
Protective Cultures in Food Products: From Science to Market

Department of Food, Nutrition and Facilities, FH Muenster, Correnstr. 25, 48149 Münster, Germany
*
Author to whom correspondence should be addressed.
Foods 2023, 12(7), 1541; https://doi.org/10.3390/foods12071541
Submission received: 1 March 2023
/
Revised: 1 April 2023
/
Accepted: 3 April 2023
/
Published: 5 April 2023
(This article belongs to the Special Issue Advance and Future Challenges to Microbial Food Safety)
Abstract
:An ultimate goal in food production is to guarantee food safety and security. Fermented food products benefit from the intrinsic capabilities of the applied starter cultures as they produce organic acids and bactericidal compounds such as hydrogen peroxide that hamper most food pathogens. In addition, highly potent small peptides, bacteriocins, are being expelled to exert antibiotic effects. Based on ongoing scientific efforts, there is a growing market of food products to which protective cultures are added exclusively for food safety and for prolonged shelf life. In this regard, most genera from the order Lactobacillales play a prominent role. Here, we give an overview on protective cultures in food products. We summarize the mode of actions of antibacterial mechanisms. We display the strategies for the isolation and characterization of protective cultures in order to have them market-ready. A survey of the growing market reveals promising perspectives. Finally, a comprehensive chapter discusses the current legislation issues concerning protective cultures, leading to the conclusion that the application of protective cultures is superior to the usage of defined bacteriocins regarding simplicity, economic costs, and thus usage in less-developed countries. We believe that further discovery of bacteria to be implemented in food preservation will significantly contribute to customer’s food safety and food security, badly needed to feed world’s growing population but also for food waste reduction in order to save substantial amounts of greenhouse gas emissions.
1. Introduction
Food safety and food security are of primary importance to fight the hunger of a reported 828 million people in 2021, so to deliver the required amounts of nutritive and highly valuable food products for everybody [1]. It has been estimated by the world health organization (WHO) and others that at least about 10% of the world’s population suffer each year from severe foodborne illnesses [2,3,4]. In addition to the affected diseased, more than 40% of food is lost along the food production chain from farm to fork, causing about 10% of total greenhouse gas emissions each year. The economic burden for food waste lost was given by an almost inconceivable 2.6 trillion USD including environmental and social costs [5,6]. One effective measure to reducing food waste and food infections would be the prolongation of the shelf life of perishable food products. How can this be achieved? In wealthy countries, the infrastructure for food transportation and food storage in companies and retail facilities, mainly done by refrigeration, assures on-time delivery of food to the customer. However, cooling and freezing of food is energy-consuming and generates unwanted emissions. In less-developed countries such food logistics are simply not affordable. Hence, there are good reasons for continuous extension of more simple and cost-effective alternatives to preserve food products.
Chemical preservation is one option, which has its limitations. Food preserved with sugar, salt, or organic acids changes texture and taste or, as in the case of sorbic acid, are not acceptable by the customer, because all-natural products that contain as few food additives as possible are en vogue [7,8]. Physical preservation requires equipment and energy for heating or cooling and has its limitations as sensory characteristics can sometimes undesirably alter the food product (i.e., high-pressure processing) [7]. Biological preservation can be achieved through food fermentation by microorganisms such as yeasts and lactic acid bacteria (LAB) [9]. This has led in human history to new, often very tasty food products, accompanied by prolonged preservation resulting in shelf-life extensions from hours or days to weeks or even months. Already about 2000 years ago, the availability of wine as a safe source for drinking supply together with the increased availability of fermented foods was a factor not to be underestimated for the Roman armies to build up the Roman empire [9,10]. When in the early 20th century Ilja Iljitsch Metchnikow described the activities of non-pathogenic, health-promoting LAB as the basis of fermented milk products such as kefir and yoghurt, it became obvious that microorganisms are the indispensable actors, the so-called starter cultures [11,12]. While fermentation generally influences the organoleptic properties of food products by secreting organic acids, ethanol, carbon dioxide, and numerous aromatic compounds, they produce concomitantly small peptides, the bacteriocins, and other bactericidal compounds such as diacetyl. The observation that species from the order Lactobacillales can be included in food products as protective cultures to fight pathogenic and food spoiling bacteria without interfering with food taste and texture paved the way for the field of bio-protection [13]. Hence, the use of protective cultures in perishable foods such as smoked salmon, minced meat, salad, and sprouts has resulted in a significant increase in shelf life and food safety.
In this review, we address the application of LAB as food protective cultures. We describe the characteristic features of protective microorganisms regarding the underlying mechanisms. We outline the strategies for strain identification and validation relating to application in diverse food products and provide a survey on the market. For the first time, we extensively attribute the legal restrictions for the use of protective cultures to provide a guideline to the interested user.
2. Antibacterial Properties
The concept of bio-protection of food by the implementation of protective cultures is widely accepted, is substantiated in numerous studies, and is implemented into a wide range of products on the market [13,14,15]. As a first precondition, the activity of a protection culture in a certain food environment must leave the organoleptic properties untouched. However, there is more to consider: (i) production of antimicrobial substances should be effective; (ii) survival during the manufacturing process must be guaranteed; (iii) bacteria must be able to function at cooling temperatures; and (iv) they should be proofed as safe regarding the absence of pathogenic genes and genes encoding resistances to antibiotics. How do they exert these properties?
The addition of live bacteria to food products as protective cultures was based on the observation that bacteria have developed a number of strategies to outplace competing species. An initial observation was that a bacterial species can overgrow in co-culture experiments other species simply by the higher cell amount of the inoculum, depriving nutrients and minerals. The so-called Jameson-Effect proved to be valid in many cases [16,17,18,19,20]. Nevertheless, when alternative nutrients are available, it does not always work [21].
Microorganisms have evolved mechanisms to survive in their ecosystems. One example is the capacity of soil-dwelling Streptomycetes that secret bio-active secondary metabolites as a survival strategy against other microorganisms, a process that is triggered by carbon regulation [22,23,24]. Among these metabolites are more than two-thirds of medically used antibiotics [25]. Members of the order Lactobacillales exert as well a multitude of measures against other microorganisms (Figure 1). Through homo- and heterofermentative fermentation of carbohydrates, they produce a variety of organic acids which they secrete as the metabolic end products [26]. While lactate and acetate are quite common, there are LAB that make propionic acid, malic acid, succinic acid, butyric acid, or formic acid [14,27].
Organic acids are entering the cell in their uncharged form where they dissociate and lower the cell internal pH [28,29]. Most bacteria keep and regulate internal pH between six and seven as long as the external pH is in the range of five to eight [29]. Further acidification disturbs the cell internal ionic balance through the disruption of cell membranes, enzyme denaturation and damage of nucleic acids. Notably, the proton gradient across the membrane collapses which inhibits ATP Synthase, proton symporters, and the proton-driven flagellar motor (Figure 1) [27,30].
Hydrogen peroxide, carbon dioxide, and diacetyl are widely produced antimicrobial agents. LAB synthesize hydrogen peroxide in the presence of oxygen and light to amounts up to 1 mM [31,32]. Among 193 examined LAB species, 37 were able to secret hydrogen peroxide [31,33]. Bacteria that do not possess enzymes for breakdown such as peroxidases or catalases are posed under stress through this oxidative compound that target thiol groups within enzymes and exert oxidative cell reactions such as uncoupling the electron transfer chain [31,34]. Escherichia coli and Salmonella enteritidis strains that possess catalase are inhibited as well, since in this case the overall enzyme activity is often not sufficient for detoxification [31,33].
Decent amounts of the butter flavor diacetyl, formed by LAB, are present in many food products [35]. Diacetyl has been shown to inhibit food pathogenic Gram-negative bacteria, molds, and yeasts in the range of 50 to 1000 ppm [35] but has been shown to be less effective against Gram-positive bacteria [36]. It has been identified as a mutagenic agent by the Ames-Test [37,38] and attributed to the inhibition of arginine utilization [35]. It should be noted that diacetyl could be toxic for humans through inhalation of this volatile compound (Figure 1) [39].
Lactic acid bacteria that produce carbon dioxide have been shown to inhibit the growth of Gram-positive bacteria and Gram-negative psychrotrophs, including Enterobacteriaceae and Listeria monocytogenes [40]. Carbon dioxide can alter bacterial communities, reduce metabolite production, and as an example improve in this way the quality of fermented kimchi [40,41]. The mechanisms comprise the creation of an anaerobic atmosphere, the decrease of pH accompanied with enzyme denaturation, and the loss of the proton gradient and cell membrane [42]. Secreted aldehydes also confer antimicrobial function. The best studied one is reuterin (3-hydroxy propionaldehyde) from Lactobacillus reuteri strains. It causes oxidative stress and interacts with thiol groups [43]. All groups of microorganisms including spores and protozoa are targeted by reuterin [44].
Of central importance is the production of numerous bio-protective peptides, the bacteriocins (Figure 1) [45,46]. The class I contains peptides with less than 10 kDa that are posttranslational modified and thus have unusual amino acids [47]. In class II, non-modified small bacteriocins are classified. They are independent from modifying cell enzymes. Peptides bigger than 10 kDa, all unmodified, are in class III. By bacteriocin-mediated pore formation, the cell wall is targeted, and damage occurs in the peptidoglycan layer [48]. Bacteriocins bind to cell envelope structures like Lipid II or to the PEP:dependent mannose phosphotransferase transport system [49]. Some bacteriocins act on the central cell components DNA, RNA, and protein metabolism (Figure 1) [45,46]. Diverse bacteriocins can exhibit a very narrow spectrum against a few species or a broad spectrum against bacteria. Some of the most notable ones are nisin, produced by Lactococcus lactis, pediocin, synthesized by Pediococcus strains, or zoocin A, a product from Streptococcus equi subsp. Zooepidemicus [50]. The screening for genes and operons encoding bacteriocins has been facilitated by the BAGEL4 database server [51]. In 2018, there have been 820 gene loci detected in 238 genomes, best studied in the genera Lactobacillus and Streptococcus [47].
Although much knowledge has been acquired through the past decades on the above-mentioned antibacterial compounds and bacteriocins, there are still research gaps to fill in that require further research efforts (Figure 1). The molecular mechanisms of the numerous antibacterial actions are still to some extent a mystery and thus await discovery. In addition, the invention of high throughput technologies to directly identify novel bacteriocins is still a drawback. However, the recent launching of a micro-bioreactor could help solving this, since the device allows parallel fermentation of 48 cultures in 0.8 mL to 2.4 mL [52].
Regarding application, there are many studies demonstrating that protective cultures can be applied in various food to enhance food safety [53,54,55,56,57,58,59,60,61,62,63,64]. A few selected studies in various food groups are given here. Meat products made of cooked ham and minced meat are most sensible towards spoilage and contamination with pathogenic bacteria. Cooked cubed ham was inoculated with a mixture of L. monocytogenes strains and was challenged with two commercial protective cultures, Lyocarni BOX-74 and Lyocarni BOX-57. Storage for 40 days at a household-representative refrigerator temperature of 8 °C revealed that Listeria have been eliminated without organoleptic changes from the ham, making this setting a proof-of-concept for cooked ham products [53]. Stephane Chaillou and co-workers applied L. sakei strains to ground beef stored at 4 °C and 8 °C, respectively. A protective effect could be demonstrated against Salmonella enterica Typhimurium and Escherichia coli O157:H7 [54]. Among marine products, smoked salmon is of particular safety concern due to contamination with L. monocytogenes and also due to a number of strong-smelling spoilage bacteria like Photobacterium phosphoreum, Brochothrix thermosphacta, and Serratia proteamaculans. It was shown in several publications that appropriate LAB could stabilize such products [55,56]. Fresh cheese has been studied by the isolation and investigation of eight LAB strains. A mixture of two strains demonstrated great potential as a protective culture for the cheese making process against listerial contamination [57]. Fruits and vegetables can be contaminated predominantly with E. coli EHEC ssp., Salmonella enterica ssp., or L. monocytogenes. Since such food is consumed raw, numerous outbreaks have happened in past decades [58,59]. The functionality of the prevention of pathogens has been shown for cut fruits, pre-cut cantaloupes, and papayas as well as in iceberg salad, cucumber, or romaine lettuce [58,60]. A quite novel approach for application is the improvement of food safety in catering by protective cultures, especially concerning vulnerable groups. By spraying protective LAB on certain dishes before serving, the risk of food infections was significantly reduced [61]. The above-mentioned selection of thorough investigations shows the potential for food produced with protective cultures to improve food safety. We will further address this on the perspectives of the market.
