.S.; Kim, K.; Lee, K.G.; Heu, S.; Ryu, S. Prevalence and genetic diversity of Bacillus cereus in dried red pepper in Korea. J. Food Prot. 2007, 70, 917–922. [Google Scholar] [CrossRef]
- Wijnands, L.M.; Dufrenne, J.B.; Rombouts, F.M.; In’t Veld, P.H.; van Leusden, F.M. Prevalence of potentially pathogenic Bacillus cereus in food commodities in The Netherlands. J. Food Prot. 2006, 69, 2587–2594. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, M.; Yu, P.; Yu, S.; Wang, J.; Guo, H.; Zhang, J.; Zhou, H.; Chen, M.; Zeng, H.; et al. Prevalence, virulence feature, antibiotic resistance and MLST typing of Bacillus cereus isolated from retail aquatic products in China. Front. Microbiol. 2020, 11, 1513. [Google Scholar] [CrossRef] [PubMed]
- Te Giffel, M.C.; Beumer, R.R.; Granum, P.E.; Rombouts, F.M. Isolation and characterisation of Bacillus cereus from pasteurised milk in household refrigerators in The Netherlands. Int. J. Food Microbiol. 1997, 34, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Berthold-Pluta, A.; Pluta, A.; Garbowska, M.; Stefańska, I. Prevalence and toxicity characterization of Bacillus cereus in food products from Poland. Foods 2019, 8, 269. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Yu, P.; Wang, J.; Li, C.; Guo, H.; Liu, C.; Kong, L.; Yu, L.; Wu, S.; Lei, T.; et al. A study on prevalence and characterization of Bacillus cereus in ready-to-eat foods in China. Front. Microbiol. 2020, 10, 3043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeßberger, N.; Krey, V.M.; Rademacher, C.; Böhm, M.E.; Mohr, A.K.; Ehling-Schulz, M.; Scherer, S.; Märtlbauer, E. From genome to toxicity: A combinatory approach highlights the complexity of enterotoxin production in Bacillus cereus. Front. Microbiol. 2015, 6, 560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA. Risks for public health related to the presence of Bacillus cereus and other Bacillus spp. including Bacillus thuringiensis in foodstuffs. EFSA J. 2016, 14, e04524. [Google Scholar] [CrossRef]
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17, 276. [Google Scholar] [CrossRef] [Green Version]
- EFSA. The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, e06406. [Google Scholar] [CrossRef]
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, 324. [Google Scholar] [CrossRef]
- EFSA. The European Union One Health 2021 Zoonoses Report. EFSA J. 2022, 20, 273. [Google Scholar] [CrossRef]
- Drobniewski, F.A. Bacillus cereus and related species. Clin. Microbiol. Rev. 1993, 6, 324–338. [Google Scholar] [CrossRef]
- Ghelardi, E.; Celandroni, F.; Salvetti, S.; Barsotti, C.; Baggiani, A.; Senesi, S. Identification and characterization of toxigenic Bacillus cereus isolates responsible for two food-poisoning outbreaks. FEMS Microbiol. Lett. 2002, 208, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Bottone, E.J. Bacillus cereus, a volatile human pathogen. Clin. Microbiol. Rev. 2010, 23, 382–398. [Google Scholar] [CrossRef] [Green Version]
- Rishi, E.; Rishi, P.; Sengupta, S.; Jambulingam, M.; Madhavan, H.N.; Gopal, L.; Therese, K.L. Acute postoperative Bacillus cereus endophthalmitis mimicking toxic anterior segment syndrome. Ophthalmology 2013, 120, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Lund, T.; De Buyser, M.L.; Granum, P.E. A new cytotoxin from Bacillus cereus that may cause necrotic enteritis. Mol. Microbiol. 2000, 38, 254–261. [Google Scholar] [CrossRef]
