

The Potential Therapeutic Role of Lactobacillaceae rhamnosus for Treatment of Inflammatory Bowel Disease

Abstract
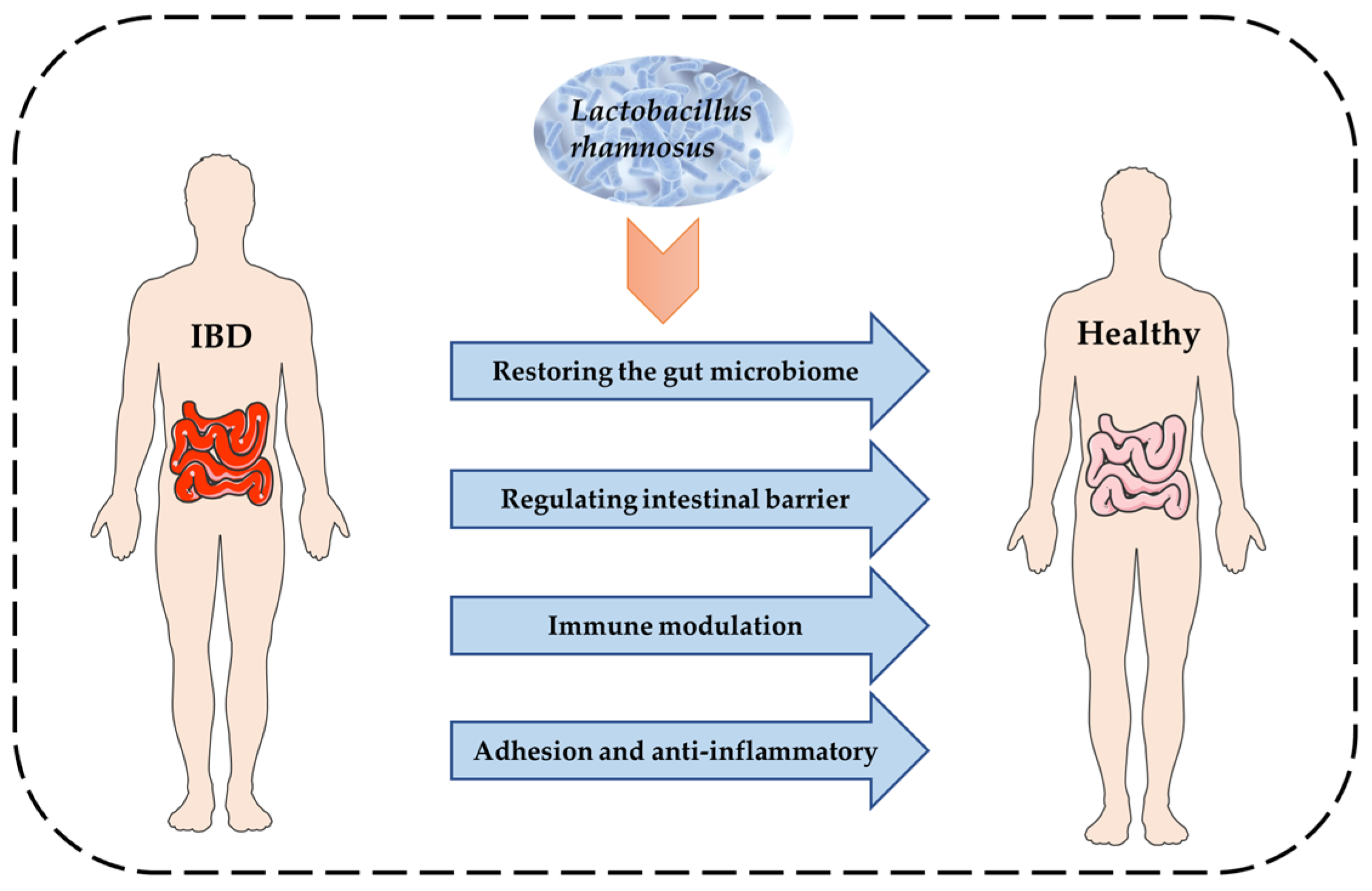
:1. Introduction
2. Molecular Mechanism of L. rhamnosus to Alleviate IBD
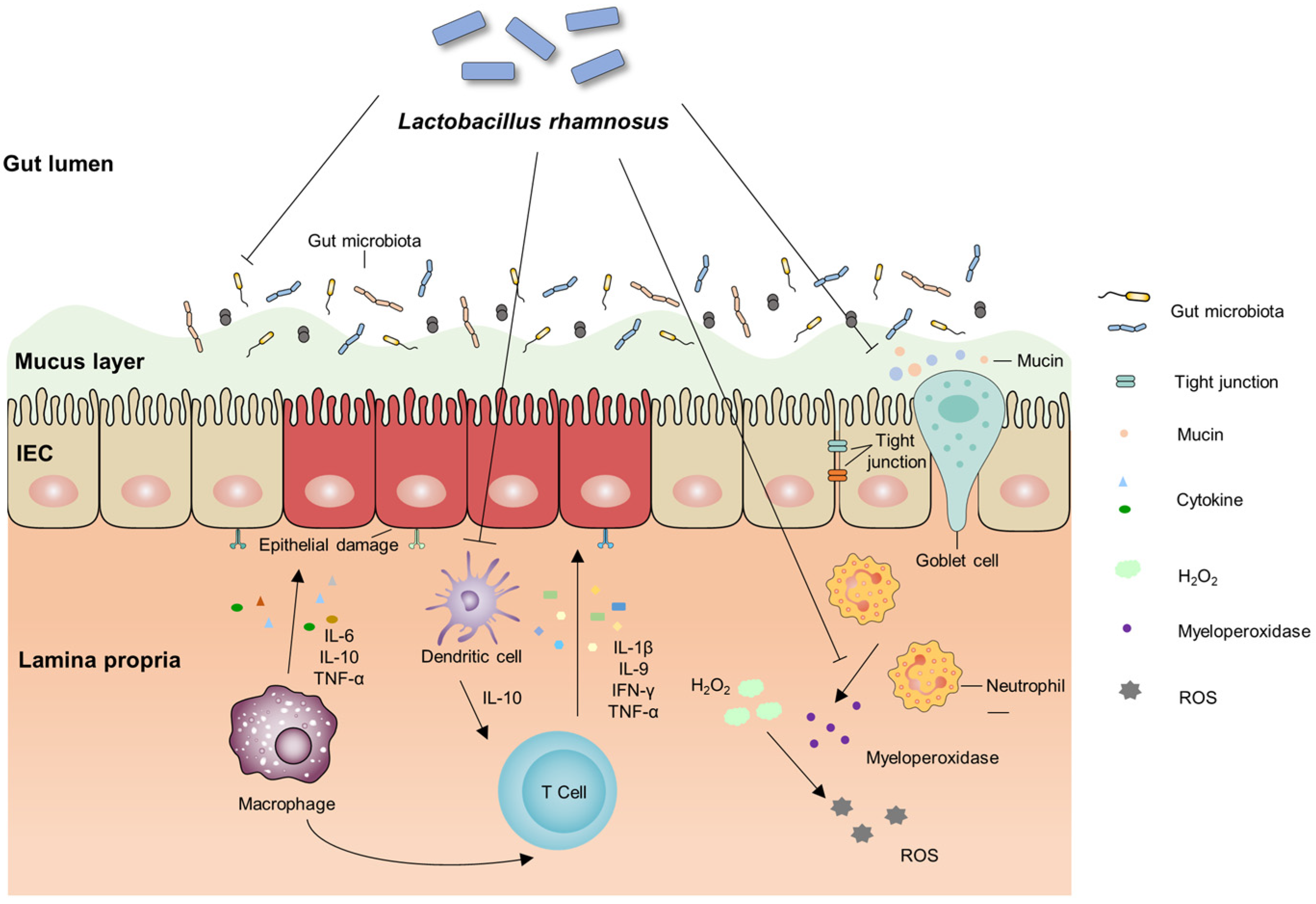
2.1. IBD and Gut Microbiota
2.1.1. Relationship between IBD and Gut Microbiota
2.1.2. Alleviating Symptoms in Animal Models by Regulating Gut Microbiota
2.2. IBD and the Intestinal Barrier
2.2.1. Relationship between IBD and Abnormal Intestinal Barrier Function
2.2.2. Ability to Restore Intestinal Barrier Function in Animal and Cellular Models
2.3. IBD and the Immune Function Modulation
2.3.1. Relationship between IBD and the Abnormal Immune Function
2.3.2. Improving Immune Disorders Caused by Colitis in Animal Models
2.4. IBD and Adhesion Anti-Inflammatory
2.4.1. IBD and Inflammation Caused by Adherent Pathogenic Bacteria
2.4.2. LGG Exerts Adhesion and Anti-Inflammatory Effects in Animals and In Vitro Models
3. Safety Issues in the Application of L rhamnosus
4. Conclusions and Outlook
Author Contributions
Funding
Conflicts of Interest
References
- Flynn, S.; Eisenstein, S. Inflammatory bowel disease presentation and diagnosis. Surg. Clin. 2019, 99, 1051–1062. [Google Scholar] [CrossRef] [PubMed]
- Kotze, P.G.; Underwood, F.E.; Damião, A.O.M.C.; Ferraz, J.G.P.; Saad-Hossne, R.; Toro, M.; Iade, B.; Bosques-Padilla, F.; Teixeira, F.V.; Juliao-Banos, F. Progression of inflammatory bowel diseases throughout Latin America and the Caribbean: A systematic review. Clin. Gastroenterol. Hepatol. 2020, 18, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Celiberto, L.S.; Bedani, R.; Rossi, E.A.; Cavallini, D.C.U. Probiotics: The scientific evidence in the context of inflammatory bowel disease. Crit. Rev. Food Sci. Nutr. 2017, 57, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Chang, E.B. Inflammatory Bowel Diseases (IBD) and the microbiome—Searching the crime scene for clues. Gastroenterology 2021, 160, 524–537. [Google Scholar] [CrossRef]
- Mirkov, M.U.; Verstockt, B.; Cleynen, I. Genetics of inflammatory bowel disease: Beyond NOD2. Lancet Gastroenterol. Hepatol. 2017, 2, 224–234. [Google Scholar] [CrossRef]
- Levine, A.; Sigall Boneh, R.; Wine, E. Evolving role of diet in the pathogenesis and treatment of inflammatory bowel diseases. Gut 2018, 67, 1726–1738. [Google Scholar] [CrossRef]
- Yan, F.; Cao, H.; Cover, T.L.; Whitehead, R.; Washington, M.K.; Polk, D.B. Soluble proteins produced by probiotic bacteria regulate intestinal epithelial cell survival and growth. Gastroenterology 2007, 132, 562–575. [Google Scholar] [CrossRef]
- Neurath, M.F. Current and emerging therapeutic targets for IBD. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 269–278. [Google Scholar] [CrossRef]
- Sandborn, W.J.; Feagan, B.G.; Marano, C.; Zhang, H.; Strauss, R.; Johanns, J.; Adedokun, O.J.; Guzzo, C.; Colombel, J.-F.; Reinisch, W. Subcutaneous golimumab induces clinical response and remission in patients with moderate-to-severe ulcerative colitis. Gastroenterology 2014, 146, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, L.R.; MacDonald, J.K. Oral 5-aminosalicylic acid for maintenance of remission in ulcerative colitis. Cochrane Database Syst. Rev. 2006, 2, 1–32. [Google Scholar]
- Ganji-Arjenaki, M.; Rafieian-Kopaei, M. Probiotics are a good choice in remission of inflammatory bowel diseases: A meta analysis and systematic review. J. Cell. Physiol. 2018, 233, 2091–2103. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, H.; Nakase, H.; Inoue, S.; Kawanami, C.; Itani, T.; Ohana, M. Efficacy of probiotic treatment with Bifidobacterium longum 536 for induction of remission in active ulcerative colitis: A randomized, double-blinded, placebo-controlled multicenter trial. Dig. Endosc. 2016, 28, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Fedorak, R.N.; Feagan, B.G.; Hotte, N.; Leddin, D.; Dieleman, L.A.; Petrunia, D.M.; Madsen, K. The probiotic VSL# 3 has anti-inflammatory effects and could reduce endoscopic recurrence after surgery for Crohn’s disease. Clin. Gastroenterol. Hepatol. 2015, 13, 928–935. [Google Scholar]
