
Valorisation of Wasted Immature Tomato to Innovative Fermented Functional Foods
 ,
,  , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Preparation of LAB Cultures
2.3. In Vitro Gastrointestinal Digestion Model
2.4. Immature Tomato Pulp Fermentation
2.5. Analytical Procedures
2.5.1. Microbial Analysis and the Survival of Viable LAB Cells
2.5.2. Titratable Acidity and Solid Soluble Content
2.5.3. Quantification of Total Phenolic Content and Antioxidant Capacity
2.5.4. Organic Acids and Phenolic Profiles
2.5.5. CIELab Colour
2.5.6. Sensory Analysis
2.6. Statistical Analysis
3. Results and Discussion
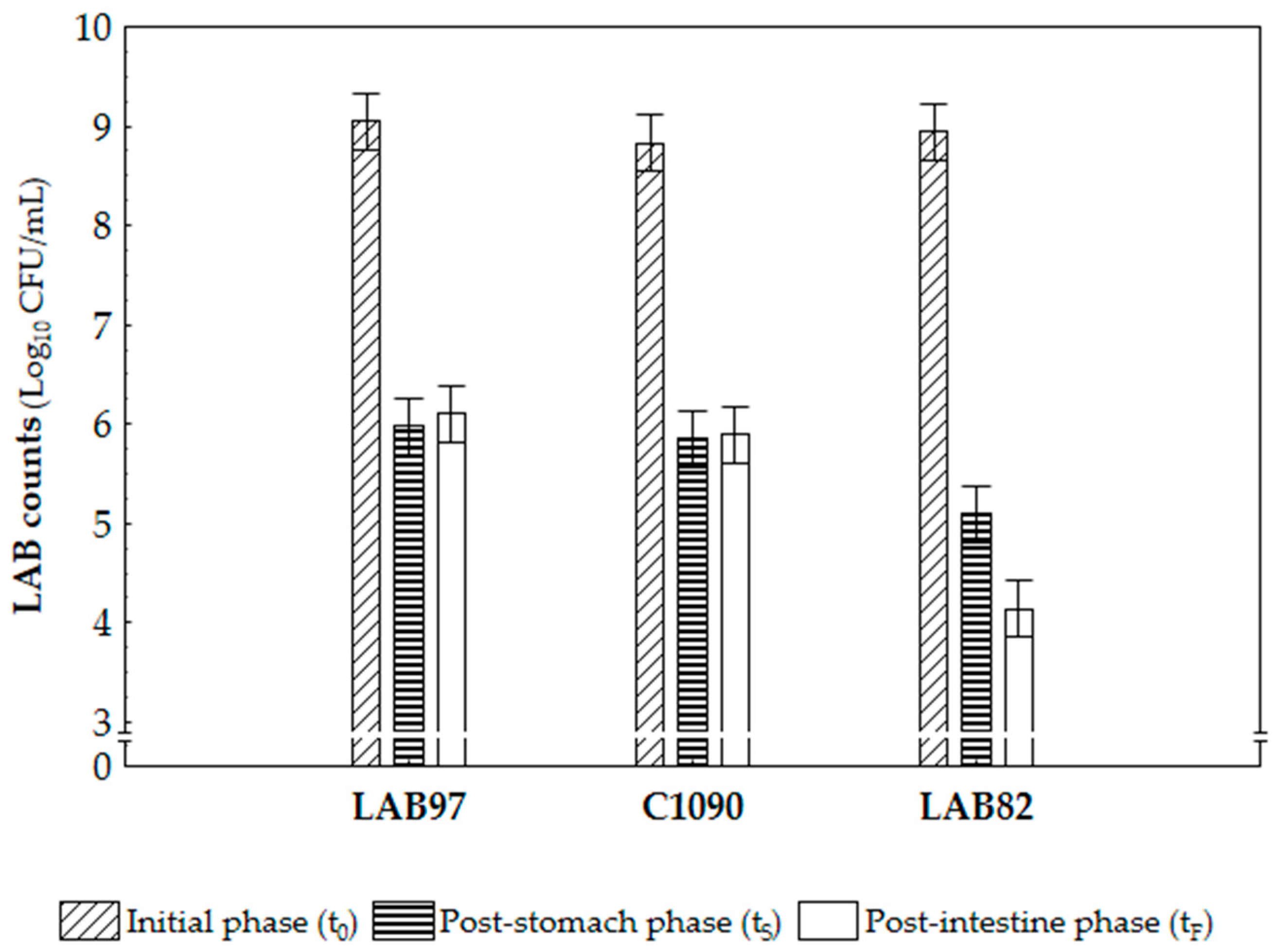
3.1. Probiotic Potential Assessment of Two LAB Cultures
3.2. Quality Assessment of Fermented Immature Tomato Ingredients Inoculated with LAB Strains as Starters
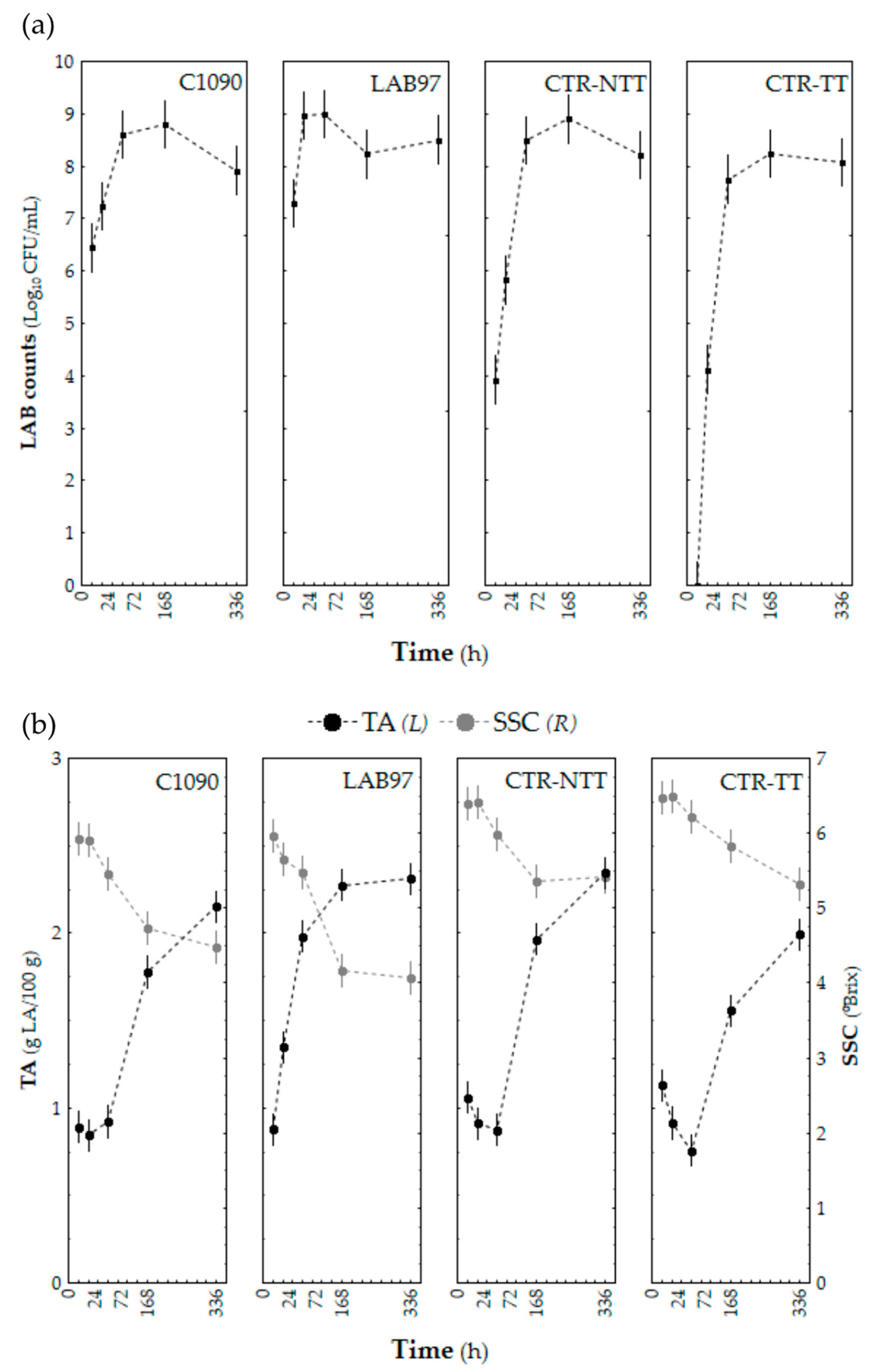
3.2.1. Microbial Growth, Acidification, and Soluble Solids Content during Fermentation
3.2.2. Organic Acids Profile
