Quality Control of Jinhua Ham from the Influence between Proteases Activities and Processing Parameters: A Review
by
 , and
, and
Shiqi Hu
1,2,3 ,
,
Xinglian Xu
1,2,3,
Wangang Zhang
1,2,3,
Chunbao Li
1,2,3 and
Guanghong Zhou
1,2,3,* 1
Key Laboratory of Meat Processing and Quality Control, Ministry of Education, Nanjing 210095, China
2
Key Laboratory of Meat Processing, Ministry of Agriculture and Rural Affairs, Nanjing 210095, China
3
Jiangsu Collaborative Innovation Center of Meat Production, Processing and Quality Control, College of Food Science and Technology, Nanjing Agricultural University, Nanjing 210095, China
*
Author to whom correspondence should be addressed.
Foods 2023, 12(7), 1454; https://doi.org/10.3390/foods12071454
Submission received: 14 February 2023
/
Revised: 21 March 2023
/
Accepted: 25 March 2023
/
Published: 29 March 2023
(This article belongs to the Section Meat)
Abstract
:Endogenous proteases are significant for Jinhua ham quality. Protein degradation affects the chemical traits, texture and the formation of flavor substances. Protease activities are affected by different process parameters, such as processing temperature, maturation time, salt content and the drying rate. They affect ham quality, which can be controlled by process parameters. The influences of key factors on Jinhua ham quality are briefly summarized, which can provide a theoretical basis for the selection of specific parameters in dry-cured ham processing. Furthermore, some suggestions are proposed for correcting and improving the flavor and textural defects of ham, yet the effectiveness depends on the operating conditions. The determination of enzyme activity is not real-time and unsupervised at the moment. Future research will focus on the determination of the actual endogenous protease activity and the quantitative relationship between the enzyme activity and main processing parameters.
1. Introduction
There are various types of dry-cured ham, and the ripening process is critical for flavor development. Among them, Jinhua ham is an important part of traditional Chinese meat products with a long history. Likewise, Parma ham from Italy, Iberian ham, and Serrano ham from Spain are also globally recognized. The unique production process for each type of dry-cured ham contributes to their characteristic color, texture and flavor [1]. In the modern procedure of Jinhua ham, salting is carried out by adding 6 g of NaCl per kilogram of raw ham. This is followed by processes such as washing, air-drying, ripening, and post-ripening [2]. It requires a few months, while the processing time for ham from Italy and Spain is even longer. For instance, Iberian dry-cured ham needs 18–24 months for manufacturing. Furthermore, compared to Parma ham and Iberian ham, the salt content is high in the finished products of Jinhua ham. It reaches 12%, while Parma ham is 4.2–6.2% and Iberian ham is around 6.5% [3,4]. It is worth noting that proteolysis is critical to the formation of ham flavor and texture, as demonstrated by numerous studies. This process is primarily induced by muscle endogenous proteases [5,6].
Protease activities are affected by a variety of factors, and the changes in either intrinsic or extrinsic factors have a probable effect on endogenous protease activity [7]. Zhao et al. [8] recorded the changes of cathepsin B and L activities during Jinhua ham processing, and they revealed that temperature, pH value and salt content had significant effects on them. Zhou et al. [9] found that sodium chloride had a significant inhibitory effect on cathepsins activities, and Jinhua ham with reduced salt exhibited higher sensory scores. Moreover, the literature from Zhou et al. [10] showed intenser taste and richer attributes in modern-processed Jinhua ham, and the activity of endogenous protease was found to vary with changes in ambient temperature and relative humidity. Arnau et al. [11] studied the addition of lipases and proteinases during the process of fermented sausages. Based on the above description, it can be found that a comprehensive understanding of the influence between proteases activities and processing parameters is necessary to optimize Jinhua ham processing [12,13]. Internal factors affecting protease activity in ham include pH level, weight, and fat content, while extrinsic factors are composed of processing temperature, maturation time, salt content, and the use of new equipment. The changes in these factors are of great practical significance in predicting and regulating the activity of endogenous protease [14,15].
The processing technology and competitiveness of Jinhua ham are considered to be inadequate when compared to Italian and Spanish hams. Currently, there is a significant amount of literature exploring salt reduction, technological modernization and the time saved in the production [16,17]. It is important to note that the activities of muscle endogenous proteases are influenced by multiple factors. However, there is a lack of information available regarding how to improve ham quality through the influence between protease activity and processing parameters. This paper summarizes the contribution of endopeptidases and exopeptidases to Jinhua ham flavor and texture, and discusses how processing parameters affect protease activity. The aim of this study was to explore the relationship between endogenous protease activity, processing parameters, and ham quality. The ultimate goal was to identify the factors that influence protease activity and optimize processing conditions to achieve higher quality products.
The rest of this paper is structured as follows: Section 2 provides information about the different endogenous proteases in Jinhua ham, including calpains, cathepsins, dipeptidases, tripeptidases and aminopeptidases. In Section 3, the contribution of endogenous proteases to the texture and flavor of Jinhua ham is clarified. Additionally, Section 4 shows the association between enzyme activity and ham processing parameters. Various influencing factors are involved, such as raw material, processing temperature, NaCl content, processing time and the application of non-destructive techniques. Finally, the conclusion of this paper is presented in Section 5.
2. Compositions and Characteristics of Proteases
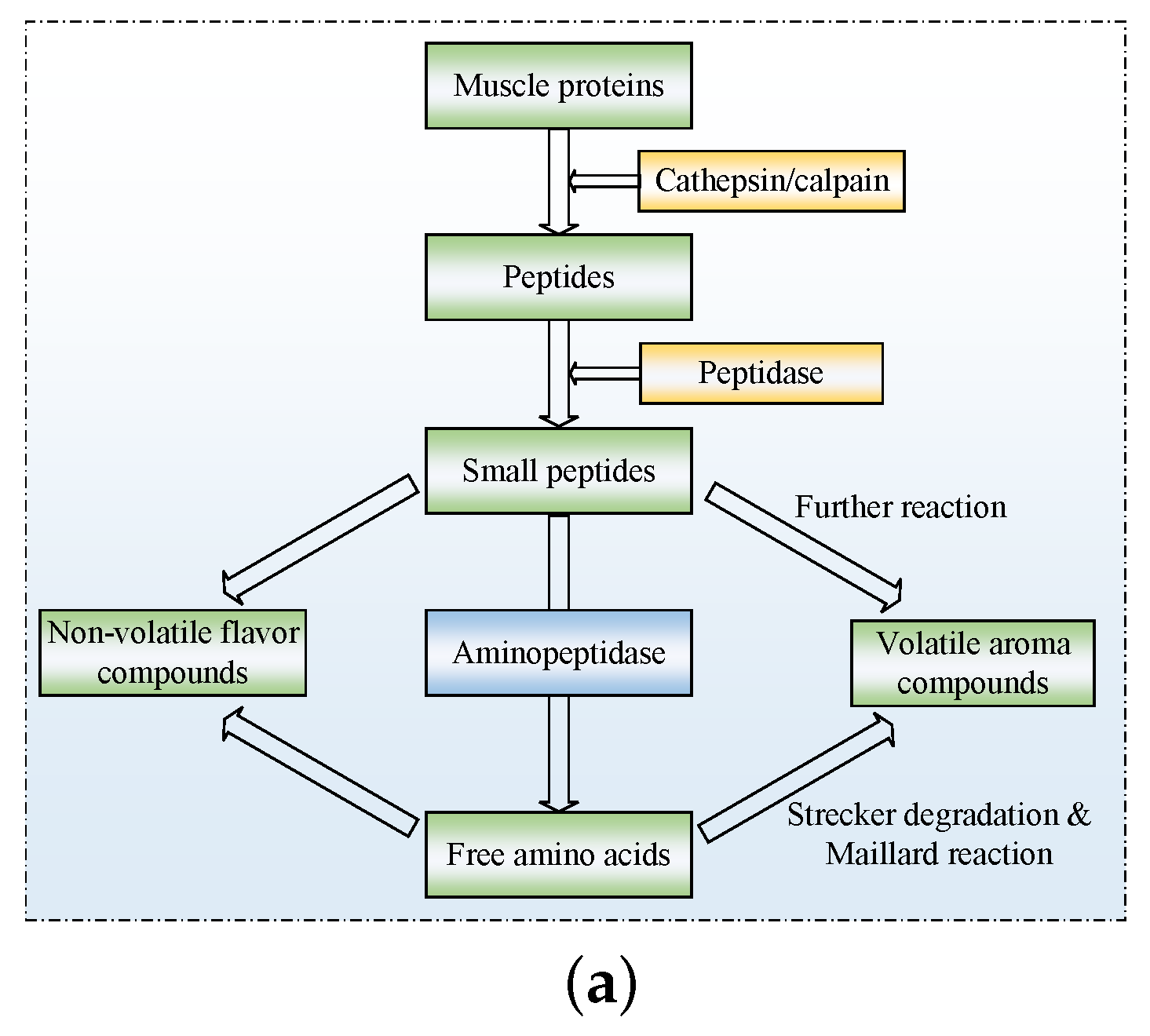
The section provides an overview of different endogenous proteases in Jinhua ham muscle. Figure 1a shows their role in the degradation process of muscle protein, explaining the formation of flavor substances. Firstly, the skeleton proteins are degraded by calpains and cathepsins. Then, the peptides produced above are degraded into small molecule peptides under the further action of dipeptidases and tripeptidases. Ultimately, they are degraded into free amino acids by aminopeptidases and carboxypeptidases.