- Petersen, A.M.; Mirsepasi, H.; Halkjær, S.I.; Mortensen, E.M.; Nordgaard-Lassen, I.; Krogfelt, K.A. Ciprofloxacin and probiotic Escherichia coli Nissle add-on treatment in active ulcerative colitis: A double-blind randomized placebo controlled clinical trial. J. Crohn’s Colitis 2014, 8, 1498–1505. [Google Scholar] [CrossRef]
- Haller, D.; Antoine, J.-M.; Bengmark, S.; Enck, P.; Rijkers, G.T.; Lenoir-Wijnkoop, I. Guidance for substantiating the evidence for beneficial effects of probiotics: Probiotics in chronic inflammatory bowel disease and the functional disorder irritable bowel syndrome. J. Nutr. 2010, 140, 690S–697S. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Liu, B.; Huang, L.; Li, Z.; Cheng, Y.; Tian, Y.; Xie, L. Probiotic consortia and their metabolites ameliorate the symptoms of inflammatory bowel diseases in a colitis mouse model. Microbiol. Spectr. 2022, 10, e00657-22. [Google Scholar] [CrossRef] [PubMed]
- Engevik, M.A.; Herrmann, B.; Ruan, W.; Engevik, A.C.; Engevik, K.A.; Ihekweazu, F.; Versalovic, J. Bifidobacterium dentium-derived y-glutamylcysteine suppresses ER-mediated goblet cell stress and reduces TNBS-driven colonic inflammation. Gut Microbes 2021, 13, 1902717. [Google Scholar] [CrossRef]
- Liu, Z.; Jiang, Z.; Zhang, Z.; Liu, T.; Fan, Y.; Liu, T.; Peng, N. Bacillus coagulans in combination with chitooligosaccharides regulates gut microbiota and ameliorates the Dss-induced colitis in mice. Microbiol. Spectr. 2022, 10, e00641-22. [Google Scholar] [CrossRef] [PubMed]
- Pascual, L.M.; Daniele, M.B.; Ruiz, F.; Giordano, W.; Pájaro, C.; Barberis, L. Lactobacillus rhamnosus L60, a potential probiotic isolated from the human vagina. J. Gen. Appl. Microbiol. 2008, 54, 141–148. [Google Scholar] [CrossRef]
- Huang, D.; Yang, B.; Chen, Y.; Stanton, C.; Ross, R.P.; Zhao, J.; Zhang, H.; Chen, W. Comparative genomic analyses of Lactobacillus rhamnosus isolated from Chinese subjects. Food Biosci. 2020, 36, 100659. [Google Scholar] [CrossRef]
- Bernardeau, M.; Vernoux, J.P.; Henri-Dubernet, S.; Guéguen, M. Safety assessment of dairy microorganisms: The Lactobacillus genus. Int. J. Food Microbiol. 2008, 126, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Kolinska, J.; Zakostelecka, M.; Zemanova, Z.; Lisa, V.; Golias, J.; Kozakova, H.; Dvorak, B. Cellular differentiation of non-transformed intestinal epithelial cells is regulated by Lactobacillus rhamnosus and L. casei strains. Physiol. Res. 2018, 67, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Rolim, F.R.L.; dos Santos, K.M.O.; de Barcelos, S.C.; do Egito, A.S.; Ribeiro, T.S.; da Conceicao, M.L.; Magnani, M.; de Oliveira, M.E.G.; do Egypto, R.d.C.R. Survival of Lactobacillus rhamnosus EM1107 in simulated gastrointestinal conditions and its inhibitory effect against pathogenic bacteria in semi-hard goat cheese. LWT Food Sci. Technol. 2015, 63, 807–813. [Google Scholar] [CrossRef]
- Yang, L.; Higginbotham, J.N.; Liu, L.; Zhao, G.; Acra, S.A.; Peek, R.M., Jr.; Polk, D.B.; Li, H.; Yan, F. Production of a Functional Factor, p40, by Lactobacillus rhamnosus GG Is Promoted by Intestinal Epithelial Cell-Secreted Extracellular Vesicles. Infect. Immun. 2019, 87, e00113-19. [Google Scholar] [CrossRef]
