
Rutin Concentration and Characterization of Rutinosidase in Perennial Buckwheat (Fagopyrum cymosum) and Its Application in Foods
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
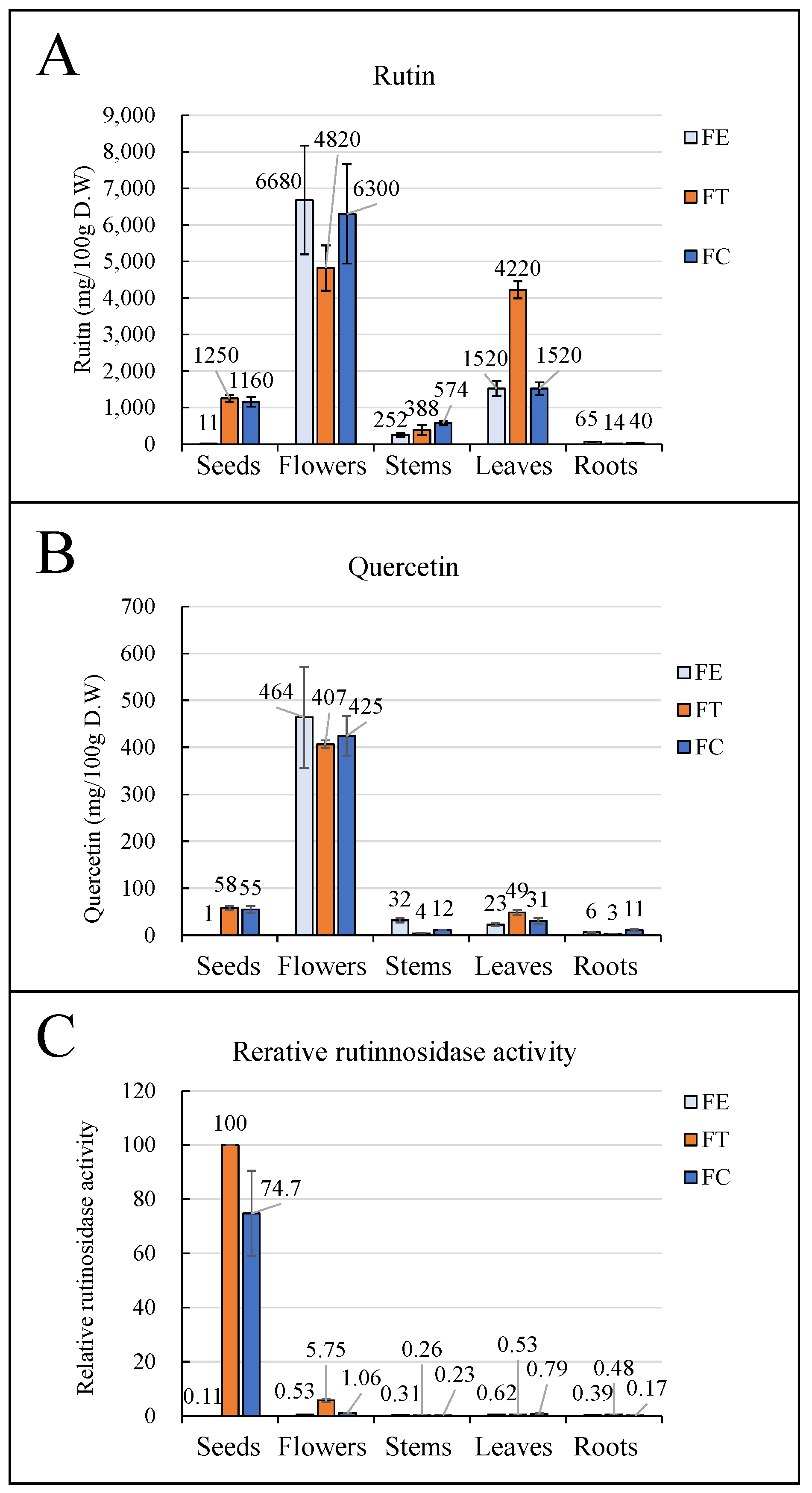
2.2. Organ Distribution of Rutin, Quercetin, and Rutinosidase Activity in FC
2.3. Purification and Characterization of Rutinosidase from FC Seeds
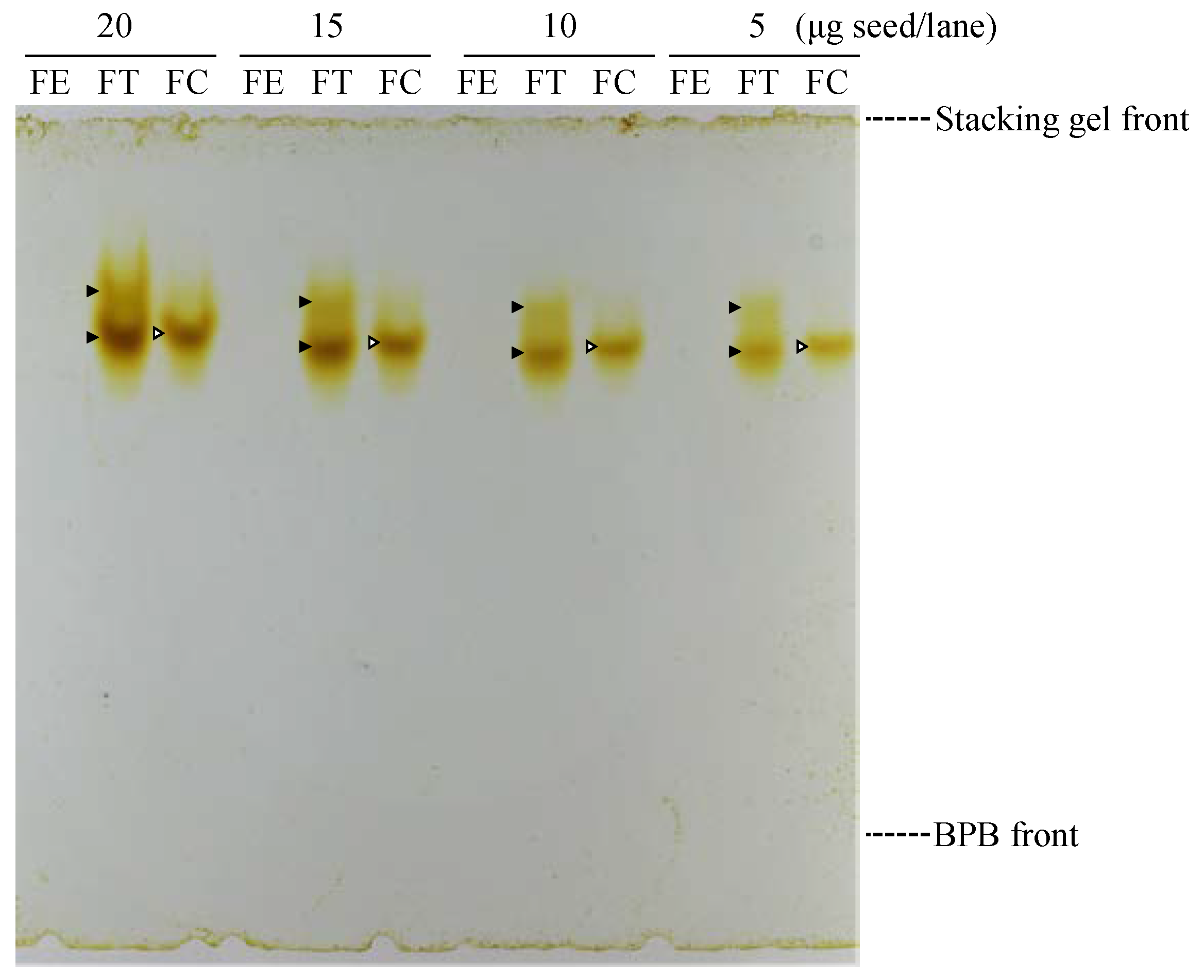
2.4. In-Gel Detection of Rutinosidase in Cultivated Buckwheat Varieties
2.5. Investigation of Rutin and Quercetin Concentration in FC Dough and Evaluation of Bitterness
3. Results
3.1. Organ Distribution of Rutin and Rutinosidase Activity
3.2. Purification and Characterization of Rutinosidase from FC Seeds
3.3. Investigation of Rutin and Quercetin Concentration in FC Dough and Evaluation of Bitterness
4. Discussion
4.1. Organ Distribution of Rutin and Rutinosidase Activity
4.2. Purification and Characterization of Rutinosidase from FC Seeds and Contribution of Rutinosidase on Rutin Hydrolysis and Bitterness in FC Dough
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Griffith, J.Q.; Couch, J.F.; Lindauer, A. Effect of rutin on increased capillary fragility in man. Proc. Soc. Exp. Biol. Med. 1944, 55, 228–229. [Google Scholar] [CrossRef]
- Awatsuhara, R.; Harada, K.; Maeda, T. Antioxidative activity of the buckwheat polyphenol rutin in combination with ovalbumin. Mol. Med. Rep. 2010, 3, 121–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitabayashi, H.; Ujihara, A.; Hirose, T.; Minami, M. Varietal differences and heritability for rutin content in common buckwheat, Fagopyrum esculentum Moench. Breed. Sci. 1995, 45, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Kreft, I.; Fabjan, N.; Yasumoto, K. Rutin content in buckwheat (Fagopyrum esculentum Moench) food materials and products. Food Chem. 2006, 98, 508–512. [Google Scholar] [CrossRef]
- Couch, J.; Naghski, J.; Krewson, C.F. Buckwheat as a source of rutin. Science 1946, 103, 197–198. [Google Scholar] [CrossRef]