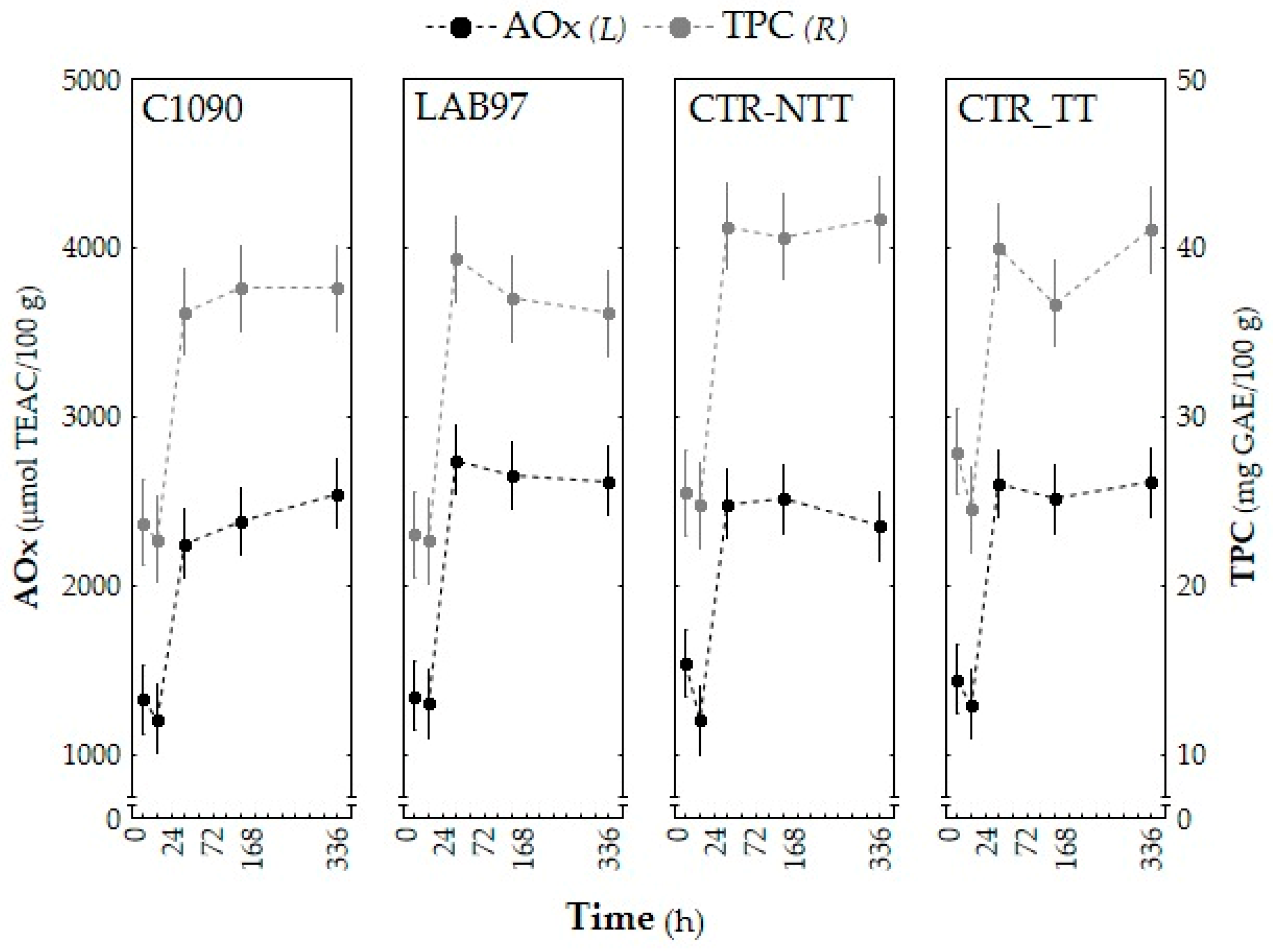
3.2.3. Bioactive Composition (AOx, TPC, and Phenolic Profile)
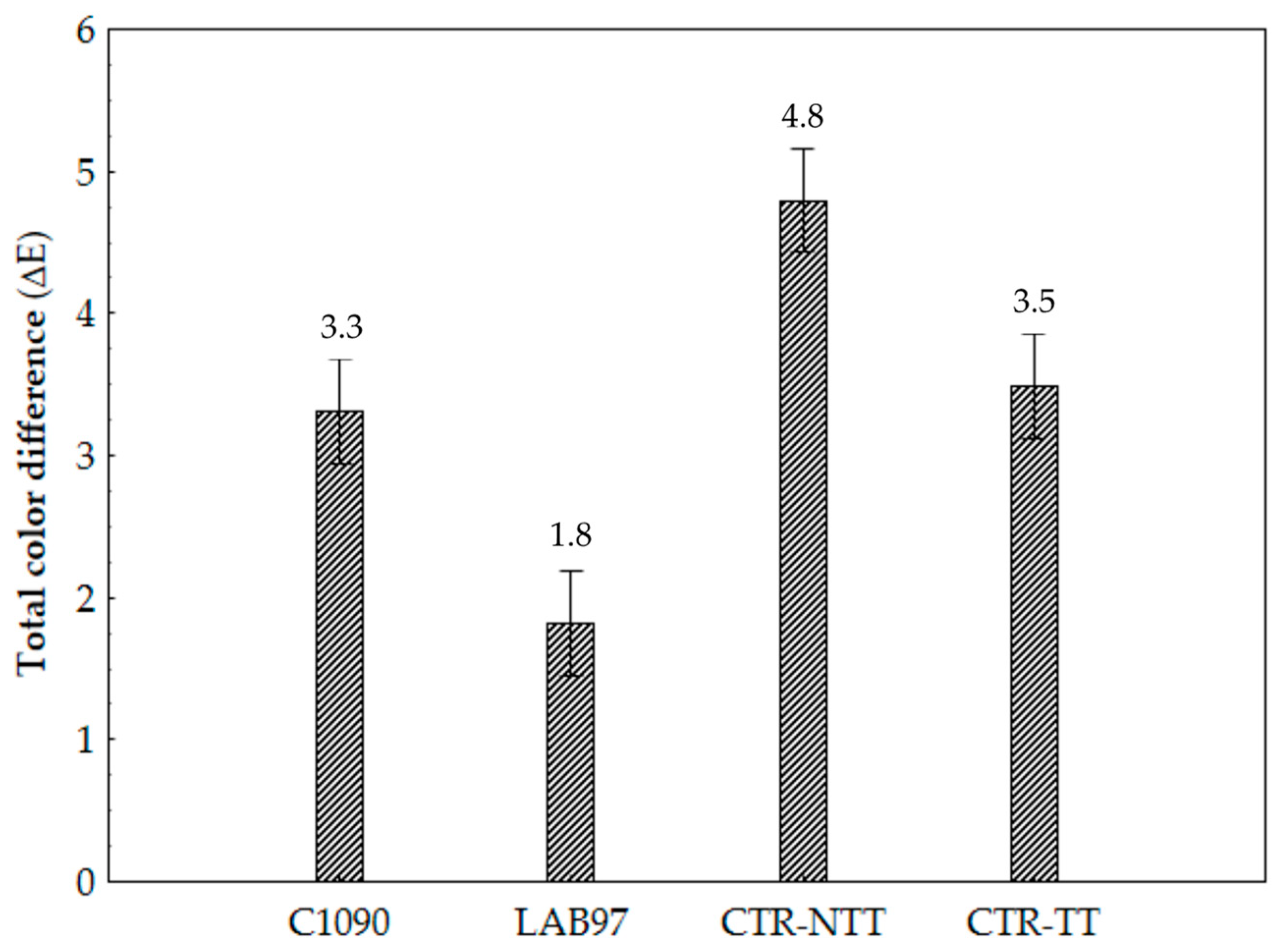
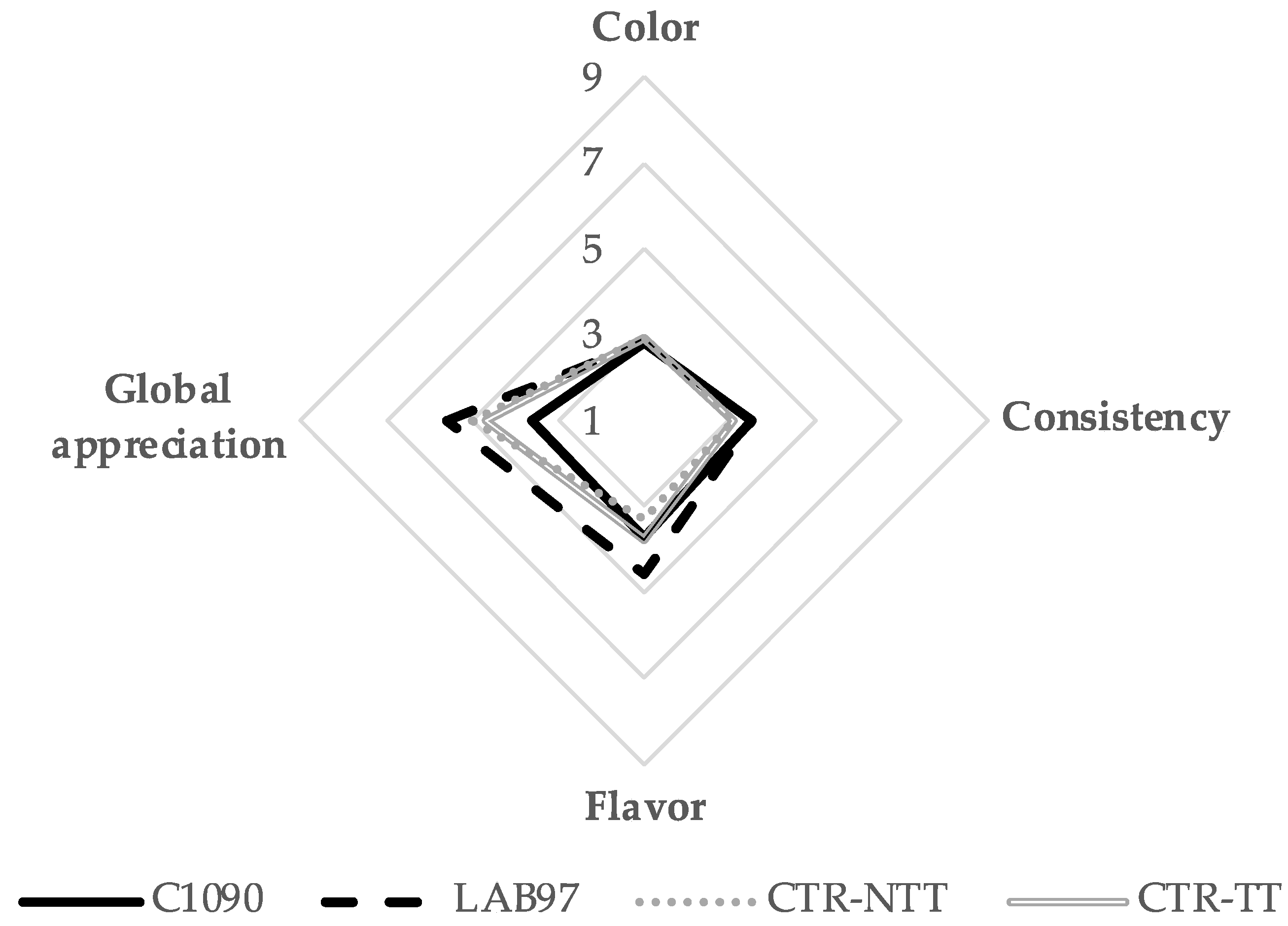
3.2.4. CIELab Colour and Sensory Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Global Food Losses and Food Waste—Extent, Causes and Prevention; FAO: Rome, Italy, 2011. [Google Scholar] [CrossRef]
- Ravindran, R.; Jaiswal, A.K. A comprehensive review on pre-treatment strategy for lignocellulosic food industry waste: Challenges and opportunities. Bioresour. Technol. 2016, 199, 92–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. Global Initiative on Food Loss and Waste Reduction. 2015. Available online: http://www.fao.org/3/a-i4068e.pdf (accessed on 24 October 2022).
- FAOSTAT. Agriculture Holdings Cultivated for the Production of Crops. 2020. Available online: http://www.fao.org/faostat/en/#data/QC/visualize (accessed on 2 December 2022).