- Claes, I.J.; Lebeer, S.; Shen, C.; Verhoeven, T.L.; Dilissen, E.; De Hertogh, G.; Bullens, D.M.; Ceuppens, J.L.; Van Assche, G.; Vermeire, S.; et al. Impact of lipoteichoic acid modification on the performance of the probiotic Lactobacillus rhamnosus GG in experimental colitis. Clin. Exp. Immunol. 2010, 162, 306–314. [Google Scholar] [CrossRef]
- Wan, M.L.; Chen, Z.; Shah, N.P.; El-Nezami, H. Effects of Lactobacillus rhamnosus GG and Escherichia coli Nissle 1917 Cell-Free Supernatants on Modulation of Mucin and Cytokine Secretion on Human Intestinal Epithelial HT29-MTX Cells. J. Food Sci. 2018, 83, 1999–2007. [Google Scholar] [CrossRef]
- Gamallat, Y.; Meyiah, A.; Kuugbee, E.D.; Hago, A.M.; Chiwala, G.; Awadasseid, A.; Bamba, D.; Zhang, X.; Shang, X.; Luo, F.; et al. Lactobacillus rhamnosus induced epithelial cell apoptosis, ameliorates inflammation and prevents colon cancer development in an animal model. Biomed. Pharmacother. 2016, 83, 536–541. [Google Scholar] [CrossRef]
- Laval, L.; Martin, R.; Natividad, J.N.; Chain, F.; Miquel, S.; de Maredsous, C.D.; Capronnier, S.; Sokol, H.; Verdu, E.F.; van Hylckama Vlieg, J.E.T.; et al. Lactobacillus rhamnosus CNCM I-3690 and the commensal bacterium Faecalibacterium prausnitzii A2-165 exhibit similar protective effects to induced barrier hyper-permeability in mice. Gut Microbes 2015, 6, 1–9. [Google Scholar] [CrossRef]
- Foster, L.M.; Tompkins, T.A.; Dahl, W.J. A comprehensive post-market review of studies on a probiotic product containing Lactobacillus helveticus R0052 and Lactobacillus rhamnosus R0011. Benef. Microbes 2011, 2, 319–334. [Google Scholar] [CrossRef]
- Stanojević, S.; Blagojević, V.; Ćuruvija, I.; Veljović, K.; Bajić, S.S.; Kotur-Stevuljević, J.; Bogdanović, A.; Petrović, R.; Vujnović, I.; Kovačević-Jovanović, V. Oral treatment with Lactobacillus rhamnosus 64 during the early postnatal period improves the health of adult rats with TNBS-induced colitis. J. Funct. Foods 2018, 48, 92–105. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, F.; Wang, W.; Sun, C.; Gao, D.; Ma, J.; Hussain, M.A.; Xu, C.; Jiang, Z.; Hou, J. Study of the alleviation effects of a combination of Lactobacillus rhamnosus and inulin on mice with colitis. Food Funct. 2020, 11, 3823–3837. [Google Scholar] [CrossRef]
- Rodrigues, R.; Guerra, G.; Soares, J.; Santos, K.; Rolim, F.; Assis, P.; Araújo, D.; de Araújo Júnior, R.F.; Garcia, V.B.; de Araújo, A.A. Lactobacillus rhamnosus EM1107 in goat milk matrix modulates intestinal inflammation involving NF-κB p65 and SOCs-1 in an acid-induced colitis model. J. Funct. Foods 2018, 50, 78–92. [Google Scholar] [CrossRef]
- Yeo, S.; Park, H.; Seo, E.; Kim, J.; Kim, B.K.; Choi, I.S.; Huh, C.S. Anti-inflammatory and gut microbiota modulatory effect of Lactobacillus rhamnosus strain LDTM 7511 in a dextran sulfate sodium-induced colitis murine model. Microorganisms 2020, 8, 845. [Google Scholar] [CrossRef] [PubMed]
- Panpetch, W.; Hiengrach, P.; Nilgate, S.; Tumwasorn, S.; Somboonna, N.; Wilantho, A.; Chatthanathon, P.; Prueksapanich, P.; Leelahavanichkul, A. Additional Candida albicans administration enhances the severity of dextran sulfate solution induced colitis mouse model through leaky gut-enhanced systemic inflammation and gut-dysbiosis but attenuated by Lactobacillus rhamnosus L34. Gut Microbes 2020, 11, 465–480. [Google Scholar] [CrossRef]
- Pang, B.; Jin, H.; Liao, N.; Li, J.; Jiang, C.; Shao, D.; Shi, J. Lactobacillus rhamnosus from human breast milk ameliorates ulcerative colitis in mice via gut microbiota modulation. Food Funct. 2021, 12, 5171–5186. [Google Scholar] [CrossRef]
- Miyauchi, E.; Morita, H.; Tanabe, S. Lactobacillus rhamnosus alleviates intestinal barrier dysfunction in part by increasing expression of zonula occludens-1 and myosin light-chain kinase in vivo. J. Dairy Sci. 2009, 92, 2400–2408. [Google Scholar] [CrossRef]
