Quality Control of Jinhua Ham from the Influence between Proteases Activities and Processing Parameters: A Review

 , and
, and
Abstract
:1. Introduction
2. Compositions and Characteristics of Proteases
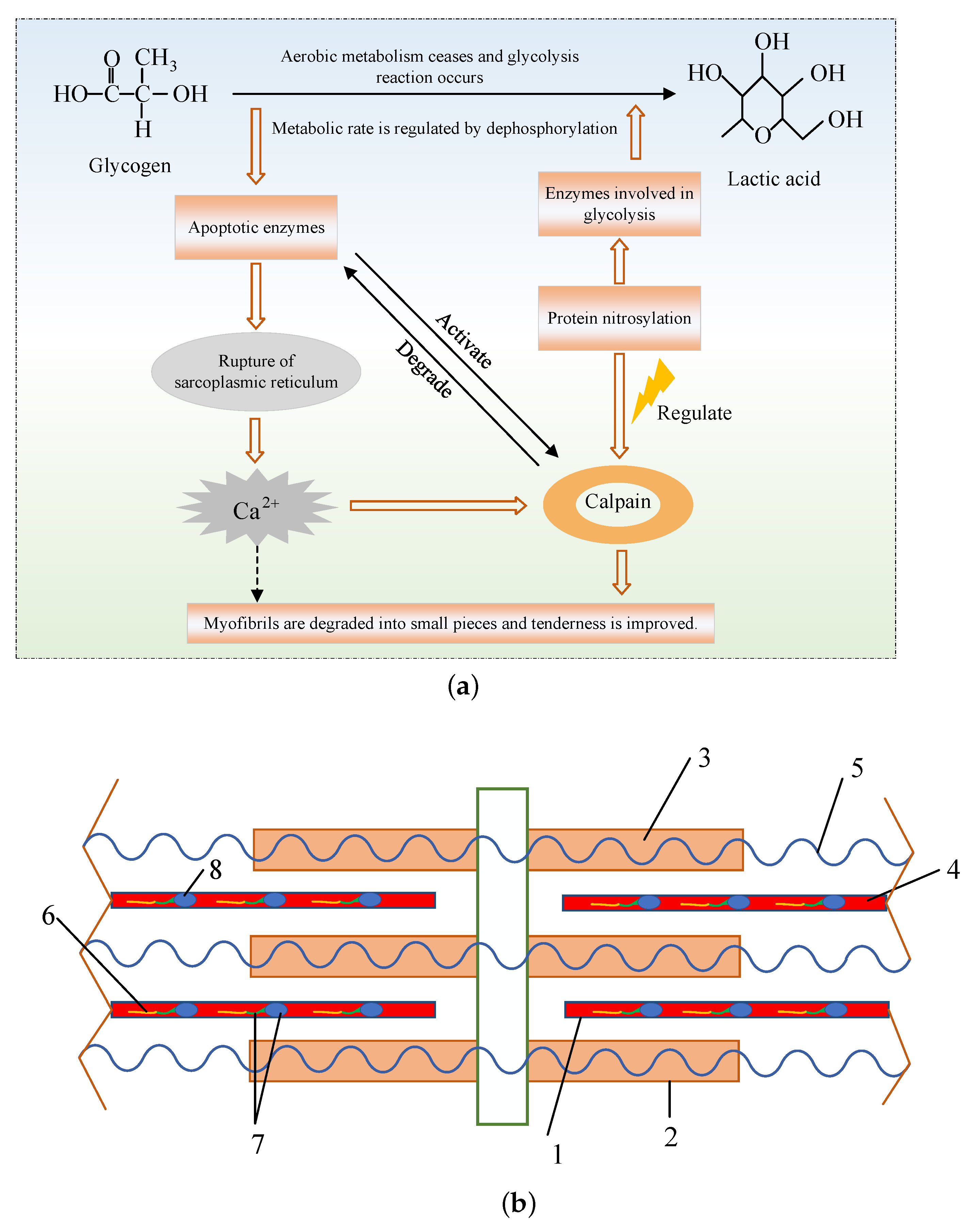
2.1. Calpains
2.2. Cathepsins
2.3. Tripeptidases and Dipeptidases
2.4. Aminopeptidases
3. Action of Proteases for Improved Ham Quality
3.1. Protein Degradation
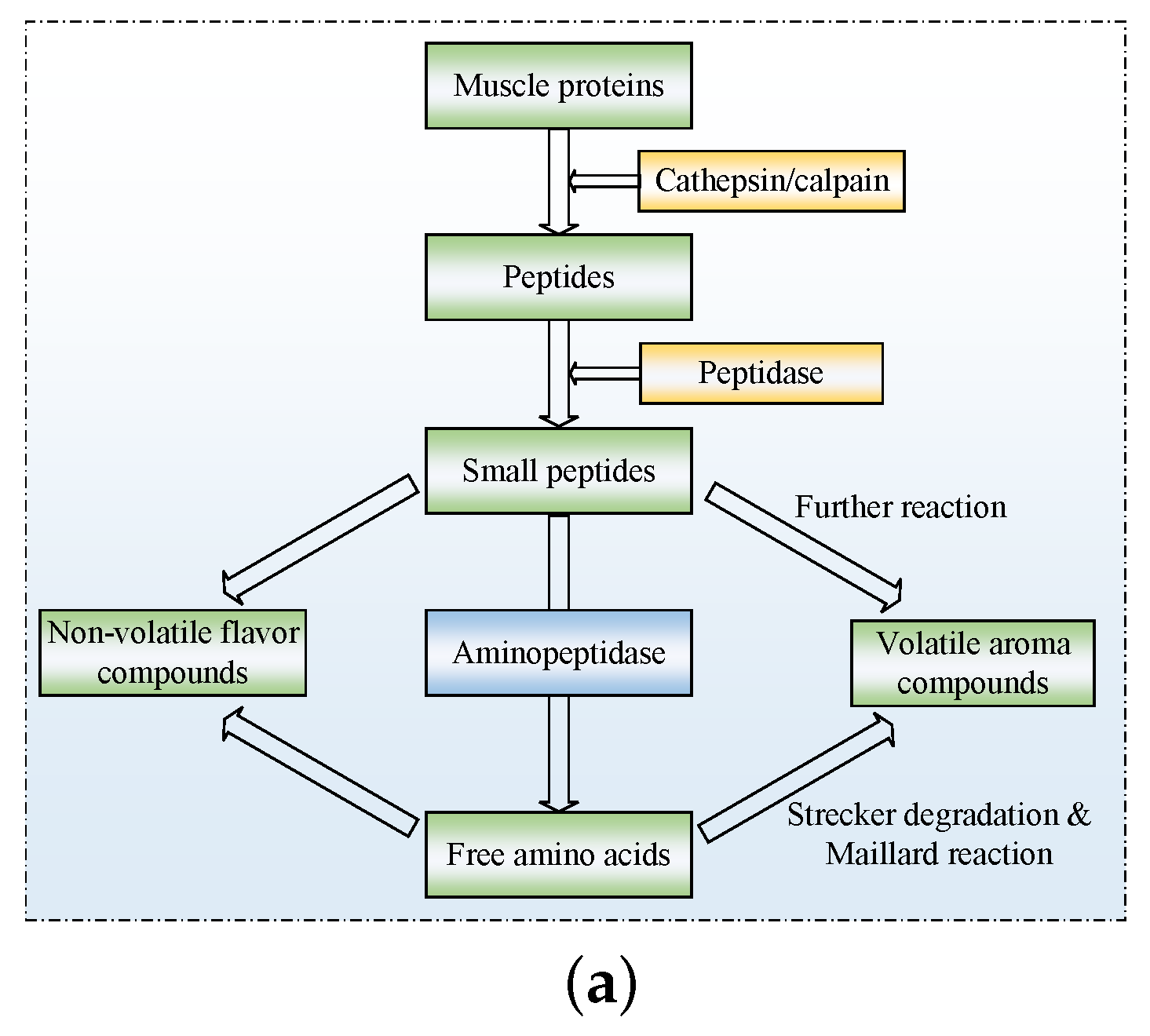
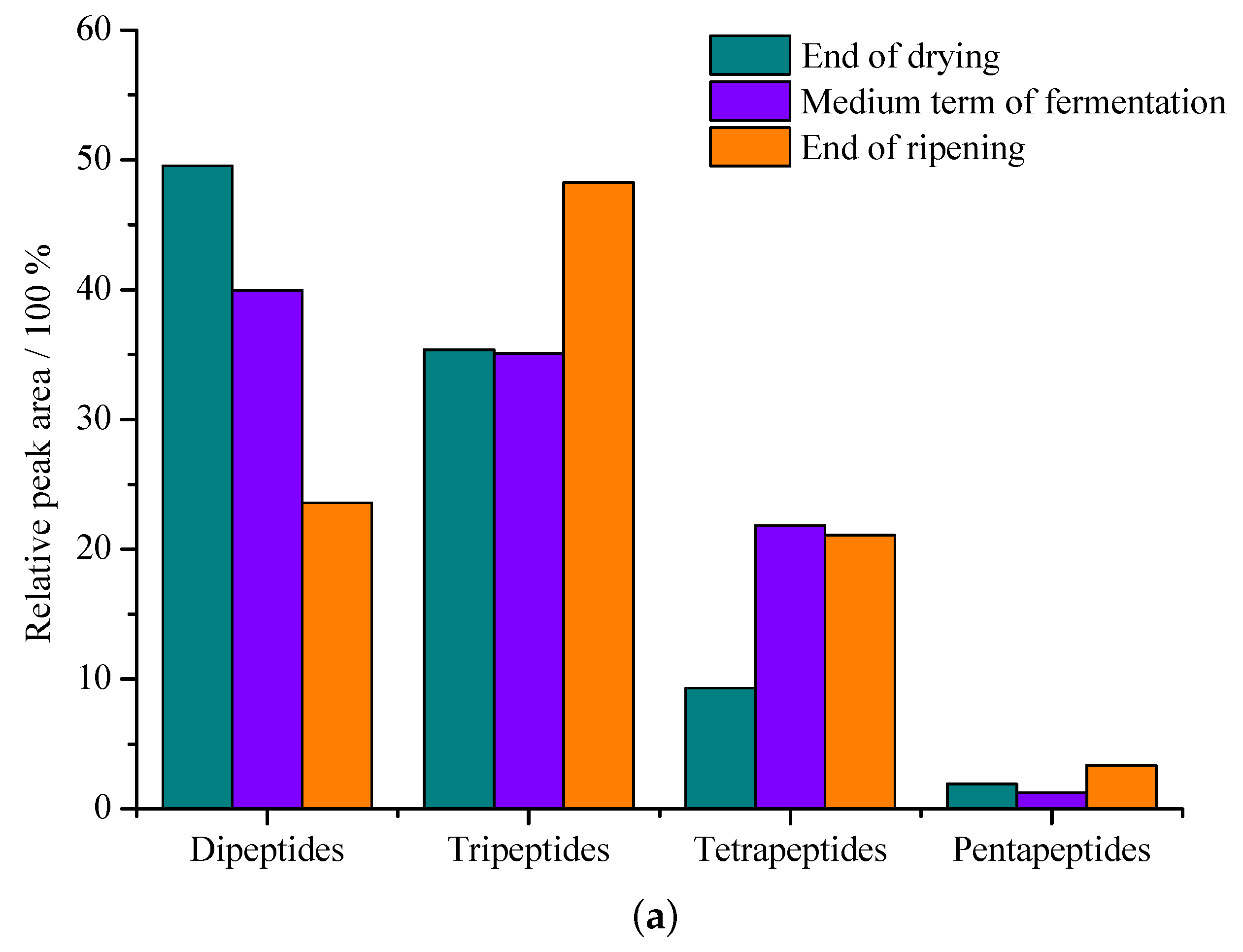
3.2. Peptide and Free Amino Acid Generation
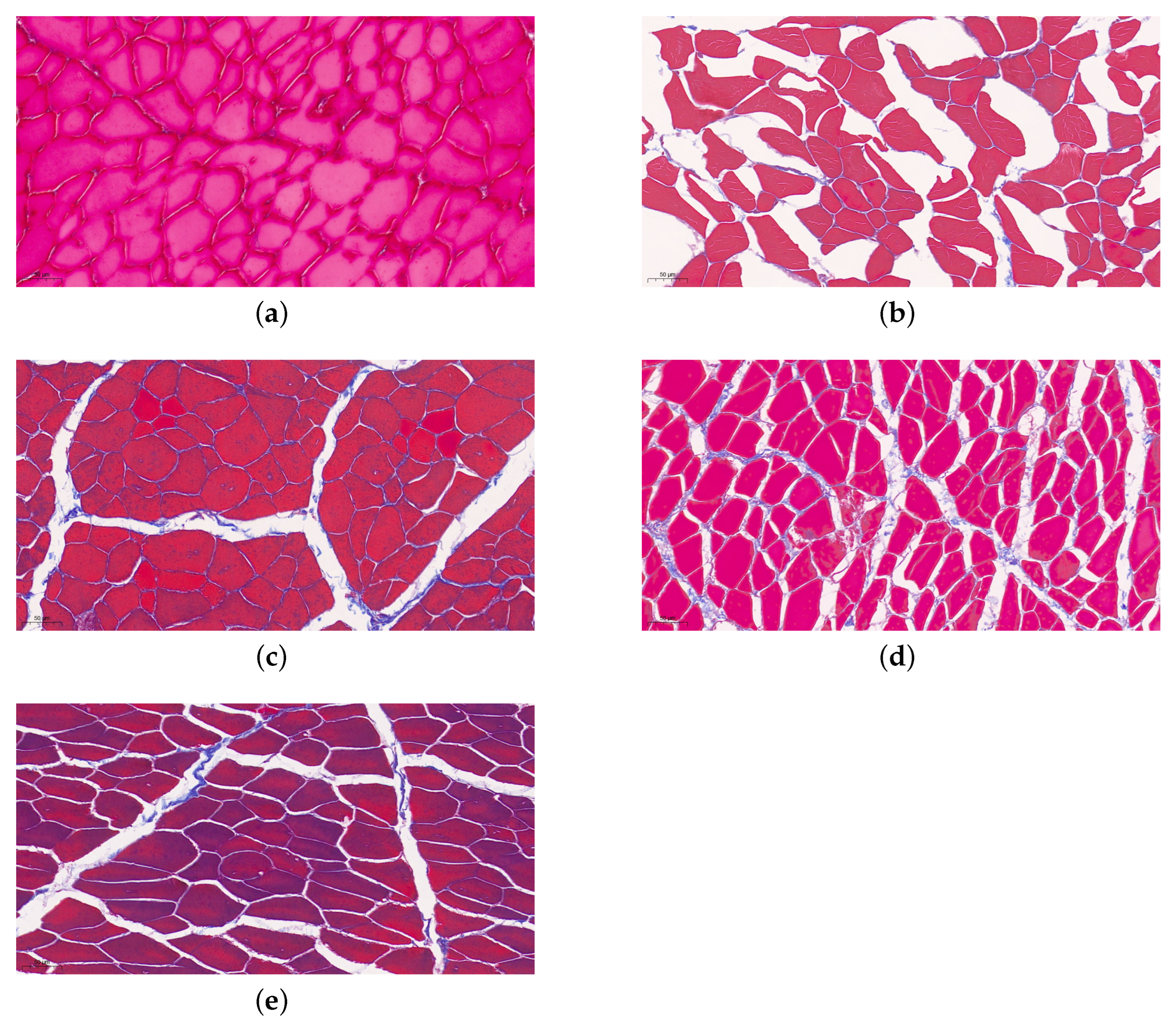
3.3. The Microstructure of Muscle
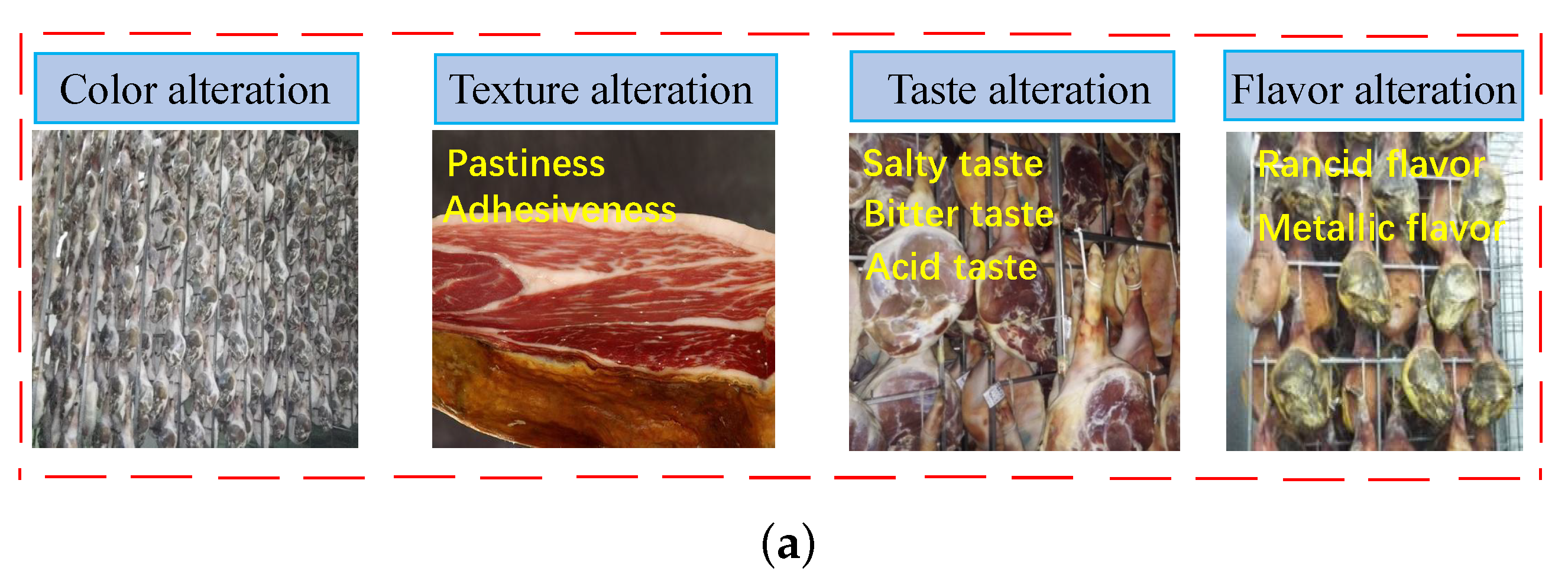
3.4. Endogenous Proteases and Texture Development
3.5. Endogenous Proteases and Flavor Development
4. Key Parameters Influencing the Activities of Muscle Endogenous Proteases
4.1. Raw Material
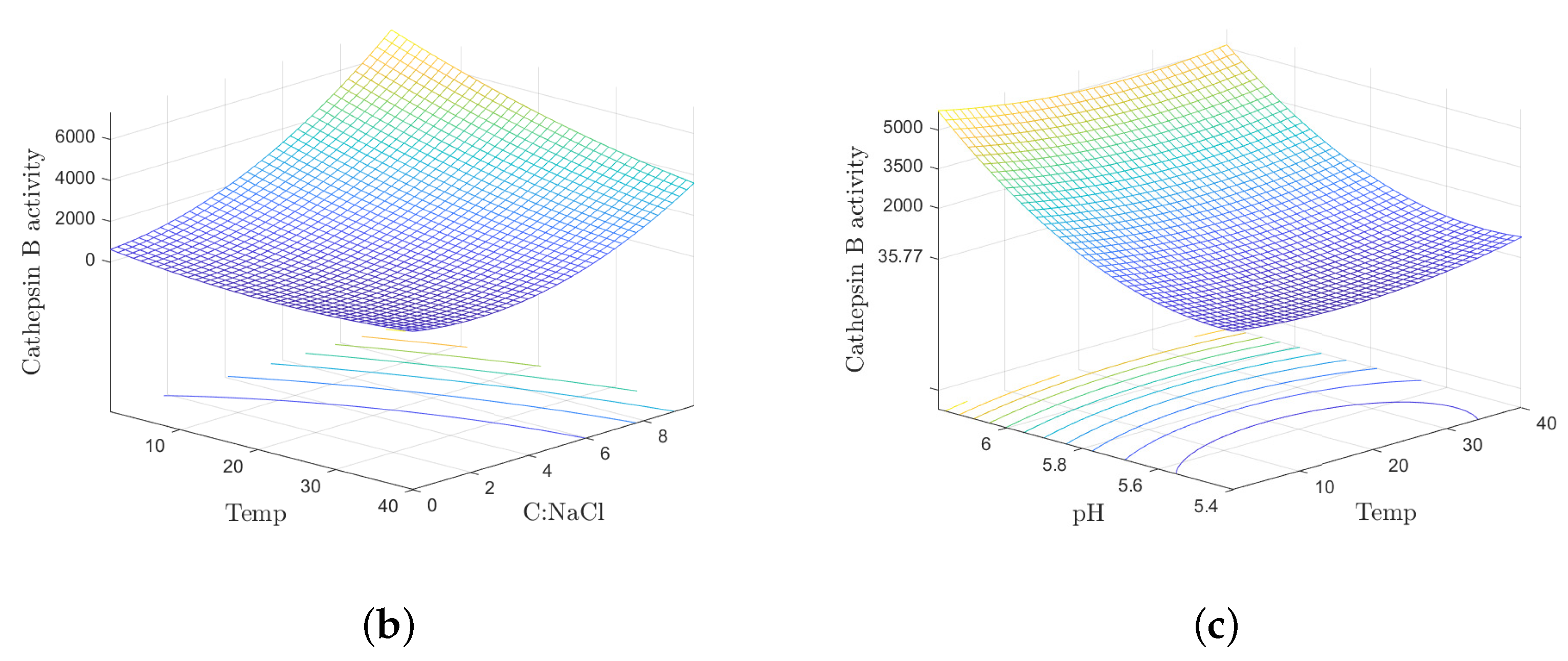
4.2. Processing Temperature
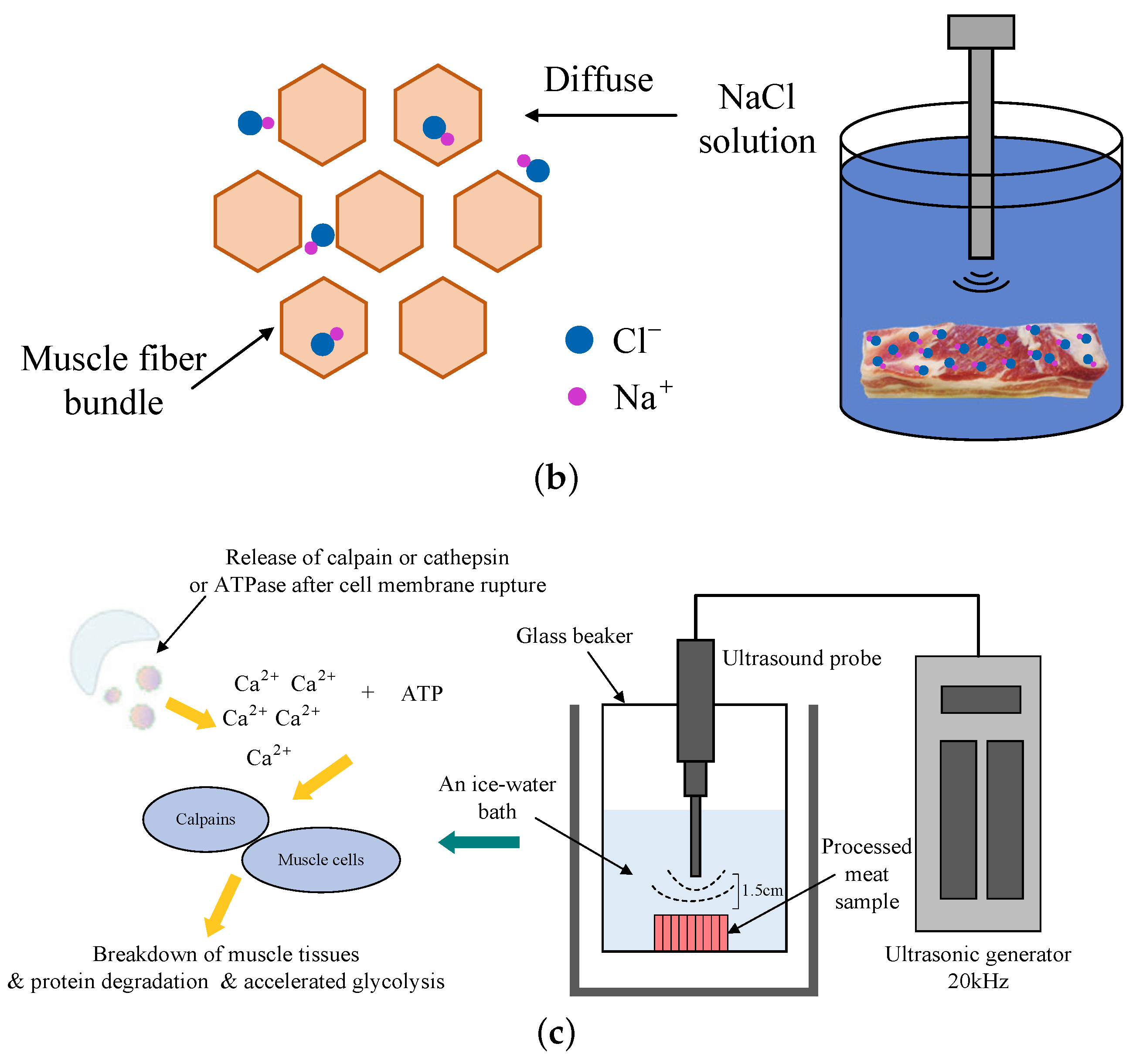
4.3. Sodium Chloride Content and Processing Time
4.4. Ultrasound-Assisted Technique and High-Pressure Processing during the Manufacture
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ramona Bosse, A.M.M.G.; Weiss, J. Recent advances in cured raw ham manufacture. Crit. Rev. Food Sci. Nutr. 2018, 4, 610–630. [Google Scholar] [CrossRef]
- Liu, D.; Bai, L.; Feng, X.; Chen, Y.P.; Zhang, D.; Yao, W.; Zhang, H.; Chen, G.; Liu, Y. Characterization of Jinhua ham aroma profiles in specific to aging time by gas chromatography-ion mobility spectrometry (GC-IMS). Meat Sci. 2020, 168, 108178. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M.; Sugawara, T.; Obiya, S.; Enomoto, A.; Kaneko, M.; Ota, S.; Soga, T.; Tomita, M. Sensory properties and metabolomic profiles of dry-cured ham during the ripening process. Food Res. Int. 2020, 129, 108850. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, Y.P.; Blank, I.; Li, F.; Li, C.; Liu, Y. GC×GC-ToF-MS and GC-IMS based volatile profile characterization of the Chinese dry-cured hams from different regions. Food Res. Int. 2021, 142, 110222. [Google Scholar] [CrossRef] [PubMed]
- MartÍn, A.; Benito, M.J.; Aranda, E.; Ruiz-Moyano, S.; Córdoba, J.J.; Córdoba, M.G. Characterization by Volatile Compounds of Microbial Deep Spoilage in Iberian Dry-Cured Ham. J. Food Sci. 2010, 75, M360–M365. [Google Scholar] [CrossRef] [PubMed]
- López-Pedrouso, M.; Pérez-Santaescolástica, C.; Franco, D.; Fulladosa, E.; Carballo, J.; Zapata, C.; Lorenzo, J.M. Comparative proteomic profiling of myofibrillar proteins in dry-cured ham with different proteolysis indices and adhesiveness. Food Chem. 2018, 244, 238–245. [Google Scholar] [CrossRef]
- Larrea, V.; Hernando, I.; Quiles, A.; Lluch, M.A.; P’erez-Munuera, I. Changes in proteins during Teruel dry-cured ham processing. Meat Sci. 2006, 74, 586–593. [Google Scholar] [CrossRef]
- Zhao, G.M.; Zhou, G.H.; Wang, Y.L.; Xu, X.L.; Huan, Y.J.; Wu, J.Q. Time-related changes in cathepsin B and L activities during processing of Jinhua ham as a function of pH, salt and temperature. Meat Sci. 2005, 70, 381–388. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Wu, J.Q.; Tang, C.B.; Li, G.; Dai, C.; Bai, Y.; Li, C.B.; Xu, X.L.; Zhou, G.H.; Cao, J.X. Comparing the proteomic profile of proteins and the sensory characteristics in Jinhua ham with different processing procedures. Food Control 2019, 106, 106694. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Bai, Y.; Wang, C.; Li, C.B.; Xu, X.L.; Pan, D.D.; Cao, J.X.; Zhou, G.H. 1H NMR-based metabolomics and sensory evaluation characterize taste substances of Jinhua ham with traditional and modern processing procedures. Food Control 2021, 126, 107873. [Google Scholar] [CrossRef]