- Pósfay-Barbe, K.M.; Schrenzel, J.; Frey, J.; Studer, R.; Korff, C.; Belli, D.C.; Parvex, P.; Rimensberger, P.C.; Schäppi, M.G. Food poisoning as a cause of acute liver failure. Pediatr. Infect. Dis. J. 2008, 27, 846–847. [Google Scholar] [CrossRef] [PubMed]
- Ehling-Schulz, M.; Guinebretiere, M.H.; Monthán, A.; Berge, O.; Fricker, M.; Svensson, B. Toxin gene profiling of enterotoxic and emetic Bacillus cereus. FEMS Microbiol. Lett. 2006, 260, 232–240. [Google Scholar] [CrossRef] [Green Version]
- Fagerlund, A.; Ween, O.; Lund, T.; Hardy, S.P.; Granum, P.E. Genetic and functional analysis of the cytK family of genes in Bacillus cereus. Microbiol.-SGM 2004, 150, 2689–2697. [Google Scholar] [CrossRef] [Green Version]
- Ehling-Schulz, M.; Svensson, B.; Guinebretiere, M.H.; Lindbäck, T.; Andersson, M.; Schulz, A.; Fricker, M.; Christiansson, A.; Granum, P.E.; Märtlbauer, E.; et al. Emetic toxin formation of Bacillus cereus is restricted to a single evolutionary lineage of closely related strains. Microbiology 2005, 151, 183–197. [Google Scholar] [CrossRef] [Green Version]
- Clavel, T.; Carlin, F.; Lairon, D.; Nguyen-The, C.; Schmitt, P. Survival of Bacillus cereus spores and vegetative cells in acid media simulating human stomach. J. Appl. Microbiol. 2004, 97, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Ceuppens, S.; Uyttendaele, M.; Drieskens, K.; Heyndrickx, M.; Rajkovic, A.; Boon, N.; Van de Wiele, T. Survival and germination of Bacillus cereus spores without outgrowth or enterotoxin production during in vitro simulation of gastrointestinal transit. Appl. Environ. Microbiol. 2012, 78, 7698–7705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajkovic, A.; Uyttendaele, M.; Vermeulen, A.; Andjelkovic, M.; Fitz-James, I.; In’t Veld, P.; Denon, Q.; Vérhe, R.; Debevere, J. Heat resistance of Bacillus cereus emetic toxin, cereulide. Lett. Appl. Microbiol. 2008, 46, 536–541. [Google Scholar] [CrossRef] [PubMed]
- ISTAT. 2020. Available online: http://dati.istat.it/Index.aspx?QueryId=25520 (accessed on 18 January 2023).
- ISO 21871:2006; Microbiology of Food and Animal Feeding Stuffs Horizontal Method for the Determination of Low Numbers of Presumptive Bacillus cereus Most Probable Number Technique and Detection Method. ISO: Geneva, Switzerland, 2006.
- PubMLST Database. Available online: https://pubmlst.org/bcereus/ (accessed on 18 January 2023).
- Guinebretière, M.-H.; Broussolle, V.; Nguyen-The, C. Enterotoxigenic profiles of food-poisoning and food-borne Bacillus cereus strains. J. Clin. Microbiol. 2002, 40, 3053–3056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asano, S.I.; Nukumizu, Y.; Bando, H.; Iizuka, T.; Yamamoto, T. Cloning of novel enterotoxin genes from Bacillus cereus and Bacillus thuringiensis. Appl. Environ. Microbiol. 1997, 63, 1054–1057. [Google Scholar] [CrossRef] [Green Version]