3. Isolation and Validation of protective LAB and Bacteriocins
LAB are ubiquitous present in the environment and thus can be isolated from a wide variety of food-associated habitats such as milk from buffalo, cow, and goat and their respective farm environments. A good source are wild fermented food products like cheese, sausages, or wine. These are made by taking advantage of the autochthonic flora. Other suitable sources are the gastrointestinal tracts of fishes or insects. Even from honeydew and pig feces, the isolation of potential new protective cultures or possible bacteriocin producers has been demonstrated [65,66,67,68,69,70]. Samples are plated onto De Man, Rogosa, Sharpe Agar (MRS), or Brain Heart Infusion Agar (BHI), sometimes supplemented with growth-stimulating additives such as cysteine, fructose, or bromphenol blue (BPB) [67,70]. New species can be readily distinguished by colony morphology in shape, size, and color. In our hands the best choice is MRS-BPB agar as a chromogenic differential medium that delivers all kind of blueish colonies (our unpublished results; Figure 2A [71,72,73].
If new strains are available as pure cultures, species identification is conventionally performed by 16S-rRNA sequencing [67,69,70,74]. However, due to the conserved behavior of the 16SrRNA gene, unambiguous identification at the species level will not always be possible. Instead, PCR amplification with primer pairs corresponding to less-conserved genes will be the solution. It should be noted that only species can be identified for which the genome has been sequenced [75,76]. Alternatively, species identification can be achieved by comparison of the in silico-generated proteolytic protein fragment pattern using matrix-assisted laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF MS). Reliable identification with this rapid detection method is particularly dependent whether the respective species has been sequenced and deposited in the database [66,70].
In a next step, the isolate is characterized in terms of antimicrobial properties and biological safety. This is addressed by (i) in vivo detection of antibacterial activity against indicator strains or (ii) by genome sequencing followed by bioinformatic searches for the presence of bacteriocin-encoding genes. For detection in vivo, the point inoculation assay (Figure 2B), the spot-on-lawn assay, or the agar-well diffusion assay is available [69,74,77,78,79,80]. These assays can also be used to clarify a sporocidal or sporostatic effect [65,81]. Since a response against an indicator organism does not necessarily have to be based solely on the action of bacteriocins, possible effects of acidification or hydrogen peroxide should be clarified first. The production of the latter can be analyzed with a 3,3’,5,5’-tetramethyl-benzidine (TMB) and catalase-containing MRS agar, in which a color change of the colony from white to dark blue occurs if hydrogen peroxide is produced (Figure 2C). In this case, the produced amounts should be quantitatively determined [82,83]. Secretion of acid can be measured with pH indicator stripes placed on the agar plate as shown in Figure 2D. The tolerance of the isolate to low pH environments (acidification) can be determined in a survival challenge test by incubation at a low pH-range between two to five [66].
In order to meet the required criteria for food safety, it is necessary to show the absence of antibiotic resistant genes and genes encoding virulence factors [66,84,85]. It is recommended to substantiate the analyses in vivo by antibiotic susceptibility testing [84]. A convenient and reliable strategy is whole genome sequencing, which is cost- and time-efficient since the invention of nanopore sequencing technology, followed by bioinformatics [51,86,87,88,89,90,91]. A versatile tool is the BAGEL4 webserver that has been established within the group of Jan Kok and Oscar Kuipers at the Groningen Biomolecular Sciences and Biotechnology Institute (GBB), Netherlands [51]. The database comprises 820 genes encoding bacteriocins [51].
While the examination of the safety criteria is ongoing, experiments could be conducted in parallel to implement the new isolated protection strain into the chosen food product. The microbiological profile should be followed to determine the shelf-life. Absence or prevention against potential food pathogens, texture analysis, and proof of unchanged organoleptic properties will complete characterization. Subsequently, the upscaling process must follow in order to commercially sell the food product in sufficient quantities.
4. Industrial Production of Protective Cultures, Bacteriocins, and Their Actual Imitations
The industrial production of protective cultures in a bioreactor follows those for LAB used as food starter cultures. The protective cultures are cultivated from a stock culture using small-volume batches of about 2 mL to 100 mL or a fed-batch process to grow the bacteria until a desired biomass concentration is reached. The biomass serves to inoculate 200 L to 50,000 L bioreactors in a batch or fed-batch process [92]. At this stage, bioreactors no longer work with defined media, but instead with cheap raw materials such as sidestream products from cereals or sugar beets to minimize costs and maximize yield (Table 1). Decisive for a high biomass during fermentation is keeping the culture in the exponential growth phase and within the appropriate environmental parameters. They include oxygen content, redox potential, stirring speed, pH, and temperature. Although LAB are acid tolerant, self-acidification primarily by lactic acid has a more negative influence on biomass formation than limitation of nutrients. Controlling pH at a value close to neutral is critical for ensuring highest growth rates and a maximum of biomass [93].
A compromise between protective culture as maximum biomass and pure bacteriocin production are fermentates. Fermentates are the bacterial cells propagated in the reactor plus medium into which bacteriocins are secreted. Under optimally adapted physical and chemical conditions in vitro, LAB are able to secrete bacteriocins into significantly higher amounts than in vivo in a food matrix [98]. However, many factors influence the secretion of large amounts of bacteriocin into the medium, for example pH [99,100], temperature [100], nutrient density, and salt content. The higher the salinity the lower the yield of the respective bacteriocin [101]. On the other hand, salt stress may also be necessary to secrete bacteriocins [102]. This poses complex challenges between biomass and bacteriocin production. Especially, the focus on bacteriocin production should result in a yield of a minimum of 50% and a purity of 90% [103]. In most cases, the yield is significantly lower and is then associated with large efforts to improve culture conditions and purification process [104,105].
An alternative approach for bacteriocin production is the molecular biology approach in which bacteriocin-encoding genes or operons are cloned into strains that are widely used in biotechnology. Since the gene sequences of numerous bacteriocins are readily available [68,106], the successful construction of recombinant bacteriocin-producing Escherichia coli and Corynebacterium glutamicum has been demonstrated [107,108,109,110,111].
5. The Legal Use of Protection Cultures and Bacteriocins in the European Union
The use of microbial protective cultures is not generally regulated as an ingredient, additive, or processing aid in the regulatory framework of the European Union (EU) with its 27 member states and the European Economic Area (EEA) and does not necessarily require approval and safety assessment. The duty of care and the sole responsibility for the specific use lies for the time being solely by the user. Further regulation is carried out outside the legal requirements of the Basic Food Regulation (EC) No. 178/2002 [112] on the basis of its technological purpose. Only Denmark and France are exceptions and register microbiological cultures on a national level as additives [113,114], regardless of their intended use, and require in part proof of efficacy and safety in addition to notification and approval. However, this does not reduce the free market access of foods treated with protective cultures from other EU countries.
In addition, food- or feed-associated microorganisms must always undergo a safety assessment and be approved by the European Food Safety Authority (EFSA) [115] in the EU if they are considered genetically modified (GMO) by Directive 2009/41/EC [116], Regulation (EC) No 1829/2003 [117], and Directive 2001/18/EC [118]. They are classified as a novel food by Regulation (EU) No 2015/2283 [119] or are considered a feed additive according to Regulation (EC) No 1831/2003 [120]. In 2006, the EU Standing Committee on the Food Chain and Animal Health proposed that microbial cultures with a technological purpose, such as food preservation, should be considered as additives [121], which would require safety assessment and approval of preservative cultures by the EFSA. This turns out to be problematic if they originally belong to or have developed from starter cultures that are not specifically regulated in the EU. Starter cultures also have the additional benefit of reducing the potential risk of pathogenic microorganisms. Therefore, a differentiation must be made according to the basic principle of the effectiveness of the protective culture.
The effect is based on displacement cause by microorganisms that have a higher colonization rate. They can outcompete nutrients through more efficient transport systems of carbon sources or by an accelerated metabolism. They also may exert superior adaptation to aerobic or anaerobic, or other growth conditions. In such cases, the activity is attributed to a non-specific protective culture [19,122,123]. This also applies to the antagonistic formation of acids such as lactic, acetic, benzoic, malic, succinic, and formic acid or ethanol, diacetyl, hydrogen peroxide, and carbon dioxide [29,35,40,124].
Specific protective cultures are distinguished from non-specific ones by their technological purpose and their action by means of bacteriocins, proteins, or protein-like compounds, such as nisin, with antagonistic activity against defined foodborne pathogens [75]. This distinction results from the European Food Additives Regulation (EC) No 1333/2008 [125] and the prohibition of the use of unauthorized substances in food according to Article 5 of Regulation (EC) No 1333/2008 [125].
The prohibition does not refer to the microorganisms themselves but to the substances produced by them. This is further differentiated in the Enzyme Regulation (EC) No. 1332/2008 [126]; the exemption from approval and regulation does not apply to the use of microorganism cultures for the targeted production of substances with a technological effect. LAB cultures, for example, can be used without prior approval as protective cultures that produce not only lactic acid but also the bacteriocin nisin (E234), which is approved as a food additive. However, the maximum permissible quantities for nisin must not be exceeded and nisin must be declared accordingly (Table 2). Apart from nisin, there are no other bacteriocins permitted as food additives in the Additives Ordinance as amended on 31 October 2022. A distinction between non-specific and specific protective cultures is not necessary for all microbial cultures; if they were not contained in foodstuffs in the European Union before 14 May 1997 [127] and were marketed in significant quantities for consumption, they all fall under the novel food Regulation (EU) No 2015/2283 [119] and require a marketing authorization.
If an application for market authorization and with it a safety assessment on the part of EFSA is required, there are two approaches depending on the organism used. In principle, the application procedure for food additives is carried out in accordance with Regulation (EC) No 1331/2008 [128]. The technical and administrative requirements for an application to be submitted are set out in Regulation (EC) No 234/2011 [129]. In addition, EFSA provides practical guidelines to support the submission of an application for marketing authorization [130].
The two approaches for safety assessment differ in terms of their depth of testing. If the EFSA already has a qualified presumption of safety (QPS) status for the microorganism used, an accelerated generic safety assessment will be performed. This procedure was introduced by the EFSA in 2007 to simplify the marketing authorization process and save resources. For a microorganism to have QPS status, the taxonomic unit of the organism used must be at species level for bacteria, fungi, and microalgae/protists and at the family level for viruses, and it must be on the QPS list [131] maintained by the EFSA. This QPS list is reviewed by the EFSA Panel of Biological Hazard (BIOHAZ) and updated every six months.
If a microorganism is used that does not have QPS status, there may be two reasons: First, the organism has never been evaluated because no dossier was ever submitted for authorization and therefore the QPS evaluation process was not initiated by EFSA. Second, the micro-organism in question was not granted QPS status following a QPS evaluation by the BIOHAZ Panel [132]. The QPS workflow by the BIOHAZ panel is shown in Figure 3. This can happen if the taxonomic status of the microorganism is ambiguous, if there is not sufficient study evidence for the microorganism, and/or if the microorganism used has potentially harmful characteristics such as pathogenicity, antibiotic resistance, or virulence factors or produces biologically active toxic secondary metabolites. However, this does not constitute a disqualification for marketing authorization, as a full safety review is foreseen for these organisms. The 2017 work by Laulund and colleagues describes possible tools that can be used to review a list of methods and procedures for assessing the safety of food cultures at the strain level and as quality assurance measures during production and in the final product [133]. These can be applied to protective cultures as well. An overview of all microorganisms, notified for marketing authorization since 2007, can be found on the list of microbiological agents as notified to the EFSA [134].
6. The Legal Use of Protection Cultures and Bacteriocins in the United States of America
In contrast to the partially undefined European legal framework for microbial cultures, the Federal Food, Drug, and Cosmetic Act [136] has provided a corresponding legal framework in the United States of America (US) since 1958, which was revised in 1997. Both legal frameworks are fundamentally different, even if at first glance there are two similar programs with the GRAS program [137] (generally recognized as safe) of the U.S. Food and Drug Administration (FDA) and the QPS list at the EFSA.
In the US, according to the Federal Food, Drug, and Cosmetic Act [136], a food additive is basically any substance that directly or indirectly becomes a component of a food or could influence the food in one way or another. Thus, unlike in the European Union, the term food additive is very broad and encompasses virtually as what comes into contact with food of a pre-approval by the FDA. The only exception is that it is a substance that has been recognized by the FDA generally recognized as safe (GRAS). Accordingly, microbial protective cultures, microbially derived ingredients such as bacteriocins can be introduced to the market either as a food additive or by obtaining GRAS status. While a food additive requires an evaluation and approval by application according to 21 CFR Part 171 [138], this is not necessary to obtain GRAS status. Here, the regulations according to 21 CFR 170.30 [139] apply, which contain some special features. The GRAS status is provided for one microorganism at a strain level and one specific application. This means that an additional GRAS status must be provided for each additional use case.
In addition, the GRAS status is not determined by the FDA, as is the case with food additives, but by qualified experts. The first distributor or manufacturer is free to decide how the expert panel is convened and staffed. An evaluation can also be carried out by the FDA if desired. The Guidance for Industry [140] assists in the implementation of the requirements under 21 CFR 170.30 [139]. If there is a GRAS conclusion by the expert panel, the marketer may make a notice of congestion to the FDA under 21 CFR 170 Subpart E [141]. However, the marketer or manufacturer is not required to do so under either the Federal Food, Drug, and Cosmetic Act [136] or 21 CFR 170.30 [139,140]. Upon submission of the GRAS conclusion, the FDA either agrees with the conclusion, and the GRAS status exists further, or denies it. However, even then it is not prohibited to use the particular culture in food under U.S. law unless the FDA issues a ban. In the event of a claim, the liability issues are different than with GRAS status granted by the FDA. Table 3 shows the main difference between the GRAS system and its European counterpart QPS.