2.1. Calpains
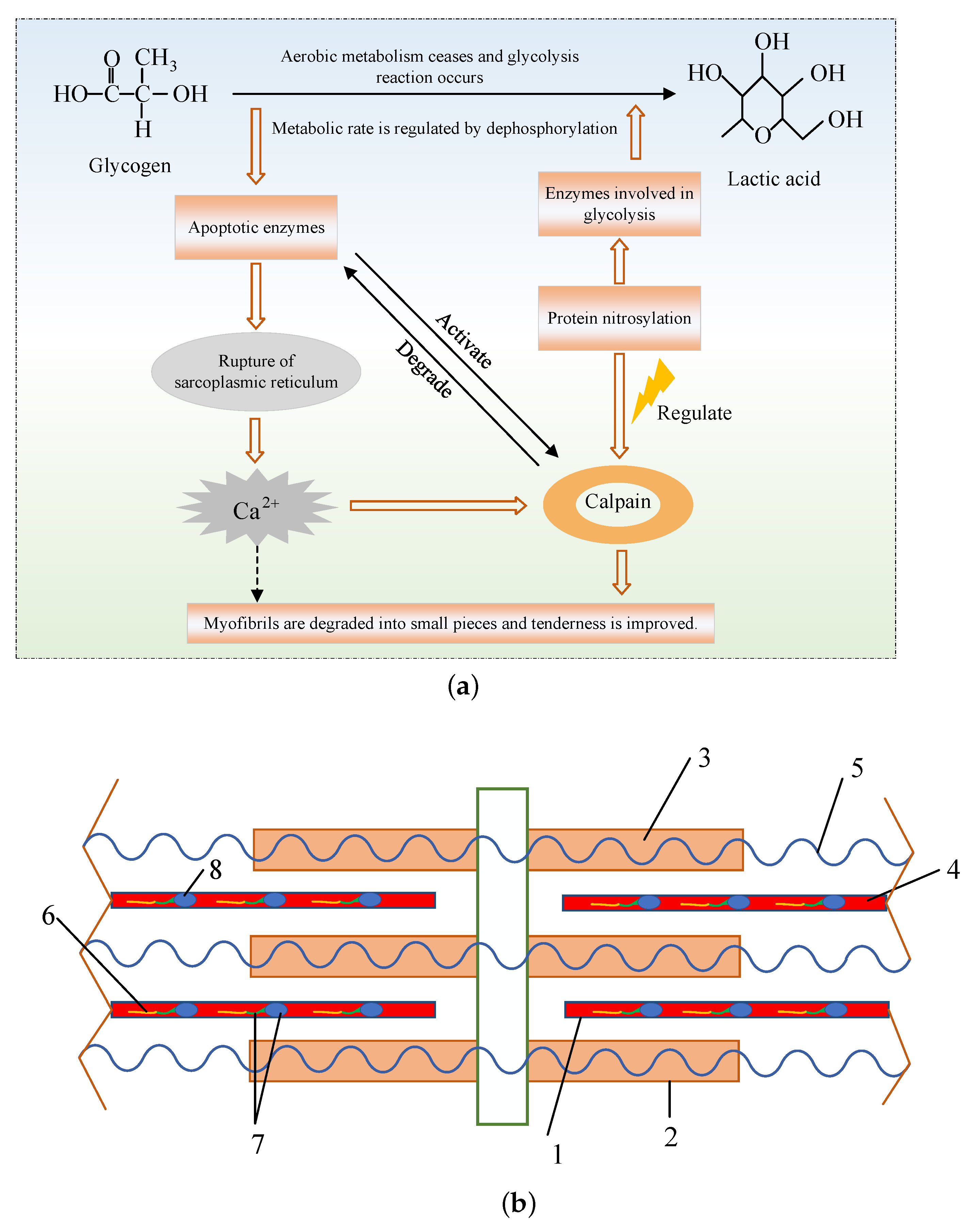
Calpain is a major contributor to meat tenderness improvement. It is a kind of protease and it can degrade various muscle proteins, mainly myofibrillar proteins. Numerous studies have found that calpain improved the tenderness of meat by degrading skeletal proteins in muscle cells such as desmin, fibrin, troponin-T and connexin. Therefore, the muscle fibers are eventually degraded and the shearing force of the skeletal muscle reduces, resulting in changes in the structure of myofibrils and the improvement of meat tenderness [19]. Calpain is a calcium ion-dependent protease and its activity can be activated by calcium ions. It can be divided into -calpain (micromolar level) and m-calpain (millimolar level) according to the calcium ion concentration required to activate the proteins. Calpain has been widely studied in the post-mortem maturation of meat. The role of calpain in the tenderization and the myofibrillar protein related to meat maturity after slaughter are shown in the Figure 2. It is extremely sensitive to temperature and pH, and the optimum temperature is about . Due to some existing factors, such as the autolysis and poor stability of calpain, and the pH condition, most of its activity is completely lost after the long maturation [20,21]. Nevertheless, what cannot be ignored is that calpains can greatly promote the degradation of the skeleton protein and the destruction of the integrity of myofibril structure, which is conducive to the further action of other proteases [22].
2.2. Cathepsins
Cathepsins are considered to play a major role in the protein degradation during the processing of Jinhua ham, especially cathepsin B and L. Unlike calpain, which plays a crucial role in the maturation and tenderization and the activity of which is almost completely lost at the end of ham’s elaboration procedure, cathepsin keeps active throughout the whole processing of dry-cured ham. At present, nearly 20 cathepsins have been isolated and identified. Some researchers have evaluated the activities of cathepsins and protein degradation as well as the enzymatic properties in hams. Partial characteristics of the common lysosome cathepsins existing in animal muscles are listed in Table 1. Venugopal et al. [24] examined the optimal pH and temperature, molecular weight and the isoelectric point of cathepsin D. Zhou et al. [25] discussed the kinetics of enzyme-catalyzed reactions of cathepsin L. Moreover, myosin, troponin, tropomyosin and actin can be degraded by cathepsin B. Myosin heavy chain, -actin, actin, and the T subunit of troponin will be degraded by cathepsin L [26].
Cathepsin B has been extracted and purified from the muscle, liver and pancreas of carp, bovine spleen, pig liver, mouse liver, human liver, salmon, tilapia, silver carp, and other fish muscles. Cathepsin B is composed of two molecular structures—namely cathepsin B1 and cathepsin B2—and the optimum temperature is about . In animals, cathepsin B usually exists in the following three forms: an inactive enzyme-endogenous inhibitor form, an inactive or low-activity precursor form, and the active mature monomer form. Cathepsin L has several complex structures. Iodoacetic acid, leupeptin, and anti-protease have a strong inhibitory effect on its activity, but aprotinin and sulfonyltoluene fluoride have no inhibitory effect [28]. It is excessively unstable at neutral and alkaline pH and easily denatured and inactivated. The optimum temperature of cathepsin L is 40–45 , and the high activity occurs at 20–60 . Its activity decrease rapidly at and cathepsin L is completely inactivated at about . Cathepsin B and L have extensive effects on protein degradation. For example, actin can be degraded into peptides released from the N-end and C-end of the protein by cathepsin B [29].
Cathepsin H possesses endonuclease and exonuclease properties with a molecular weight of 26–28 . It has maximum vitality at pH . Kaur et al. [30] confirmed that cathepsin H existed in animal skeletal muscle. In addition, for degrading muscle proteins and generating taste substances, cathepsin H is critical in Jinhua ham [31]. Cathepsin D is less stable than cathepsin B and L. Sárraga et al. [32] found that its activity decreased quickly during the production of dry-cured ham, which showed that the residual activity reached 10–20% of the raw hams after processing for 2.5 months. Cathepsin D may only be active during the first few months of processing.
2.3. Tripeptidases and Dipeptidases
In dry-cured ham, small peptides in the muscle are produced by peptidyl peptidases, including tripeptidylpeptidase (TPP) and dipeptidylpeptidase (DPP). TPP is an enzyme that can hydrolyze different tripeptides from the amino terminus of proteins and polypeptides. TPP I is glycoprotein and exists in the lysosome organelles with an optimal pH of . Substrate Gly-Pro-X can be hydrolyzed, where X is any kind of amino acid and hydrophobic amino acids are preferred [33]. In addition, TPP I is considered to be an aspartic peptidase, and its sequence has a certain similarity to carboxypeptidase belonging to the peptidase family. TPP II is an oligomer composed of subunits and the optimum pH is 6.5–7.5. TPP II can hydrolyze peptides to produce a variety of tripeptides, and Ala-Ala-Phe is a typical tripeptide preferentially hydrolyzed by TPP II [9].
DPP is always highly active throughout ham processing because of its diverse types, wide range of action and good stability [34]. It is the protease that generates dipeptide products from the amino terminal under the hydrolysis of peptides and proteins. DPP can be divided into four different types: DPP I, DPP II, DPP III and DPP IV. In terms of distribution, DPP I and DPP II are found in the lysosome. DPP III is present in the cytoplasm, and DPP IV is located on the cell membrane. Among all dipeptidases, DPP I exhibits high activity and is the most active enzyme. DPP I has good stability and plays an important role in the whole processing of dry-cured hams [35]. Wang et al. [36] have obtained a higher degree of protein degradation employing the raw material with higher DPP I activity, which is related to a higher amount of peptide production. It is consistent with a fact that DPP 1 activity is needed in the flavor formation process. DPP II plays a certain role, mainly before the period of ripening rather than throughout the whole ham processing [37]. DPP III reaches its maximum activity at a pH of 8.0 and a temperature of . Its activity decreases rapidly under acidic conditions and decreases significantly when the optimum temperature is exceeded [38]. DPP IV is a dipeptidase that is second only to DPP I in activity and it has a positive effect on the proteolysis.
2.4. Aminopeptidases
Free amino acids are due to the action of aminopeptidases, which are crucial in the formation of ham taste. Aminopeptidases cannot be separated from the proteolytic action when talking about ham ripening. Aminopeptidases are mainly involved in the later stages of dry-cured hams, which usually exist in the cytoplasm with complex structures and various types. Five main types are closely related to the processing of Jinhua ham and have been studied—namely alanyl amino peptidase (AAP), arginyl amino peptidase (RAP), leucyl amino peptidase (LAP), tyrosine acylaminopeptidase (TAP) and pyroglutamylaminopeptidase (p GAP) [39]. All of them show high activity between 30 and 40 and pH 5.0–7.0, which is similar to the ripening conditions of Jinhua ham. The analysis of potential activities of various aminopeptidases showed that AAP, RAP and LAP are active and maintain high residual vitality [18]. In the dry-cured hams, they are the main reasons for the accumulation of free amino acids. The actual effects of the three aminopeptidases are also different due to their substrate specificity.
AAP is the most abundant and important aminopeptidase in skeletal muscle sarcoplasm, accounting for 80–83% of the total activity. It has broad substrate specificity for aromatic amide bonds, aliphatic amide bonds and basic amide bonds. In comparison, the substrate range of LAP and RAP is much narrower. RAP is specific for basic amide bonds and it mainly hydrolyzes peptides with alkaline amino acids ends such as arginine and lysine [40]. The activity of LAP is very stable and high activity is still displayed in the later stage of dry-cured ham processing, so it plays a leading role in the formation of free amino acids. The concentration of free amino acids in the muscles continues to increase during the whole processing, especially for glutamic acid, arginine, alanine, aspartic acid, leucine, valine and lysine. The aminopeptidases activities of AAP, RAP and LAP are closely related to this increase in the Jinhua hams.
3. Action of Proteases for Improved Ham Quality
3.1. Protein Degradation
Intense proteolysis during the processing of Jinhua ham had been clarified [41,42]. Zhao et al. [26] determined the content of total nitrogen, total soluble nitrogen, non-protein nitrogen and soluble protein nitrogen for analyzing muscle protein degradation. The results showed that the soluble proteins are mainly degraded in the complex hydrolysis process, and are degraded faster than insoluble proteins. Sforza et al. [43] suggested that proteolytic enzymes lead to an accumulation of low molecular weight compounds. Zhao et al. [44] also indicated that the total concentration of free amino acids increased significantly during the storage of Jinhua ham, and the decisive role of aminopeptidases in the production of free amino acids was confirmed.
Yang et al. [45] found that higher moisture content promoted protein hydrolysis and the pH increased slightly to the range of 6.3–6.5, which was suitable for the action of many endogenous proteases in the muscle. Proteolysis index (PI) expresses the degree of proteolysis. Zhou et al. [46] ascertained that the diverse processing materials and processing technologies were important reasons for the difference in PI values. The significantly higher PI value is found in the defective ham, indicating more intense protein degradation. Simultaneously, cathepsin B and B + L with higher residual activity were detected in defective ham. Cathepsins led to stronger degradation in sarcoplasmic proteins and myofibril proteins.
3.2. Peptide and Free Amino Acid Generation
Small molecular peptides and free amino acids contribute to the ham flavor. Flavor is one of the most important quality indicators of meat products, and it is also a significant factor in attracting consumers. In the last 20 years, plenty of work has been carried out on the formation mechanism of dry-cured ham flavor. The results indicate that proteolysis and lipolysis are the main biochemical reactions in the generation of flavor or flavor precursors in ham processing. Proteolysis is dominated by endogenous proteases in view of the low microbial counts found inside hams [47]. The key part of dry-cured ham taste consists of small molecule peptides and free amino acids. Umami, sweetness, sourness and bitterness properties depend on their content and proportion. Figure 1a shows that the resulting taste-related peptides and free amino acids provide a taste for hams.