- Arnau, J.; Serra, X.; Comaposada, J.; Gou, P.; Garriga, M. Technologies to shorten the drying period of dry-cured meat products. Meat Sci. 2007, 77, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.Y.; Wang, C.; Tang, C.B.; Dai, C.; Bai, Y.; Yu, X.B.; Li, C.B.; Xu, X.L.; Zhou, G.H.; Cao, J.X. Label-free proteomics reveals the mechanism of bitterness and adhesiveness in Jinhua ham. Food Chem. 2019, 297, 125012. [Google Scholar] [CrossRef]
- Petrova, I.; Aasen, I.M.; Rustad, T.; Eikevik, T.M. Manufacture of dry-cured ham: A review. Part 1. Biochemical changes during the technological process. Eur. Food Res. Technol. 2015, 241, 587–599. [Google Scholar] [CrossRef]
- Marušić, R.N.; Poljanec, I.; Petričević, S.; Mora, L.; Medić, H. Influence of Muscle Type on Physicochemical Parameters, Lipolysis, Proteolysis, and Volatile Compounds throughout the Processing of Smoked Dry-Cured Ham. Foods 2021, 10, 1228. [Google Scholar] [CrossRef] [PubMed]
- Xiang, A.; Wang, Y.; Sun, B.; Xu, X.; Zhou, G.; Zhou, C.; Geng, F.; Pan, D.; Cao, J. Peptidomic insights on structural protein hydrolysis by exopeptidase activities and its effects on bitterness defects of dry-cured ham. Int. J. Food Sci. Technol. 2023, 58, 755–765. [Google Scholar] [CrossRef]
- Inguglia, E.S.; Zhang, Z.; Tiwari, B.K.; Kerry, J.P.; Burgess, C.M. Salt reduction strategies in processed meat products—A review. Trends Food Sci. Technol. 2017, 59, 70–78. [Google Scholar] [CrossRef]
- Petrova, I.; Bantle, M.; Eikevik, T.M. Manufacture of dry-cured ham: A review. Part 2. Drying kinetics, modeling and equipment. Eur. Food Res. Technol. 2015, 241, 447–458. [Google Scholar] [CrossRef]
- Zhou, G.H.; Zhao, G.M. Biochemical changes during processing of traditional Jinhua ham. Meat Sci. 2007, 77, 114–120. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Wang, Y.; Pan, D.D.; Cao, J.X.; Chen, Y.J.; Liu, Y.; Sun, Y.Y.; Ou, C.R. The changes in the proteolysis activity and the accumulation of free amino acids during chinese traditional dry-cured loins processing. Food Sci. Biotechnol. 2017, 26, 679–687. [Google Scholar] [CrossRef]
- Dang, Y.; Gao, X.; Ma, F.; Wu, X. Comparison of umami taste peptides in water-soluble extractions of Jinhua and Parma hams. LWT-Food Sci. Technol. 2015, 60, 1179–1186. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Pan, D.D.; Bai, Y.; Li, C.B.; Xu, X.L.; Zhou, G.H.; Cao, J.X. Evaluating endogenous protease of salting exudates during the salting process of Jinhua ham. LWT-Food Sci. Technol. 2019, 101, 76–82. [Google Scholar] [CrossRef]
- Zhu, C.Z.; Tian, W.; Li, M.Y.; Liu, Y.X.; Zhao, G.M. Separation and identification of peptides from dry-cured Jinhua ham. Int. J. Food Prop. 2018, 20, S2980–S2989. [Google Scholar] [CrossRef]
- Chéret, R.; Delbarre-Ladrat, C.; Lamballerie-Anton, M.D.; Verrez-Bagnis, V. Calpain and cathepsin activities in post mortem fish and meat muscles. Food Chem. 2007, 101, 1474–1479. [Google Scholar] [CrossRef] [Green Version]