7. The Legal Use of Protection Cultures and Bacteriocins in Latin America
In Mexico, the use of microbial protective cultures in food products is regulated by the Federal Commission for Protection against Sanitary Risks (COFEPRIS) under the Ministry of Health. The use of these cultures is permitted only when they are considered safe for human consumption and do not alter the nutritional value of the food product. The manufacturer of the protective culture must provide evidence that the culture is safe and effective for use in the specific food product. This requires GRAS status on the part of the FDA or its homologation. Further, a sample of the microbiological culture used must be deposited in the strain collection for microorganisms of the National Institute of Forestry, Agriculture and Livestock Research (INIFAP) [142].
The use of microbial protective cultures in Brazil is regulated by the National Health Surveillance Agency (ANVISA). ANVISA requires that manufacturers of protective cultures provide evidence that the culture is safe and effective for use in food products. Since the revision of Resolution RDC 27/2010: Categories of food and packaging that require pre-market approval by Anvisa by Resolution—RDC No. 240, of 26 July 2018 the registration and approval of microbial cultures as technical adjuvants is no longer required [143].
In Peru, the General Directorate of Environmental Health (DIGESA) of the Ministry of Health is responsible for regulating the food industry. Currently, there is no regulatory framework for the use of microbial protection cultures in food.
8. The Legal Use of Protection Cultures and Bacteriocins in Asia
The regulatory framework for microbial-protective cultures in China is primarily governed by the Food Safety Law, which sets the basic principles for food safety, and the Regulations for the Administration of Food Additives, which provides specific requirements for the use of food additives, including microbial protective cultures. The regulatory authorities in China are the National Health Commission and the Department of Food Safety Standards, Monitoring and Evaluation, which since 2010 has had a separate regulation for food cultures, including protective cultures. From this date, all new strains placed on the market must be on the official list. The list was last updated on 18 August 2022 [144].
Some other countries have also established positive lists for food and protective crops. Examples include Thailand and Malaysia. In Thailand, industrially used cultures are considered food additives. The regulatory framework requires the submission of strain level documentation for registration with the Food and Drug Administration [145]. Traditional fermented foods (spontaneous fermentation) without the use of food cultures are not subject to registration. Since 2014, Malaysia has provided a “New Regulations 26B” with its Malaysian Food Act, which provides the regulatory framework for “Microbial Cultures for Food Fermentation”. Protective cultures also fall under this regulation [146].
9. Market Overview
The economic importance of protective cultures for food processing should be appreciated. The global market volume in 2022 was already 1.3 billion USD [147] in the food and beverage sector. Consumers now expect not only safe food but also healthy food that is as natural as possible [148]. In addition, social goals such as sustainable ecological management and the reduction of food waste are increasingly prompting food companies to take action. Companies can meet these challenges through the innovative use of microbial protection cultures. Several studies have shown that the use of protective cultures on food can extend the shelf life of products such as fish or meat, thus avoiding food waste [56,75,149,150,151,152]. The potential elimination of refrigeration contributes to carbon reduction and sustainability goals and enables safe food in regions without refrigeration infrastructure. Prolonged storage of grapes (12 days) and meat (2 days) at room temperature, rather than in refrigeration, after treatment with a protective culture or its ferment, demonstrated the potential benefits in this case [153,154]. In addition, the elimination of artificial preservatives brings healthier and more natural products to the market. Consequently, there is a great development and growth potential for protective cultures in the food sector, which market research companies estimate to have an annual growth rate between 4.3% and 23.3% by the end of the decade [147,155,156]. Market development is not only driven by further growth in Europe and North America but also by the Asian market, which is home to 60% of the world’s population and is developing consumer needs comparable to those in Europe and the US [147,155].
However, the market for protective cultures is quite fragmented and is not dominated by large international companies. In addition to finished products, start-ups and medium-sized companies in particular develop customer- and product-specific cultures [147,155]. Protective cultures are primarily offered as freeze-dried or frozen preparations and as ferments. The only bacteriocin used in pure form is nisin. Table 4 shows a compilation of ready-to-use commercial protective cultures available in the European Union categorized by food application. As shown in Figure 4, almost 60% of the listed commercial protective cultures address a single category, that of milk and dairy products, and currently represent the largest area of application for protective cultures.
10. Conclusions
The implementation of protective cultures of lactic acid bacteria in diverse perishable food products contributes substantially to food protection, food safety and food security. The here thoroughly discussed regulatory frameworks show that the use of foods harboring protective cultures is superior to the application of specifically defined agents, such as a pure bacteriocins. This accounts even more regarding the production costs. The current state of knowledge through studies providing proof of principle for all categories of risky foods shows that a wealth of protective cultures is available. Furthermore, nature still has an untapped unlimited reservoir for more protective bacterial species that remain to uncover. Bio-protection could thus contribute to nourish the hundreds of millions of starving people and to reduce significantly greenhouse gas emissions by food waste reduction. The biggest challenge to making this true is probably to speed up the transfer of scientific knowledge towards food production companies.
Author Contributions
Conceptualization, S.W.F.; methodology, S.W.F.; validation, S.W.F. and F.T.; formal analysis, S.W.F.; investigation, S.W.F.; resources, S.W.F.; data curation, S.W.F.; writing—original draft preparation, S.W.F.; writing—review and editing, S.W.F. and F.T.; visualization, S.W.F. and F.T.; supervision, F.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We are grateful to our students Leonie Bertels, Anna Euler, Nikola Johannsen, Frank Meyer, and Jennifer Wachtarczyk for experimental assistance. We thank Nadine Mariani Corea and Jan Kok for expert assistance and many helpful discussions. We are grateful to Till Sander-Titgemeyer for professional graphical design support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAO. The State of Food Security and Nutrition in the World 2022; FAO: Rome, Italy, 2022; ISBN 978-92-5-136499-4. [Google Scholar]
- Kirk, M.D.; Pires, S.M.; Black, R.E.; Caipo, M.; Crump, J.A.; Devleesschauwer, B.; Döpfer, D.; Fazil, A.; Fischer-Walker, C.L.; Hald, T.; et al. World Health Organization Estimates of the Global and Regional Disease Burden of 22 Foodborne Bacterial, Protozoal, and Viral Diseases, 2010: A Data Synthesis. PLoS Med. 2015, 12, e1001921. [Google Scholar] [CrossRef] [Green Version]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.-A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne Illness Acquired in the United States—Major Pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Pires, S.M.; Desta, B.N.; Mughini-Gras, L.; Mmbaga, B.T.; Fayemi, O.E.; Salvador, E.M.; Gobena, T.; Majowicz, S.E.; Hald, T.; Hoejskov, P.S.; et al. Burden of Foodborne Diseases: Think Global, Act Local. Curr. Opin. Food Sci. 2021, 39, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Liu, G.; Parfitt, J.; Liu, X.; Van Herpen, E.; Stenmarck, Å.; O’Connor, C.; Östergren, K.; Cheng, S. Missing Food, Missing Data? A Critical Review of Global Food Losses and Food Waste Data. Environ. Sci. Technol. 2017, 51, 6618–6633. [Google Scholar] [CrossRef]
- Food Wastage Footprint. Food Wastage Footprint Full-Cost Accounting: Final Report; Food Wastage Footprint: Rome, Italy, 2014; ISBN 978-92-5-108512-7. [Google Scholar]
- Rahman, S. (Ed.) Handbook of Food Preservation, 2nd ed.; Food Science and Technology; CRC Press: Boca Raton, FL, USA, 2007; ISBN 978-1-57444-606-7. [Google Scholar]
- Zhong, Y.; Wu, L.; Chen, X.; Huang, Z.; Hu, W. Effects of Food-Additive-Information on Consumers’ Willingness to Accept Food with Additives. Int. J. Environ. Res. Public Health 2018, 15, 2394. [Google Scholar] [CrossRef] [Green Version]
- Hutkins, R.W. Microbiology and Technology of Fermented Foods, 2nd ed.; IFT PRESS Series; Wiley Blackwell: Hoboken, NJ, USA, 2019; ISBN 978-1-119-02756-0. [Google Scholar]
- Cheung, C. Managing Food Storage in the Roman Empire. Quat. Int. 2021, 597, 63–75. [Google Scholar] [CrossRef]
- Metchnikoff, É. The Prolongation of Life- Optimistic Studies; The Knickerbocker Press: New York, NY, USA; London, UK, 1908. [Google Scholar]
- Vasiljevic, T.; Shah, N.P. Probiotics—From Metchnikoff to Bioactives. Int. Dairy. J. 2008, 18, 714–728. [Google Scholar] [CrossRef]
- Hansen, E.B. Commercial Bacterial Starter Cultures for Fermented Foods of the Future. Int. J. Food Microbiol. 2002, 78, 119–131. [Google Scholar] [CrossRef]
- Ibrahim, S.A.; Ayivi, R.D.; Zimmerman, T.; Siddiqui, S.A.; Altemimi, A.B.; Fidan, H.; Esatbeyoglu, T.; Bakhshayesh, R.V. Lactic Acid Bacteria as Antimicrobial Agents: Food Safety and Microbial Food Spoilage Prevention. Foods 2021, 10, 3131. [Google Scholar] [CrossRef]
- Ben Said, L.; Gaudreau, H.; Dallaire, L.; Tessier, M.; Fliss, I. Bioprotective Culture: A New Generation of Food Additives for the Preservation of Food Quality and Safety. Ind. Biotechnol. 2019, 15, 138–147. [Google Scholar] [CrossRef]
- Jay, J.M. Microorganisms in Fresh Ground Meats: The Relative Safety of Products with Low versus High Numbers. Meat Sci. 1996, 43, 59–66. [Google Scholar] [CrossRef]
- Mellefont, L.A.; McMeekin, T.A.; Ross, T. Effect of Relative Inoculum Concentration on Listeria Monocytogenes Growth in Co-Culture. Int. J. Food Microbiol. 2008, 121, 157–168. [Google Scholar] [CrossRef]
- Jameson, J.E. A Discussion of the Dynamics of Salmonella Enrichment. J. Hyg. 1962, 60, 193–207. [Google Scholar] [CrossRef] [Green Version]
- Bolívar, A.; Tarlak, F.; Costa, J.C.C.P.; Cejudo-Gómez, M.; Bover-Cid, S.; Zurera, G.; Pérez-Rodríguez, F. A New Expanded Modelling Approach for Investigating the Bioprotective Capacity of Latilactobacillus Sakei CTC494 against Listeria Monocytogenes in Ready-to-Eat Fish Products. Food Res. Int. 2021, 147, 110545. [Google Scholar] [CrossRef]
- Serra-Castelló, C.; Costa, J.C.C.P.; Jofré, A.; Bolívar, A.; Pérez-Rodríguez, F.; Bover-Cid, S. A Mathematical Model to Predict the Antilisteria Bioprotective Effect of Latilactobacillus Sakei CTC494 in Vacuum Packaged Cooked Ham. Int. J. Food Microbiol. 2022, 363, 109491. [Google Scholar] [CrossRef]