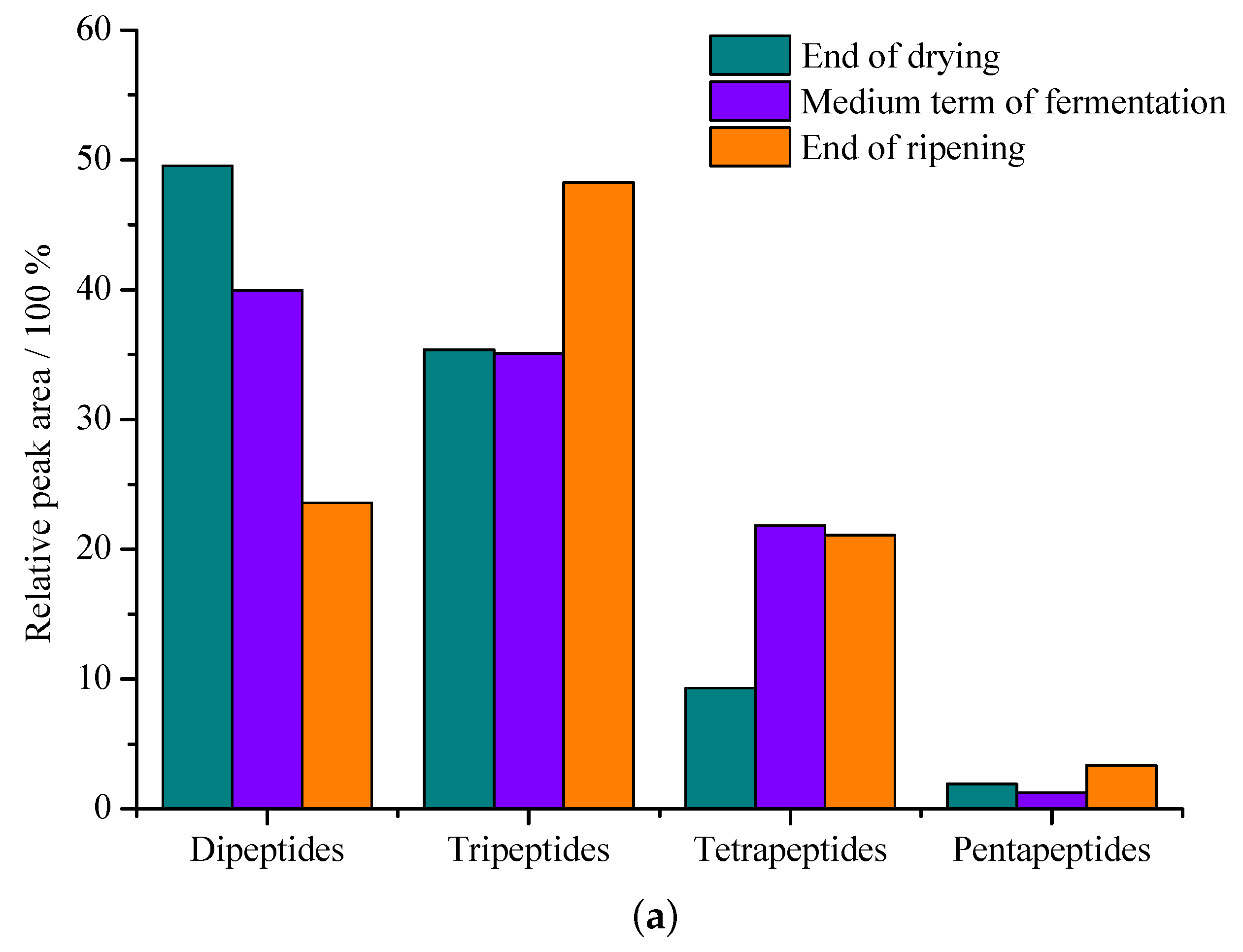
In many investigations, free amino acids, polypeptides and oligopeptides provide a unique flavor and quality in dry-cured hams, and they originate from extensive proteolysis of endogenous peptidases. A large amount of peptides increase after maturation and the change trend of small peptides in Jinhua ham is shown in Figure 3a. The accumulation of peptides derived from myosin is similar to the change in the total peptide content number, and the content of small peptides increases significantly, especially in the latter stages of ripening and post-ripening. Xing et al. [48] extracted and identified 213 peptides from Jinhua ham, and showed that polypeptide substances are mainly generated from myosin, troponin and actin. Huan et al. [49] surveyed the processing of Jinhua ham and found that 43, 46 and 63 major small peptides were obtained at different processing stages with molecular weights ranging from 204.1 to 1774.0 Da.
Recently, the use of mass spectrometry (MS) and peptomics methods has been able to better identify the source of the protein and the degradation site of the peptide. Mora et al. [50] identified the peptide sequences using LC-MS/MS. The dipeptides of DR, EN, HP, PL, IM, IQ, VF and the tripeptides of ALN, DPN, GHP, HGG, KLR, VGS were produced by creatine kinase degradation. The protein fragments of AQ, II, RG and IIP, RGA could originate from glyceraldehyde 3-Phosphate [51]. In addition, analysis of the correlation between the breaking sites of small peptides and enzymes in ham muscle provide better knowledge of the role of exopeptidase, which is shown in Figure 3b.
Furthermore, the relationship between various small peptides and the taste characteristics of ham is evident, and mainly depends on the length of the peptide chain itself, the composition and sequence of amino acids, and the original taste of their constituent amino acids [52]. The sensory properties of small peptides (generally peptides with molecular weight less than ) contain sweetness, sourness, saltiness, umami and bitterness. Aristoy et al. [53] focused on the taste characteristics of the peptides with a molecular weight less than 3 KDa, and they pointed out that the peptides with a molecular weight of about 1.8 kD exhibited a bitter taste. However, the molecular weight between 1.5 and 1.7 kD has an umami taste, and lower peptides (below ) possess a slightly sour taste. By the action of muscle dipeptidases especially DPP I and DPP II [54], some di-peptides such as Ala–Gln, Arg–Gly, Asn–Pro, lle–Leu, Ala–Gly, Ser–Gly and Ser–Gln are released from the N-terminal probably. In addition, tripeptides such as Ile–Ile–Pro, Arg–Gly–Ala, Gly–Asn–Pro, Gly–Ala–Gly and Gly–Pro–Gly are detected and released through the action of TPP I.
3.3. The Microstructure of Muscle
Not only protein composition, but also the structure, is affected by the action of proteolytic enzymes. It is noteworthy that protein degradation leads to the production of protein polymers or fragments [55]. Wu et al. [56] examined the changes in protein structure during the heating process of meat products. Hydrogen bonds, hydrophobicity and disulfide bonds are the forces for stabilizing the protein structure. Sun et al. [57] identified the structural changes and discussed the effects of protein oxidation on the myofibril protein emulsification. It is proved that the addition of oxidants enhanced the surface hydrophobicity of myofibrillar protein. Therefore, the investigation into microstructure in ham muscle tissue is as important as postmortem storage, which provides valuable information for ham manufacture. In previous research, we performed SDS-PAGE electrophoresis experiments on the sarcoplasmic and myofibril proteins extracted from Jinhua ham. The analysis showed that the electrophoresis band of sarcoplasmic and myofibrillar proteins decreased sharply, which was due to the important role of cathepsin B and L in the processing of Jinhua ham.
Furthermore, Harkouss et al. [58] determined the texture characteristics of dry-cured ham and highlighted the effect of endogenous proteolytic enzymes on muscle ultrastructure by electron microscopy. The model and relationship between structural parameters and proteolysis were investigated, which can be applied to any processing stages of Jinhua ham. As shown in Figure 4, the muscle fibers during different processing periods change gradually in the histological characteristics. In the beginning, the cross-section of the ham muscle tissue is relatively dense and the tissue structure is complete. The internal muscle fibers are thin and tightly arranged, which indicates that the higher the density of muscle fiber, the better the water retention and the finer the meat quality. After salting, moisture loss is accelerated with time, leading to muscle fibers atrophy and large gaps. On the contrary, the cross-section of ham muscle tissue tends to loosen and muscle fibers become thicker. Moreover, the hams, which have an excessive proteolysis due to marked protease activity, are prone to showing a softer tissue structure. In these mushy samples, the integrity of muscle cells and connective tissues disappears and the typical cellular structure can not be easily observed.
3.4. Endogenous Proteases and Texture Development
Texture defects of dry-cured ham induced by endogenous proteases are highlighted, such as pastiness, adhesiveness and softness. The myofibrillar proteins are destroyed through proteolysis, which constitutes the texture of dry-cured ham. Generally speaking, the texture property of Jinhua ham is a comprehensive impression that is mainly comprised of hardness, brittleness, adhesiveness, elasticity and cohesion. In Jinhua ham, the hardness, adhesiveness and chewiness increase significantly (), yet there is no significant difference in elasticity. It suggested that the difference in texture is related to the water content, salt content and the degree of protein hydrolysis [60]. What is more, excessive proteolytic activity may create damage in the structure, leading to pastiness and adhesiveness. Pastiness is an oral sensation described as the feeling of flour and water paste during mastication. This is the major texture problem in dry-cured ham with a 10% incidence in the industrial production. Hernández-Ramos et al. [61] displayed that the proteolytic index was the best parameter for characterizing the relationship between processing conditions and texture of dry-cured ham. Another reason for softness is incomplete balance of moisture inside the muscle and the insufficient drying, which results in excessive moisture. Zhou et al. [12] showed that the adhesiveness was negatively correlated with the moisture content, and reducing the moisture content was conductive to improving the adhesiveness of dry-cured ham. In contrast, stiffness means that the structure of dry-cured ham is too hard, which results from the excessive drying of the dry-cured ham processing. High-pressure processing (HPP) treatment has been confirmed to decrease both pastiness and viscosity values [43]. After rheological analysis, Coll-Brasas et al. [62] proposed an in-vitro approach, which allowed faster, reproducible, and less expensive measurements for the pastiness defect in dry-cured ham. Contreras et al. [63] examined the use of ultrasound to assess pastiness and determined dry-cured ham non-destructively. This technology is effective at quickly detecting and separating defective hams to ensure product homogeneity and high quality.