- Venugopal, A.; Siva Kumar, N. Biochemical characterization of cathepsin D from the mussel Lamellidens corrianus. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2014, 169, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.Y.; Chang, X.N.; Bao, S.S.; Song, L.; Zhu, B.W.; Dong, X.P.; Zong, Y.; Li, D.M.; Zhang, M.M.; Liu, Y.X.; et al. Purification and partial characterisation of a cathepsin L-like proteinase from sea cucumber (Stichopus japonicus) and its tissue distribution in body wall. Food Chem. 2014, 158, 192–199. [Google Scholar] [CrossRef]
- Zhao, G.M.; Tian, W.; Liu, Y.X.; Zhou, G.H.; Xu, X.L.; Li, M.Y. Proteolysis in biceps femoris during Jinhua ham processing. Meat Sci. 2008, 79, 39–45. [Google Scholar] [CrossRef]
- Wu, Y.; Cao, S.M. Study on endogenous protease and protein degradation of dry-salted Decapterus maruadsi. CyTA-J. Food 2018, 16, 350–356. [Google Scholar] [CrossRef] [Green Version]
- Zhong, C.; Cai, Q.F.; Liu, G.M.; Sun, L.C.; Hara, K.; Su, W.J.; Cao, M.J. Purification and characterisation of cathepsin L from the skeletal muscle of blue scad (Decapterus maruadsi) and comparison of its role with myofibril-bound serine proteinase in the degradation of myofibrillar proteins. Food Chem. 2012, 133, 1560–1568. [Google Scholar] [CrossRef]
- Ge, L.; Xu, Y.; Xia, W.; Jiang, Q. Synergistic action of cathepsin B, L, D and calpain in disassembly and degradation of myofibrillar protein of grass carp. Food Res. Int. 2018, 109, 481–488. [Google Scholar] [CrossRef]
- Kaur, L.; Hui, S.X.; Boland, M. Changes in Cathepsin Activity during Low-Temperature Storage and Sous Vide Processing of Beef Brisket. Food Sci. Anim. Resour. 2020, 40, 415–425. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Xia, Q.; He, J.; Sun, Y.Y.; Dang, Y.L.; Zhou, G.H.; Geng, F.; Pan, D.D.; Cao, J.X. Insights into ultrasonic treatment on the mechanism of proteolysis and taste improvement of defective dry-cured ham. Food Chem. 2022, 388, 133059. [Google Scholar] [CrossRef] [PubMed]
- Sárraga, C.; Gil, M.; GarcÍa-Regueiro, J.A. Comparison of calpain and cathepsin (B, L and D) activities during dry-cured ham processing from heavy and light large white pigs. J. Science Food Agric. 1993, 1, 71–75. [Google Scholar] [CrossRef]
- Gallego, M.; Mora, L.; Toldrá, F. The relevance of dipeptides and tripeptides in the bioactivity and taste of dry-cured ham. Food Prod. Process. Nutr. 2019, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Sentandreu, M.; Toldrá, F. Dipeptidyl peptidase activities along the processing of Serrano dry-cured ham. Eur. Food Res. Technol. 2001, 213, 83–87. [Google Scholar] [CrossRef]
- Mora, L.; Sentandreu, M.A.; Koistinen, K.M.; Fraser, P.D.; Toldrá, F.; Bramley, P.M. Naturally Generated Small Peptides Derived from Myofibrillar Proteins in Serrano Dry-Cured Ham. J. Agric. Food Chem. 2009, 57, 3228–3234. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, Y.; Zhang, J.; Li, X.; Lin, Z.; Ma, C. Proteolysis, protein oxidation and protease activity in dry-cured Xuanwei ham during the salting stages. Int. J. Food Sci. Technol. 2011, 46, 1370–1377. [Google Scholar] [CrossRef]