- Cornu, M.; Billoir, E.; Bergis, H.; Beaufort, A.; Zuliani, V. Modeling Microbial Competition in Food: Application to the Behavior of Listeria Monocytogenes and Lactic Acid Flora in Pork Meat Products. Food Microbiol. 2011, 28, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Rigali, S.; Titgemeyer, F.; Barends, S.; Mulder, S.; Thomae, A.W.; Hopwood, D.A.; van Wezel, G.P. Feast or Famine: The Global Regulator DasR Links Nutrient Stress to Antibiotic Production by Streptomyces. EMBO Rep. 2008, 9, 670–675. [Google Scholar] [CrossRef] [Green Version]
- Molloy, S. From Start to Finish for Streptomyces. Nat. Rev. Microbiol. 2008, 6, 569. [Google Scholar] [CrossRef]
- Nothaft, H.; Rigali, S.; Boomsma, B.; Swiatek, M.; McDowall, K.J.; van Wezel, G.P.; Titgemeyer, F. The Permease Gene NagE2 Is the Key to N. -Acetylglucosamine Sensing and Utilization in Streptomyces Coelicolor and Is Subject to Multi-Level Control. Mol. Microbiol. 2010, 75, 1133–1144. [Google Scholar] [CrossRef]
- Van Bergeijk, D.A.; Terlouw, B.R.; Medema, M.H.; van Wezel, G.P. Ecology and Genomics of Actinobacteria: New Concepts for Natural Product Discovery. Nat. Rev. Microbiol. 2020, 18, 546–558. [Google Scholar] [CrossRef]
- Lindgren, S.E.; Dobrogosz, W.J. Antagonistic Activities of Lactic Acid Bacteria in Food and Feed Fermentations. FEMS Microbiol. Lett. 1990, 87, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Van de Guchte, M.M.; Serror, P.P.; Chervaux, C.; Smokvina, T.; Ehrlich, S.S.; Maguin, E. Stress Responses in Lactic Acid Bacteria. Antonie Van Leeuwenhoek 2002, 82, 187. [Google Scholar] [CrossRef] [PubMed]
- Lund, P.A.; De Biase, D.; Liran, O.; Scheler, O.; Mira, N.P.; Cetecioglu, Z.; Fernández, E.N.; Bover-Cid, S.; Hall, R.; Sauer, M.; et al. Understanding How Microorganisms Respond to Acid PH Is Central to Their Control and Successful Exploitation. Front. Microbiol. 2020, 11, 556140. [Google Scholar] [CrossRef] [PubMed]
- Hirshfield, I.N.; Terzulli, S.; O’Byrne, C. Weak Organic Acids: A Panoply of Effects on Bacteria. Sci. Prog. 2003, 86, 245–270. [Google Scholar] [CrossRef] [PubMed]
- Wadhwa, N.; Berg, H.C. Bacterial Motility: Machinery and Mechanisms. Nat. Rev. Microbiol. 2022, 20, 161–173. [Google Scholar] [CrossRef]
- Nakajima, H.; Toba, T.; Ito, A.; Kudo, S.; Sato, S.; Sato, Y. The Screening of Hydrogen Peroxide-Producing Lactic Acid Bacteria and Their Application to Inactivating Psychrotrophic Food-Borne Pathogens. Curr. Microbiol. 2003, 47, 231–236. [Google Scholar] [CrossRef]
- Hertzberger, R.; Arents, J.; Dekker, H.L.; Pridmore, R.D.; Gysler, C.; Kleerebezem, M.; de Mattos, M.J.T. H2O2 Production in Species of the Lactobacillus Acidophilus Group: A Central Role for a Novel NADH-Dependent Flavin Reductase. Appl. Environ. Microbiol. 2014, 80, 2229–2239. [Google Scholar] [CrossRef] [Green Version]
- Khurshid, H. Antimicrobial Properties of Hydrogen Peroxide and Potash Alum Alone and in Combination against Clinical Bacterial Isolates. Pure Appl. Biol. 2019, 8, 2238–2247. [Google Scholar] [CrossRef]
- Andrés, C.M.C.; Pérez de la Lastra, J.M.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. Chemistry of Hydrogen Peroxide Formation and Elimination in Mammalian Cells, and Its Role in Various Pathologies. Stresses 2022, 2, 256–274. [Google Scholar] [CrossRef]
- Lanciotti, R.; Patrignani, F.; Bagnolini, F.; Guerzoni, M.E.; Gardini, F. Evaluation of Diacetyl Antimicrobial Activity against Escherichia Coli, Listeria Monocytogenes and Staphylococcus Aureus. Food Microbiol. 2003, 20, 537–543. [Google Scholar] [CrossRef]
- Jay, J.M. Antimicrobial Properties of Diacetylt. Appl. Environ. Microbiol. 1982, 44, 525–532. [Google Scholar] [CrossRef] [Green Version]
- Bjeldanes, L.F.; Chew, H. Mutagenicity of 1,2-Dicarbonyl Compounds: Maltol, Kojic Acid, Diacetyl and Related Substances. Mutat. Res. Genet. Toxicol. 1979, 67, 367–371. [Google Scholar] [CrossRef]
- Whittaker, P.; Clarke, J.J.; San, R.H.C.; Begley, T.H.; Dunkel, V.C. Evaluation of the Butter Flavoring Chemical Diacetyl and a Fluorochemical Paper Additive for Mutagenicity and Toxicity Using the Mammalian Cell Gene Mutation Assay in L5178Y Mouse Lymphoma Cells. Food Chem. Toxicol. 2008, 46, 2928–2933. [Google Scholar] [CrossRef]
- Shibamoto, T. Diacetyl: Occurrence, Analysis, and Toxicity. J. Agric. Food Chem. 2014, 62, 4048–4053. [Google Scholar] [CrossRef]
- Park, D.H. Effects of Carbon Dioxide on Metabolite Production and Bacterial Communities during Kimchi Fermentation. Biosci. Biotechnol. Biochem. 2018, 82, 1234–1242. [Google Scholar] [CrossRef]
- Martley, F.G.; Crow, V.L. Open Texture in Cheese: The Contributions of Gas Production by Microorganisms and Cheese Manufacturing Practices. J. Dairy Res. 1996, 63, 489–507. [Google Scholar] [CrossRef]
- De Vuyst, L.; Vandamme, E.J. Antimicrobial Potential of Lactic Acid Bacteria. In Bacteriocins of Lactic Acid Bacteria: Microbiology, Genetics and Applications; De Vuyst, L., Vandamme, E.J., Eds.; Springer: Boston, MA, USA, 1994; pp. 91–142. ISBN 978-1-4615-2668-1. [Google Scholar]
- Schaefer, L.; Auchtung, T.A.; Hermans, K.E.; Whitehead, D.; Borhan, B.; Britton, R.A. The Antimicrobial Compound Reuterin (3-Hydroxypropionaldehyde) Induces Oxidative Stress via Interaction with Thiol Groups. Microbiology 2010, 156, 1589–1599. [Google Scholar] [CrossRef] [Green Version]
- Vimont, A.; Fernandez, B.; Ahmed, G.; Fortin, H.-P.; Fliss, I. Quantitative Antifungal Activity of Reuterin against Food Isolates of Yeasts and Moulds and Its Potential Application in Yogurt. Int. J. Food Microbiol. 2019, 289, 182–188. [Google Scholar] [CrossRef]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A Viable Alternative to Antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef]
- Heilbronner, S.; Krismer, B.; Brötz-Oesterhelt, H.; Peschel, A. The Microbiome-Shaping Roles of Bacteriocins. Nat. Rev. Microbiol. 2021, 19, 726–739. [Google Scholar] [CrossRef]
- Alvarez-Sieiro, P.; Montalbán-López, M.; Mu, D.; Kuipers, O.P. Bacteriocins of Lactic Acid Bacteria: Extending the Family. Appl. Microbiol. Biotechnol. 2016, 100, 2939–2951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, C.C.G.; Silva, S.P.M.; Ribeiro, S.C. Application of Bacteriocins and Protective Cultures in Dairy Food Preservation. Front. Microbiol. 2018, 9, 594. [Google Scholar] [CrossRef] [PubMed]
- Kjos, M.; Nes, I.F.; Diep, D.B. Mechanisms of Resistance to Bacteriocins Targeting the Mannose Phosphotransferase System. Appl. Env. Microbiol. 2011, 77, 3335–3342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gargis, A.S.; O’Rourke, A.-L.D.; Sloan, G.L.; Simmonds, R.S. Prevalence and Acquisition of the Genes for Zoocin A and Zoocin A Resistance in Streptococcus Equi Subsp. Zooepidemicus. J. Mol. Evol. 2009, 68, 498–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Heel, A.J.; de Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: A User-Friendly Web Server to Thoroughly Mine RiPPs and Bacteriocins. Nucleic Acids Res. 2018, 46, W278–W281. [Google Scholar] [CrossRef]
- Balasubramanian, S.; Chen, J.; Wigneswaran, V.; Bang-Berthelsen, C.H.; Jensen, P.R. Droplet-Based Microfluidic High Throughput Screening of Corynebacterium Glutamicum for Efficient Heterologous Protein Production and Secretion. Front. Bioeng. Biotechnol. 2021, 9, 668531. [Google Scholar] [CrossRef]
- Iacumin, L.; Cappellari, G.; Colautti, A.; Comi, G. Listeria Monocytogenes Survey in Cubed Cooked Ham Packaged in Modified Atmosphere and Bioprotective Effect of Selected Lactic Acid Bacteria. Microorganisms 2020, 8, 898. [Google Scholar] [CrossRef]
- Chaillou, S.; Christieans, S.; Rivollier, M.; Lucquin, I.; Champomier-Vergès, M.C.; Zagorec, M. Quantification and Efficiency of Lactobacillus Sakei Strain Mixtures Used as Protective Cultures in Ground Beef. Meat Sci. 2014, 97, 332–338. [Google Scholar] [CrossRef]
- Vescovo, M.; Scolari, G.; Zacconi, C. Inhibition of Listeria Innocua Growth by Antimicrobial-Producing Lactic Acid Cultures in Vacuum-Packed Cold-Smoked Salmon. Food Microbiol. 2006, 23, 689–693. [Google Scholar] [CrossRef]
- Leroi, F.; Cornet, J.; Chevalier, F.; Cardinal, M.; Coeuret, G.; Chaillou, S.; Joffraud, J.-J. Selection of Bioprotective Cultures for Preventing Cold-Smoked Salmon Spoilage. Int. J. Food Microbiol. 2015, 213, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Coelho, M.C.; Silva, C.C.G.; Ribeiro, S.C.; Dapkevicius, M.L.N.E.; Rosa, H.J.D. Control of Listeria Monocytogenes in Fresh Cheese Using Protective Lactic Acid Bacteria. Int. J. Food Microbiol. 2014, 191, 53–59. [Google Scholar] [CrossRef]
- Agriopoulou, S.; Stamatelopoulou, E.; Sachadyn-Król, M.; Varzakas, T. Lactic Acid Bacteria as Antibacterial Agents to Extend the Shelf Life of Fresh and Minimally Processed Fruits and Vegetables: Quality and Safety Aspects. Microorganisms 2020, 8, 952. [Google Scholar] [CrossRef]
- Köckerling, E.; Karrasch, L.; Schweitzer, A.; Razum, O.; Krause, G. Public Health Research Resulting from One of the World’s Largest Outbreaks Caused by Entero-Hemorrhagic Escherichia Coli in Germany 2011: A Review. Front. Public Health 2017, 5, 332. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Wolf, G.; Hammes, W.P. Indigenous Microorganisms from Iceberg Lettuce with Adherence and Antagonistic Potential for Use as Protective Culture. Innov. Food Sci. Emerg. Technol. 2006, 7, 294–301. [Google Scholar] [CrossRef]
- Rodgers, S. Novel Applications of Live Bacteria in Food Services: Probiotics and Protective Cultures. Trends Food Sci. Technol. 2008, 19, 188–197. [Google Scholar] [CrossRef]
- Makhal, S.; Kanawjia, S.K.; Giri, A. Effect of MicroGARD on Keeping Quality of Direct Acidified Cottage Cheese. J. Food Sci. Technol. 2015, 52, 936–943. [Google Scholar] [CrossRef] [Green Version]
- Quinto, E.J.; Marín, J.M.; Schaffner, D.W. Effect of the Competitive Growth of Lactobacillus Sakei MN on the Growth Kinetics of Listeria Monocytogenes Scott A in Model Meat Gravy. Food Control. 2016, 63, 34–45. [Google Scholar] [CrossRef]
- Mejlholm, O.; Dalgaard, P. Modelling and Predicting the Simultaneous Growth of Listeria Monocytogenes and Psychrotolerant Lactic Acid Bacteria in Processed Seafood and Mayonnaise-Based Seafood Salads. Food Microbiol. 2015, 46, 1–14. [Google Scholar] [CrossRef]
- Zhao, S.; Hao, X.; Yang, F.; Wang, Y.; Fan, X.; Wang, Y. Antifungal Activity of Lactobacillus plantarum ZZUA493 and Its Application to Extend the Shelf Life of Chinese Steamed Buns. Foods 2022, 11, 195. [Google Scholar] [CrossRef] [PubMed]
- Lauková, A.; Focková, V.; Pogány Simonová, M. Enterococcus Mundtii Isolated from Slovak Raw Goat Milk and Its Bacteriocinogenic Potential. Int. J. Environ. Res. Public Health 2020, 17, 9504. [Google Scholar] [CrossRef]
- Tulini, F.L.; Hymery, N.; Haertlé, T.; Le Blay, G.; De Martinis, E.C.P. Screening for Antimicrobial and Proteolytic Activities of Lactic Acid Bacteria Isolated from Cow, Buffalo and Goat Milk and Cheeses Marketed in the Southeast Region of Brazil. J. Dairy Res. 2016, 83, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Pang, H.; Wang, L.; Ma, C.; Wu, G.; Liu, Y.; Guan, Y.; Zhang, M.; Qin, G.; Tan, Z. Bacteriocin-Producing Lactic Acid Bacteria Strains with Antimicrobial Activity Screened from Bamei Pig Feces. Foods 2022, 11, 709. [Google Scholar] [CrossRef] [PubMed]
- Tuntisuwanno, N.; Charernjiratrakul, W.; Bovornruengroj, N.; Borvornruengroj, P. Selection of Biopreservative-Produced Lactic Acid Bacteria from Chilled Seafood Products. CMUJNS 2014, 13, 459–468. [Google Scholar] [CrossRef]