3.5. Endogenous Proteases and Flavor Development
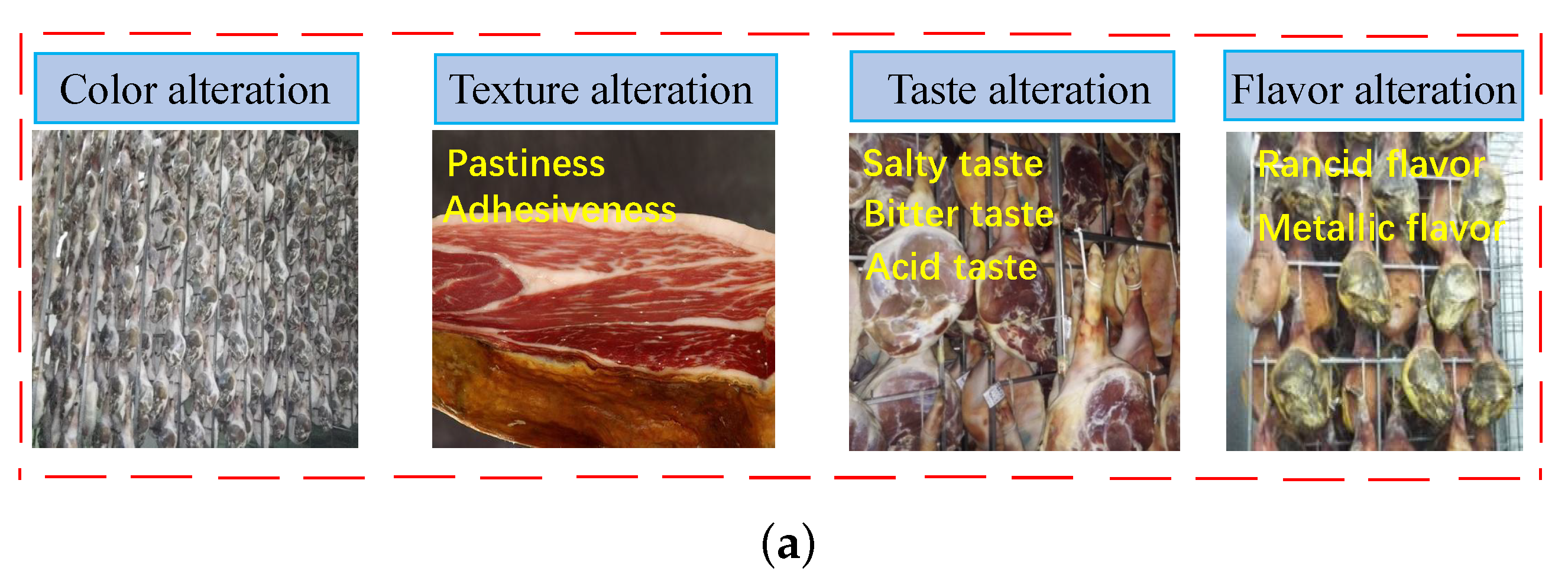
Strong endogenous enzymatic action is often a key factor in the development of sensory defects. In the time-consuming production of Jinhua ham, the degradation of muscle protein and degradation products will affect the texture, aroma and taste of the ham directly, which is dominated by the role of endogenous proteases [64]. The common defects in the ham quality are shown in Figure 5a. First of all, excessive saltness in the finished products is an urgent problem for the development of ham manufacturing. The defect in the ham quality is excessive salt, which is caused by the high salt content in the finished products. Meanwhile, NaCl is a potent inhibitor of most proteases involved in ham muscle and higher salt concentration suppresses putrid microorganisms reproduction inside and on the surface. However, the incidence of excessively soft and mushy hams will increase when sodium chloride is reduced, and the shelf life and safety of products is also not as expected [65]. Therefore, it is crucial to coordinate the endogenous protease activity and the salt amount.
Besides excessive saltiness, bitterness and sourness in Jinhua ham have usually produced an undesirable taste experience, and they generally appear with higher levels of proteolysis. The bitter hydrophobic free amino acids and short peptides accumulate massively in the highly proteolyzed hams. In an early study by Virgili et al. [69], high bitterness perception in aged ham was accompanied by pronounced endopeptidase activity, which was associated with high levels of methionine, asparagine, and isoleucine. This is in agreement with other research [70,71], so the ham can be effectively prevented from having a bitter taste by controlling the proteolytic index.
Additionally, dry-cured ham has also had an insufficient or even poor flavor. This is caused by an insufficient processing time to complete the necessary biochemical reactions, especially protein and lipid degradation reactions. As a result, the accumulation of flavor precursors and flavor substances in the muscle is scarce. In order to get round the quality defect of insufficient flavor, it is necessary to ensure a longer period for maturation (at least three months). Therefore, the selection of processing parameters is intricate and comprehensive. The biochemical changes that occur in the muscle tissues are quite intricate, so it is necessary to implement a complete and systematic control of the entire processing parameters to prevent the quality defects.
4. Key Parameters Influencing the Activities of Muscle Endogenous Proteases
4.1. Raw Material
Raw materials lead to different enzyme activities, including the breed, age, physical and chemical properties of the muscles. GarcÍa-Garrido et al. [72] showed that the reasons leading to different enzyme activity in raw materials are breed, age, physical and chemical properties of muscles. In several different ham types, Parma ham is made from Large White, Landrace and Duroc. Typical Spanish Iberian ham is mainly taken from Pure Iberian and Duroc males, and China Jinhua ham is made from “Liangtou Wu” pigs. The quality of the raw materials affects the rate and extent of biochemical reactions. Muscle properties, including pH, water content, salt concentration and tissue composition, are important factors affecting enzymatic activity. The biceps femoris (BF) and semimembranosus (SM) are two important muscle types with which to track the characteristics of dry-cured ham. The former is located inside and it is covered with a thick layer of subcutaneous fat one side, which slows the permeability of sodium chloride. The slow increase of salt in BF muscles throughout the process contributes to higher proteolytic activity [58]. On the contrary, SM muscles are close to the surface without fat coverage, resulting in fast absorption during salting. These two muscles have different protease activities, as well as intensive proteolysis [73]. Moreover, pig sex is also a factor in salt absorption and processing losses in dry-cured ham. The castration helps to improve the sensory quality of dry-cured ham [74].
PH value has an influence on the rate of an enzyme-catalyzed reaction directly and it plays a crucial role in ham manufacture. The pH value of raw material recommended for ham manufacture are between 5.6 and 6.2. With regard to the influence on enzymes, a low pH in hams is more prone to releasing cathepsins from lysosomes into muscle and enhances the activity of certain enzymes [75]. High pH values accelerate the growth and reproduction of microorganisms, which are also not suitable to be regarded as raw materials for Jinhua ham manufacture. For example, Guerrero et al. [76] discovered that the pastiness occurrence is associated with PSE meat. In response to the problem of softness in the dry-cured ham, the activity of cathepsin in fresh hind legs demand to be checked. Specifically, for legs with overhigh cathepsin activity, the amount of sodium chloride should be increased to inhibit protein degradation.
4.2. Processing Temperature
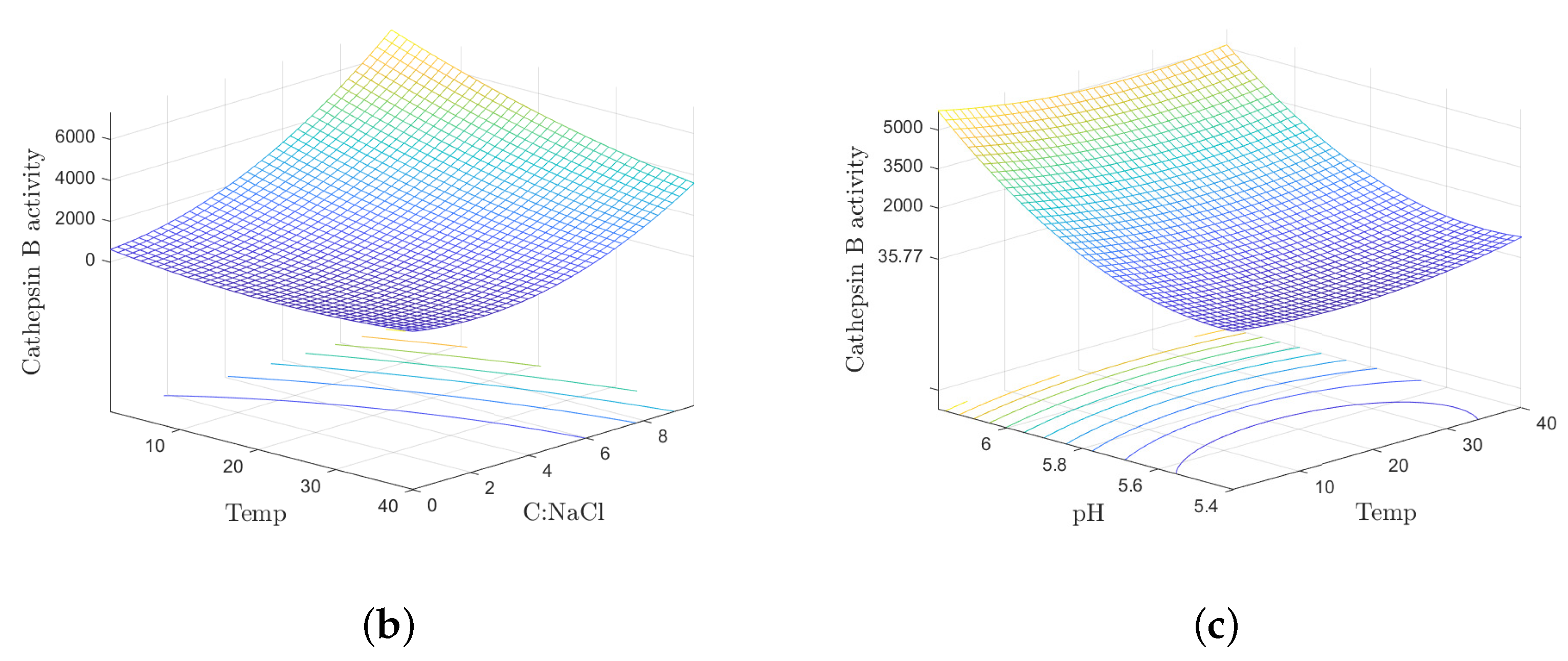
The temperature during the processing of Jinhua ham affects the enzyme activity directly, and has been listed in Table 2. Some researchers have found that the protease is the most active in the optimum temperature range with the pH, salinity and other conditions unchanged. For example, Moin et al. [77] reported that the optimum temperature of cathepsin B was about . Hu et al. [59] detected that cathepsins activities varied with temperature and increased under higher temperature at the beginning. In addition, a proper temperature is beneficial for shortening the ripening time, producing the special aromatic flavor and improving the quality of the final products. For example, Pérez-Santaescolástica et al. [78] applied a mild thermal treatment (around ) to avoid high instrumental adhesiveness and pastiness due to excessive proteolysis. Interestingly, as shown in Figure 1b,c, Zhao et al. [8] studied and found that there was an interactive effect between temperature and pH value. Specifically, the continuous decrease of water activity caused the decrease of protease activities, and the increment of maturation temperature increases the enzyme activities within a certain range. In the production, the management of drying rate is accomplished by relative humidity and the temperature of drying air. As mentioned in Table 2, the temperature is kept low until the water activity fall down to a level that suppresses microbial growth, which suggests a relatively low enzyme activity. During the period of drying, the temperature is increased to allow the increment in the proteases activities. It is an important stage for endogenous proteases to kick in.
4.3. Sodium Chloride Content and Processing Time
Sodium chloride plays an important role in regulating endogenous proteolysis. It is an indispensable material for salting processing, which functions mainly in the ways of seasoning, antiseptic and water retention improvement. However, excessive sodium intake will lead to high blood pressure and increase the incidence of heart disease and stroke [80]. The proper NaCl concentration is beneficial to the shelf life, safety characteristics, and the eating quality of cured meat products. However, lower cathepsin activity and excessive saltness mainly result from high salt content in the finished products. The increase in salt content and the loss of moisture result in the increment of hardness and chewiness of ham [81]. Thence, the problem of reducing salt content without affecting the palatability and safety of meat products has always been a hot research topic. Toldrá et al. [42] indicated that sodium chloride was a strong inhibitor for protease activity. Zhao et al. [8] also demonstrated that cathepsin B was susceptible to salt content, and the inhibitory effect will be enhanced with an increase in temperature. Specifically, higher temperatures increase the inhibitory effect on enzyme activity. Interestingly, Zhou et al. [21] found that salting treatment accelerated the release of cathepsin B from lysosomes and demonstrated that cathepsin B was sensitive to NaCl.