- Iwaniak, A.; Hrynkiewicz, M.; Bucholska, J.; Minkiewicz, P.; Darewicz, M. Understanding the nature of bitter-taste di- and tripeptides derived from food proteins based on chemometric analysis. J. Food Biochem. 2019, 43, e12500. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, P.; Muenzner, M.; Kästorf, M.; Santos, K.; Hartmann, T.; Dienelt, A.; Rehfeld, L.; Bergmann, A. A novel and highly efficient purification procedure for native human dipeptidyl peptidase 3 from human blood cell lysate. PLoS ONE 2019, 14, e0220866. [Google Scholar] [CrossRef] [Green Version]
- Toldrá, F.; Aristoy, M.C.; Flores, M. Contribution of muscle aminopeptidases to flavor development in dry-cured ham. Food Res. Int. 2000, 33, 181–185. [Google Scholar] [CrossRef]
- Petrova, I.; Tolstorebrov, I.; Mora, L.; Toldrá, F.; Eikevik, T.M. Evolution of proteolytic and physico-chemical characteristics of Norwegian dry-cured ham during its processing. Meat Sci. 2016, 121, 243–249. [Google Scholar] [CrossRef] [Green Version]
- Toldrá, F.; Flores, M. The role of muscle proteases and lipases in flavor development during the processing of dry-cured ham. Crit. Rev. Food Sci. Nutr. 1998, 38, 331–352. [Google Scholar] [CrossRef] [PubMed]
- Toldrá, F. The role of muscle enzymes in dry-cured meat products with different drying conditions. Trends Food Sci. Technol. 2006, 17, 164–168. [Google Scholar] [CrossRef]
- Sforza, S.; Pigazzani, A.; Motti, M.; Porta, C.; Virgili, R.; Galaverna, G.; Dossena, A.; Marchelli, R. Oligopeptides and free amino acids in Parma hams of known cathepsin B activity. Food Chem. 2001, 75, 267–273. [Google Scholar] [CrossRef]
- Zhao, G.M.; Zhou, G.H.; Tian, W.; Xu, X.L.; Wang, Y.L.; Luo, X. Changes of alanyl aminopeptidase activity and free amino acid contents in biceps femoris during processing of Jinhua ham. Meat Sci. 2005, 71, 612–619. [Google Scholar] [CrossRef]
- Yang, H.; Ma, C.; Qiao, F.; Song, Y.; Du, M. Lipolysis in intramuscular lipids during processing of traditional Xuanwei ham. Meat Sci. 2005, 71, 670–675. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Le, Y.; Zheng, Y.Y.; Wang, J.J.; Li, G.; Bai, Y.; Li, C.B.; Xu, X.L.; Zhou, G.H.; Cao, J.X. Characterizing the effect of free amino acids and volatile compounds on excessive bitterness and sourness in defective dry-cured ham. LWT-Food Sci. Technol. 2020, 123, 109071. [Google Scholar] [CrossRef]
- Marušić, N.; Vidaček, S.; Janči, T.; Petrak, T.; Medič, H. Determination of volatile compounds and quality parameters of traditional Istrian dry-cured ham. Meat Sci. 2014, 96, 1409–1416. [Google Scholar] [CrossRef]
- Xing, L.; Liu, R.; Gao, X.; Zheng, J.; Wang, C.; Zhou, G.; Zhang, W. The proteomics homology of antioxidant peptides extracted from dry-cured Xuanwei and Jinhua ham. Food Chem. 2018, 266, 420–426. [Google Scholar] [CrossRef]
- Huan, Y.; Zhou, G.; Zhao, G.; Xu, X.; Peng, Z. Changes in flavor compounds of dry-cured Chinese Jinhua ham during processing. Meat Sci. 2005, 71, 291–299. [Google Scholar] [CrossRef]
- Mora, L.; Gallego, M.; Aristoy, M.C.; Fraser, P.D.; Toldrá, F. Peptides naturally generated from ubiquitin-60S ribosomal protein as potential biomarkers of dry-cured ham processing time. Food Control 2015, 48, 102–107. [Google Scholar] [CrossRef] [Green Version]