- Leisner, C.P. Climate change impacts on food security- focus on perennial cropping systems and nutritional value. Plant Sci. 2020, 293, 110412. [Google Scholar] [CrossRef]
- Lv, X.; Wan, K.; Liang, J.; Feng, Y.; Dao, J.; Wu, S.; Zhang, L.; Yang, X.; Lian, X.; Huang, L.; et al. Sustained productivity and agronomic potential of perennial rice. Nat. Sustain. 2023, 6, 28–38. [Google Scholar] [CrossRef]
- Cox, T.S.; Bender, M.; Picone, C.; Tassel, D.L.V.; Holland, J.B.; Brummer, E.C.; Zoeller, B.E.; Jacson, W. Breeding Perennial Grain Crops. Crit. Rev. Plant Sci. 2002, 21, 59–91. [Google Scholar] [CrossRef]
- Cowart, B.J. Development of taste perceptions in humans: Sensitivity and preference throughout the life span. Psychol. Bull. 1981, 90, 43–73. [Google Scholar] [CrossRef]
- Yasuda, T.; Masaki, K.; Kashiwagi, T. An enzyme degrading rutin in Tartary buckwheat seeds. J. Jpn. Soc. Food Sci. Technol. 1992, 39, 994–1000. [Google Scholar] [CrossRef]
- Suzuki, T.; Honda, Y.; Funatsuki, W.; Nakatsuka, K. Purification and characterization of flavonol 3-glucosidase, and its activity during ripening in Tartary buckwheat seeds. Plant Sci. 2002, 163, 417–423. [Google Scholar] [CrossRef]
- Suzuki, T.; Morishita, T.; Mukasa, Y.; Takigawa, S.; Yokota, S.; Ishiguro, K.; Noda, T. Discovery and genetic analysis of non-bitter Tartary buckwheat (Fagopyrum tataricum Gaertn.) with trace-rutinosidase activity. Breed. Sci. 2014, 64, 339–343. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Morishita, T.; Mukasa, Y.; Takigawa, S.; Yokota, S.; Ishiguro, K.; Noda, T. Breeding of “Manten-Kirari”, a non-bitter and trace-rutinosidase variety of Tartary buckwheat (Fagopyrum tataricum Gaertn.). Breed. Sci. 2014, 64, 344–350. [Google Scholar] [CrossRef] [Green Version]
- Katsu, K.; Suzuki, T.; Fujino, K.; Morishita, T.; Noda, T. Development of a DNA marker for variety discrimination specific to‘Manten-Kirari’ based on an NGS-RNA sequence in Tartary buckwheat (Fagopyrum Tataricum). Food Chem. 2019, 295, 51–57. [Google Scholar] [CrossRef]