When it comes to the processing time, the activity of endogenous proteases is dynamic with the extension of time. Zhang et al. [82] found that the temperature and humidity were directly related to the dehydration rate and processing time. Ding et al. [83] showed that muscle protein undergone strong hydrolysis and the content of free amino acid increased over time. Salazar et al. [84] also proved that the content of free amino acids was positively correlated with the processing time. Moreover, the content of non-protein nitrogen and peptides increased over time. As described above, the endogenous protease activity in the hams is directly affected by the water activity, pH and temperature, and they are time-varying. During the whole processing of the Jinhua ham, the muscle pH increase slightly with time, and the temperature fluctuates in the range of 0 –37 . A significant decrease in endogenous protease activity occurs after salting. An important reduction of moisture content is typically observed, and Pugliese et al. [85] suggested that the salt content gradually increased over time. High sodium chloride content inhibited the growth of spoilage bacteria and produces a negative impact on the activity of endogenous proteases. Some proteases are relatively stable during the post-ripening period and not completely inactivated, which is due to the rising temperature and the destruction of organelles to release lysosomal proteases. The treatment time affects the molecular weight of peptides produced in ham. Hams with a shorter processing time have a higher molecular weight, while the molecular weight of hams with a longer processing time is smaller. This is because that large molecular weight peptides are further degraded into small molecular peptides with time [86].
4.4. Ultrasound-Assisted Technique and High-Pressure Processing during the Manufacture
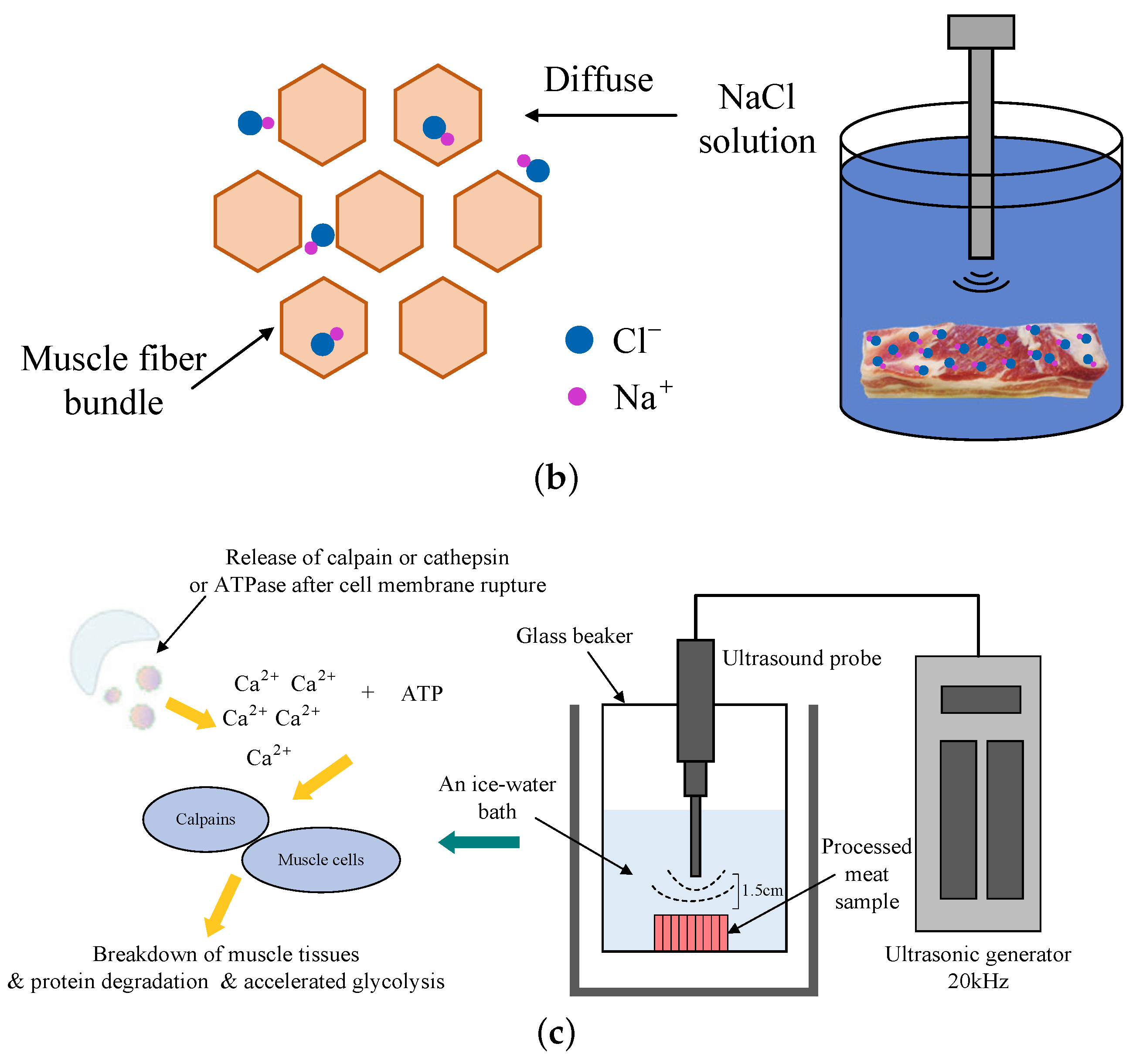
There are some non-destructive techniques that have been evaluated with regard to their implementation in the manufacturing of dry-cured ham, including irradiation, magnetic resonance imaging, infrared spectroscopy (IR), high pressure processing (HPP), and ultrasound (US). Kang et al. [87] found that the diffusion coefficient of NaCl or water increases significantly with the increase of ultrasonic intensity in the range of 2.39–20.96 , which is shown in Figure 5b. Furthermore, Refs. [88,89] showed that the ultrasound treatment caused damage to lysosomal membrane, which promoted the release of cathepsins and further improved their activities (Figure 5c).
In addition to the application in salting, there have been many reports of potential techniques for correcting and improving the organoleptic and textural attributes of defective ham. Sensory defects of ham are highly related to protein degradation catalyzed by endogenous proteases, which are located in the muscle tissues. The application of high-pressure technology has a key impact on the organoleptic characteristics of final products, which is based on the fact that high pressure will promote the changes in protein unfolding, amino acids releasing and cross-link between proteins. Buckow et al. [90] found that the activity of cathepsin B and L increased gradually by increasing the pressure to , which was due to the release of enzymes from muscle cells by the high pressure. When the processing pressure was increased to , the hardness increased and the pastiness and adhesiveness decreased in the dry-cured ham [91]. Rivas-Cañedo et al. [92] observed a higher proteolysis under and , resulting in further accumulation of free amino acids. After that, they also detected that the activity of cathepsin D would increase in the temperature range (33 –70 ), indicating that high pressure promoted the rupture of lysosomal membranes and accelerated the release of cathepsins. Zhou et al. [93] illustrated that ultrasound-assisted thermal treatment improved the activities of aminopeptidases, which further accelerated the degradation of polypeptides into free amino acids and then the taste attributes were enriched. It is worth noting that the appropriate operating conditions should be selected, or else the nutrients may be degraded and a peculiar smell generated, causing damage to the quality of the products [94]. For instance, Kang et al. [95] showed that inappropriate processing significantly increased the lipid and protein oxidation and led to high-level protein structure changes during salting.
5. Conclusions
The activity of muscle endogenous proteases can be detected in the whole process, and they play an important role in Jinhua ham flavor and texture. The regulation of enzyme activities can be achieved by changing the processing conditions. At the optimum temperature or pH, the protease is the most active. Salting and decreased water activity in muscle are associated with the decrease in enzymatic activity. In addition, the length of treatment time is positively correlated with the degree of proteolysis, that is, the accumulation of flavor substances. Some new and developing processing techniques accelerate the release of endogenous proteases from cell membranes, such as high-pressure technology and ultrasound. However, it is a challenge to decide on appropriate processing parameters. In the future, for Jinhua hams, the quantitative relationship needs to be further explored between key process parameters and endogenous proteases activities. Additionally, the interaction between various factors involved in ham manufacturing cannot be ignored, and a more accurate method for enzyme activity determination is needed.
Author Contributions
S.H. designed the study and interpreted the results. S.H. and C.L. investigated test data and drafted the manuscript. S.H., W.Z. innovated the data processing software. G.Z. and X.X. conceived the project. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (No. 31972097) and Jiangsu Key Research and Development Plan (Modern Agriculture) (No. BE2020302).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Ramona Bosse, A.M.M.G.; Weiss, J. Recent advances in cured raw ham manufacture. Crit. Rev. Food Sci. Nutr. 2018, 4, 610–630. [Google Scholar] [CrossRef]
- Liu, D.; Bai, L.; Feng, X.; Chen, Y.P.; Zhang, D.; Yao, W.; Zhang, H.; Chen, G.; Liu, Y. Characterization of Jinhua ham aroma profiles in specific to aging time by gas chromatography-ion mobility spectrometry (GC-IMS). Meat Sci. 2020, 168, 108178. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M.; Sugawara, T.; Obiya, S.; Enomoto, A.; Kaneko, M.; Ota, S.; Soga, T.; Tomita, M. Sensory properties and metabolomic profiles of dry-cured ham during the ripening process. Food Res. Int. 2020, 129, 108850. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, Y.P.; Blank, I.; Li, F.; Li, C.; Liu, Y. GC×GC-ToF-MS and GC-IMS based volatile profile characterization of the Chinese dry-cured hams from different regions. Food Res. Int. 2021, 142, 110222. [Google Scholar] [CrossRef] [PubMed]
- MartÍn, A.; Benito, M.J.; Aranda, E.; Ruiz-Moyano, S.; Córdoba, J.J.; Córdoba, M.G. Characterization by Volatile Compounds of Microbial Deep Spoilage in Iberian Dry-Cured Ham. J. Food Sci. 2010, 75, M360–M365. [Google Scholar] [CrossRef] [PubMed]
- López-Pedrouso, M.; Pérez-Santaescolástica, C.; Franco, D.; Fulladosa, E.; Carballo, J.; Zapata, C.; Lorenzo, J.M. Comparative proteomic profiling of myofibrillar proteins in dry-cured ham with different proteolysis indices and adhesiveness. Food Chem. 2018, 244, 238–245. [Google Scholar] [CrossRef]
- Larrea, V.; Hernando, I.; Quiles, A.; Lluch, M.A.; P’erez-Munuera, I. Changes in proteins during Teruel dry-cured ham processing. Meat Sci. 2006, 74, 586–593. [Google Scholar] [CrossRef]
- Zhao, G.M.; Zhou, G.H.; Wang, Y.L.; Xu, X.L.; Huan, Y.J.; Wu, J.Q. Time-related changes in cathepsin B and L activities during processing of Jinhua ham as a function of pH, salt and temperature. Meat Sci. 2005, 70, 381–388. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Wu, J.Q.; Tang, C.B.; Li, G.; Dai, C.; Bai, Y.; Li, C.B.; Xu, X.L.; Zhou, G.H.; Cao, J.X. Comparing the proteomic profile of proteins and the sensory characteristics in Jinhua ham with different processing procedures. Food Control 2019, 106, 106694. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Bai, Y.; Wang, C.; Li, C.B.; Xu, X.L.; Pan, D.D.; Cao, J.X.; Zhou, G.H. 1H NMR-based metabolomics and sensory evaluation characterize taste substances of Jinhua ham with traditional and modern processing procedures. Food Control 2021, 126, 107873. [Google Scholar] [CrossRef]
- Arnau, J.; Serra, X.; Comaposada, J.; Gou, P.; Garriga, M. Technologies to shorten the drying period of dry-cured meat products. Meat Sci. 2007, 77, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.Y.; Wang, C.; Tang, C.B.; Dai, C.; Bai, Y.; Yu, X.B.; Li, C.B.; Xu, X.L.; Zhou, G.H.; Cao, J.X. Label-free proteomics reveals the mechanism of bitterness and adhesiveness in Jinhua ham. Food Chem. 2019, 297, 125012. [Google Scholar] [CrossRef]
- Petrova, I.; Aasen, I.M.; Rustad, T.; Eikevik, T.M. Manufacture of dry-cured ham: A review. Part 1. Biochemical changes during the technological process. Eur. Food Res. Technol. 2015, 241, 587–599. [Google Scholar] [CrossRef]
- Marušić, R.N.; Poljanec, I.; Petričević, S.; Mora, L.; Medić, H. Influence of Muscle Type on Physicochemical Parameters, Lipolysis, Proteolysis, and Volatile Compounds throughout the Processing of Smoked Dry-Cured Ham. Foods 2021, 10, 1228. [Google Scholar] [CrossRef] [PubMed]