- Gustaw, K.; Michalak, M.; Polak-Berecka, M.; Waśko, A. Isolation and Characterization of a New Fructophilic Lactobacillus Plantarum FPL Strain from Honeydew. Ann. Microbiol. 2018, 68, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.M.; Lee, Y. A Differential Medium for Lactic Acid-Producing Bacteria in a Mixed Culture. Lett. Appl. Microbiol. 2008, 46, 676–681. [Google Scholar] [CrossRef]
- Ricciardi, A.; Parente, E.; Tramutola, A.; Guidone, A.; Ianniello, R.G.; Pavlidis, D.; Tsakalidou, E.; Zotta, T. Evaluation of a Differential Medium for the Preliminary Identification of Members of the Lactobacillus Plantarum and Lactobacillus Casei Groups. Ann. Microbiol. 2015, 65, 1649–1658. [Google Scholar] [CrossRef]
- Galat, A.; Dufresne, J.; Combrisson, J.; Thépaut, J.; Boumghar-Bourtchai, L.; Boyer, M.; Fourmestraux, C. Novel Method Based on Chromogenic Media for Discrimination and Selective Enumeration of Lactic Acid Bacteria in Fermented Milk Products. Food Microbiol. 2016, 55, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Fijan, S.; Šulc, D.; Steyer, A. Study of the In Vitro Antagonistic Activity of Various Single-Strain and Multi-Strain Probiotics against Escherichia Coli. Int. J. Environ. Res. Public Health 2018, 15, 1539. [Google Scholar] [CrossRef] [Green Version]
- Aymerich, T.; Martín, B.; Garriga, M.; Hugas, M. Microbial Quality and Direct PCR Identification of Lactic Acid Bacteria and Nonpathogenic Staphylococci from Artisanal Low-Acid Sausages. Appl. Environ. Microbiol. 2003, 69, 12. [Google Scholar] [CrossRef] [Green Version]
- Ben Amor, K.; Vaughan, E.E.; de Vos, W.M. Advanced Molecular Tools for the Identification of Lactic Acid Bacteria. J. Nutr. 2007, 137, 741S–747S. [Google Scholar] [CrossRef] [Green Version]
- Yadav, M.K.; Singh, B.; Tiwari, S.K. Comparative Analysis of Inhibition-Based and Indicator-Independent Colorimetric Assay for Screening of Bacteriocin-Producing Lactic Acid Bacteria. Probiotics Antimicrob. Proteins 2021, 9, 687–695. [Google Scholar] [CrossRef]
- Adeniyi, B.A.; Adetoye, A.; Ayeni, F.A. Antibacterial Activities of Lactic Acid Bacteria Isolated from Cow Faeces against Potential Enteric Pathogens. Afr. Health Sci. 2015, 15, 888–895. [Google Scholar] [CrossRef] [Green Version]
- Vijay Simha, B.; Sood, S.K.; Kumariya, R.; Garsa, A.K. Simple and Rapid Purification of Pediocin PA-1 from Pediococcus Pentosaceous NCDC 273 Suitable for Industrial Application. Microbiol. Res. 2012, 167, 544–549. [Google Scholar] [CrossRef]
- Hettiarachchi, S.A.; Lee, S.-J.; Lee, Y.; Kwon, Y.-K.; Zoysa, M.D.; Moon, S.; Jo, E.; Kim, E.; Kang, D.-H.; Heo, S.-J.; et al. A Rapid and Efficient Screening Method for Antibacterial Compound-Producing Bacteria. J. Microbiol. Biotechnol. 2017, 27, 1441–1448. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Guimaraes, A.; Venancio, A. Antifungal Effect of Organic Acids from Lactic Acid Bacteria on Penicillium Nordicum. Food Addit. Contam. Part A Chem. Anal. Control. 2018, 35, 1803–1818. [Google Scholar]
- Tomás, M.S.J.; Claudia Otero, M.; Ocaña, V.; Elena Nader-Macías, M. Production of Antimicrobial Substances by Lactic Acid Bacteria I: Determination of Hydrogen Peroxide. In Methods in Molecular Biology; Spencer, J.F.T., Ed.; Public Health Microbiology: Methods and Protocols; Humana Press: Totowa, NJ, USA, 2004; Volume 268, pp. 337–346. ISBN 978-1-59259-766-6. [Google Scholar]
- Tomás, M.S.J.; Bru, E.; Nader-Macías, M.E. Comparison of the Growth and Hydrogen Peroxide Production by Vaginal Probiotic Lactobacilli under Different Culture Conditions. Am. J. Obstet. Gynecol. 2003, 188, 35–44. [Google Scholar] [CrossRef]
- Muñoz-Atienza, E.; Gómez-Sala, B.; Araújo, C.; Campanero, C.; del Campo, R.; Hernández, P.E.; Herranz, C.; Cintas, L.M. Antimicrobial Activity, Antibiotic Susceptibility and Virulence Factors of Lactic Acid Bacteria of Aquatic Origin Intended for Use as Probiotics in Aquaculture. BMC Microbiol. 2013, 13, 15. [Google Scholar] [CrossRef] [Green Version]
- Sirichoat, A.; Flórez, A.B.; Vázquez, L.; Buppasiri, P.; Panya, M.; Lulitanond, V.; Mayo, B. Antibiotic Susceptibility Profiles of Lactic Acid Bacteria from the Human Vagina and Genetic Basis of Acquired Resistances. Int. J. Med. Sci. 2020, 21, 2594. [Google Scholar] [CrossRef] [Green Version]
- Panthee, S.; Paudel, A.; Blom, J.; Hamamoto, H.; Sekimizu, K. Complete Genome Sequence of Weissella Hellenica 0916-4-2 and Its Comparative Genomic Analysis. Front. Microbiol. 2019, 10, 1619. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, J.M.; Martínez, M.I.; Kok, J. Pediocin PA-1, a Wide-Spectrum Bacteriocin from Lactic Acid Bacteria. Crit. Rev. Food Sci. Nutr. 2002, 42, 91–121. [Google Scholar] [CrossRef] [Green Version]
- Yi, L.; Qi, T.; Hong, Y.; Deng, L.; Zeng, K. Screening of Bacteriocin-Producing Lactic Acid Bacteria in Chinese Homemade Pickle and Dry-Cured Meat, and Bacteriocin Identification by Genome Sequencing. LWT 2020, 125, 109177. [Google Scholar] [CrossRef]
- Papadimitriou, K.; Venieraki, A.; Tsigkrimani, M.; Katinakis, P.; Skandamis, P.N. Whole-Genome Sequence Data of the Proteolytic and Bacteriocin Producing Strain Enterococcus Faecalis PK23 Isolated from the Traditional Halitzia Cheese Produced in Cyprus. Data Brief 2021, 38, 107437. [Google Scholar] [CrossRef] [PubMed]
- Rezaei Javan, R.; van Tonder, A.J.; King, J.P.; Harrold, C.L.; Brueggemann, A.B. Genome Sequencing Reveals a Large and Diverse Repertoire of Antimicrobial Peptides. Front. Microbiol. 2018, 9, 2012. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, J.; Albano, H.; Silva, B.; Almeida, M.H.; Nogueira, T.; Teixeira, P. Characterization of a Lactiplantibacillus Plantarum R23 Isolated from Arugula by Whole-Genome Sequencing and Its Bacteriocin Production Ability. Int. J. Environ. Res. Public Health 2021, 18, 5515. [Google Scholar] [CrossRef] [PubMed]
- Taskila, S. Industrial Production of Starter Cultures. In Starter Cultures in Food Production; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 79–100. ISBN 978-1-118-93379-4. [Google Scholar]
- Giraud, E.; Lelong, B.; Raimbault, M. Influence of PH and Initial Lactate Concentration on the Growth of Lactobacillus Plantarum. Appl. Microbiol. Biotechnol. 1991, 36, 96–99. [Google Scholar] [CrossRef]
- Partanen, L.; Marttinen, N.; Alatossava, T. Fats and Fatty Acids as Growth Factors for Lactobacillus Delbrueckii. Syst. Appl. Microbiol. 2001, 24, 500–506. [Google Scholar] [CrossRef]
- Morishita, T.; Deguchi, Y.; Yajima, M.; Sakurai, T.; Yura, T. Multiple Nutritional Requirements of Lactobacilli: Genetic Lesions Affecting Amino Acid Biosynthetic Pathways. J. Bacteriol. 1981, 148, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Cocaign-Bousquet, M.; Garrigues, C.; Novak, L.; Lindley, N.d.; Loublere, P. Rational Development of a Simple Synthetic Medium for the Sustained Growth of Lactococcus Lactis. J. Appl. Bacteriol. 1995, 79, 108–116. [Google Scholar] [CrossRef]
- Mora-Villalobos, J.A.; Montero-Zamora, J.; Barboza, N.; Rojas-Garbanzo, C.; Usaga, J.; Redondo-Solano, M.; Schroedter, L.; Olszewska-Widdrat, A.; López-Gómez, J.P. Multi-Product Lactic Acid Bacteria Fermentations: A Review. Fermentation 2020, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Leroy, F.; De Vuyst, L. Simulation of the Effect of Sausage Ingredients and Technology on the Functionality of the Bacteriocin-Producing Lactobacillus Sakei CTC 494 Strain. Int. J. Food Microbiol. 2005, 100, 141–152. [Google Scholar] [CrossRef]
- Yang, R.; Johnson, M.C.; Ray, B. Novel Method to Extract Large Amounts of Bacteriocins from Lactic Acid Bacteria. Appl. Env. Microbiol. 1992, 58, 3355–3359. [Google Scholar] [CrossRef] [Green Version]
- Leroy, F.; de Vuyst, L. Temperature and PH Conditions That Prevail during Fermentation of Sausages Are Optimal for Production of the Antilisterial Bacteriocin Sakacin, K. Appl. Environ. Microbiol. 1999, 65, 974–981. [Google Scholar] [CrossRef] [Green Version]
- Verluyten, J.; Messens, W.; De Vuyst, L. Sodium Chloride Reduces Production of Curvacin A, a Bacteriocin Produced by Lactobacillus Curvatus Strain LTH 1174, Originating from Fermented Sausage. Appl. Env. Microbiol. 2004, 70, 2271–2278. [Google Scholar] [CrossRef] [Green Version]
- Delgado, A.; Brito, D.; Peres, C.; Noé-Arroyo, F.; Garrido-Fernández, A. Bacteriocin Production by Lactobacillus Pentosus B96 Can Be Expressed as a Function of Temperature and NaCl Concentration. Food Microbiol. 2005, 22, 521–528. [Google Scholar] [CrossRef]
- Schöbitz, R.P.; Bórquez, P.A.; Costa, M.E.; Ciampi, L.R.; Brito, C.S. Bacteriocin like Substance Production by Carnobacterium Piscicola in a Continuous System with Three Culture Broths. Study of Antagonism against Listeria Monocytogenes on Vacuum Packaged Salmon. Braz. J. Microbiol. 2006, 37, 52–57. [Google Scholar] [CrossRef] [Green Version]
- Jack, R.W.; Tagg, J.R.; Ray, B. Bacteriocins of Gram-Positive Bacteria. Microbiol. Rev. 1995, 59, 171–200. [Google Scholar] [CrossRef]
- Wayah, S.B.; Philip, K. Characterization, Yield Optimization, Scale up and Biopreservative Potential of Fermencin SA715, a Novel Bacteriocin from Lactobacillus Fermentum GA715 of Goat Milk Origin. Microb. Cell. Factories 2018, 17, 125. [Google Scholar] [CrossRef] [Green Version]
- Dougherty, B.A.; Hill, C.; Weidman, J.F.; Richardson, D.R.; Venter, J.C.; Ross, R.P. Sequence and Analysis of the 60 Kb Conjugative, Bacteriocin-Producing Plasmid PMRC01 from Lactococcus Lactis DPC3147. Mol. Microbiol. 1998, 29, 1029–1038. [Google Scholar] [CrossRef]
- Weixler, D.; Berghoff, M.; Ovchinnikov, K.V.; Reich, S.; Goldbeck, O.; Seibold, G.M.; Wittmann, C.; Bar, N.S.; Eikmanns, B.J.; Diep, D.B.; et al. Recombinant Production of the Lantibiotic Nisin Using Corynebacterium Glutamicum in a Two-Step Process. Microb. Cell. Factories 2022, 21, 11. [Google Scholar] [CrossRef]
- Weixler, D.; Goldbeck, O.; Seibold, G.M.; Eikmanns, B.J.; Riedel, C.U. Towards Improved Resistance of Corynebacterium Glutamicum against Nisin. bioRxiv 2021, arXiv:2021.08.09.454123. [Google Scholar]
- Goldbeck, O.; Desef, D.N.; Ovchinnikov, K.V.; Perez-Garcia, F.; Christmann, J.; Sinner, P.; Crauwels, P.; Weixler, D.; Cao, P.; Becker, J.; et al. Establishing Recombinant Production of Pediocin PA-1 in Corynebacterium Glutamicum. Metab. Eng. 2021, 68, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Mesa-Pereira, B.; Rea, M.C.; Cotter, P.D.; Hill, C.; Ross, R.P. Heterologous Expression of Biopreservative Bacteriocins with a View to Low Cost Production. Front. Microbiol. 2018, 9, 1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Ibarreche, M.; Field, D.; Ross, R.P.; Hill, C. A Bioengineered Nisin Derivative to Control Streptococcus Uberis Biofilms. Appl. Env. Microbiol. 2021, 87, e00391-21. [Google Scholar] [CrossRef] [PubMed]
- European Parliament. Regulation (EC) No 178/2002 of the European Parliament and of the Council of 28 January 2002 Laying Down the General Principles and Requirements of Food Law, Establishing the European Food Safety Authority and Laying Down Procedures in Matters of Food Safety; European Parliament: Strasbourg, France, 2002; Volume 31. [Google Scholar]
- Wessels, S.; Axelsson, L.; Bech Hansen, E.; De Vuyst, L.; Laulund, S.; Lähteenmäki, L.; Lindgren, S.; Mollet, B.; Salminen, S.; von Wright, A. The Lactic Acid Bacteria, the Food Chain, and Their Regulation. Trends Food Sci. Technol. 2004, 15, 498–505. [Google Scholar] [CrossRef]
- Danish Veterinary and Food Administration. List of Notified Microbial Cultures Applied in Food. Oktober 2016. Available online: https://www.foedevarestyrelsen.dk/SiteCollectionDocuments/Kemi%20og%20foedevarekvalitet/Liste%20over%20anmeldte%20mikrobielle%20kulturer%20oktober%202016.pdf (accessed on 25 February 2023).