- Han, M.; Liao, W.; Si, X.; Bai, C.; Gai, Z. Protective Effects of Lacticaseibacillus rhamnosus Hao9 on Dextran Sulphate Sodium-Induced Ulcerative Colitis in Mice. J. Appl. Microbiol. 2022, 133, 2039–2049. [Google Scholar] [CrossRef]
- Martin, R.; Chamignon, C.; Mhedbi-Hajri, N.; Chain, F.; Derrien, M.; Escribano-Vazquez, U.; Garault, P.; Cotillard, A.; Pham, H.P.; Chervaux, C.; et al. The potential probiotic Lactobacillus rhamnosus CNCM I-3690 strain protects the intestinal barrier by stimulating both mucus production and cytoprotective response. Sci. Rep. 2019, 9, 5398. [Google Scholar] [CrossRef] [PubMed]
- Zocco, M.; Dal Verme, L.Z.; Cremonini, F.; Piscaglia, A.; Nista, E.; Candelli, M.; Novi, M.; Rigante, D.; Cazzato, I.; Ojetti, V. Efficacy of Lactobacillus GG in maintaining remission of ulcerative colitis. Aliment. Pharmacol. Ther. 2006, 23, 1567–1574. [Google Scholar] [CrossRef]
- Gupta, P.; Andrew, H.; Kirschner, B.S.; Guandalini, S. Is Lactobacillus GG helpful in children with Crohn’s disease? Results of a preliminary, open-label study. J. Pediatr. Gastroenterol. Nutr. 2000, 31, 453–457. [Google Scholar] [CrossRef]
- Schultz, M.; Timmer, A.; Herfarth, H.H.; Sartor, R.B.; Vanderhoof, J.A.; Rath, H.C. Lactobacillus GG in inducing and maintaining remission of Crohn’s disease. BMC Gastroenterol. 2004, 4, 5. [Google Scholar] [CrossRef] [PubMed]
- Malin, M.; Suomalainen, H.; Saxelin, M.; Isolauri, E. Promotion of IgA immune response in patients with Crohn’s disease by oral bacteriotherapy with Lactobacillus GG. Ann. Nutr. Metab. 1996, 40, 137–145. [Google Scholar] [CrossRef]
- Prantera, C.; Scribano, M.; Falasco, G.; Andreoli, A.; Luzi, C. Ineffectiveness of probiotics in preventing recurrence after curative resection for Crohn’s disease: A randomised controlled trial with Lactobacillus GG. Gut 2002, 51, 405–409. [Google Scholar] [CrossRef]
- Gosselink, M.P.; Schouten, W.R.; van Lieshout, L.; Hop, W.C.; Laman, J.D.; Ruseler-van Embden, J.G. Delay of the first onset of pouchitis by oral intake of the probiotic strain Lactobacillus rhamnosus GG. Dis. Colon Rectum 2004, 47, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Rutgeerts, P.; Peeters, M.; Hiele, M.; Vantrappen, G.; Pennincx, F.; Aerts, R.; Kerremans, R.; Goboes, K. Effect of faecal stream diversion on recurrence of Crohn’s disease in the neoterminal ileum. Lancet 1991, 338, 771–774. [Google Scholar] [CrossRef] [PubMed]
- Casellas, F.; Borruel, N.; Papo, M.; Guarner, F.; Antolín, M.; Videla, S.; Malagelada, J.-R. Antiinflammatory Effects of Enterically Coated Amoxicillin-Clavulanic Acid in Active Ulcerative Colitis. Inflamm. Bowel Dis. 1998, 4, 1–5. [Google Scholar] [CrossRef]
- Sartor, R.B. Therapeutic manipulation of the enteric microflora in inflammatory bowel diseases: Antibiotics, probiotics, and prebiotics. Gastroenterology 2004, 126, 1620–1633. [Google Scholar] [CrossRef]
- Lucas López, R.; Grande Burgos, M.J.; Gálvez, A.; Pérez Pulido, R. The human gastrointestinal tract and oral microbiota in inflammatory bowel disease: A state of the science review. APMIS 2017, 125, 3–10. [Google Scholar] [CrossRef]
- Jostins, L.; Ripke, S.; Weersma, R.K.; Duerr, R.H.; McGovern, D.P.; Hui, K.Y.; Lee, J.C.; Philip Schumm, L.; Sharma, Y.; Anderson, C.A. Host–microbe interactions have shaped the genetic architecture of inflammatory bowel disease. Nature 2012, 491, 119–124. [Google Scholar] [CrossRef]
- Zeng, M.; Inohara, N.; Nuñez, G. Mechanisms of inflammation-driven bacterial dysbiosis in the gut. Mucosal Immunol. 2017, 10, 18–26. [Google Scholar] [CrossRef]
- Hirano, A.; Umeno, J.; Okamoto, Y.; Shibata, H.; Ogura, Y.; Moriyama, T.; Torisu, T.; Fujioka, S.; Fuyuno, Y.; Kawarabayasi, Y. Comparison of the microbial community structure between inflamed and non-inflamed sites in patients with ulcerative colitis. J. Gastroenterol. Hepatol. 2018, 33, 1590–1597. [Google Scholar] [CrossRef]