- Xiang, A.; Wang, Y.; Sun, B.; Xu, X.; Zhou, G.; Zhou, C.; Geng, F.; Pan, D.; Cao, J. Peptidomic insights on structural protein hydrolysis by exopeptidase activities and its effects on bitterness defects of dry-cured ham. Int. J. Food Sci. Technol. 2023, 58, 755–765. [Google Scholar] [CrossRef]
- Inguglia, E.S.; Zhang, Z.; Tiwari, B.K.; Kerry, J.P.; Burgess, C.M. Salt reduction strategies in processed meat products—A review. Trends Food Sci. Technol. 2017, 59, 70–78. [Google Scholar] [CrossRef]
- Petrova, I.; Bantle, M.; Eikevik, T.M. Manufacture of dry-cured ham: A review. Part 2. Drying kinetics, modeling and equipment. Eur. Food Res. Technol. 2015, 241, 447–458. [Google Scholar] [CrossRef]
- Zhou, G.H.; Zhao, G.M. Biochemical changes during processing of traditional Jinhua ham. Meat Sci. 2007, 77, 114–120. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Wang, Y.; Pan, D.D.; Cao, J.X.; Chen, Y.J.; Liu, Y.; Sun, Y.Y.; Ou, C.R. The changes in the proteolysis activity and the accumulation of free amino acids during chinese traditional dry-cured loins processing. Food Sci. Biotechnol. 2017, 26, 679–687. [Google Scholar] [CrossRef]
- Dang, Y.; Gao, X.; Ma, F.; Wu, X. Comparison of umami taste peptides in water-soluble extractions of Jinhua and Parma hams. LWT-Food Sci. Technol. 2015, 60, 1179–1186. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Pan, D.D.; Bai, Y.; Li, C.B.; Xu, X.L.; Zhou, G.H.; Cao, J.X. Evaluating endogenous protease of salting exudates during the salting process of Jinhua ham. LWT-Food Sci. Technol. 2019, 101, 76–82. [Google Scholar] [CrossRef]
- Zhu, C.Z.; Tian, W.; Li, M.Y.; Liu, Y.X.; Zhao, G.M. Separation and identification of peptides from dry-cured Jinhua ham. Int. J. Food Prop. 2018, 20, S2980–S2989. [Google Scholar] [CrossRef]
- Chéret, R.; Delbarre-Ladrat, C.; Lamballerie-Anton, M.D.; Verrez-Bagnis, V. Calpain and cathepsin activities in post mortem fish and meat muscles. Food Chem. 2007, 101, 1474–1479. [Google Scholar] [CrossRef] [Green Version]
- Venugopal, A.; Siva Kumar, N. Biochemical characterization of cathepsin D from the mussel Lamellidens corrianus. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2014, 169, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.Y.; Chang, X.N.; Bao, S.S.; Song, L.; Zhu, B.W.; Dong, X.P.; Zong, Y.; Li, D.M.; Zhang, M.M.; Liu, Y.X.; et al. Purification and partial characterisation of a cathepsin L-like proteinase from sea cucumber (Stichopus japonicus) and its tissue distribution in body wall. Food Chem. 2014, 158, 192–199. [Google Scholar] [CrossRef]
- Zhao, G.M.; Tian, W.; Liu, Y.X.; Zhou, G.H.; Xu, X.L.; Li, M.Y. Proteolysis in biceps femoris during Jinhua ham processing. Meat Sci. 2008, 79, 39–45. [Google Scholar] [CrossRef]
- Wu, Y.; Cao, S.M. Study on endogenous protease and protein degradation of dry-salted Decapterus maruadsi. CyTA-J. Food 2018, 16, 350–356. [Google Scholar] [CrossRef] [Green Version]
- Zhong, C.; Cai, Q.F.; Liu, G.M.; Sun, L.C.; Hara, K.; Su, W.J.; Cao, M.J. Purification and characterisation of cathepsin L from the skeletal muscle of blue scad (Decapterus maruadsi) and comparison of its role with myofibril-bound serine proteinase in the degradation of myofibrillar proteins. Food Chem. 2012, 133, 1560–1568. [Google Scholar] [CrossRef]
- Ge, L.; Xu, Y.; Xia, W.; Jiang, Q. Synergistic action of cathepsin B, L, D and calpain in disassembly and degradation of myofibrillar protein of grass carp. Food Res. Int. 2018, 109, 481–488. [Google Scholar] [CrossRef]
- Kaur, L.; Hui, S.X.; Boland, M. Changes in Cathepsin Activity during Low-Temperature Storage and Sous Vide Processing of Beef Brisket. Food Sci. Anim. Resour. 2020, 40, 415–425. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Xia, Q.; He, J.; Sun, Y.Y.; Dang, Y.L.; Zhou, G.H.; Geng, F.; Pan, D.D.; Cao, J.X. Insights into ultrasonic treatment on the mechanism of proteolysis and taste improvement of defective dry-cured ham. Food Chem. 2022, 388, 133059. [Google Scholar] [CrossRef] [PubMed]
- Sárraga, C.; Gil, M.; GarcÍa-Regueiro, J.A. Comparison of calpain and cathepsin (B, L and D) activities during dry-cured ham processing from heavy and light large white pigs. J. Science Food Agric. 1993, 1, 71–75. [Google Scholar] [CrossRef]
- Gallego, M.; Mora, L.; Toldrá, F. The relevance of dipeptides and tripeptides in the bioactivity and taste of dry-cured ham. Food Prod. Process. Nutr. 2019, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Sentandreu, M.; Toldrá, F. Dipeptidyl peptidase activities along the processing of Serrano dry-cured ham. Eur. Food Res. Technol. 2001, 213, 83–87. [Google Scholar] [CrossRef]
- Mora, L.; Sentandreu, M.A.; Koistinen, K.M.; Fraser, P.D.; Toldrá, F.; Bramley, P.M. Naturally Generated Small Peptides Derived from Myofibrillar Proteins in Serrano Dry-Cured Ham. J. Agric. Food Chem. 2009, 57, 3228–3234. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, Y.; Zhang, J.; Li, X.; Lin, Z.; Ma, C. Proteolysis, protein oxidation and protease activity in dry-cured Xuanwei ham during the salting stages. Int. J. Food Sci. Technol. 2011, 46, 1370–1377. [Google Scholar] [CrossRef]
- Iwaniak, A.; Hrynkiewicz, M.; Bucholska, J.; Minkiewicz, P.; Darewicz, M. Understanding the nature of bitter-taste di- and tripeptides derived from food proteins based on chemometric analysis. J. Food Biochem. 2019, 43, e12500. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, P.; Muenzner, M.; Kästorf, M.; Santos, K.; Hartmann, T.; Dienelt, A.; Rehfeld, L.; Bergmann, A. A novel and highly efficient purification procedure for native human dipeptidyl peptidase 3 from human blood cell lysate. PLoS ONE 2019, 14, e0220866. [Google Scholar] [CrossRef] [Green Version]
- Toldrá, F.; Aristoy, M.C.; Flores, M. Contribution of muscle aminopeptidases to flavor development in dry-cured ham. Food Res. Int. 2000, 33, 181–185. [Google Scholar] [CrossRef]
- Petrova, I.; Tolstorebrov, I.; Mora, L.; Toldrá, F.; Eikevik, T.M. Evolution of proteolytic and physico-chemical characteristics of Norwegian dry-cured ham during its processing. Meat Sci. 2016, 121, 243–249. [Google Scholar] [CrossRef] [Green Version]
- Toldrá, F.; Flores, M. The role of muscle proteases and lipases in flavor development during the processing of dry-cured ham. Crit. Rev. Food Sci. Nutr. 1998, 38, 331–352. [Google Scholar] [CrossRef] [PubMed]
- Toldrá, F. The role of muscle enzymes in dry-cured meat products with different drying conditions. Trends Food Sci. Technol. 2006, 17, 164–168. [Google Scholar] [CrossRef]
- Sforza, S.; Pigazzani, A.; Motti, M.; Porta, C.; Virgili, R.; Galaverna, G.; Dossena, A.; Marchelli, R. Oligopeptides and free amino acids in Parma hams of known cathepsin B activity. Food Chem. 2001, 75, 267–273. [Google Scholar] [CrossRef]
- Zhao, G.M.; Zhou, G.H.; Tian, W.; Xu, X.L.; Wang, Y.L.; Luo, X. Changes of alanyl aminopeptidase activity and free amino acid contents in biceps femoris during processing of Jinhua ham. Meat Sci. 2005, 71, 612–619. [Google Scholar] [CrossRef]
- Yang, H.; Ma, C.; Qiao, F.; Song, Y.; Du, M. Lipolysis in intramuscular lipids during processing of traditional Xuanwei ham. Meat Sci. 2005, 71, 670–675. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Le, Y.; Zheng, Y.Y.; Wang, J.J.; Li, G.; Bai, Y.; Li, C.B.; Xu, X.L.; Zhou, G.H.; Cao, J.X. Characterizing the effect of free amino acids and volatile compounds on excessive bitterness and sourness in defective dry-cured ham. LWT-Food Sci. Technol. 2020, 123, 109071. [Google Scholar] [CrossRef]
- Marušić, N.; Vidaček, S.; Janči, T.; Petrak, T.; Medič, H. Determination of volatile compounds and quality parameters of traditional Istrian dry-cured ham. Meat Sci. 2014, 96, 1409–1416. [Google Scholar] [CrossRef]
- Xing, L.; Liu, R.; Gao, X.; Zheng, J.; Wang, C.; Zhou, G.; Zhang, W. The proteomics homology of antioxidant peptides extracted from dry-cured Xuanwei and Jinhua ham. Food Chem. 2018, 266, 420–426. [Google Scholar] [CrossRef]
- Huan, Y.; Zhou, G.; Zhao, G.; Xu, X.; Peng, Z. Changes in flavor compounds of dry-cured Chinese Jinhua ham during processing. Meat Sci. 2005, 71, 291–299. [Google Scholar] [CrossRef]
- Mora, L.; Gallego, M.; Aristoy, M.C.; Fraser, P.D.; Toldrá, F. Peptides naturally generated from ubiquitin-60S ribosomal protein as potential biomarkers of dry-cured ham processing time. Food Control 2015, 48, 102–107. [Google Scholar] [CrossRef] [Green Version]
- Mora, L.; Gallego, M.; Toldrá, F. Degradation of myosin heavy chain and its potential as a source of natural bioactive peptides in dry-cured ham. Food Biosci. 2019, 30, 100416. [Google Scholar] [CrossRef]
- Gallego, M.; Toldr’a, F.; Mora, L. Quantification and in silico analysis of taste dipeptides generated during dry-cured ham processing. Food Chem. 2022, 370, 130977. [Google Scholar] [CrossRef] [PubMed]
- Aristoy, M.C.; Fidel, T. Isolation of flavor peptides from raw pork meat and dry-cured ham. Dev. Food Sci. 1995, 37, 1323–1344. [Google Scholar]
- Mora, L.; Gallego, M.; Escudero, E.; Reig, M.; Aristoy, M.C.; Toldrá, F. Small peptides hydrolysis in dry-cured meats. Int. J. Food Microbiol. 2015, 212, 9–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arroyo-Maya, I.J.; Campos-Terán, J.; Hernández-Arana, A.; McClements, D.J. Characterization of flavonoid-protein interactions using fluorescence spectroscopy: Binding of pelargonidin to dairy proteins. Food Chem. 2016, 213, 431–439. [Google Scholar] [CrossRef]
- Wu, M.; Xiong, Y.L.; Chen, J.; Tang, X.; Zhou, G. Rheological and Microstructural Properties of Porcine Myofibrillar Protein-Lipid Emulsion Composite Gels. J. Food Sci. 2009, 74, E207–E217. [Google Scholar] [CrossRef]
- Sun, W.; Zhou, F.; Sun, D.W.; Zhao, M. Effect of Oxidation on the Emulsifying Properties of Myofibrillar Proteins. Food Bioprocess Technol. 2012, 6, 1703–1712. [Google Scholar] [CrossRef]