- Mora, L.; Gallego, M.; Toldrá, F. Degradation of myosin heavy chain and its potential as a source of natural bioactive peptides in dry-cured ham. Food Biosci. 2019, 30, 100416. [Google Scholar] [CrossRef]
- Gallego, M.; Toldr’a, F.; Mora, L. Quantification and in silico analysis of taste dipeptides generated during dry-cured ham processing. Food Chem. 2022, 370, 130977. [Google Scholar] [CrossRef] [PubMed]
- Aristoy, M.C.; Fidel, T. Isolation of flavor peptides from raw pork meat and dry-cured ham. Dev. Food Sci. 1995, 37, 1323–1344. [Google Scholar]
- Mora, L.; Gallego, M.; Escudero, E.; Reig, M.; Aristoy, M.C.; Toldrá, F. Small peptides hydrolysis in dry-cured meats. Int. J. Food Microbiol. 2015, 212, 9–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arroyo-Maya, I.J.; Campos-Terán, J.; Hernández-Arana, A.; McClements, D.J. Characterization of flavonoid-protein interactions using fluorescence spectroscopy: Binding of pelargonidin to dairy proteins. Food Chem. 2016, 213, 431–439. [Google Scholar] [CrossRef]
- Wu, M.; Xiong, Y.L.; Chen, J.; Tang, X.; Zhou, G. Rheological and Microstructural Properties of Porcine Myofibrillar Protein-Lipid Emulsion Composite Gels. J. Food Sci. 2009, 74, E207–E217. [Google Scholar] [CrossRef]
- Sun, W.; Zhou, F.; Sun, D.W.; Zhao, M. Effect of Oxidation on the Emulsifying Properties of Myofibrillar Proteins. Food Bioprocess Technol. 2012, 6, 1703–1712. [Google Scholar] [CrossRef]
- Harkouss, R.; Astruc, T.; Lebert, A.; Gatellier, P.; Loison, O.; Safa, H.; Portanguen, S.; Parafita, E.; Mirade, P.S. Quantitative study of the relationships among proteolysis, lipid oxidation, structure and texture throughout the dry-cured ham process. Food Chem. 2015, 166, 522–530. [Google Scholar] [CrossRef]
- Hu, S.; Zhou, G.; Xu, X.; Zhang, W.; Li, C. Contribution of cathepsin B and L to endogenous proteolysis in the course of modern Jinhua ham processing. Food Control 2021, 135, 108584. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Y.; He, J.; Pan, D.; Wang, H.; Cao, J. Evaluating the profile of myofibrillar proteins and its relationship with tenderness among five styles of dry-cured hams. Int. J. Food Sci. Technol. 2021, 56, 259–268. [Google Scholar] [CrossRef]
- Hernández-Ramos, P.; Vivar-Quintana, A.M.; Revilla, I.; González-Martín, M.I.; Hernández-Jiménez, M.; Martínez-Martín, I. Prediction of Sensory Parameters of Cured Ham: A Study of the Viability of the Use of NIR Spectroscopy and Artificial Neural Networks. Sensors 2020, 20, 5624. [Google Scholar] [CrossRef] [PubMed]
- Coll-Brasas, E.; Laguna, L.; Tárrega, A.; Arnau, J.; Claret, A.; Fulladosa, E. Evaluation of dry-cured ham pastiness through rheological measurements of aqueous extracts obtained in vitro mimicking the mastication process. LWT-Food Sci. Technol. 2021, 145, 111272. [Google Scholar] [CrossRef]
- Contreras, M.; Benedito, J.; Quiles, A.; Lorenzo, J.M.; Fulladosa, E.; Gou, P.; Garcia-Perez, J.V. Assessing the textural defect of pastiness in dry-cured pork ham using chemical, microstructural, textural and ultrasonic analyses. J. Food Eng. 2020, 265, 109690. [Google Scholar] [CrossRef]
- Hu, S.; Zhou, G.; Xu, X.; Zhang, W.; Li, C. Insight into the impacts of Jinhua ham processing conditions on cathepsin B activity and conformation changes based on molecular simulation. LWT-Food Sci. Technol. 2023, 174, 114459. [Google Scholar] [CrossRef]