- Nishino, K.; Nishida, A.; Inoue, R.; Kawada, Y.; Ohno, M.; Sakai, S.; Inatomi, O.; Bamba, S.; Sugimoto, M.; Kawahara, M. Analysis of endoscopic brush samples identified mucosa-associated dysbiosis in inflammatory bowel disease. J. Gastroenterol. 2018, 53, 95–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scaldaferri, F.; Gerardi, V.; Lopetuso, L.R.; Del Zompo, F.; Mangiola, F.; Boškoski, I.; Bruno, G.; Petito, V.; Laterza, L.; Cammarota, G. Gut microbial flora, prebiotics, and probiotics in IBD: Their current usage and utility. BioMed Res. Int. 2013, 2013, 435268. [Google Scholar] [CrossRef] [PubMed]
- Hager, C.L.; Ghannoum, M.A. The mycobiome: Role in health and disease, and as a potential probiotic target in gastrointestinal disease. Dig. Liver Dis. 2017, 49, 1171–1176. [Google Scholar] [CrossRef]
- Wan, C.; Qian, W.W.; Liu, W.; Pi, X.; Tang, M.T.; Wang, X.L.; Gu, Q.; Li, P.; Zhou, T. Exopolysaccharide from Lactobacillus rhamnosus ZFM231 alleviates DSS-induced colitis in mice by regulating gut microbiota. J. Sci. Food Agric. 2022, 102, 7087–7097. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Laukoetter, M.G.; Nava, P.; Nusrat, A. Role of the intestinal barrier in inflammatory bowel disease. World J. Gastroenterol. 2008, 14, 401. [Google Scholar] [CrossRef]
- Clayburgh, D.R.; Shen, L.; Turner, J.R. A porous defense: The leaky epithelial barrier in intestinal disease. Lab. Investig. 2004, 84, 282–291. [Google Scholar] [CrossRef]
- Turpin, W.; Lee, S.-H.; Garay, J.A.R.; Madsen, K.L.; Meddings, J.B.; Bedrani, L.; Power, N.; Espin-Garcia, O.; Xu, W.; Smith, M.I. Increased intestinal permeability is associated with later development of Crohn’s disease. Gastroenterology 2020, 159, 2092–2100.e5. [Google Scholar] [CrossRef]
- Torres, J.; Petralia, F.; Sato, T.; Wang, P.; Telesco, S.E.; Strauss, R.; Li, X.-j.; Laird, R.M.; Gutierrez, R.L.; Porter, C.K. Serum biomarkers identify patients who will develop inflammatory bowel diseases up to 5 years before diagnosis. Gastroenterology 2020, 159, 96–104. [Google Scholar] [CrossRef]
- Lechuga, S.; Ivanov, A.I. Disruption of the epithelial barrier during intestinal inflammation: Quest for new molecules and mechanisms. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2017, 1864, 1183–1194. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Li, Z.-R.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate enhances the intestinal barrier by facilitating tight junction assembly via activation of AMP-activated protein kinase in Caco-2 cell monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Candela, M.; Perna, F.; Carnevali, P.; Vitali, B.; Ciati, R.; Gionchetti, P.; Rizzello, F.; Campieri, M.; Brigidi, P. Interaction of probiotic Lactobacillus and Bifidobacterium strains with human intestinal epithelial cells: Adhesion properties, competition against enteropathogens and modulation of IL-8 production. Int. J. Food Microbiol. 2008, 125, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Krishna Rao, R.; Samak, G. Protection and restitution of gut barrier by probiotics: Nutritional and clinical implications. Current Nutr. Food Sci. 2013, 9, 99–107. [Google Scholar] [CrossRef]
- Seth, A.; Yan, F.; Polk, D.B.; Rao, R. Probiotics ameliorate the hydrogen peroxide-induced epithelial barrier disruption by a PKC-and MAP kinase-dependent mechanism. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1060–G1069. [Google Scholar] [CrossRef]
- Uhlig, H.H.; Powrie, F. Translating immunology into therapeutic concepts for inflammatory bowel disease. Annu. Rev. Immunol. 2018, 36, 755–781. [Google Scholar] [CrossRef]