- Løvdal, T.; Van Droogenbroeck, B.; Eroglu, E.C.; Kaniszewski, S.; Agati, G.; Verheul, M.; Skipnes, D. Valorization of tomato surplus and waste fractions: A case study using Norway, Belgium, Poland, and Turkey as examples. Foods 2019, 8, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Oliveira, M.F.M.L.; de Cepeda, M.S.; Bernabéu, R. Evolution of the processed tomato sector in Portugal and Spain: Comparative analysis. Emir. J. Food Agric. 2019, 31, 470–476. [Google Scholar] [CrossRef]
- Simões, S.; Santos, R.; Bento-Silva, A.; Santos, M.V.; Mota, M.; Duarte, N.; Sousa, I.; Raymundo, A.; Prista, C. Improving nutritional quality of unripe tomato through fermentation by a consortium of yeast and lactic acid bacteria. J. Sci. Food Agric. 2021, 102, 1422–1429. [Google Scholar] [CrossRef]
- Lu, Z.; Wang, J.; Gao, R.; Ye, F.; Zhao, G. Sustainable valorisation of tomato pomace: A comprehensive review. Trends Food Sci. Technol. 2019, 86, 172–187. [Google Scholar] [CrossRef]
- Santos, R.; Sousa, I.; Raymundo, A.; Silva, J.S.; Prista, C.; Mota, M. Development of a fermented green tomato base for dressings and sauces with high nutritional value. Acta Hortic. 2019, 1233, 239–246. [Google Scholar] [CrossRef]
- Shah, N.N.; Singhal, R.S. Fermented Fruits and Vegetables. In Current Developments in Biotechnology and Bioengineering: Food and Beverages Industry; Elsevier B.V.: Amsterdam, The Netherlands, 2017; pp. 45–89. [Google Scholar] [CrossRef]
- Dahiya, D.; Nigam, P.S. Use of Characterized Microorganisms in Fermentation of Non-Dairy-Based Substrates to Produce Probiotic Food for Gut-Health and Nutrition. Fermentation 2022, 9, 1. [Google Scholar] [CrossRef]
- Szutowska, J. Functional properties of lactic acid bacteria in fermented fruit and vegetable juices: A systematic literature review. Eur. Food Res. Technol. 2020, 246, 357–372. [Google Scholar] [CrossRef]
- Li, Q.; Gänzle, M.G. Host-adapted lactobacilli in food fermentations: Impact of metabolic traits of host adapted lactobacilli on food quality and human health. Curr. Opin. Food Sci. 2020, 31, 71–80. [Google Scholar] [CrossRef]
- Jampaphaeng, K.; Ferrocino, I.; Giordano, M.; Rantsiou, K.; Maneerat, S.; Cocolin, L. Microbiota dynamics and volatilome profile during stink bean fermentation (Sataw-Dong) with Lactobacillus plantarum KJ03 as a starter culture. Food Microbiol. 2018, 76, 91–102. [Google Scholar] [CrossRef]
- Soccol, C.R.; Vandenberghe, L.P.S.; Spier, M.R.; Medeiros, A.B.P.; Yamaguishi, C.T.; De Dea Lindner, J.; Pandey, A.; Thomaz-Soccol, V. The potential of probiotics: A review. Food Technol. Biotechnol. 2010, 48, 413–434. [Google Scholar]
- Terpou, A.; Papadaki, A.; Lappa, I.K.; Kachrimanidou, V.; Bosnea, L.A.; Kopsahelis, N. Probiotics in Food Systems: Significance and Emerging Strategies Towards Improved Viability and Delivery of Enhanced Beneficial Value. Nutrients 2019, 11, 1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, C.; Guerin, M.; Souidi, K.; Remize, F. Lactic fermented fruit or vegetable juices: Past, present and future. Beverages 2020, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Angmo, K.; Kumari, A.; Savitri; Bhalla, T.C. Probiotic characterization of lactic acid bacteria isolated from fermented foods and beverage of Ladakh. LWT-Food Sci. Technol. 2016, 66, 428–435. [Google Scholar] [CrossRef]
- Pereira, N.; Alegria, C.; Aleixo, C.; Martins, P.; Gonçalves, E.M.; Abreu, M. Selection of autochthonous LAB strains of unripe green tomato towards the production of highly nutritious lacto-fermented ingredients. Foods 2021, 10, 2916. [Google Scholar] [CrossRef]
- Fernández De Palencia, P.; López, P.; Corbí, A.L.; Peláez, C.; Requena, T. Probiotic strains: Survival under simulated gastrointestinal conditions, in vitro adhesion to Caco-2 cells and effect on cytokine secretion. Eur. Food Res. Technol. 2008, 227, 1475–1484. [Google Scholar] [CrossRef] [Green Version]
- Moreira, L. Produção de Azeitona de Mesa ao Natural Fermentada por Estirpes de Bactérias Lácticas Potencialmente Probióticas. Master’s Thesis, Instituto Superior de Agronomia da Universidade de Lisboa, Lisboa, Portugal, 2013. [Google Scholar]
- ISO 15214:1998; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Emumeration of Mesophilic Lactic Acid Bacteria—Colony-Count Technique at 30 °C. ISO: Geneva, Switzerland, 1998.