- European Parliament. Regulation (EU) 2019/1381 of the European Parliament and of the Council of 20 June 2019 on the Transparency and Sustainability of the EU Risk Assessment in the Food Chain and Amending Regulations (EC) No 178/2002, (EC) No 1829/2003, (EC) No 1831/2003, (EC) No 2065/2003, (EC) No 1935/2004, (EC) No 1331/2008, (EC) No 1107/2009, (EU) 2015/2283 and Directive 2001/18/EC (Text with EEA Relevance); European Parliament: Strasbourg, France, 2019; p. 231. [Google Scholar]
- European Parliament. Directive 2009/41/EC of the European Parliament and of the Council of 6 May 2009 on the Contained Use of Genetically Modified Micro-Organisms (Recast) (Text with EEA Relevance); European Parliament: Strasbourg, France, 2009; p. 125. [Google Scholar]
- European Parliament. Regulation (EC) No 1829/2003 of the European Parliament and of the Council of 22 September 2003 on Genetically Modified Food and Feed (Text with EEA Relevance); European Parliament: Strasbourg, France, 2003; p. 268. [Google Scholar]
- European Parliament. Directive 2001/18/EC of the European Parliament and of the Council of 12 March 2001 on the Deliberate Release into the Environment of Genetically Modified Organisms and Repealing Council Directive 90/220/EEC—Commission Declaration; European Parliament: Strasbourg, France, 2001; p. 106. [Google Scholar]
- European Parliament. Regulation (EU) 2015/2283 of the European Parliament and of the Council of 25 November 2015 on Novel Foods, Amending Regulation (EU) No 1169/2011 of the European Parliament and of the Council and Repealing Regulation (EC) No 258/97 of the European Parliament and of the Council and Commission Regulation (EC) No 1852/2001 (Text with EEA Relevance); European Parliament: Strasbourg, France, 2015; p. 327. [Google Scholar]
- European Parliament. Regulation (EC) No 1831/2003 of the European Parliament and of the Council of 22 September 2003 on Additives for Use in Animal Nutrition (Text with EEA Relevance); European Parliament: Strasbourg, France, 2003; p. 268. [Google Scholar]
- European Parliament. Directorate-General Health and Consumer Protection (SANCO) D1(06)D/413447, Summery Record of the Standing Committee on the Food Chain and Animal Health, Held in Brussels on 14 December 2006; European Parliament: Strasbourg, France, 2006. [Google Scholar]
- Stecchini, M.L.; Sarais, I.; de Bertoldi, M. The Influence of Lactobacillus Plantarum Culture Inoculation on the Fate of Staphylococcus Aureus and Salmonella typhimurium in Montasio Cheese. Int. J. Food Microbiol. 1991, 14, 99–109. [Google Scholar] [CrossRef]
- Bolívar, A.; Correia Peres Costa, J.C.; Posada-Izquierdo, G.D.; Bover-Cid, S.; Zurera, G.; Pérez-Rodríguez, F. Quantifying the Bioprotective Effect of Lactobacillus Sakei CTC494 against Listeria Monocytogenes on Vacuum Packaged Hot-Smoked Sea Bream. Food Microbiol. 2021, 94, 103649. [Google Scholar] [CrossRef]
- Juven, B.J.; Pierson, M.D. Antibacterial Effects of Hydrogen Peroxide and Methods for Its Detection and Quantitation. J. Food Prot. 1996, 59, 1233–1241. [Google Scholar] [CrossRef]
- European Parliament. Regulation (EC) No 1333/2008 of the European Parliament and of the Council of 16 December 2008 on Food Additives (Text with EEA Relevance); European Parliament: Strasbourg, France, 2008; Volume 354. [Google Scholar]
- European Parliament. Regulation (EC) No 1332/2008 of the European Parliament and of the Council of 16 December 2008 on Food Enzymes and Amending Council Directive 83/417/EEC, Council Regulation (EC) No 1493/1999, Directive 2000/13/EC, Council Directive 2001/112/EC and Regulation (EC) No 258/97 (Text with EEA Relevance); European Parliament: Strasbourg, France, 2008; Volume 354. [Google Scholar]
- European Parliament. Regulation (EC) No 258/97 of the European Parliament and of the Council of 27 January 1997 Concerning Novel Foods and Novel Food Ingredients; European Parliament: Strasbourg, France, 1997; Volume 43. [Google Scholar]
- European Parliament. Regulation (EC) No 1331/2008 of the European Parliament and of the Council of 16 December 2008 Establishing a Common Authorisation Procedure for Food Additives, Food Enzymes and Food Flavourings (Text with EEA Relevance)Text with EEA Relevance; European Parliament: Strasbourg, France, 2021. [Google Scholar]
- European Parliament. Commission Regulation (EU) No 234/2011 of 10 March 2011 Implementing Regulation (EC) No 1331/2008 of the European Parliament and of the Council Establishing a Common Authorisation Procedure for Food Additives, Food Enzymes and Food Flavourings (Text with EEA Relevance)Text with EEA Relevance; European Parliament: Strasbourg, France, 2021. [Google Scholar]
- EFSA. EFSA Panel on Food Additives and Nutrient Sources added to Food (ANS) Guidance for Submission for Food Additive Evaluations. EFSA J. 2012, 10, 2760. [Google Scholar] [CrossRef]
- Panel, E.B.; Koutsoumanis, K.; Allende, A.; Alvarez-Ordonez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; De Cesare, A.; Hilbert, F.; Lindqvist, R.; et al. Updated List of QPS-Recommended Microorganisms for Safety Risk Assessments Carried Out by EFSA 2023; Wiley: Hoboken, NJ, USA, 2023. [Google Scholar]
- EFSA Panel on Biological Hazards (BIOHAZ); Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Girones, R.; Herman, L.; Koutsoumanis, K.; Lindqvist, R.; et al. Scientific Opinion on the Update of the List of QPS-Recommended Biological Agents Intentionally Added to Food or Feed as Notified to EFSA. EFSA J. 2017, 15, e04664. [Google Scholar] [CrossRef] [Green Version]
- Laulund, S.; Wind, A.; Derkx, P.M.F.; Zuliani, V. Regulatory and Safety Requirements for Food Cultures. Microorganisms 2017, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Panel, E.B.; Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; De Cesare, A.; Hilbert, F.; Lindqvist, R.; et al. Microbiological Agents as Notified to EFSA 2023; Wiley: Hoboken, NJ, USA, 2023. [Google Scholar]
- Herman, L.; Chemaly, M.; Cocconcelli, P.S.; Fernandez, P.; Klein, G.; Peixe, L.; Prieto, M.; Querol, A.; Suarez, J.E.; Sundh, I.; et al. The Qualified Presumption of Safety Assessment and Its Role in EFSA Risk Evaluations: 15 Years Past. FEMS Microbiol. Lett. 2019, 366, fny260. [Google Scholar] [CrossRef]
- Federal Food, Drug, and Cosmetic Act, 21 U.S.C. § 301. Available online: https://www.govinfo.gov/content/pkg/USCODE-2021-title21/pdf/USCODE-2021-title21-chap9-subchapII-sec321.pdf (accessed on 25 February 2023).
- Substances Generally Recognized as Safe—Final Rule. Available online: file:///C:/Users/MDPI/Downloads/Substances-Generally-Recognized-as-Safe-(Final-Rule)-Regulatory-Impact-Analysis.pdf (accessed on 25 February 2023).
- Food Additive Petitions, 21 CFR Part 171. Available online: https://www.ecfr.gov/current/title-21/chapter-I/subchapter-B/part-171?toc=1 (accessed on 26 February 2023).
- 21 Food Additives CFR Part 170.30. Available online: https://www.ecfr.gov/current/title-21/chapter-I/subchapter-B/part-170 (accessed on 26 February 2023).
- Nutrition, C. for F.S. and A. Guidance for Industry: Regulatory Framework for Substances Intended for Use in Human Food or Animal Food on the Basis of the Generally Recognized as Safe (GRAS) Provision of the Federal Food, Drug, and Cosmetic Act 2022. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/guidance-industry-regulatory-framework-substances-intended-use-human-food-or-animal-food-basis (accessed on 26 February 2023).
- Generally Recognized as Safe (GRAS) Notice, 21 CFR Part 170 Subpart E. Available online: https://www.ecfr.gov/current/title-21/chapter-I/subchapter-B/part-170/subpart-E?toc=1 (accessed on 25 February 2023).
- AMEPRES. Probiótica, Suplementos Alimenticios y Alimentos Funcionales. Las Diferencias y Semejanzas, Así Como Su Entorno Regulatorio. Available online: https://amepresmexico.org.mx/wp-content/uploads/2018/07/Boletin-No14_OK.pdf (accessed on 31 March 2023).
- Gência Nacional de Vigilância Sanitária—Anvisa. Resolution RDC 27/2010: Categories of Foods and Packaging That Require a Pre-Market Approval by Anvisa; 2010; Volume RDC. Available online: http://antigo.anvisa.gov.br/legislacao#/visualizar/28564 (accessed on 31 March 2023).
- National Health Commission China. List of Strains That Can Be Used in Foods; 2022. Available online: http://www.nhc.gov.cn/sps/s7892/202208/1d6c229d6f744b35827e98161c146afb/files/f7177b32d93142f9827dcc55fb6889c2.pdf?_x_tr_sl=auto&_x_tr_tl=de&_x_tr_hl=de&_x_tr_pto=wapp&_x_tr_sch=http (accessed on 31 March 2023).
- Ministry of Public Health Thailand. Volume 133, Special Part 298, Government Gazette, 20 December 2016, Notification of the Ministry of Public Health (No. 381) B.E. 2559 on Food Additives (No. 4) 2016. Available online: http://food.fda.moph.go.th/law/data/announ_moph/P381.pdf (accessed on 31 March 2023).
- LAWS OF MALAYSIA. Regulation No.437—1985 Malaysian Food Act Version of 2014. Available online: https://importlicensing.wto.org/sites/default/files/members/91/Regulation%20No.437%20-%20Food%201985_01.01.2014.pdf (accessed on 31 March 2023).
- Future Market Insights. Food and Beverage Protective Cultures Market. 2022. Available online: https://www.futuremarketinsights.com/reports/food-and-beverage-protective-cultures-market (accessed on 26 February 2023).
- Grunert, K.G. Food Quality and Safety: Consumer Perception and Demand. Eur. Rev. Agric. Econ. 2005, 32, 369–391. [Google Scholar] [CrossRef]
- Inglin, R.C.; Delbrück, A.I.; Fässler, B.; Siebenmann, K.E.; Lacroix, C.; Stevens, M.J.A.; Meile, L. An Approach to Select Lactobacillus Isolates as Protective Cultures for Food Fermentations. J. Food Saf. 2018, 38, e12483. [Google Scholar] [CrossRef]
- Xu, M.M.; Kaur, M.; Pillidge, C.J.; Torley, P.J. Effect of Protective Cultures on Spoilage Bacteria and the Quality of Vacuum-Packaged Lamb Meat. Food Biosci. 2022, 50, 102148. [Google Scholar] [CrossRef]
- Xu, M.M. Protective Cultures as Natural Antimicrobials for Fresh Meat Shelf-Life Extension: Their Application and Consumer Acceptance. Ph.D. Thesis, RMIT University, Melbourne, VIC, Australia, 2022. [Google Scholar]
- Kim, C.R.; Hearnsberger, J.O.; Vickery, A.P.; White, C.H.; Marshall, D.L. Sodium Acetate and Bifidobacteria Increase Shelf-Life of Refrigerated Catfish Fillets. J. Food Sci. 1995, 60, 25–27. [Google Scholar] [CrossRef]
- Dopazo, V.; Luz, C.; Mañes, J.; Quiles, J.M.; Carbonell, R.; Calpe, J.; Meca, G. Bio-Preservative Potential of Microorganisms Isolated from Red Grape against Food Contaminant Fungi. Toxins 2021, 13, 412. [Google Scholar] [CrossRef]
- Le, N.T.T.; Bach, L.G.; Nguyen, D.C.; Le, T.H.X.; Pham, K.H.; Nguyen, D.H.; Hoang Thi, T.T. Evaluation of Factors Affecting Antimicrobial Activity of Bacteriocin from Lactobacillus Plantarum Microencapsulated in Alginate-Gelatin Capsules and Its Application on Pork Meat as a Bio-Preservative. Int. J. Environ. Res. Public Health 2019, 16, 1017. [Google Scholar] [CrossRef] [Green Version]
- Mordor Intelligence. Marktanalyse für Schutzkulturen, Größe, Anteil (2022—2027); Mordor Intelligence: Telangana, India, 2022. [Google Scholar]
- MDF ltd. Protective Cultures Market Size, Growth, Share | 2023–2028; Market Data Forecast: Telangana, India, 2022. [Google Scholar]
Figure 1.