- Fricker, M.; Messelhäusser, U.; Busch, U.; Scherer, S.; Ehling-Schulz, M. Diagnostic real-time PCR assays for the detection of emetic Bacillus cereus strains in foods and recent food-borne outbreaks. Appl. Environ. Microbiol. 2007, 73, 1892–1898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehling-Schulz, M.; Vukov, N.; Schulz, A.; Shaheen, R.; Andersson, M.; Märtlbauer, E.; Scherer, S. Identification and partial characterization of the nonribosomal peptide synthetase gene responsible for cereulide production in emetic Bacillus cereus. Appl. Environ. Microbiol. 2005, 71, 105–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, B.M.; Hendriksen, N.B. Detection of enterotoxic Bacillus cereus and Bacillus thuringiensis strains by PCR analysis. Appl. Environ. Microbiol. 2001, 67, 185–189. [Google Scholar] [CrossRef] [Green Version]
- Andersson, A.; Ronner, U.; Granum, P.E. What problems does the food industry have with the spore-forming pathogens Bacillus cereus and Clostridium perfringens? Int. J. Food Microbiol. 1995, 28, 145–155. [Google Scholar] [CrossRef]
- Svensson, B.; Ekelund, K.; Ogura, H.; Christiansson, A. Characterisation of Bacillus cereus isolated from milk silo tanks at eight different dairy plants. Int. Dairy J. 2004, 14, 17–27. [Google Scholar] [CrossRef]
- Liu, X.Y.; Hu, Q.; Xu, F.; Ding, S.Y.; Zhu, K. Characterization of Bacillus cereus in dairy products in China. Toxins 2020, 12, 454. [Google Scholar] [CrossRef] [PubMed]
- Yobouet, B.; Kouamé-Sina, S.; Dadié, A.; Makita, K.; Grace, D.; Djè, K.; Bonfoh, B. Contamination of raw milk with Bacillus cereus from farm to retail in Abidjan, Côte d’Ivoire and possible health implications. Dairy Sci. Technol. 2013, 94, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Reis, A.L.S.; Montanhini, M.T.M.; Bittencourt, J.V.M.; Destro, M.T.; Bersot, L.S. Gene detection and toxin production evaluation of hemolysin BL of Bacillus cereus isolated from milk and dairy products marketed in Brazil. Braz. J. Microbiol. 2013, 44, 1195–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adame-Gómez, R.; Muñoz-Barrios, S.; Castro-Alarcón, N.; Leyva-Vázquez, M.A.; Toribio-Jiménez, J.; Ramírez-Peralta, A. Prevalence of the strains of Bacillus cereus group in artisanal Mexican cheese. Foodborne Pathog. Dis. 2020, 17, 8–14. [Google Scholar] [CrossRef]
- Acai, P.; Valík, Ľ.; Liptáková, D. Quantitative risk assessment of Bacillus cereus in pasteurised milk produced in the Slovak Republic. Czech. J. Food Sci. 2014, 32, 122–131. [Google Scholar] [CrossRef] [Green Version]
- Fei, P.; Yuan, X.; Zhao, S.; Yang, T.; Xiang, J.; Chen, X.; Zhou, L.; Ji, M. Prevalence and genetic diversity of Bacillus cereus isolated from raw milk and cattle farm environments. Curr. Microbiol. 2019, 76, 1355–1360. [Google Scholar] [CrossRef]
- Lücking, G.; Stoeckel, M.; Atamer, Z.; Hinrichs, J.; Ehling-Schulz, M. Characterization of aerobic spore-forming bacteria associated with industrial dairy processing environments and product spoilage. Int. J. Food Microbiol. 2013, 166, 270–279. [Google Scholar] [CrossRef]
- Tirloni, E.; Ghelardi, E.; Celandroni, F.; Bernardi, C.; Stella, S. Effect of dairy product environment on the growth of Bacillus cereus. J. Dairy Sci. 2017, 100, 7026–7034. [Google Scholar] [CrossRef] [PubMed]
- Carlin, F.; Fricker, M.; Pielaat, A.; Heisterkamp, S.; Shaheen, R.; Salonen, M.S.; Svensson, B.; Nguyen-The, C.; Ehling-Schulz, M. Emetic toxin-producing strains of Bacillus cereus show distinct characteristics within the Bacillus cereus group. Int. J. Food Microbiol. 2006, 109, 132–138. [Google Scholar] [CrossRef] [Green Version]
- Regione Puglia. Available online: https://www.patpuglia.it/ (accessed on 18 January 2023).