- ISO 21527-1:2008; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Yeasts and Moulds—Part 1: Colony Count Technique in Products with Water Activity Greater than 0, 95. ISO: Geneva, Switzerland, 2008.
- NP 1421; Foodstuffs Derived from Fruits and Vegetables: Determination of Acidity. Instituto Português da Qualidade: Lisbon, Portugal, 1977.
- Alegria, C.; Gonçalves, E.M.; Moldão-Martins, M.; Abreu, M. Influence of a heat-shock pre-treatment on wound-induced phenolic biosynthesis as an alternative strategy towards fresh-cut carrot processing. Food Sci. Technol. Int. 2021, 28, 421–429. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Panda, A.; Alvarenga, N.; Silva, J.L.; Partidário, A.; Lageiro, M.; Roseiro, C.; Dias, J. Influence of cocoa origin on the nutritional characterization of chocolate. Eur. Food Res. Technol. 2022, 248, 2569–2577. [Google Scholar] [CrossRef]
- Petitjean-Freytet, C.; Carnat, A.; Lamaison, J. Teneurs en flavonoides et en dérivés hydroxycinnamiques de la fleur de Sambucus nigra L. J. Pharm. Belg. 1991, 46, 241–246. [Google Scholar]
- ISO 8586-1:1993; Sensory Analysis—General Guidance for the Selection, Training and Monitoring of Assessors. ISO: Geneva, Switzerland, 1993.
- ISO 13299:2016; Sensory Analysis—Methodology—General Guidance for Establishing a Sensory Profile. ISO: Geneva, Switzerland, 2016.
- Lim, J. Hedonic scaling: A review of methods and theory. Food Qual. Prefer. 2011, 22, 733–747. [Google Scholar] [CrossRef]
- StatSoft Inc. STATISTICA (Data Analysis Software System); Version 8.0; StatSoft Inc.: Tulsa, OK, USA, 2007. [Google Scholar]
- Sun, Y.; Zhang, S.; Li, H.; Zhu, J.; Liu, Z.; Hu, X.; Yi, J. Assessments of Probiotic Potentials of Lactiplantibacillus plantarum Strains Isolated from Chinese Traditional Fermented Food: Phenotypic and Genomic Analysis. Front. Microbiol. 2022, 13, 895132. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.; Sunita; Shukla, P. Probiotic potential of Weissella paramesenteroides MYPS5.1 isolated from customary dairy products and its therapeutic application. 3 Biotech 2022, 12, 9. [Google Scholar] [CrossRef] [PubMed]
- Paula, A.T.; Jeronymo-Ceneviva, A.B.; Silva, L.F.; Todorov, S.D.; Franco, B.D.G.M.; Penna, A.L.B. Leuconostoc mesenteroides SJRP55: A potential probiotic strain isolated from Brazilian water buffalo mozzarella cheese. Ann Microbiol. 2014, 65, 899–910. [Google Scholar] [CrossRef]
- Sathyapriya, A.; Anitha, A. Assessment of Probiotic Potential Leuconostoc mesenteroides from Natural Microbiota. Int. J. Sci. Res. Biol. Sci. 2019, 5, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Abushelaibi, A.; Al-Mahadin, S.; El-Tarabily, K.; Shah, N.P.; Ayyash, M. Characterization of potential probiotic lactic acid bacteria isolated from camel milk. LWT-Food Sci. Technol. 2017, 79, 316–325. [Google Scholar] [CrossRef]
- Bah, A.; Ferjani, R.; Fhoula, I.; Gharbi, Y.; Najjari, A.; Boudabous, A.; Ouzari, H.I. Microbial community dynamic in tomato fruit during spontaneous fermentation and biotechnological characterization of indigenous lactic acid bacteria. Ann. Microbiol. 2018, 69, 41–49. [Google Scholar] [CrossRef]
- Bartkiene, E.; Vidmantiene, D.; Juodeikiene, G.; Viskelis, P.; Urbonaviciene, D. Lactic Acid Fermentation of Tomato: Effects on cis/trans Lycopene Isomer Ratio, β-Carotene Mass Fraction and Formation of L(+)- and D(–)-Lactic Acid. Food Technol. Biotechnol. 2013, 51, 471–478. [Google Scholar]
- Alberto, M.R.; Perera, M.F.; Arena, M.E. Lactic Acid Fermentation of Peppers. Food Nutr. Sci. 2013, 4, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Bartkiene, E.; Juodeikiene, G.; Zadeike, D.; Viskelis, P. The Use of Tomato Powder Fermented with Pediococcus pentosaceus and Lactobacillus sakei for the Ready-to-Cook Minced Meat Product Quality Improvement. Food Technol. Biotechnol. 2015, 53, 163–170. [Google Scholar] [CrossRef]
- Di Cagno, R.; Filannino, P.; Gobbetti, M. Vegetable and Fruit Fermentation by Lactic Acid Bacteria. In Biotechnology of Lactic Acid Bacteria: Novel Applications, 2nd ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016; pp. 216–230. [Google Scholar]