- Hütter, J.; Eriksson, M.; Johannssen, T.; Klopfleisch, R.; von Smolinski, D.; Gruber, A.D.; Seeberger, P.H.; Lepenies, B. Role of the C-type lectin receptors MCL and DCIR in experimental colitis. PLoS ONE 2014, 9, e103281. [Google Scholar] [CrossRef]
- Glocker, E.-O.; Frede, N.; Perro, M.; Sebire, N.; Elawad, M.; Shah, N.; Grimbacher, B. Infant colitis—It’s in the genes. Lancet 2010, 376, 1272. [Google Scholar] [CrossRef]
- Bron, P.A.; Van Baarlen, P.; Kleerebezem, M. Emerging molecular insights into the interaction between probiotics and the host intestinal mucosa. Nat. Rev. Microbiol. 2012, 10, 66–78. [Google Scholar] [CrossRef]
- Sutmuller, R.P.; den Brok, M.H.; Kramer, M.; Bennink, E.J.; Toonen, L.W.; Kullberg, B.-J.; Joosten, L.A.; Akira, S.; Netea, M.G.; Adema, G.J. Toll-like receptor 2 controls expansion and function of regulatory T cells. J. Clin. Investig. 2006, 116, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Littman, D.R.; Rudensky, A.Y. Th17 and regulatory T cells in mediating and restraining inflammation. Cell 2010, 140, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Schnell, A.; Huang, L.; Singer, M.; Singaraju, A.; Barilla, R.M.; Regan, B.M.L.; Bollhagen, A.; Thakore, P.I.; Dionne, D.; Delorey, T.M.; et al. Stem-like intestinal Th17 cells give rise to pathogenic effector T cells during autoimmunity. Cell 2021, 184, 6281–6298. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Lopes, J.E.; Chong, M.M.; Ivanov, I.I.; Min, R.; Victora, G.D.; Shen, Y.; Du, J.; Rubtsov, Y.P.; Rudensky, A.Y. TGF-β-induced Foxp3 inhibits TH17 cell differentiation by antagonizing RORγt function. Nature 2008, 453, 236–240. [Google Scholar] [CrossRef]
- Jia, L.; Wu, R.; Han, N.; Fu, J.; Luo, Z.; Guo, L.; Su, Y.; Du, J.; Liu, Y. Porphyromonas gingivalis and Lactobacillus rhamnosus GG regulate the Th17/Treg balance in colitis via TLR4 and TLR2. Clin. Transl. Immunol. 2020, 9, e1213. [Google Scholar] [CrossRef] [PubMed]
- Han, S.-K.; Shin, Y.-J.; Lee, D.-Y.; Kim, K.M.; Yang, S.-J.; Kim, D.S.; Choi, J.-W.; Lee, S.; Kim, D.-H. Lactobacillus rhamnosus HDB1258 modulates gut microbiota-mediated immune response in mice with or without lipopolysaccharide-induced systemic inflammation. BMC Microbiol. 2021, 21, 146. [Google Scholar] [CrossRef]
- Vong, L.; Lorentz, R.J.; Assa, A.; Glogauer, M.; Sherman, P.M. Probiotic Lactobacillus rhamnosus inhibits the formation of neutrophil extracellular traps. J. Immunol. 2014, 192, 1870–1877. [Google Scholar] [CrossRef]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef]
- Lin, P.W.; Myers, L.E.; Ray, L.; Song, S.C.; Nasr, T.R.; Berardinelli, A.J.; Kundu, K.; Murthy, N.; Hansen, J.M.; Neish, A.S. Lactobacillus rhamnosus blocks inflammatory signaling in vivo via reactive oxygen species generation. Free Radic. Biol. Med. 2009, 47, 1205–1211. [Google Scholar] [CrossRef]
- Ganguli, K.; Collado, M.C.; Rautava, J.; Lu, L.; Satokari, R.; von Ossowski, I.; Reunanen, J.; de Vos, W.M.; Palva, A.; Isolauri, E.; et al. Lactobacillus rhamnosus GG and its SpaC pilus adhesin modulate inflammatory responsiveness and TLR-related gene expression in the fetal human gut. Pediatr. Res. 2015, 77, 528–535. [Google Scholar] [CrossRef]
- Ardita, C.S.; Mercante, J.W.; Kwon, Y.M.; Luo, L.; Crawford, M.E.; Powell, D.N.; Jones, R.M.; Neish, A.S. Epithelial adhesion mediated by pilin SpaC is required for Lactobacillus rhamnosus GG-induced cellular responses. Appl. Environ. Microbiol. 2014, 80, 5068–5077. [Google Scholar] [CrossRef]
- Smythies, L.E.; Sellers, M.; Clements, R.H.; Mosteller-Barnum, M.; Meng, G.; Benjamin, W.H.; Orenstein, J.M.; Smith, P.D. Human intestinal macrophages display profound inflammatory anergy despite avid phagocytic and bacteriocidal activity. J. Clin. Investig. 2005, 115, 66–75. [Google Scholar] [CrossRef]