- Tirloni, E.; Stella, S.; Bernardi, C.; Sand Rosshaug, P. A new predictive model for the description of the growth of Salmonella spp. in Italian fresh ricotta cheese. LWT 2021, 143, 111163. [Google Scholar] [CrossRef]
- Lorusso, V.; Dambrosio, A.; Quaglia, N.C.; Parisi, A.; La Salandra, G.; Lucifora, G.; Mula, G.; Virgilio, S.; Carosielli, L.; Rella, A.; et al. Verocytotoxin-producing Escherichia coli O26 in raw water buffalo (Bubalus bubalis) milk products in Italy. J. Food Prot. 2009, 72, 1705–1708. [Google Scholar] [CrossRef]
- Finlay, W.J.; Logan, N.A.; Sutherland, A.D. Bacillus cereus produces most emetic toxin at lower temperatures. Lett. Appl. Microbiol. 2000, 31, 385–389. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Opinion of the Scientific Panel on biological hazards (BIOHAZ) on Bacillus cereus and other Bacillus spp. in foodstuffs. EFSA J. 2005, 3, 175. [Google Scholar] [CrossRef]
- Carlin, F.; Albagnac, C.; Rida, A.; Guinebretière, M.H.; Couvert, O.; Nguyen-The, C. Variation of cardinal growth parameters and growth limits according to phylogenetic affiliation in the Bacillus cereus group. Consequences for risk assessment. Food Microbiol. 2013, 33, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, R.J.; de Louvois, J.; Donovan, T.; Little, C.; Nye, K.; Ribeiro, C.D.; Richards, J.; Roberts, D.; Bolton, F.J. Guidelines for the microbiological quality of some ready-to-eat foods sampled at the point of sale. PHLS Advisory Committee for Food and Dairy Products. Commun. Dis. Public Health 2000, 3, 163–167. [Google Scholar] [PubMed]
- EU. COMMISSION REGULATION (EC) No 1441/2007 of 5 December 2007 amending Regulation (EC) No 2073/2005 on microbiological criteria for foodstuffs. Off. J. Eur. Union 2007, L 322/12. [Google Scholar]
- Cardazzo, B.; Negrisolo, E.; Carraro, L.; Alberghini, L.; Patarnello, T.; Giaccone, V. Multiple-locus sequence typing and analysis of toxin genes in Bacillus cereus food-borne isolates. Appl. Environ. Microbiol. 2008, 74, 850–860. [Google Scholar] [CrossRef] [Green Version]
- Didelot, X.; Barker, M.; Falush, D.; Priest, F.G. Evolution of pathogenicity in the Bacillus cereus group. Syst. Appl. Microbiol. 2009, 32, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yu, X.; Zhan, L.; Chen, J.; Zhang, Y.; Zhang, J.; Chen, H.; Zhang, Z.; Zhang, Y.; Lu, Y.; et al. Multilocus sequence type profiles of Bacillus cereus isolates from infant formula in China. Food Microbiol. 2017, 62, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Collier, F.A.; Elliot, S.L.; Ellis, R.J. Spatial variation in Bacillus thuringiensis/cereus populations within the phyllosphere of broad-leaved dock (Rumex obtusifolius) and surrounding habitats. FEMS Microbiol. Ecol. 2005, 54, 417–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helgason, E.; Caugant, D.A.; Lecadet, M.M.; Chen, Y.; Mahillon, J.; Lövgren, A.; Hegna, I.; Kvaløy, K.; Kolstø, A.B. Genetic diversity of Bacillus cereus/B. thuringiensis isolates from natural sources. Curr. Microbiol. 1998, 37, 80–87. [Google Scholar] [CrossRef]
- Priest, F.G.; Barker, M.; Baillie, L.W.; Holmes, E.C.; Maiden, M.C. Population structure and evolution of the Bacillus cereus group. J. Bacteriol. 2004, 186, 7959–7970. [Google Scholar] [CrossRef] [Green Version]
- Priest, F.G.; Goodfellow, M.; Todd, C. A numerical classification of the genus Bacillus. J. Gen. Microbiol. 1988, 134, 1847–1882. [Google Scholar] [CrossRef] [Green Version]
- Agata, N.; Ohta, M.; Yokoyama, K. Production of Bacillus cereus emetic toxin (cereulide) in various foods. Int. J. Food Microbiol. 2002, 73, 23–27. [Google Scholar] [CrossRef]