- Harkouss, R.; Astruc, T.; Lebert, A.; Gatellier, P.; Loison, O.; Safa, H.; Portanguen, S.; Parafita, E.; Mirade, P.S. Quantitative study of the relationships among proteolysis, lipid oxidation, structure and texture throughout the dry-cured ham process. Food Chem. 2015, 166, 522–530. [Google Scholar] [CrossRef]
- Hu, S.; Zhou, G.; Xu, X.; Zhang, W.; Li, C. Contribution of cathepsin B and L to endogenous proteolysis in the course of modern Jinhua ham processing. Food Control 2021, 135, 108584. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Y.; He, J.; Pan, D.; Wang, H.; Cao, J. Evaluating the profile of myofibrillar proteins and its relationship with tenderness among five styles of dry-cured hams. Int. J. Food Sci. Technol. 2021, 56, 259–268. [Google Scholar] [CrossRef]
- Hernández-Ramos, P.; Vivar-Quintana, A.M.; Revilla, I.; González-Martín, M.I.; Hernández-Jiménez, M.; Martínez-Martín, I. Prediction of Sensory Parameters of Cured Ham: A Study of the Viability of the Use of NIR Spectroscopy and Artificial Neural Networks. Sensors 2020, 20, 5624. [Google Scholar] [CrossRef] [PubMed]
- Coll-Brasas, E.; Laguna, L.; Tárrega, A.; Arnau, J.; Claret, A.; Fulladosa, E. Evaluation of dry-cured ham pastiness through rheological measurements of aqueous extracts obtained in vitro mimicking the mastication process. LWT-Food Sci. Technol. 2021, 145, 111272. [Google Scholar] [CrossRef]
- Contreras, M.; Benedito, J.; Quiles, A.; Lorenzo, J.M.; Fulladosa, E.; Gou, P.; Garcia-Perez, J.V. Assessing the textural defect of pastiness in dry-cured pork ham using chemical, microstructural, textural and ultrasonic analyses. J. Food Eng. 2020, 265, 109690. [Google Scholar] [CrossRef]
- Hu, S.; Zhou, G.; Xu, X.; Zhang, W.; Li, C. Insight into the impacts of Jinhua ham processing conditions on cathepsin B activity and conformation changes based on molecular simulation. LWT-Food Sci. Technol. 2023, 174, 114459. [Google Scholar] [CrossRef]
- Aaslyng, M.D.; Vestergaard, C.; Koch, A.G. The effect of salt reduction on sensory quality and microbial growth in hotdog sausages, bacon, ham and salami. Meat Sci. 2014, 96, 47–55. [Google Scholar] [CrossRef]
- Zhou, C.; Xia, Q.; L, D.; He, J.; Sun, Y.; Dang, Y.; Geng, F.; Pan, D.; Cao, J.; Zhou, G. Recent developments in off-odor formation mechanism and the potential regulation by starter cultures in dry-cured ham. Crit. Rev. Food Sci. Nutr. 2022, 9, 1–15. [Google Scholar] [CrossRef]
- López-Pedrouso, M.; Pérez-Santaescolástica, C.; Franco, D.; Carballo, J.; Garcia-Perez, J.V.; Benedito, J.; Zapata, C.; Lorenzo, J.M. Proteomic footprint of ultrasound intensification on sliced dry-cured ham subjected to mild thermal conditions. J. Proteom. 2019, 193, 123–130. [Google Scholar] [CrossRef]
- Kang, D.c.; Wang, A.r.; Zhou, G.h.; Zhang, W.g.; Xu, S.m.; Guo, G.p. Power ultrasonic on mass transport of beef: Effects of ultrasound intensity and NaCl concentration. Innov. Food Sci. Emerg. Technol. 2016, 35, 36–44. [Google Scholar] [CrossRef]
- Virgili, R.; Schivazappa, C.; Parolari, G.; Bordini, C.S.; Degni, M. Proteases in fresh pork muscle and their influence on bitter taste formation in dry-cured ham. J. Food Biochem. 1998, 22, 53–63. [Google Scholar] [CrossRef]
- Jiménez-Colmenero, F.; Ventanas, J.; Toldrá, F. Nutritional composition of dry-cured ham and its role in a healthy diet. Meat Sci. 2010, 84, 585–593. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Tang, C.B.; Wang, C.; Dai, C.; Bai, Y.; Yu, X.B.; Li, C.B.; Xu, X.L.; Zhou, G.H.; Cao, J.X. Insights into the evolution of myosin light chain isoforms and its effect on sensory defects of dry-cured ham. Food Chem. 2020, 315, 126318. [Google Scholar] [CrossRef] [PubMed]
- GarcÍa-Garrido, J.A.; Quiles-Zafra, R.; Tapiador, J.; Luque De Castro, M.D. Activity of cathepsin B, D, H and L in Spanish dry-cured ham of normal and defective texture. Meat Sci. 2000, 56, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-RamÍrez, J.; Arnau, J.; Serra, X.; Gou, P. Effect of pH24, NaCl content and proteolysis index on the relationship between water content and texture parameters in biceps femoris and semimembranosus muscles in dry-cured ham. Meat Sci. 2006, 72, 185–194. [Google Scholar] [CrossRef]
- Bañón, S.; Gil, M.D.; Garrido, M.D. The effects of castration on the eating quality of dry-cured ham. Meat Sci. 2003, 3, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Čandek-Potokar, M.; Škrlep, M. Factors in pig production that impact the quality of dry-cured ham: A review. Animal 2012, 6, 327–338. [Google Scholar] [CrossRef]
- Guerrero, L.; Gobantes, I.; Oliver, M.A.; Arnau, J.; Dolors Guȧrdia, M.; Elvira, J.; Riu, P.; Grėbol, N.; Monfort, J.M. Green hams electrical impedance spectroscopy (EIS) measures and pastiness prediction of dry cured hams. Meat Sci. 2004, 66, 289–294. [Google Scholar] [CrossRef]
- Moin, K.; Day, N.A.; Sameni, M.; Hasnain, S.; Hirama, T.; Sloane, B.F. Human tumour cathepsin B. Comparison with normal liver cathepsin B. Biochem. J. 1992, 285, 427–434. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Santaescolástica, C.; Carballo, J.; Fulladosa, E.; Garcia-Perez José, V.; Benedito, J.; Lorenzo, J.M. Application of temperature and ultrasound as corrective measures to decrease the adhesiveness in dry-cured ham. Influence on free amino acid and volatile compound profile. Food Res. Int. 2018, 114, 140–150. [Google Scholar] [CrossRef]
- Zhou, C.; Zhan, G.; Pan, D.; Zhou, G.; Wang, Y.; He, J.; Cao, J. Charactering the spoilage mechanism of “three sticks” of Jinhua ham. Food Sci. Hum. Wellness 2022, 11, 1322–1330. [Google Scholar] [CrossRef]
- Bhat, Z.F.; Morton, J.D.; Mason, S.L.; Bekhit, A.E.D.A. The application of pulsed electric field as a sodium reducing strategy for meat products. Food Chem. 2020, 306, 125622. [Google Scholar] [CrossRef]
- Caballero, D.; Antequera, T.; Caro, A.; Duran, M.L.; Perez-Palacios, T. Data Mining on MRI-Computational Texture Features to Predict Sensory Characteristics in Ham. Food Bioprocess Technol. 2016, 9, 699–708. [Google Scholar] [CrossRef]
- Zhang, J.; Zhen, Z.; Zhang, W.; Zeng, T.; Zhou, G. Effect of intensifying high-temperature ripening on proteolysis, lipolysis and flavor of Jinhua ham. J. Sci. Food Agric. 2009, 89, 834–842. [Google Scholar] [CrossRef]
- Ding, X.; Wang, G.; Zou, Y.; Zhao, Y.; Ge, C.; Liao, G. Evaluation of small molecular metabolites and sensory properties of Xuanwei ham salted with partial replacement of NaCl by KCl. Meat Sci. 2021, 175, 108465. [Google Scholar] [CrossRef] [PubMed]
- Salazar, E.; Cayuela, J.M.; Abellán, A.; Bueno-Gavilá, E.; Tejada, L. Fatty Acids and Free Amino Acids Changes during Processing of a Mediterranean Native Pig Breed Dry-Cured Ham. Foods 2020, 9, 1170. [Google Scholar] [CrossRef]
- Pugliese, C.; Sirtori, F.; Škrlep, M.; Piasentier, E.; Calamai, L.; Franci, O.; Čandek Potokar, M. The effect of ripening time on the chemical, textural, volatile and sensorial traits of Bicep femoris and Semimembranosus muscles of the Slovenian dry-cured ham Kraški pršut. Meat Sci. 2015, 100, 58–68. [Google Scholar] [CrossRef]
- Sforza, S.; Galaverna, G.; Schivazappa, C.; Marchelli, R.; Dossena, A.; Virgili, R. Effect of extended aging of parma dry-cured ham on the content of oligopeptides and free amino acids. J. Agric. Food Chem. 2006, 54, 9422–9429. [Google Scholar] [CrossRef]
- Kang, Z.L.; Zou, Y.F.; Xu, X.L.; Zhu, C.Z.; Wang, P.; Zhou, G.H. Effect of a beating process, as a means of reducing salt content in Chinese-style meatballs (kung-wan): A physico-chemical and textural study. Meat Sci. 2014, 96, 147–152. [Google Scholar] [CrossRef]
- Zhang, J.; Toldrá, F.; Zhang, W.; Yin, Y.; Zhu, Z. Study on the effects and mechanisms of ultrasound on the peptide profile and taste of unsmoked bacon using peptidomics and bioinformatics. Food Chem. 2023, 414, 135764. [Google Scholar] [CrossRef]
- Wang, L.; Li, J.; Teng, S.; Zhang, W.; Purslow, P.P.; Zhang, R. Changes in collagen properties and cathepsin activity of beef M. semitendinosus by the application of ultrasound during post-mortem aging. Meat Sci. 2022, 185, 108718. [Google Scholar] [CrossRef]
- Buckow, R.; Sikes, A.; Tume, R. Effect of High Pressure on Physicochemical Properties of Meat. Crit. Rev. Food Sci. Nutr. 2013, 53, 770–786. [Google Scholar] [CrossRef]
- Coll-Brasas, E.; Arnau, J.; Gou, P.; Lorenzo, J.M.; GarcÍa-Pérez, J.V.; Fulladosa, E. Effect of high pressure processing temperature on dry-cured hams with different textural characteristics. Meat Sci. 2019, 152, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Cañedo, A.; Martínez-Onandi, N.; Gaya, P.; Nuñez, M.; Picon, A. Effect of high-pressure processing and chemical composition on lipid oxidation, aminopeptidase activity and free amino acids of Serrano dry-cured ham. Meat Sci. 2021, 172, 108349. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.Y.; Pan, D.D.; Cao, J.X.; Zhou, G.H. A comprehensive review on molecular mechanism of defective dry-cured ham with excessive pastiness, adhesiveness, and bitterness by proteomics insights. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3838–3857. [Google Scholar] [CrossRef] [PubMed]
- Pingret, D.; Fabiano-Tixier, A.S.; Chemat, F. Degradation during application of ultrasound in food processing: A review. Food Control 2013, 31, 593–606. [Google Scholar] [CrossRef]
- Kang, D.C.; Zou, Y.H.; Cheng, Y.P.; Xing, L.J.; Zhou, G.H.; Zhang, W.G. Effects of power ultrasound on oxidation and structure of beef proteins during curing processing. Ultrason. Sonochem. 2016, 33, 47–53. [Google Scholar] [CrossRef]
Figure 1.