- Darfeuille-Michaud, A.; Neut, C.; Barnich, N.; Lederman, E.; Di Martino, P.; Desreumaux, P.; Gambiez, L.; Joly, B.; Cortot, A.; Colombel, J.-F. Presence of adherent Escherichia coli strains in ileal mucosa of patients with Crohn’s disease. Gastroenterology 1998, 115, 1405–1413. [Google Scholar] [CrossRef]
- O’Brien, C.L.; Bringer, M.-A.; Holt, K.E.; Gordon, D.M.; Dubois, A.L.; Barnich, N.; Darfeuille-Michaud, A.; Pavli, P. Comparative genomics of Crohn’s disease-associated adherent-invasive Escherichia coli. Gut 2017, 66, 1382–1389. [Google Scholar] [CrossRef] [PubMed]
- Bétis, F.; Brest, P.; Hofman, V.; Guignot, J.; Bernet-Camard, M.-F.; Rossi, B.; Servin, A.; Hofman, P. The Afa/Dr adhesins of diffusely adhering Escherichia coli stimulate interleukin-8 secretion, activate mitogen-activated protein kinases, and promote polymorphonuclear transepithelial migration in T84 polarized epithelial cells. Infect. Immun. 2003, 71, 1068–1074. [Google Scholar] [CrossRef] [PubMed]
- Harrington, S.M.; Strauman, M.C.; Abe, C.M.; Nataro, J.P. Aggregative adherence fimbriae contribute to the inflammatory response of epithelial cells infected with enteroaggregative Escherichia coli. Cell. Microbiol. 2005, 7, 1565–1578. [Google Scholar] [CrossRef] [PubMed]
- Steiner, T.S.; Nataro, J.P.; Poteet-Smith, C.E.; Smith, J.A.; Guerrant, R.L. Enteroaggregative Escherichia coli expresses a novel flagellin that causes IL-8 release from intestinal epithelial cells. J. Clin. Investig. 2000, 105, 1769–1777. [Google Scholar] [CrossRef]
- Sharma, R.; Tesfay, S.; Tomson, F.L.; Kanteti, R.P.; Viswanathan, V.; Hecht, G. Balance of bacterial pro-and anti-inflammatory mediators dictates net effect of enteropathogenic Escherichia coli on intestinal epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G685–G694. [Google Scholar] [CrossRef]
- Zhou, X.; Girón, J.A.; Torres, A.G.; Crawford, J.A.; Negrete, E.; Vogel, S.N.; Kaper, J.B. Flagellin of enteropathogenic Escherichia coli stimulates interleukin-8 production in T84 cells. Infect. Immun. 2003, 71, 2120–2129. [Google Scholar] [CrossRef]
- Sun, J.; Le, G.W.; Shi, Y.H.; Su, G.W. Factors involved in binding of Lactobacillus plantarum Lp6 to rat small intestinal mucus. Lett. Appl. Microbiol. 2007, 44, 79–85. [Google Scholar] [CrossRef]
- Collado, M.; Meriluoto, J.; Salminen, S. Role of commercial probiotic strains against human pathogen adhesion to intestinal mucus. Lett. Appl. Microbiol. 2007, 45, 454–460. [Google Scholar] [CrossRef]
- Pagnini, C.; Corleto, V.D.; Martorelli, M.; Lanini, C.; D’Ambra, G.; Di Giulio, E.; Delle Fave, G. Mucosal adhesion and anti-inflammatory effects of Lactobacillus rhamnosus GG in the human colonic mucosa: A proof-of-concept study. World J. Gastroenterol. 2018, 24, 4652. [Google Scholar] [CrossRef] [PubMed]
- von Ossowski, I.; Reunanen, J.; Satokari, R.; Vesterlund, S.; Kankainen, M.; Huhtinen, H.; Tynkkynen, S.; Salminen, S.; de Vos, W.M.; Palva, A. Mucosal adhesion properties of the probiotic Lactobacillus rhamnosus GG SpaCBA and SpaFED pilin subunits. Appl. Environ. Microbiol. 2010, 76, 2049–2057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamil, R.; Geier, M.S.; Butler, R.N.; Howarth, G.S. Lactobacillus rhamnosus GG exacerbates intestinal ulceration in a model of indomethacin-induced enteropathy. Dig. Dis. Sci. 2007, 52, 1247–1252. [Google Scholar] [CrossRef]
- Zein, E.; Karaa, S.; Chemaly, A.; Saidi, I.; Daou-Chahine, W.; Rohban, R. Lactobacillus rhamnosus septicemia in a diabetic patient associated with probiotic use: A case report. Proc. Ann. Biol. Clin. 2008, 66, 195–198. [Google Scholar]
- Shanahan, F. A commentary on the safety of probiotics. Gastroenterol. Clin. 2012, 41, 869–876. [Google Scholar] [CrossRef]