- Vassileva, M.; Torii, K.; Oshimoto, M.; Okamoto, A.; Agata, N.; Yamada, K.; Hasegawa, T.; Ohta, M. A new phylogenetic cluster of cereulide-producing Bacillus cereus strains. J. Clin. Microbiol. 2007, 45, 1274–1277. [Google Scholar] [CrossRef] [Green Version]
- Dierick, K.; Van Coillie, E.; Swiecicka, I.; Meyfroidt, G.; Devlieger, H.; Meulemans, A.; Hoedemaekers, G.; Fourie, L.; Heyndrickx, M.; Mahillon, J. Fatal family outbreak of Bacillus cereus-associated food poisoning. J. Clin. Microbiol. 2005, 43, 4277–4279. [Google Scholar] [CrossRef] [Green Version]
- Wetterhall, M.; Grönberg, A.; Grönlund, S.; Björkman, T.; Sandberg, L.; Musunuri, S.; Chaloupka, K.; Gammell, P. Removal of B. cereus cereulide toxin from monoclonal antibody bioprocess feed via two-step Protein A affinity and multimodal chromatography. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2019, 1118–1119, 194–202. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, Y.; Liu, X.; Xia, X.; Ding, S.; Zhu, K. Evaluation of the toxicity and toxicokinetics of cereulide from an emetic Bacillus cereus strain of milk origin. Toxins 2016, 8, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glasset, B.; Herbin, S.; Guillier, L.; Cadel-Six, S.; Vignaud, M.L.; Grout, J.; Pairaud, S.; Michel, V.; Hennekinne, J.A.; Ramarao, N.; et al. Bacillus cereus-induced food-borne outbreaks in France, 2007 to 2014: Epidemiology and genetic characterisation. Eurosurveillance 2016, 21, 30413. [Google Scholar] [CrossRef] [Green Version]
- Sánchez Chica, J.; Correa, M.M.; Aceves-Diez, A.E.; Rasschaert, G.; Heyndrickx, M.; Castañeda-Sandoval, L.M. Genomic and toxigenic heterogeneity of Bacillus cereus sensu lato isolated from ready-to-eat foods and powdered milk in day care centers in Colombia. Foodborne Pathog. Dis. 2020, 17, 340–347. [Google Scholar] [CrossRef] [Green Version]
- Castiaux, V.; Laloux, L.; Schneider, Y.J.; Mahillon, J. Screening of cytotoxic B. cereus on differentiated Caco-2 cells and in co-culture with mucus-secreting (HT29-MTX) Cells. Toxins 2016, 8, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biesta-Peters, E.G.; Dissel, S.; Reij, M.W.; Zwietering, M.H.; In’t Veld, P.H. Characterization and exposure assessment of emetic Bacillus cereus and cereulide production in food products on the Dutch market. J. Food Prot. 2016, 79, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Chica, J.; Correa, M.M.; Aceves-Diez, A.E.; Castañeda-Sandoval, L.M. Enterotoxin gene distribution and genotypes of Bacillus cereus sensu lato isolated from Cassava starch. Toxins 2021, 13, 131. [Google Scholar] [CrossRef] [PubMed]
- Økstad, O.A.; Kolstø, A.-B. Genomics of Bacillus Species. In Genomics of Foodborne Bacterial Pathogens; Wiedmann, M., Zhang, W., Eds.; Springer: New York, NY, USA, 2011; pp. 29–53. [Google Scholar] [CrossRef]
- Ellouze, M.; Buss Da Silva, N.; Rouzeau-Szynalski, K.; Coisne, L.; Cantergiani, F.; Baranyi, J. Modeling Bacillus cereus growth and cereulide formation in cereal-, dairy-, meat-, vegetable-based food and culture medium. Front. Microbiol. 2021, 12, 639546. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Target | Gene | Primer Sequence (5′-3′) | Reference |
|---|---|---|---|
| hblA | hblA_F | GTGCAGATGTTGATGCCGAT | [40] |
| HBLA_R | ATGCCACTGCGTGGACATAT | ||
| hblC | L2A_F | AATGGTCATCGGAACTCTAT | [40] |
| L2B_R | CTCGCTGTTCTGCTGTTAAT | ||
| hblD | L1A_F | AATCAAGAGCTGTCACGAAT | [40] |
| L1B_R | CACCAATTGACCATGCTAAT | ||
| nheA | nheA_F | TACGCTAAGGAGGGGCA | [40] |
| nheA_R | GTTTTTATTGCTTCATCGGCT | ||
| nheB | nheB_F | CTATCAGCACTTATGGCAG | [32] |
| nheB_R | ACTCCTAGCGGTGTTCC | ||
| nheC | nheC_F | CGGTAGTGATTGCTGGG | [32] |
| nheC_R | CAGCATTCGTACTTGCCAA | ||
| cytK | cytK_F | ACAGATATCGGKCAAAATGC | [26] |
| cytK_R | TCCAACCCAGTTWSCAGTTC | ||
| entFM | entFM_F | ATGAAAAAAGTAATTTGCAGG | [41] |