- Suzuki, T.; Morishita, T.; Takigawa, S.; Noda, T.; Ishiguro, K.; Otsuka, S. Development of Novel Detection Method for Rutinosidase in Tartary Buckwheat (Fagopyrum tataricum Gaertn.). Plants 2022, 11, 320. [Google Scholar] [CrossRef]
- Ikuma, T.; Kakeuchi, K.; Sagisaka, K.; Takasawa, T. Coomassie brilliant blue G250 dye-binding microassay for protein. Obihirochikusandaigaku Gakujutsukenkyuuhoukoku. 2002, 23, 18–26. [Google Scholar]
- Suzuki, T.; Honda, Y.; Funatsuki, E.; Nakatsuka, K. In-gel detection and study of the role of flavonol 3-glucosidase in the bitter taste generation in tartary buckwheat. J. Sci. Food Agric. 2004, 84, 1691–1694. [Google Scholar] [CrossRef]
- Ishiguro, K.; Morishita, T.; Ashizawa, J.; Suzuki, T.; Noda, T. Antioxidative activities in rutin rich noodles and cookies made with a trace rutinosidase variety of Tartary buckwheat (Fagopyrum tataricum Gaertn.), ‘Manten-Kirari’. Food Sci. Technol. Res. 2016, 22, 557–562. [Google Scholar] [CrossRef] [Green Version]
- Narikawa, T.; Shinoyama, H.; Fujii, T. A β-Rutinosidase from Penicillium rugulosum IFO 7242 That Is a Peculiar Flavonoid Glycosidase. Biosci. Biotechnol. Biochem. 2000, 64, 1317–1319. [Google Scholar] [CrossRef]
- Suzuki, T.; Watanabe, M.; Iki, M.; Aoyagi, Y.; Kim, S.J.; Mukasa, Y.; Yokota, S.; Takigawa, S.; Hashimoto, N.; Noda, T.; et al. Time-course study and effects of drying method on concentrations of γ-aminobutyric acid, flavonoids, antho-cyanin, and 200-Hydroxynicotianamine in leaves of Buckwheats. J. Agric. Food Chem. 2007, 57, 259–264. [Google Scholar] [CrossRef]
- Liu, N.; Zeller, F.J.; Chen, Q.F. The flavonoid content in leaves and inflorescences of the wild perennial Fagopyrum cymosum complex. Genet. Resour. Crop. Evol. 2013, 60, 825–838. [Google Scholar] [CrossRef]
- Johnson, A.E. Photosensitizing toxins from plants and their biologic effects. In Handbook of Natural Toxins; Keeler, R.F., Tu, A.T., Eds.; Plant and Fungal Toxins; Marcel Dekker: New York, NY, USA, 1983; Volume 1, pp. 345–359. [Google Scholar]