- Ouwehand, A.C.; Saxelins, M.; Salminen, S. Phenotypic differences between commercial Lactobacillus rhamnosus GG and L. rhamnosus strains recovered from blood. Clin. Infect. Dis. 2004, 39, 1858–1860. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Ref. | Numbers, Model, Age | Moulding Method | Probiotic Strains | CFU/Dose, Duration | Effects |
|---|---|---|---|---|---|
| [30] | 25, Dark Agouti rats, 30 d | TNBS | L. rhamnosus 64 | 3 × 106 CFU, daily, 27 d | damage score↓, immune cell infiltration↓, cytokine↓, MPO activity↓. |
| [31] | 90, BALB/c mice, 7 w | DSS | L. rhamnosus 1.0320 | 2 × 108 CFU, daily, 28 d | the DAI score↓, hemoglobin content↑, MPO activity↓, IL-1β↓, IL-6↓, TNF-α↓ and IL-10↑. |
| [32] | 48, Wistar rats, 10 w | Acid | L. rhamnosus EM1107 | 109 CFU, daily, 17 d | TNF-α↓, myeloperoxidase↓, IL-1β↓and oxidative stress↑. IL-17↓, NF-κB p65↓, MMP-2↓, MMP-9↓, and iNOS↓, SOCs-1↑, ZO-1↑ and mucin-2↑. |
| [33] | 32, C57BL/6J mice, 6 w | DSS | L. rhamnosus LDTM 7511 | 109 CFU, daily, 14 d | colon length↑, spleen weight↓, Lcn-2↓, MPO↓, CRP↑, relatively intact colonic architecture, Chao1 index↑, Shannon index↑. |
| [34] | 40, C57BL/6 mice, 8 w | DSS | L. rhamnosus L34 | 1 × 107 CFU, once every 3 d, 14 d | the gut local inflammation↓, gut-leakage severity↓, fecal dysbiosis↓ and systemic inflammation↓. |
| [35] | 40, C57BL/6Cnc mice, 8 w | DSS | L. rhamnosus SHA113 | 109 CFU, 9 d | SCFA-producing genera↑, UC-related genera↓. |
| [36] | 16, BALB/c mice, 6 w | DSS | L. rhamnosus OLL2838 | 107 CFU, 3 d | Body weight↑, and colon length↑, expression of zonula occludens-1 and myosin light-chain kinase↑ |
| [37] | 40, C57BL/6J mice, 5 w | DSS | L. rhamnosus Hao9 | 109 CFU, 7 d | DAI↓, colon length↑, alleviated colonic pathological variations, histological scores↓, TNF-α, IL-6, and IL-1β ↓, IL-10↑. |
| [38] | 50, C57BL/6 mice | DNBS | L. rhamnosus I-3690 | 5 × 109 CFU, 10 d | macroscopic scores↓, cytokine levels↓, colon and ileum MPO activities↓ |
| Ref. | Numbers of Patients | Disease | Probiotic Strains | CFU/Dose, Duration | Effects |
|---|---|---|---|---|---|
| [39] | 187 adults | UC | LGG | 18 × 109 CFU, 12 m | LGG treatment is more efficient than standard treatment in extending relapse-free time. |
| [40] | 4 children | CD | LGG | 1010 CFU, twice a day, 6 m | The median activity index of children with CD at 4 weeks was 73% lower than at baseline. |
| [41] | 11 adults | CD | LGG | 2 × 109 CFU, daily, 6 m | The median time to relapse was on average 4 weeks longer in the LGG group than in the placebo group. |
| [42] | 14 children | CD | LGG | 1010 CFU, twice a day, 10 d | Oral administration of LGG increased the intestinal IgA immune response, thus promoting the intestinal immune barrier. |
| [43] | 37 adults | CD | LGG | 6 × 109 CFU, twice a day, 52 w | LGG appears to neither prevent one-year endoscopic recurrence nor reduce the severity of recurrent disease. |
| [44] | 117 adults | CD&UC | LGG | 1.4 × 1010 CFU, daily, 52 w | Patients taking daily LGG had a lower frequency of first postoperative inflammatory bowel disease complications. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, H.; Yu, L.; Tian, F.; Chen, W.; Zhai, Q. The Potential Therapeutic Role of Lactobacillaceae rhamnosus for Treatment of Inflammatory Bowel Disease. Foods 2023, 12, 692. https://doi.org/10.3390/foods12040692
Guo H, Yu L, Tian F, Chen W, Zhai Q. The Potential Therapeutic Role of Lactobacillaceae rhamnosus for Treatment of Inflammatory Bowel Disease. Foods. 2023; 12(4):692. https://doi.org/10.3390/foods12040692
Chicago/Turabian StyleGuo, Hang, Leilei Yu, Fengwei Tian, Wei Chen, and Qixiao Zhai. 2023. "The Potential Therapeutic Role of Lactobacillaceae rhamnosus for Treatment of Inflammatory Bowel Disease" Foods 12, no. 4: 692. https://doi.org/10.3390/foods12040692