| entFM_R | TTAGTATGCTTTTGTGTAACC | ||
| entS | entS_F | GGTTTAGCAGCAGCTTCTGTAGCTGGCG | [41] |
| entFM_R | CTTGTCCAACTACTTGTAGCACTTGGCC | ||
| ces | ces_F | GGTGACACATTATCATATAAGGTG | [42] |
| ces_R | GTAAGCGAACCTGTCTGTAACAACA |
| Source | ST | CC | Genes Encoding for Enterotoxins | Cereulide Encoding Gene | Genetic Profile | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| hblA | hblC | hblD | nheA | nheB | nheC | cytK | entFM | entS | ces | ||||
| Caciocavallo | 4 | ST-142 complex | + | + | + | + | + | + | + | + | + | + | 1 |
| Caciocavallo | 33 | + | + | + | + | + | + | + | + | + | + | 1 | |
| Caciocavallo | 1967 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Mozzarella | 2491 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 1967 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 1967 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 24 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 800 | ST-111 complex | + | + | + | + | + | + | + | + | + | - | 2 |
| Caciocavallo | 2010 | ST-142 complex | + | + | + | + | + | + | + | + | + | - | 2 |
| Caciocavallo | 2010 | ST-142 complex | + | + | + | + | + | + | + | + | + | - | 2 |
| Caciocavallo | 1967 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 2010 | ST-142 complex | + | + | + | + | + | + | + | + | + | - | 2 |
| Caciocavallo | 24 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 2013 | ST-23 complex | + | + | + | + | + | + | + | + | + | - | 2 |
| Ricotta | 1967 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 2041 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 24 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 24 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 24 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 1355 | ST-18 complex | + | + | + | + | + | + | + | + | + | - | 2 |
| Caciocavallo | 33 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 33 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 33 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 2083 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 15 | ST-8 complex | + | + | + | + | + | + | + | + | + | - | 2 |
| Caciocavallo | 2095 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 4 | ST-142 complex | + | + | + | + | + | + | + | + | + | - | 2 |
| Caciocavallo | 197 | ST-23 complex | + | + | + | + | + | + | + | + | + | - | 2 |
| Caciocavallo | 262 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 205 | ST-205 complex | + | + | + | + | + | + | + | + | + | - | 2 |
| Caciocavallo | 2031 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 2031 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 1986 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 1986 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 1986 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 1986 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 1986 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 1986 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 1986 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 1986 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 500 | + | + | + | + | + | + | + | + | + | - | 2 | |
| Caciocavallo | 4 | ST-142 complex | + | + | + | + | + | + | + | - | + | - | 3 |
| Caciocavallo | 24 | + | + | + | + | + | + | + | - | + | - | 3 | |
| Mozzarella | 142 | + | + | + | + | + | + | - | + | + | + | 4 | |
| Mozzarella | 2667 | + | + | + | + | + | + | - | + | + | + | 4 | |