- Kitabayashi, H.; Ujihara, A.; Hirose, T.; Minami, M. On the genotypic differences for rutin content in tartary buckwheat, Fagopyrum tataricum Gaerth. Breed. Sci. 1995, 45, 189–194. [Google Scholar]
- Nishimura, M.; Ohkawara, T.; Sato, Y.; Satoh, H.; Suzuki, T.; Ishiguro, K.; Noda, T.; Morishita, T.; Nishihira, J. Effectiveness of rutin-rich Tartary buckwheat (Fagopyrum tataricum Gaertn.) ‘Manten-Kirari’ in body weight reduction related to its antioxidant properties: A randomised, double-blind, placebo-controlled study. J. Funct. Foods 2016, 26, 460–469. [Google Scholar] [CrossRef] [Green Version]
- Fujita, K.; Yoshihashi, T. Heat-treatment of Tartary Buckwheat (Fagopyrum tataricum Gaertn.) Provides Dehulled and Gelatinized Product with Denatured Rutinosidase. Food Sci. Technol. Res. 2019, 25, 613–618. [Google Scholar] [CrossRef]
- Morishita, T.; Ishiguro, K.; Noda, T.; Suzuki, T. The effect of grain moisture contents on the roll milling characteristics of Tartary buckwheat cultivar ‘Manten-Kirari’. Food Sci. Technol. Res. 2020, 23, 536–546. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Morishita, T.; Takigawa, S.; Noda, T.; Ishiguro, K.; Otsuka, S. Effect of Sodium Bicarbonate on Rutin Residual Ratio in Tartary Buckwheat (Fagopyrum tataricum Gaertn.) Dough. Food Sci. Technol. Res. 2020, 26, 597–603. [Google Scholar] [CrossRef]
- Krotov, A.S.; Dranenko, E.T. Amphidiploid of Buckwheat F. giganteum Krotov sp. Nova. Biull Vses Ord Lenina Inst Rastenievod Im NI Vavilova 1973, 41–44. (In Russian). From Plant Breed. Abst. 1974, 44, 1722 [Google Scholar]
- Chen, Q.F.; Hsam, S.L.K.; Zeller, F.A. Study of cytology, isozyme and interspecific hybridization on the big-achene group of buckwheat species (Fagopyrum, Polygonaceae). Crop. Sci. 2004, 44, 1511–1518. [Google Scholar] [CrossRef]