- Di Cagno, R.; Minervini, G.; Rizzello, C.G.; De Angelis, M.; Gobbetti, M. Effect of lactic acid fermentation on antioxidant, texture, color and sensory properties of red and green smoothies. Food Microbiol. 2011, 28, 1062–1071. [Google Scholar] [CrossRef]
- Agius, C.; von Tucher, S.; Poppenberger, B.; Rozhon, W. Quantification of sugars and organic acids in tomato fruits. MethodsX 2018, 5, 537–550. [Google Scholar] [CrossRef]
- Panda, S.H.; Ray, R.C. Lactic acid fermentation of β-carotene rich sweet potato (Ipomoea batatas L.) into lacto-juice. Plant Foods Hum. Nutr. 2007, 62, 65–70. [Google Scholar] [CrossRef]
- Panda, S.H.; Parmanick, M.; Ray, R.C. Lactic acid fermentation of sweet potato (Ipomoea Batatas L.) into pickles. J. Food Process. Preserv. 2007, 31, 83–101. [Google Scholar] [CrossRef]
- Gomes, R.J.; Borges, M.F.; Rosa, M.F.; Castro-Gómez, R.J.H.; Spinosa, W.A. Acetic acid bacteria in the food industry: Systematics, characteristics and applications. Food Technol. Biotechnol. 2018, 56, 139–151. [Google Scholar] [CrossRef]
- Torres, C.A.; Romero, L.A.; Diaz, R.I. Quality and sensory attributes of apple and quince leathers made without preservatives and with enhanced antioxidant activity. LWT 2015, 62, 996–1003. [Google Scholar] [CrossRef]
- Adebo, O.A.; aMedina-Meza, I.G. Impact of fermentation on the phenolic compounds and antioxidant activity of whole cereal grains: A mini review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Zhou, J.; Fan, L.; Qin, Z.; Chen, Q.; Zhao, L. Antioxidant properties of a vegetable–fruit beverage fermented with two Lactobacillus plantarum strains. Food Sci. Biotechnol. 2018, 27, 1719–1726. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Chen, R.; Chen, W.; Chen, H.; Zhang, G. Comparative Evaluation of the Antioxidant Capacities, Organic Acids, and Volatiles of Papaya Juices Fermented by Lactobacillus acidophilus and Lactobacillus plantarum. J. Food Qual. 2018, 2018, 9490435. [Google Scholar] [CrossRef] [Green Version]
- Kachouri, F.; Ksontini, H.; Kraiem, M.; Setti, K.; Mechmeche, M.; Hamdi, M. Involvement of antioxidant activity of Lactobacillus plantarum on functional properties of olive phenolic compounds. J. Food Sci. Technol. 2015, 52, 7924–7933. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chen, H.; Chen, W.; Zhong, Q.; Zhang, G.; and Chen, W. Beneficial effects of tomato juice fermented by Lactobacillus plantarum and Lactobacillus casei: Antioxidation, antimicrobial effect, and volatile profiles. Molecules 2018, 23, 2366. [Google Scholar] [CrossRef] [Green Version]
- Panda, S.K.; Behera, S.K.; Witness Qaku, X.; Sekar, S.; Ndinteh, D.T.; Nanjundaswamy, H.M.; Ray, R.C.; Kayitesi, E. Quality enhancement of prickly pears (Opuntia sp.) juice through probiotic fermentation using Lactobacillus fermentum—ATCC 9338. LWT-Food Sci. Technol. 2016, 75, 453–459. [Google Scholar] [CrossRef]
- Ricci, A.; Marrella, M.; Hadj Saadoun, J.; Bernini, V.; Godani, F.; Dameno, F.; Neviani, E.; Lazzi, C. Development of lactic acid-fermented tomato products. Microorganisms 2020, 8, 1192. [Google Scholar] [CrossRef] [PubMed]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M.T.; Wu, M.; Sackey, A.S.; Xiao, L.; Tahir, H.E. Effect of Lactobacillus strains on phenolic profile, color attributes and antioxidant activities of lactic-acid-fermented mulberry juice. Food Chem. 2018, 250, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Hur, S.J.; Lee, S.Y.; Kim, Y.C.; Choi, I.; Kim, G.B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, H.; Curiel, J.A.; Landete, J.M.; de las Rivas, B.; de Felipe, F.L.; Gómez-Cordovés, C.; Mancheño, J.M.; Muñoz, R. Food phenolics and lactic acid bacteria. Int. J. Food Microbiol. 2009, 132, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Chu, S.C.; Chen, C. Effects of origins and fermentation time on the antioxidant activities of kombucha. Food Chem. 2006, 98, 502–507. [Google Scholar] [CrossRef]