Mode of antibacterial actions by LAB protective cultures. Shown are the major molecules that exert antibacterial activity and the identified targets.
Figure 1.
Mode of antibacterial actions by LAB protective cultures. Shown are the major molecules that exert antibacterial activity and the identified targets.

Figure 2.
Lactic acid bacteria: (A) Diverse blueish colonies of LAB species on mMRS-BPB differential agar. (B) In point inoculation assay of LAB (center colony) against a pathogenic indicator strain (bacterial lawn). Left without zone of inhibition (no bacteria-killing activity), right with zone of inhibition (bacteria-killing activity). (C) Detection of hydrogen peroxide with TMB; left, light colony showing no secretion; right, blue colony indicating secretion. (D) Detection of acid production by LAB (MRS agar plates); pH stripes: orange, acidification to pH 4.1; red, no acidification pH 6.1.
Figure 2.
Lactic acid bacteria: (A) Diverse blueish colonies of LAB species on mMRS-BPB differential agar. (B) In point inoculation assay of LAB (center colony) against a pathogenic indicator strain (bacterial lawn). Left without zone of inhibition (no bacteria-killing activity), right with zone of inhibition (bacteria-killing activity). (C) Detection of hydrogen peroxide with TMB; left, light colony showing no secretion; right, blue colony indicating secretion. (D) Detection of acid production by LAB (MRS agar plates); pH stripes: orange, acidification to pH 4.1; red, no acidification pH 6.1.

Figure 3.
Workflow diagram describing how EFSA Units incorporate the qualified presumption of safety (QPS) status (from Herman et. al. under CC BY 4.0 conditions) [135].
Figure 3.
Workflow diagram describing how EFSA Units incorporate the qualified presumption of safety (QPS) status (from Herman et. al. under CC BY 4.0 conditions) [135].

Figure 4.
Commercially available protective cultures by food application.


{kind=link}
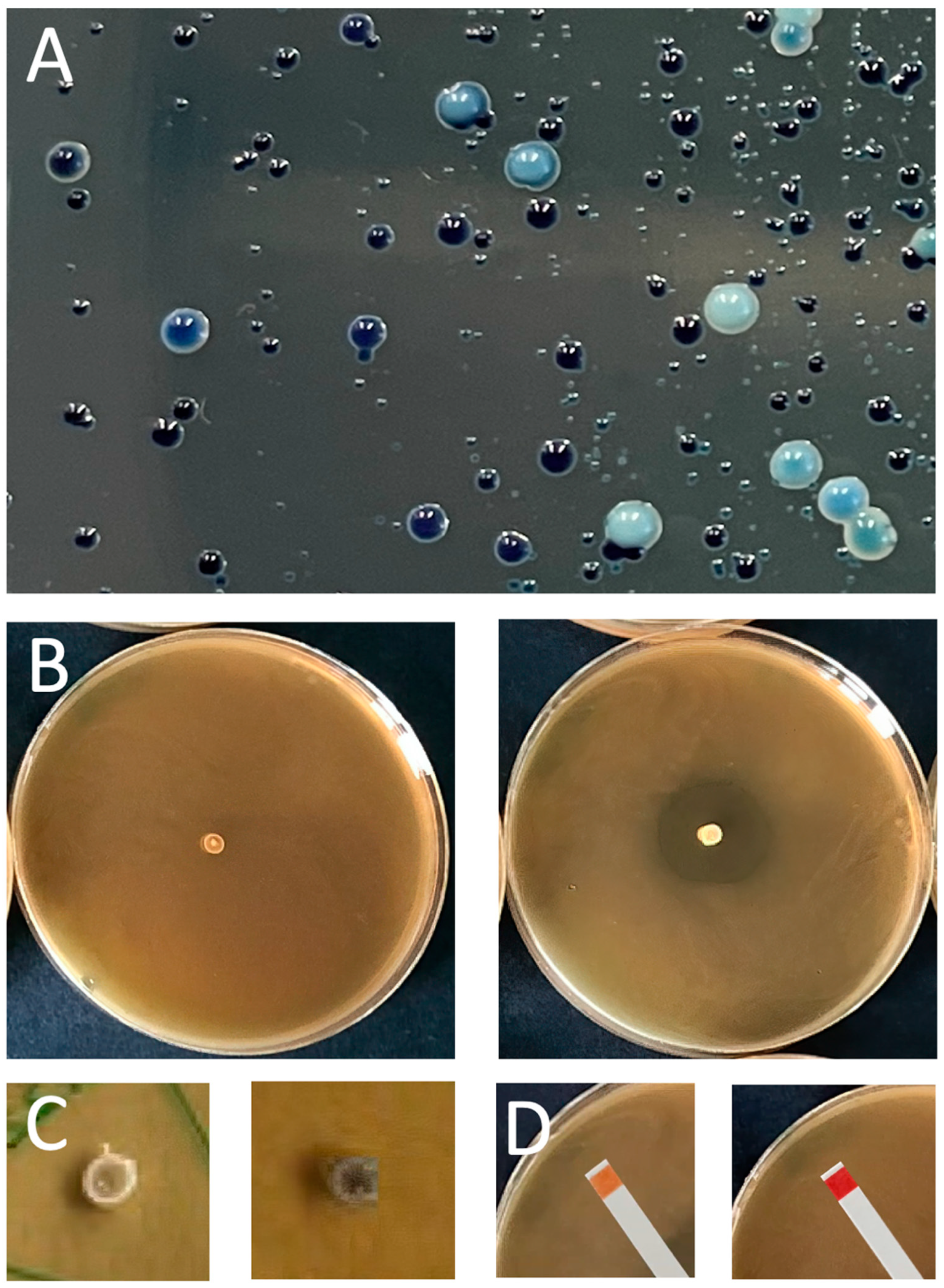{kind=link}
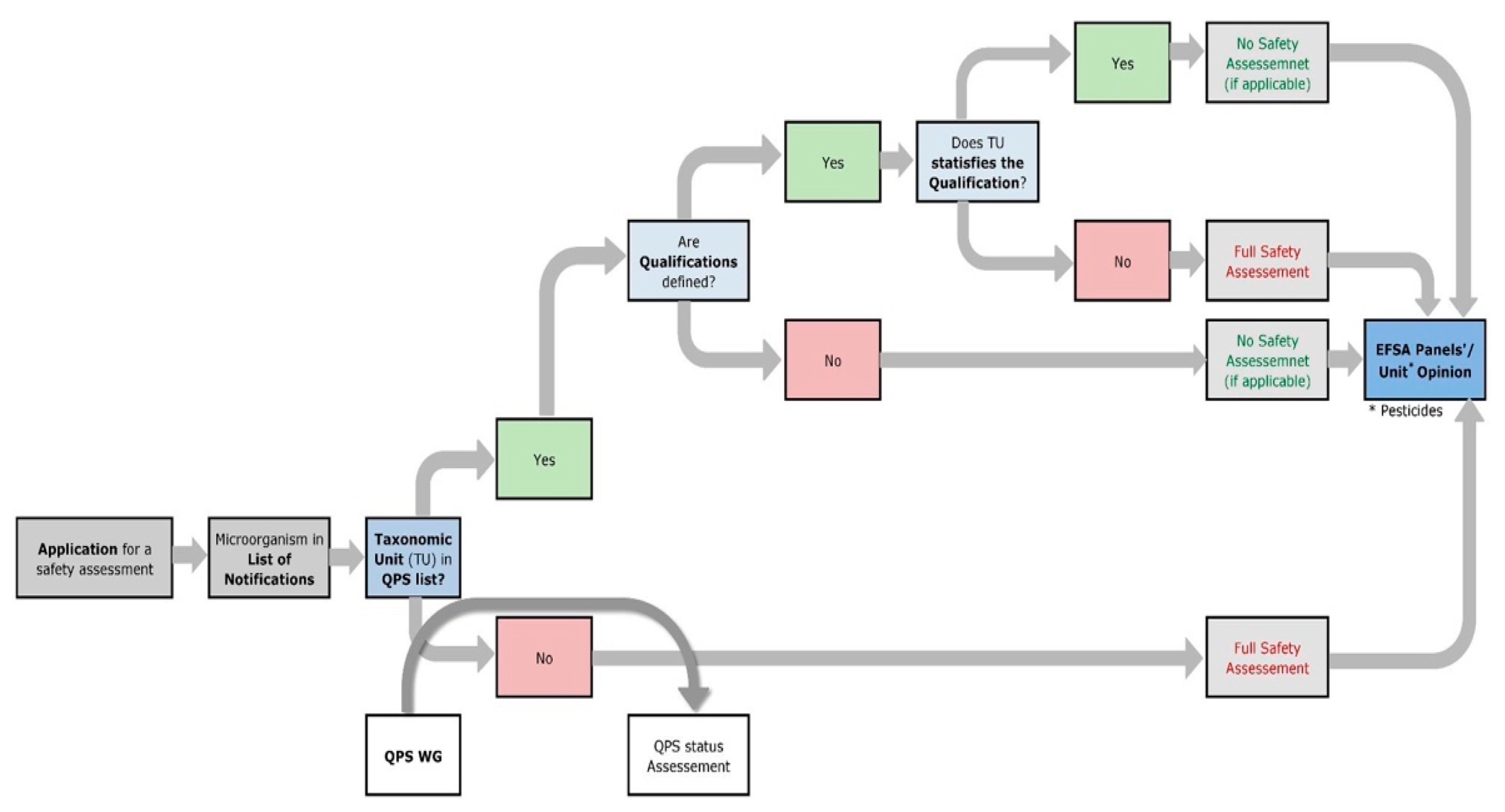{kind=link}
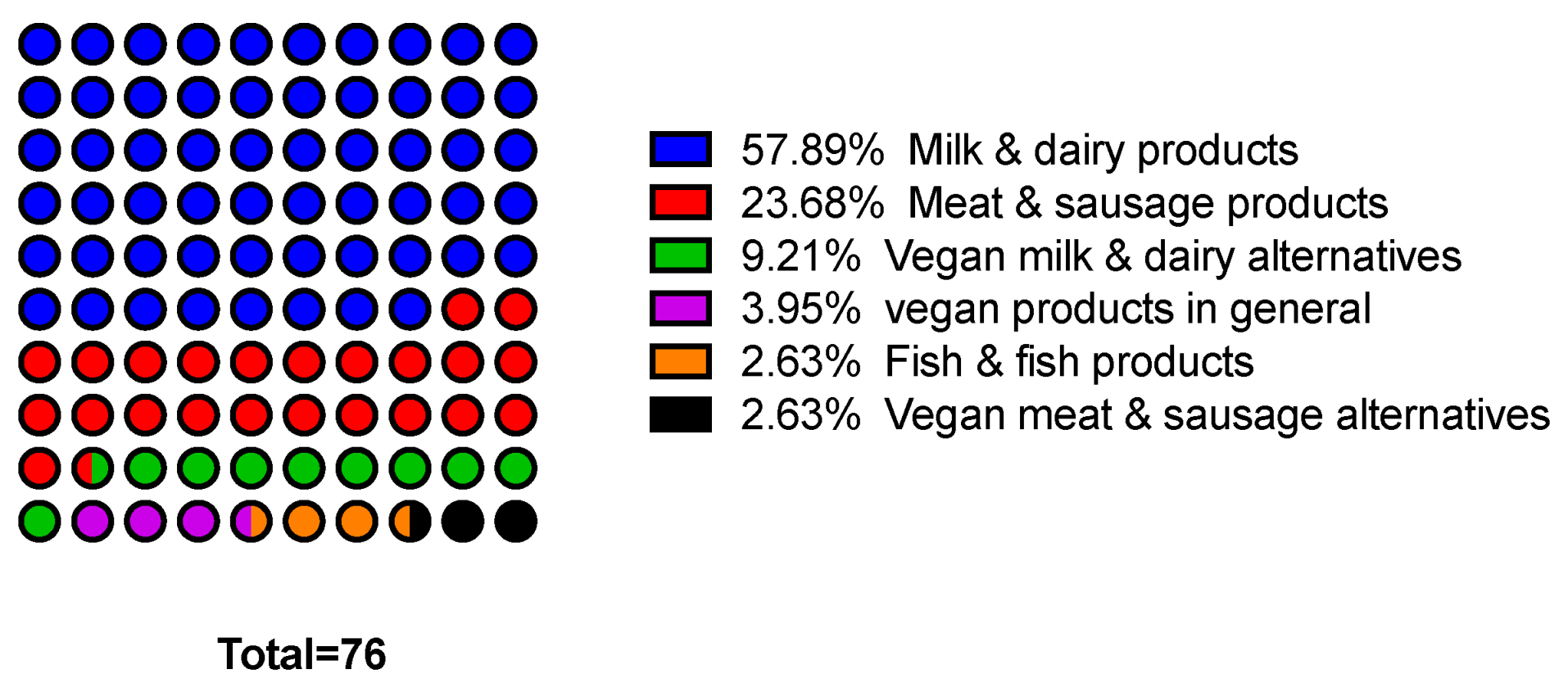{kind=link}
| Ingredient | Source |
|---|---|
| carbohydrate | glucose or lactose (skim)milk or whey cellobiose–xylose mixture) sugar molasses |
| amino acids or peptones | mainly added though peptones, yeast extracts or proteins (milk) |
| fatty acids | tween, acetate |
| nucleosides/nucleotides | yeast extracts, meat |
| minerals | mainly added as salts |
| vitamins | yeast extracts, meat extracts, and peptones |
| citrate | purified dairy-based ingredient |
Table 2.