- Aaslyng, M.D.; Vestergaard, C.; Koch, A.G. The effect of salt reduction on sensory quality and microbial growth in hotdog sausages, bacon, ham and salami. Meat Sci. 2014, 96, 47–55. [Google Scholar] [CrossRef]
- Zhou, C.; Xia, Q.; L, D.; He, J.; Sun, Y.; Dang, Y.; Geng, F.; Pan, D.; Cao, J.; Zhou, G. Recent developments in off-odor formation mechanism and the potential regulation by starter cultures in dry-cured ham. Crit. Rev. Food Sci. Nutr. 2022, 9, 1–15. [Google Scholar] [CrossRef]
- López-Pedrouso, M.; Pérez-Santaescolástica, C.; Franco, D.; Carballo, J.; Garcia-Perez, J.V.; Benedito, J.; Zapata, C.; Lorenzo, J.M. Proteomic footprint of ultrasound intensification on sliced dry-cured ham subjected to mild thermal conditions. J. Proteom. 2019, 193, 123–130. [Google Scholar] [CrossRef]
- Kang, D.c.; Wang, A.r.; Zhou, G.h.; Zhang, W.g.; Xu, S.m.; Guo, G.p. Power ultrasonic on mass transport of beef: Effects of ultrasound intensity and NaCl concentration. Innov. Food Sci. Emerg. Technol. 2016, 35, 36–44. [Google Scholar] [CrossRef]
- Virgili, R.; Schivazappa, C.; Parolari, G.; Bordini, C.S.; Degni, M. Proteases in fresh pork muscle and their influence on bitter taste formation in dry-cured ham. J. Food Biochem. 1998, 22, 53–63. [Google Scholar] [CrossRef]
- Jiménez-Colmenero, F.; Ventanas, J.; Toldrá, F. Nutritional composition of dry-cured ham and its role in a healthy diet. Meat Sci. 2010, 84, 585–593. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Tang, C.B.; Wang, C.; Dai, C.; Bai, Y.; Yu, X.B.; Li, C.B.; Xu, X.L.; Zhou, G.H.; Cao, J.X. Insights into the evolution of myosin light chain isoforms and its effect on sensory defects of dry-cured ham. Food Chem. 2020, 315, 126318. [Google Scholar] [CrossRef] [PubMed]
- GarcÍa-Garrido, J.A.; Quiles-Zafra, R.; Tapiador, J.; Luque De Castro, M.D. Activity of cathepsin B, D, H and L in Spanish dry-cured ham of normal and defective texture. Meat Sci. 2000, 56, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-RamÍrez, J.; Arnau, J.; Serra, X.; Gou, P. Effect of pH24, NaCl content and proteolysis index on the relationship between water content and texture parameters in biceps femoris and semimembranosus muscles in dry-cured ham. Meat Sci. 2006, 72, 185–194. [Google Scholar] [CrossRef]
- Bañón, S.; Gil, M.D.; Garrido, M.D. The effects of castration on the eating quality of dry-cured ham. Meat Sci. 2003, 3, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Čandek-Potokar, M.; Škrlep, M. Factors in pig production that impact the quality of dry-cured ham: A review. Animal 2012, 6, 327–338. [Google Scholar] [CrossRef]
- Guerrero, L.; Gobantes, I.; Oliver, M.A.; Arnau, J.; Dolors Guȧrdia, M.; Elvira, J.; Riu, P.; Grėbol, N.; Monfort, J.M. Green hams electrical impedance spectroscopy (EIS) measures and pastiness prediction of dry cured hams. Meat Sci. 2004, 66, 289–294. [Google Scholar] [CrossRef]
- Moin, K.; Day, N.A.; Sameni, M.; Hasnain, S.; Hirama, T.; Sloane, B.F. Human tumour cathepsin B. Comparison with normal liver cathepsin B. Biochem. J. 1992, 285, 427–434. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Santaescolástica, C.; Carballo, J.; Fulladosa, E.; Garcia-Perez José, V.; Benedito, J.; Lorenzo, J.M. Application of temperature and ultrasound as corrective measures to decrease the adhesiveness in dry-cured ham. Influence on free amino acid and volatile compound profile. Food Res. Int. 2018, 114, 140–150. [Google Scholar] [CrossRef]