| Caciocavallo | 2002 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Caciocavallo | 1833 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Mozzarella | 278 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Milk | 59 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Mozzarella | 1967 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Milk | 56 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Ricotta | 1355 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Mozzarella | 414 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Mozzarella | 24 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Mozzarella | 2660 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Mozzarella | 2661 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Ricotta | 142 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Mozzarella | 12 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Ricotta | 12 | ST-23 complex | + | + | + | + | + | + | - | + | + | - | 5 |
| Ricotta | 2683 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Mozzarella | 818 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Mozzarella | 23 | ST-23 complex | + | + | + | + | + | + | - | + | + | - | 5 |
| Mozzarella | 2038 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Mozzarella | 2038 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Ricotta | 2664 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Ricotta | 23 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Ricotta | 1756 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Milk | 1665 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Mozzarella | 12 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Mozzarella | 2683 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Mozzarella | 562 | ST-111 complex | + | + | + | + | + | + | - | + | + | - | 5 |
| Ricotta | 24 | + | + | + | + | + | + | - | + | + | - | 5 | |
| Caciocavallo | 2002 | + | + | + | + | + | + | - | + | - | - | 6 | |
| Caciocavallo | 2026 | + | + | + | + | + | + | - | + | - | - | 6 | |
| Caciocavallo | 632 | + | + | + | + | + | + | - | + | - | - | 6 | |
| Mozzarella | 2660 | + | + | + | + | + | + | - | + | - | - | 6 | |
| Caciocavallo | 2031 | + | + | + | + | + | + | - | + | + | + | 7 | |
| Milk | 1810 | + | + | + | + | + | + | - | + | + | - | 8 | |
| Mozzarella | 551 | + | + | + | + | + | + | - | + | + | - | 8 | |
| Mozzarella | 2669 | + | + | + | + | + | + | - | + | + | - | 8 | |
| Mozzarella | 2261 | + | + | + | + | + | + | - | + | + | - | 8 | |
| Caciocavallo | 2036 | + | + | + | + | + | + | - | - | + | - | 9 | |
| Caciocavallo | 2031 | + | + | + | + | + | - | - | + | + | - | 10 | |
| Caciocavallo | 33 | + | + | + | - | + | + | + | - | + | - | 11 | |
| Caciocavallo | 2036 | + | + | + | - | - | - | - | + | - | - | 12 | |
| Mozzarella | 1986 | + | + | - | + | + | + | + | + | + | - | 13 | |
| Caciocavallo | 8 | ST-8 complex | + | + | - | + | + | - | + | + | + | - | 14 |
| Caciocavallo | 4 | ST-142 complex | + | - | + | + | + | + | + | + | + | - | 15 |
| Caciocavallo | 33 | + | - | + | + | + | + | + | + | + | - | 15 | |
| Ricotta | 12 | + | - | + | + | + | + | - | + | + | - | 16 | |
| Mozzarella | 2666 | + | - | + | + | + | + | - | + | + | + | 17 | |
| Mozzarella | 1578 | ST-97 complex | + | - | + | + | + | + | - | + | + | - | 18 |
| Caciocavallo | 12 | ST-23 complex | + | - | + | + | + | + | - | - | + | - | 19 |
| Caciocavallo | 2033 | ST-23 complex | + | - | + | + | + | + | - | - | + | - | 19 |
| Ricotta | 1063 | + | - | - | + | + | + | - | + | + | - | 20 | |
| Caciocavallo | 1986 | - | + | + | + | + | + | + | + | + | + | 21 | |
| Caciocavallo | 2084 | ST-365 complex | - | + | + | + | - | + | - | + | + | - | 22 |
| Caciocavallo | 34 | - | - | + | + | + | + | + | + | + | - | 23 | |
| Caciocavallo | 34 | - | - | + | + | + | + | + | + | + | - | 23 | |
| Caciocavallo | 34 | - | - | + | + | + | + | + | + | + | - | 23 | |
| Caciocavallo | 34 | - | - | + | + | + | + | + | + | + | - | 23 | |