Allowed addition of nisin in food, natural occurrence in the products not included, source Regulation (EC) 1333/2008.
Table 2.
Allowed addition of nisin in food, natural occurrence in the products not included, source Regulation (EC) 1333/2008.
| Category | Category No. | Maximum Quantity | Restriction/Place of Use |
|---|---|---|---|
| other cream products | 01.6.3 | 10 mg/L bzw. mg/kg | clotted cream products |
| unripened cheese, (except products of category 16) | 01.7.2 | 10 mg/L or mg/kg | mascarpone |
| ripened cheese | 01.7.2 | 12.5 mg/L or mg/kg | |
| processed cheese | 01.7.5 | 12.5 mg/L or mg/kg | |
| cheese products (except category 16) | 01.7.6 | 12.5 mg/L or mg/kg | ripened and processed products only |
| eggs and egg products processed | 10.2 | 6.25 mg/L or mg/kg | pasteurized liquid only (egg white, yolk or whole egg) |
| dessert foods, excluding products of categories 1, 3 and 4 | 16 | 3 mg/L or mg/kg | semolina and tapioca puddings and similar products only |
Table 3.
Differences between the GRAS guidelines (FDA, USA) and the QPS system.
| GRAS | QPS |
|---|---|
| food additives and microorganisms | microorganisms only |
| FDA or external experts determine GRAS status | EFSA determines QPS status |
| open to all types of food additives | limited to microorganisms associated with regulated food and feed products only |
| applicants request a GRAS status | EFSA requests evaluation of new taxonomic units within the scope of an internal mandate |
| describes a specific substance or microorganism at the strain level | describes microorganisms at the taxonomic unit level |
| case-by-case safety assessment at the strain level | general safety assessment at the taxonomic unit level. |
| open tool to all applicants | internal tool by EFSA |
Table 4.
Commercially available protective microbial cultures for various applications.
| Food Application | Product Name | Microorganism | Manufacturer |
|---|---|---|---|
| Fish & fish products | Lyoflora FP-18 | Carnobacterium spp. | SACCO |
| Lyoflora FP-50 | Carnobacterium spp. | SACCO | |
| Meat & sausage products | AVO raw sausage protective culture | Lactobacillus plantarum | AVO-Werke August Beisse GmbH |
| Lyocarni BMX-37 | Carnobacterium spp., Lactobacillus sakei | SACCO | |
| Lyocarni BOM-13 | Carnobacterium spp., Lactobacillus sakei | SACCO | |
| Lyocarni BOX-74 | Carnobacterium spp., Lactobacillus sakei | SACCO | |
| Lyocarni BXH-69 | Carnobacterium spp., Lactobacillus sakei | SACCO | |
| Lyoflora FP-18 | Carnobacterium spp. | SACCO | |
| Lyoflora FP-50 | Carnobacterium spp | SACCO | |
| M-Culture RS 103 | Lactobacillus curvatus subsp. curvatus, Lactobacillus plantarum, Lactobacillus sakei, Staphylococcus carnosus | M-Foodgroup GmbH | |
| M-Culture RS 107 | Lactobacillus curvatus subsp. curvatus, Lactobacillus plantarum, Lactobacillus sakei, Staphylococcus carnosus | M-Foodgroup GmbH | |
| M-Culture RS 38 | Lactobacillus curvatus subsp. curvatus, Lactobacillus plantarum, Pediococcus pentosaceus, Staphylococcus carnosus, Staphylococcus xylosus, Debaryomyces hansenii | M-Foodgroup GmbH | |
| M-Culture RS 49 FM | Debaryomyces hansenii, Lactobacillus sakei, Lactobacillus plantarum | M-Foodgroup GmbH | |
| M-Culture Safe GDS 3349 | Leuconostoc Carnosum | M-Foodgroup GmbH | |
| ProtectSTART | Staphylococcus xylosus, Staphylococcus carnosus, Leuconostoc citreum | Moguntia food group | |
| SaferSTART | Leuconostoc citeum | Moguntia food group | |
| Protective cultures for raw sausage | Lactobacillus sakai, Pediococcus pentosaceus, Staphylococcus xylosus, Staphylococcus carnosus, Debaryomyces hansenii | Würzteufel GmbH | |
| Protective culture Ham-Protect | Lactobacillus plantarum, Staphylococcus carnosus, Staphylococcus xylosus | Würzteufel GmbH | |
| Meat & sausage products | Protective culture PROTECT ONE | Staphylococcus xylosus, Staphylococcus carnosus, Lactobacillus plantarum bac+ | Würzteufel GmbH |
| StartStar SAFE Protective Cultures | unknown culture with Bacteriocin producer | Holkof GmbH | |
| Milk & dairy products | AC line DY 4P13 | unknown mixture of LAB | SACCO |
| AC line LC 4P1 | unknown mixture of LAB | SACCO | |
| AC line LCP 4P2 | unknown mixture of LAB | SACCO | |
| AC line MO K4P04 | unknown mixture of LAB without Lactococcus lactis | SACCO | |
| AC line MO L4P03 | unknown mixture of LAB without Lactococcus lactis | SACCO | |
| AC line MO N4P01 | unknown mixture of LAB with Lactococcus lactis as nisin producer | SACCO | |
| AC line MO N4P02 | unknown mixture of LAB with Lactococcus lactis as nisin producer | SACCO | |
| AL line CNBAL | unkown mixture of LAB with Carnobacterium divergens V41 and Carnobacterium piscicola SF668 | SACCO | |
| AL line LPAL | unknown mixture of LAB | SACCO | |
| AOSM line BGP1 | unknown mixture of LAB | SACCO | |
| AOSM line LR B | unknown mixture of LAB with Lactobacillus rhamnosus | SACCO | |
| AYM line CLP C | unknown mixture of LAB | SACCO | |
| AYM line LPR A | unknown mixture of LAB | SACCO | |
| AYM line LR B | unknown mixture of LAB with Lactobacillus rhamnosus | SACCO | |
| AYM line LR4 PD | unknown mixture of LAB | SACCO | |
| BIOPROX CP63 | Lactobacillus rhamnosus, Lactiplantibacillus plantarum | bioprox | |
| BIOPROX L135 | Lactococcus lactis | bioprox | |
| BIOPROX P 83 | Lactiplantibacillus plantarum | bioprox | |
| BIOPROX P 94 | Lactobacillus plantarum subsp. Plantarum | bioprox | |
| BIOPROX RP 80 | Lactobacillus rhamnosus, Lactiplantibacillus plantarum | bioprox | |
| BIOPROX SP 86 | Limosilactobacillus fermentum | bioprox | |
| BIOPROX Z100 | Lactiplantibacillus plantarum | bioprox | |
| BS-10 | Lactococcus lactis subsp. lactis | CHR Hansen | |
| FreshQ 11 | Lactobacillus rhamnosus | CHR Hansen | |
| FreshQ 2 | Lactobacillus rhamnosus | CHR Hansen | |
| FreshQ 4 | Lactobacillus rhamnosus | CHR Hansen | |
| FreshQ Cheese 1 | Lactobacillus rhamnosus | CHR Hansen | |
| HOLDBAC GP10 | Pediococcus acidilactici | IFF | |
| Milk & dairy products | HOLDBAC GP20 FRO 500DCU | Lactococcus lactis subsp. lactis (nisin producer), Lactobacillus paracasei, Lactobacillus plantarum | IFF |
| HOLDBAC GP21 FRO 500DCU | Lactococcus lactis subsp. lactis (nisin producer), Lactobacillus rhamnosus, Lactobacillus plantarum | IFF | |
| HOLDBAC LC | Lactobacillus rhamnosus | IFF | |
| Holdbac LC Lyo 100 DCU—Schutzkultur | Lactobacillus rhamnosus | DANISCO (IFF) | |
| HOLDBAC Listeria | Lactobacillus plantarum | IFF | |
| Holdbac YM-B | Lactobacillus rhamnosus, Propionibacterium freudenreichii subsp. Shermanii | IFF | |
| Holdbac YM-B LYO 100 DCU—Schutzkultur | Lactobacillus rhamnosus, Propionibacterium freudenreichii subsp. shermanii | DANISCO (IFF) | |
| HOLDBAC YM-C | Lactobacillus paracasei, Propionibacterium freudenreichii subsp. Shermanii | IFF | |
| HOLDBAC YM-C PLUS | Propionibacterium freudenreichii, Lactobacillus paracasei | IFF | |
| HOLDBAC YM-SUSTAIN | Lactobacillus plantarum, Lactobacillus paracasei | IFF | |
| HOLDBAC YM-XPK | Lactobacillus plantarum | IFF | |
| HOLDBAC YM-XPM | Lactobacillus plantarum, Lactobacillus paracasei | IFF | |
| HOLDBAC YM-XTEND | Lactobacillus plantarum, Lactobacillus rhamnosus | IFF | |
| LC|100 DCU | Lactobacillus rhamnosus | DANISCO (IFF) | |
| Pure Appeal 01 | Lactobacillus paracasei, Streptococcus thermophilus | CHR Hansen | |
| YM-B|100 DCU | Lactobacillus rhamnosus, Propionibacterium freudenreichii subsp. Shermanii | DANISCO (IFF) | |
| vegan products in general | HOLDBAC YM-VEGE | Propionibacterium freudenreichii subsp. shermanii, Lactobacillus rhamnosus | IFF |
| Vega FreshQ 101 | Lactobacillus rhamnosus | CHR Hansen | |
| Vega SafePro 01 data | Lactobacillus sakei | CHR Hansen | |
| Vegan meat & sausage alternatives | Safe M-Culture Vegan | Leuconostoc carnosum | M-Foodgroup GmbH |
| Protective culture PROTECT ONE | Staphylococcus xylosus, Staphylococcus carnosus, Lactobacillus plantarum bac+ | Würzteufel GmbH | |
| Vegan milk & dairy alternatives | Lyofast BGP 1 | unknown mixture of LAB | SACCO |
| Lyofast CLP C | unknown mixture of LAB | SACCO | |
| Vegan milk & dairy alternatives | Lyofast CNBAL | unknown mixture of LAB with Carnobacterium divergens V41 and Carnobacterium piscicola SF668 | SACCO |
| Lyofast LPAL | unknown mixture of LAB | SACCO | |
| Lyofast LPR A | unknown mixture of LAB | SACCO | |
| Lyofast BGP 1 | unknown mixture of LAB with Lactobacillus rhamnosus | SACCO | |
| Lyofast CLP C | unknown mixture of LAB | SACCO | |
| Lyofast LR B | unknown mixture of LAB | SACCO | |
| Lyofast LR4 PD | unknown mixture of LAB | SACCO |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fischer, S.W.; Titgemeyer, F. Protective Cultures in Food Products: From Science to Market. Foods 2023, 12, 1541. https://doi.org/10.3390/foods12071541
AMA Style
Fischer SW, Titgemeyer F. Protective Cultures in Food Products: From Science to Market. Foods. 2023; 12(7):1541. https://doi.org/10.3390/foods12071541
Chicago/Turabian StyleFischer, Sebastian W., and Fritz Titgemeyer. 2023. "Protective Cultures in Food Products: From Science to Market" Foods 12, no. 7: 1541. https://doi.org/10.3390/foods12071541
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.