- Zhou, C.; Zhan, G.; Pan, D.; Zhou, G.; Wang, Y.; He, J.; Cao, J. Charactering the spoilage mechanism of “three sticks” of Jinhua ham. Food Sci. Hum. Wellness 2022, 11, 1322–1330. [Google Scholar] [CrossRef]
- Bhat, Z.F.; Morton, J.D.; Mason, S.L.; Bekhit, A.E.D.A. The application of pulsed electric field as a sodium reducing strategy for meat products. Food Chem. 2020, 306, 125622. [Google Scholar] [CrossRef]
- Caballero, D.; Antequera, T.; Caro, A.; Duran, M.L.; Perez-Palacios, T. Data Mining on MRI-Computational Texture Features to Predict Sensory Characteristics in Ham. Food Bioprocess Technol. 2016, 9, 699–708. [Google Scholar] [CrossRef]
- Zhang, J.; Zhen, Z.; Zhang, W.; Zeng, T.; Zhou, G. Effect of intensifying high-temperature ripening on proteolysis, lipolysis and flavor of Jinhua ham. J. Sci. Food Agric. 2009, 89, 834–842. [Google Scholar] [CrossRef]
- Ding, X.; Wang, G.; Zou, Y.; Zhao, Y.; Ge, C.; Liao, G. Evaluation of small molecular metabolites and sensory properties of Xuanwei ham salted with partial replacement of NaCl by KCl. Meat Sci. 2021, 175, 108465. [Google Scholar] [CrossRef] [PubMed]
- Salazar, E.; Cayuela, J.M.; Abellán, A.; Bueno-Gavilá, E.; Tejada, L. Fatty Acids and Free Amino Acids Changes during Processing of a Mediterranean Native Pig Breed Dry-Cured Ham. Foods 2020, 9, 1170. [Google Scholar] [CrossRef]
- Pugliese, C.; Sirtori, F.; Škrlep, M.; Piasentier, E.; Calamai, L.; Franci, O.; Čandek Potokar, M. The effect of ripening time on the chemical, textural, volatile and sensorial traits of Bicep femoris and Semimembranosus muscles of the Slovenian dry-cured ham Kraški pršut. Meat Sci. 2015, 100, 58–68. [Google Scholar] [CrossRef]
- Sforza, S.; Galaverna, G.; Schivazappa, C.; Marchelli, R.; Dossena, A.; Virgili, R. Effect of extended aging of parma dry-cured ham on the content of oligopeptides and free amino acids. J. Agric. Food Chem. 2006, 54, 9422–9429. [Google Scholar] [CrossRef]
- Kang, Z.L.; Zou, Y.F.; Xu, X.L.; Zhu, C.Z.; Wang, P.; Zhou, G.H. Effect of a beating process, as a means of reducing salt content in Chinese-style meatballs (kung-wan): A physico-chemical and textural study. Meat Sci. 2014, 96, 147–152. [Google Scholar] [CrossRef]
- Zhang, J.; Toldrá, F.; Zhang, W.; Yin, Y.; Zhu, Z. Study on the effects and mechanisms of ultrasound on the peptide profile and taste of unsmoked bacon using peptidomics and bioinformatics. Food Chem. 2023, 414, 135764. [Google Scholar] [CrossRef]
- Wang, L.; Li, J.; Teng, S.; Zhang, W.; Purslow, P.P.; Zhang, R. Changes in collagen properties and cathepsin activity of beef M. semitendinosus by the application of ultrasound during post-mortem aging. Meat Sci. 2022, 185, 108718. [Google Scholar] [CrossRef]
- Buckow, R.; Sikes, A.; Tume, R. Effect of High Pressure on Physicochemical Properties of Meat. Crit. Rev. Food Sci. Nutr. 2013, 53, 770–786. [Google Scholar] [CrossRef]