| Caciocavallo | 509 | - | - | + | + | + | + | + | - | + | - | 24 | |
| Caciocavallo | 1097 | - | - | + | + | + | + | - | + | + | - | 25 | |
| Caciocavallo | 1097 | - | - | + | + | + | + | - | + | + | - | 25 | |
| Caciocavallo | 2028 | - | - | + | + | - | + | - | + | - | - | 26 | |
| Caciocavallo | 2062 | - | - | + | - | + | + | - | + | + | + | 27 | |
| Caciocavallo | 2116 | - | - | + | - | - | - | + | - | - | - | 28 | |
| Caciocavallo | 34 | - | - | - | + | + | + | + | + | + | - | 29 | |
| Caciocavallo | 34 | - | - | - | + | + | + | + | + | + | - | 29 | |
| Caciocavallo | 34 | - | - | - | + | + | + | + | + | + | - | 29 | |
| Caciocavallo | 34 | - | - | - | + | + | + | + | + | + | - | 29 | |
| Caciocavallo | 34 | - | - | - | + | + | + | + | + | + | - | 29 | |
| Mozzarella | 1420 | - | - | - | + | + | + | - | + | + | - | 30 | |
| Ricotta | 120 | - | - | - | + | + | + | - | + | + | - | 30 | |
| Mozzarella | 1655 | - | - | - | + | + | + | - | + | + | - | 30 | |
| Mozzarella | 1007 | - | - | - | + | + | + | - | + | + | - | 30 | |
| Caciocavallo | 92 | - | - | - | + | + | + | - | + | + | - | 30 | |
| Mozzarella | 92 | - | - | - | + | + | + | - | + | + | - | 30 | |
| Mozzarella | 2671 | - | - | - | + | + | + | - | + | + | - | 30 | |
| Mozzarella | 92 | - | - | - | + | + | + | - | + | + | - | 30 | |
| Ricotta | 1223 | - | - | - | + | + | + | - | + | + | - | 30 | |
| Mozzarella | 26 | - | - | - | + | + | + | - | + | + | + | 31 | |
| Caciocavallo | 26 | - | - | - | + | + | + | - | + | + | + | 31 | |
| Caciocavallo | 1097 | - | - | - | + | + | + | - | + | + | - | 32 | |
| Ricotta | 164 | - | - | - | + | + | + | - | + | + | - | 32 | |
| Mozzarella | 1097 | - | - | - | + | + | + | - | + | + | - | 32 | |
| Mozzarella | 26 | - | - | - | + | + | + | - | + | + | - | 32 | |
| Ricotta | 2675 | - | - | - | + | + | + | - | + | + | - | 32 | |
| Mozzarella | 2670 | - | - | - | + | + | + | - | + | + | - | 32 | |
| Caciocavallo | 371 | - | - | - | + | + | + | - | + | + | - | 32 | |
| Ricotta | 2682 | - | - | - | + | + | + | - | + | + | - | 32 | |
| Mozzarella | 369 | - | - | - | + | + | + | - | + | + | - | 33 | |
| Caciocavallo | 1097 | - | - | - | + | + | + | - | + | - | - | 34 | |
| Caciocavallo | 34 | - | - | - | + | + | - | + | + | + | - | 35 | |
| Caciocavallo | 996 | - | - | - | + | + | - | - | + | + | - | 36 | |
| Caciocavallo | 1263 | ST-365 complex | - | - | - | + | + | - | - | + | + | - | 36 |
| Caciocavallo | 2062 | - | - | - | - | + | + | - | + | + | - | 37 | |
| Caciocavallo | 2062 | - | - | - | - | + | + | - | + | + | - | 37 | |
| Mozzarella | 26 | - | - | - | - | - | - | - | - | + | + | 38 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bianco, A.; Normanno, G.; Capozzi, L.; Del Sambro, L.; Di Fato, L.; Miccolupo, A.; Di Taranto, P.; Caruso, M.; Petruzzi, F.; Ali, A.; et al. High Genetic Diversity and Virulence Potential in Bacillus cereus sensu lato Isolated from Milk and Cheeses in Apulia Region, Southern Italy. Foods 2023, 12, 1548. https://doi.org/10.3390/foods12071548
Bianco A, Normanno G, Capozzi L, Del Sambro L, Di Fato L, Miccolupo A, Di Taranto P, Caruso M, Petruzzi F, Ali A, et al. High Genetic Diversity and Virulence Potential in Bacillus cereus sensu lato Isolated from Milk and Cheeses in Apulia Region, Southern Italy. Foods. 2023; 12(7):1548. https://doi.org/10.3390/foods12071548
Chicago/Turabian StyleBianco, Angelica, Giovanni Normanno, Loredana Capozzi, Laura Del Sambro, Laura Di Fato, Angela Miccolupo, Pietro Di Taranto, Marta Caruso, Fiorenza Petruzzi, Ashraf Ali, and et al. 2023. "High Genetic Diversity and Virulence Potential in Bacillus cereus sensu lato Isolated from Milk and Cheeses in Apulia Region, Southern Italy" Foods 12, no. 7: 1548. https://doi.org/10.3390/foods12071548