The formation of flavor substances in Jinhua ham and the interactive effect between processing parameters. (a) Formation of flavor substances in Jinhua ham and the role of endogenous protease [18]. (b) Effects of temperature and pH value on cathepsin B activity. (c) Effects of temperature and salt content on cathepsin B activity [8].
Figure 1.
The formation of flavor substances in Jinhua ham and the interactive effect between processing parameters. (a) Formation of flavor substances in Jinhua ham and the role of endogenous protease [18]. (b) Effects of temperature and pH value on cathepsin B activity. (c) Effects of temperature and salt content on cathepsin B activity [8].


Figure 2.
The role of calpains in meat tenderization and their effect on the structure of myofibrils [23]. (a) The mechanism of meat maturation involving calpains. (b) The structure of myofibrils (myofibrillar protein related to postmortem maturation are marked separately: 1-thick myofilament; 2-thin myofilament; 3-myosin; 4-Actin; 5-Titin; 6-Tropomyosin; 7-Troponin complex; 8-Troponin.
Figure 2.
The role of calpains in meat tenderization and their effect on the structure of myofibrils [23]. (a) The mechanism of meat maturation involving calpains. (b) The structure of myofibrils (myofibrillar protein related to postmortem maturation are marked separately: 1-thick myofilament; 2-thin myofilament; 3-myosin; 4-Actin; 5-Titin; 6-Tropomyosin; 7-Troponin complex; 8-Troponin.

Figure 3.
The change trend of small peptides in Jinhua ham and the breaking sites of enzymes. (a) Comparison of changes of main small peptides relative area peaks from the three periods of Jinhua ham [48]. (b) Primary sequence of fast and slow troponin T proteins from porcine skeletal muscle. Letters in bold indicate the position of the identified peptides in the protein. Black arrows indicate endo and exo cleavage sites of the peptides identified in this study. Arrows marked with an asterisk indicate agreement with calpains cleavage site reported [51].
Figure 3.
The change trend of small peptides in Jinhua ham and the breaking sites of enzymes. (a) Comparison of changes of main small peptides relative area peaks from the three periods of Jinhua ham [48]. (b) Primary sequence of fast and slow troponin T proteins from porcine skeletal muscle. Letters in bold indicate the position of the identified peptides in the protein. Black arrows indicate endo and exo cleavage sites of the peptides identified in this study. Arrows marked with an asterisk indicate agreement with calpains cleavage site reported [51].


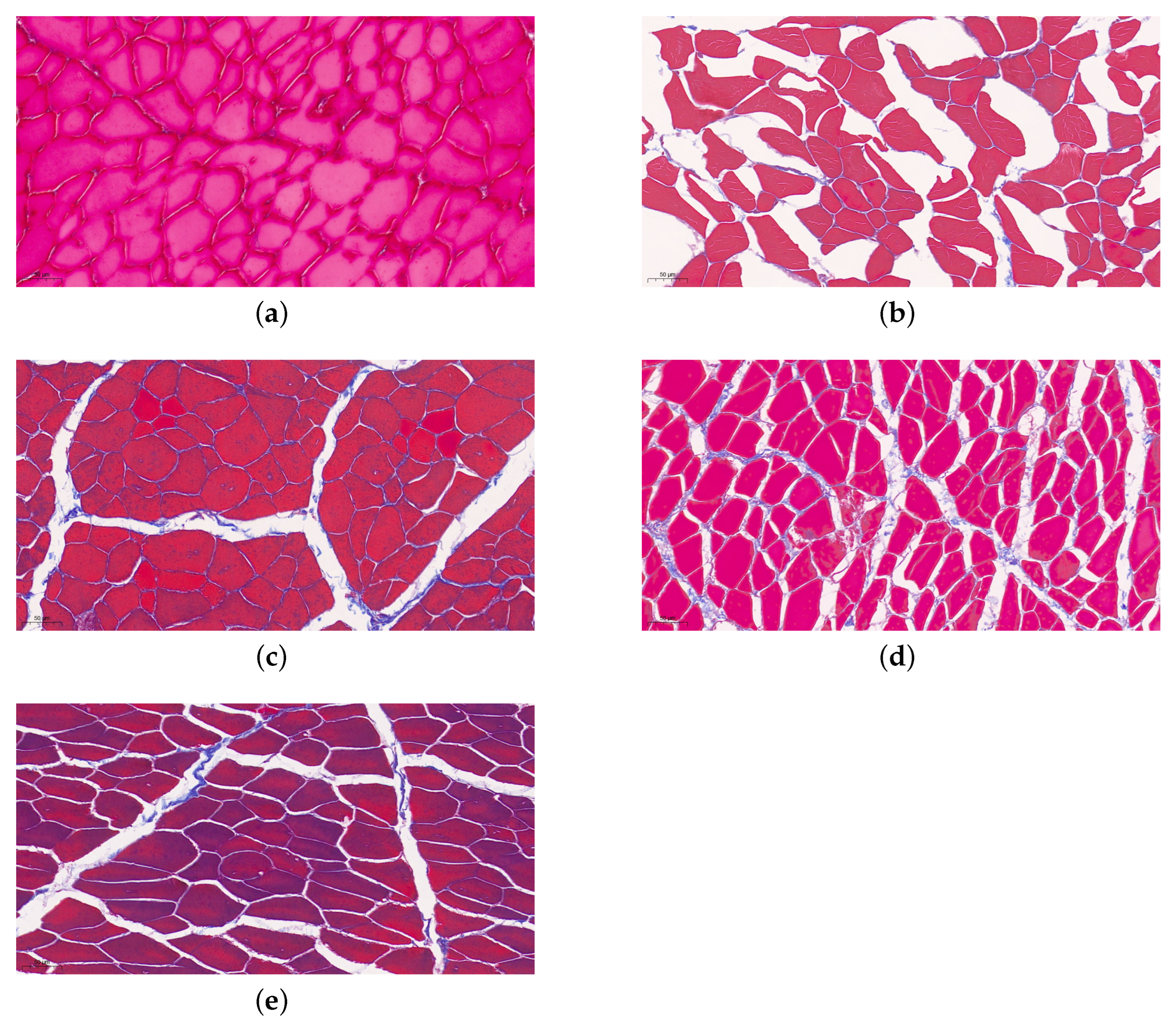
Figure 4.
Masson staining results show the changes of muscle fibers in Jinhua ham from raw ham to finished stage (20×). (a–e) Identical letters represent the period of raw ham, post-salting stage, the medium term of ripening, the latter stage of ripening and the one-year-old hams respectively [59].
Figure 4.
Masson staining results show the changes of muscle fibers in Jinhua ham from raw ham to finished stage (20×). (a–e) Identical letters represent the period of raw ham, post-salting stage, the medium term of ripening, the latter stage of ripening and the one-year-old hams respectively [59].

Figure 5.
The possible sensory defects in Jinhua ham, application and principle of ultrasound in meat products processing. (a) The possible sensory defects in Jinhua ham [66]. (b) Schematic diagram of NaCl diffusion during the salting processing in the muscle promoted by ultrasound wave. (c) The processing of the release and activation of proteases promoted by ultrasound [67,68].
Figure 5.
The possible sensory defects in Jinhua ham, application and principle of ultrasound in meat products processing. (a) The possible sensory defects in Jinhua ham [66]. (b) Schematic diagram of NaCl diffusion during the salting processing in the muscle promoted by ultrasound wave. (c) The processing of the release and activation of proteases promoted by ultrasound [67,68].



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Partial characteristics of the common lysosome cathepsins existing in animal muscles [24,27].
| Name | EC Number | Classification | Molecular Size | Optimal pH | Isoelectric Point | Feature |
|---|---|---|---|---|---|---|
| Cathepsin B | 3.4.22.1 | Cysteine protease | 25–30 KDa | 5.5–6.5 | 4.5–5.5 | Peptidyl dipeptidase activity. |
| Cathepsin L | 3.4.22.15 | Cysteine protease | 24–28 KDa | 5.5–6.5 | 5.0–6.3 | Strong proteolytic activity; weaker activity on synthetic substrates. |
| Cathepsin D | 3.4.23.5 | Aspartic protease | 30–45 KDa | 3.0–5.0 | 6.8 | One of the most abundant cathepsins in the lysosome. |
| Cathepsin H | 3.4.22.16 | Cysteine protease | 23–28 KDa | 6.5–6.8 | 6.0–7.1 | Strong aminopeptidase activity. |
| Cathepsin S | 3.4.22.27 | Cysteine protease | 24 KDa | 6.0–6.5 | 6.3–7.0 | Vigorous and stable in weak alkaline environment (pH 7.5). |
| Cathepsin E | 3.4.23.34 | Aspartic protease | 42 KDa | 3.0–3.5 | 4.1 | Exist only in macrophages of skeletal muscle. |
| Stage | Weight Loss | Temperature | Humidity | Time |
|---|---|---|---|---|
| Raw ham | 0% | 0–4 | 80–90% | 2 d |
| Salting | 0–4 | 70–80% | 15 d | |
| Pickling | 14–16% | 0–4 | 75–85% | 40 d |
| Washing and air drying | 10–15 | 60–70% | 7 d | |
| Maturation−1 | 10–20 | 65–80% | 30 d | |
| Maturation−2 | 28–30% | 18–30 | 55–65% | 40 d |
| Maturation−3 | 35–36% | 30–35 | 60–65% | 30 d |
| Post-ripening | 40–42% | 20–25 | 60–80% | 30 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hu, S.; Xu, X.; Zhang, W.; Li, C.; Zhou, G. Quality Control of Jinhua Ham from the Influence between Proteases Activities and Processing Parameters: A Review. Foods 2023, 12, 1454. https://doi.org/10.3390/foods12071454
AMA Style
Hu S, Xu X, Zhang W, Li C, Zhou G. Quality Control of Jinhua Ham from the Influence between Proteases Activities and Processing Parameters: A Review. Foods. 2023; 12(7):1454. https://doi.org/10.3390/foods12071454
Chicago/Turabian StyleHu, Shiqi, Xinglian Xu, Wangang Zhang, Chunbao Li, and Guanghong Zhou. 2023. "Quality Control of Jinhua Ham from the Influence between Proteases Activities and Processing Parameters: A Review" Foods 12, no. 7: 1454. https://doi.org/10.3390/foods12071454
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.