- Coll-Brasas, E.; Arnau, J.; Gou, P.; Lorenzo, J.M.; GarcÍa-Pérez, J.V.; Fulladosa, E. Effect of high pressure processing temperature on dry-cured hams with different textural characteristics. Meat Sci. 2019, 152, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Cañedo, A.; Martínez-Onandi, N.; Gaya, P.; Nuñez, M.; Picon, A. Effect of high-pressure processing and chemical composition on lipid oxidation, aminopeptidase activity and free amino acids of Serrano dry-cured ham. Meat Sci. 2021, 172, 108349. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.Y.; Pan, D.D.; Cao, J.X.; Zhou, G.H. A comprehensive review on molecular mechanism of defective dry-cured ham with excessive pastiness, adhesiveness, and bitterness by proteomics insights. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3838–3857. [Google Scholar] [CrossRef] [PubMed]
- Pingret, D.; Fabiano-Tixier, A.S.; Chemat, F. Degradation during application of ultrasound in food processing: A review. Food Control 2013, 31, 593–606. [Google Scholar] [CrossRef]
- Kang, D.C.; Zou, Y.H.; Cheng, Y.P.; Xing, L.J.; Zhou, G.H.; Zhang, W.G. Effects of power ultrasound on oxidation and structure of beef proteins during curing processing. Ultrason. Sonochem. 2016, 33, 47–53. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | EC Number | Classification | Molecular Size | Optimal pH | Isoelectric Point | Feature |
|---|---|---|---|---|---|---|
| Cathepsin B | 3.4.22.1 | Cysteine protease | 25–30 KDa | 5.5–6.5 | 4.5–5.5 | Peptidyl dipeptidase activity. |
| Cathepsin L | 3.4.22.15 | Cysteine protease | 24–28 KDa | 5.5–6.5 | 5.0–6.3 | Strong proteolytic activity; weaker activity on synthetic substrates. |
| Cathepsin D | 3.4.23.5 | Aspartic protease | 30–45 KDa | 3.0–5.0 | 6.8 | One of the most abundant cathepsins in the lysosome. |
| Cathepsin H | 3.4.22.16 | Cysteine protease | 23–28 KDa | 6.5–6.8 | 6.0–7.1 | Strong aminopeptidase activity. |
| Cathepsin S | 3.4.22.27 | Cysteine protease | 24 KDa | 6.0–6.5 | 6.3–7.0 | Vigorous and stable in weak alkaline environment (pH 7.5). |
| Cathepsin E | 3.4.23.34 | Aspartic protease | 42 KDa | 3.0–3.5 | 4.1 | Exist only in macrophages of skeletal muscle. |
| Stage | Weight Loss | Temperature | Humidity | Time |
|---|---|---|---|---|
| Raw ham | 0% | 0–4 | 80–90% | 2 d |
| Salting | 0–4 | 70–80% | 15 d | |
| Pickling | 14–16% | 0–4 | 75–85% | 40 d |
| Washing and air drying | 10–15 | 60–70% | 7 d | |
| Maturation−1 | 10–20 | 65–80% | 30 d | |
| Maturation−2 | 28–30% | 18–30 | 55–65% | 40 d |
| Maturation−3 | 35–36% | 30–35 | 60–65% | 30 d |
| Post-ripening | 40–42% | 20–25 | 60–80% | 30 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, S.; Xu, X.; Zhang, W.; Li, C.; Zhou, G. Quality Control of Jinhua Ham from the Influence between Proteases Activities and Processing Parameters: A Review. Foods 2023, 12, 1454. https://doi.org/10.3390/foods12071454
Hu S, Xu X, Zhang W, Li C, Zhou G. Quality Control of Jinhua Ham from the Influence between Proteases Activities and Processing Parameters: A Review. Foods. 2023; 12(7):1454. https://doi.org/10.3390/foods12071454
Chicago/Turabian StyleHu, Shiqi, Xinglian Xu, Wangang Zhang, Chunbao Li, and Guanghong Zhou. 2023. "Quality Control of Jinhua Ham from the Influence between Proteases Activities and Processing Parameters: A Review" Foods 12, no. 7: 1454. https://doi.org/10.3390/foods12071454