- Noda, T.; Ishiguro, K.; Suzuki, T.; Morishita, T. Roasted Tartary Buckwheat Bran as a Material for Producing Rutin-Rich Tea Beverages. Plants 2021, 10, 2662. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Volume | Total Protein | Total Activity | Specific Activity | Purification | Yield | |
|---|---|---|---|---|---|---|
| Purification step | (mL) | (mg) | (μkat) | (μkat/mg protein) | (fold) | (%) |
| Crude extract | 542 | 196 | 553 | 2.82 | 1.00 | 100 |
| Q-Sepharose Fast Flow | 24.0 | 26.9 | 295 | 11.0 | 3.89 | 53.3 |
| Hiload16/600 Superdex 200pg | 18.0 | 5.18 | 115 | 22.3 | 7.90 | 20.9 |
| Mono Q 5/50 GL (5–400 mM LiCl) | 7.0 | 0.33 | 28.5 | 86.4 | 30.6 | 5.15 |
| Mono Q 5/50 GL (80–120 mM LiCl) | 7.0 | 0.19 | 24.0 | 130 | 46.1 | 4.34 |
| Km | (mM) | 0.367 ± 0.0235 |
| Vmax | (U/mg) | 36.2 ± 1.72 |
| Data are means ± SD (n = 3). | ||
| Minutes after Addition of Water to Flour | ||
|---|---|---|
| 0 | 10 | |
| Rutin | 100 ± 2.40 | 0.160 ± 0.029 |
| Quercetin | 0.014 ± 0.004 | 100 ± 8.03 |
| Data are means ± SD (n = 3) Dough was stored at 25 °C Rutin at 0 min = 100 Quercetin at 10 min = 100 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, T.; Kurokoh, R.; Murakami, S.; Takahashi, N.; Matsuura, A.; Katsu, K.; Murata, K. Rutin Concentration and Characterization of Rutinosidase in Perennial Buckwheat (Fagopyrum cymosum) and Its Application in Foods. Foods 2023, 12, 1417. https://doi.org/10.3390/foods12071417
Suzuki T, Kurokoh R, Murakami S, Takahashi N, Matsuura A, Katsu K, Murata K. Rutin Concentration and Characterization of Rutinosidase in Perennial Buckwheat (Fagopyrum cymosum) and Its Application in Foods. Foods. 2023; 12(7):1417. https://doi.org/10.3390/foods12071417
Chicago/Turabian StyleSuzuki, Tatsuro, Rio Kurokoh, Shogo Murakami, Naohiro Takahashi, Asana Matsuura, Kenjiro Katsu, and Kouhei Murata. 2023. "Rutin Concentration and Characterization of Rutinosidase in Perennial Buckwheat (Fagopyrum cymosum) and Its Application in Foods" Foods 12, no. 7: 1417. https://doi.org/10.3390/foods12071417