- Vallverdú-Queralt, A.; Jáuregui, O.; Medina-Rémon, A.; Andrés-Lacueva, C.; Lamuela-Raventós, R.M. Improved characterization of tomato polyphenols using liquid chromatography/electrospray ionization linear ion trap quadrupole Orbitrap mass spectrometry and liquid chromatography/electrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 2986–2992. [Google Scholar] [CrossRef]
- Chaudhary, P.; Sharma, A.; Singh, B.; Nagpal, A.K. Bioactivities of phytochemicals present in tomato. J. Food Sci. Technol. 2018, 55, 2833–2849. [Google Scholar] [CrossRef] [PubMed]
- Hamdan, N.; Lee, C.H.; Wong, S.L.; Fauzi, C.E.; Zamri, N.M.A.; Lee, T.H. Prevention of Enzymatic Browning by Natural Extracts and Genome-Editing: A Review on Recent Progress. Molecules 2022, 27, 1101. [Google Scholar] [CrossRef]
- Fonseca, H.C.; Sousa Melo, D.; Ramos, C.L.; Dias, D.R.; Schwan, R.F. Probiotic Properties of Lactobacilli and Their Ability to Inhibit the Adhesion of Enteropathogenic Bacteria to Caco-2 and HT-29 Cells. Probiotics Antimicrob. Proteins 2021, 13, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Salmerón, I.; Thomas, K.; Pandiella, S.S. Effect of potentially probiotic lactic acid bacteria on the physicochemical composition and acceptance of fermented cereal beverages. J. Funct. Foods 2015, 15, 106–115. [Google Scholar] [CrossRef]
- Todorov, S.D.; Franco, B.D.G. Lactobacillus plantarum: Characterization of the species and application in food production. Food Rev. Int. 2010, 26, 205–229. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Time (h) | Oxalic Acid (mg/100 g) | Citric Acid (mg/100 g) | Lactic Acid (mg/100 g) | Acetic Acid (mg/100 g) |
|---|---|---|---|---|---|
| C1090 | 0 | 13.3 ± 1.3 bcd | 945.7 ± 56.0 def | 0.0 ± 0.0 a | 0.0 ± 0.0 a |
| 24 | 12.9 ± 0.2 bcd | 886.0 ± 121.9 cde | 0.0 ± 0.0 a | 68.2 ± 13.6 abcd | |
| 72 | 12.3 ± 1.1 abcd | 711.1 ± 141.3 c | 212.9 ± 15.2 bc | 125.3 ± 4.2 bcde | |
| 168 | 14.4 ± 0.7 cde | 125.5 ± 24.2 a | 485.3 ± 49.6 ef | 141.6 ± 23.4 cdef | |
| 336 | 12.6 ± 0.9 abcd | 77.3 ± 23.1 a | 526.0 ± 110.2 efg | 157.6 ± 35.8 defg | |
| LAB97 | 0 | 12.0 ± 1.4 abcd | 907.0 ± 114.9 cdef | 0.0 ± 0.0 a | 0.0 ± 0.0 a |
| 24 | 13.7 ± 0.2 bcde | 981.8 ± 17.3 ef | 258.6 ± 2.3 c | 0.0 ± 0.0 a | |
| 72 | 11.6 ± 1.2 ab | 863.1 ± 46.8 cde | 413.1 ± 13.1 de | 29.3 ± 1.8 ab | |
| 168 | 16.3 ± 0.9 e | 954.2 ± 28.6 def | 579.5 ± 25.2 fg | 64.6 ± 6.7 abcd | |
| 336 | 13.1 ± 0.6 bcd | 758.9 ± 17.5 cd | 525.1 ± 0.9 efg | 37.8 ± 5.5 abc | |
| CTR-NTT | 0 | 12.0 ± 0.9 abcd | 973.7 ± 97.7 ef | 0.0 ± 0.0 a | 0.0 ± 0.0 a |
| 24 | 12.1 ± 0.2 abcd | 1089.7 ± 2.5 f | 0.0 ± 0.0 a | 28.9 ± 5.6 ab | |
| 72 | 12.1 ± 0.0 abcd | 83.2 ± 16.3 a | 194.4 ± 6.6 bc | 191.9 ± 10.5 efg | |
| 168 | 10.1 ± 0.1 a | 33.2 ± 1.6 a | 424.4 ± 47.6 de | 212.9 ± 7.6 efg | |
| 336 | 11.7 ± 1.3 abc | 88.1 ± 8.3 a | 607.9 ± 12.0 g | 259.0 ± 13.5 g | |
| CTR-TT | 0 | 13.8 ± 0.3 bcde | 934.3 ± 128.8 def | 0.0 ± 0.0 a | 0.0 ± 0.0 a |
| 24 | 14.5 ± 0.2 de | 1017.8 ± 29.3 ef | 0.0 ± 0.0 a | 117.9 ± 6.6 bcde | |
| 72 | 13.8 ± 1.0 bcde | 498.0 ± 7.4 b | 122.4 ± 14.0 b | 148.0 ± 30.6 def | |
| 168 | 12.6 ± 1.3 abcd | 135.0 ± 24.6 a | 309.2 ± 65.0 cd | 241.1 ± 88.5 fg | |
| 336 | 11.8 ± 0.9 abc | 46.6 ± 4.5 a | 469.0 ± 91.1 ef | 258.8 ± 110.1 g |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, N.; Farrokhi, M.; Vida, M.; Lageiro, M.; Ramos, A.C.; Vieira, M.C.; Alegria, C.; Gonçalves, E.M.; Abreu, M. Valorisation of Wasted Immature Tomato to Innovative Fermented Functional Foods. Foods 2023, 12, 1532. https://doi.org/10.3390/foods12071532
Pereira N, Farrokhi M, Vida M, Lageiro M, Ramos AC, Vieira MC, Alegria C, Gonçalves EM, Abreu M. Valorisation of Wasted Immature Tomato to Innovative Fermented Functional Foods. Foods. 2023; 12(7):1532. https://doi.org/10.3390/foods12071532
Chicago/Turabian StylePereira, Nelson, Mahsa Farrokhi, Manuela Vida, Manuela Lageiro, Ana Cristina Ramos, Margarida C. Vieira, Carla Alegria, Elsa M. Gonçalves, and Marta Abreu. 2023. "Valorisation of Wasted Immature Tomato to Innovative Fermented Functional Foods" Foods 12, no. 7: 1532. https://doi.org/10.3390/foods12071532