
Glycemic Responses of Milk and Plant-Based Drinks: Food Matrix Effects


Abstract
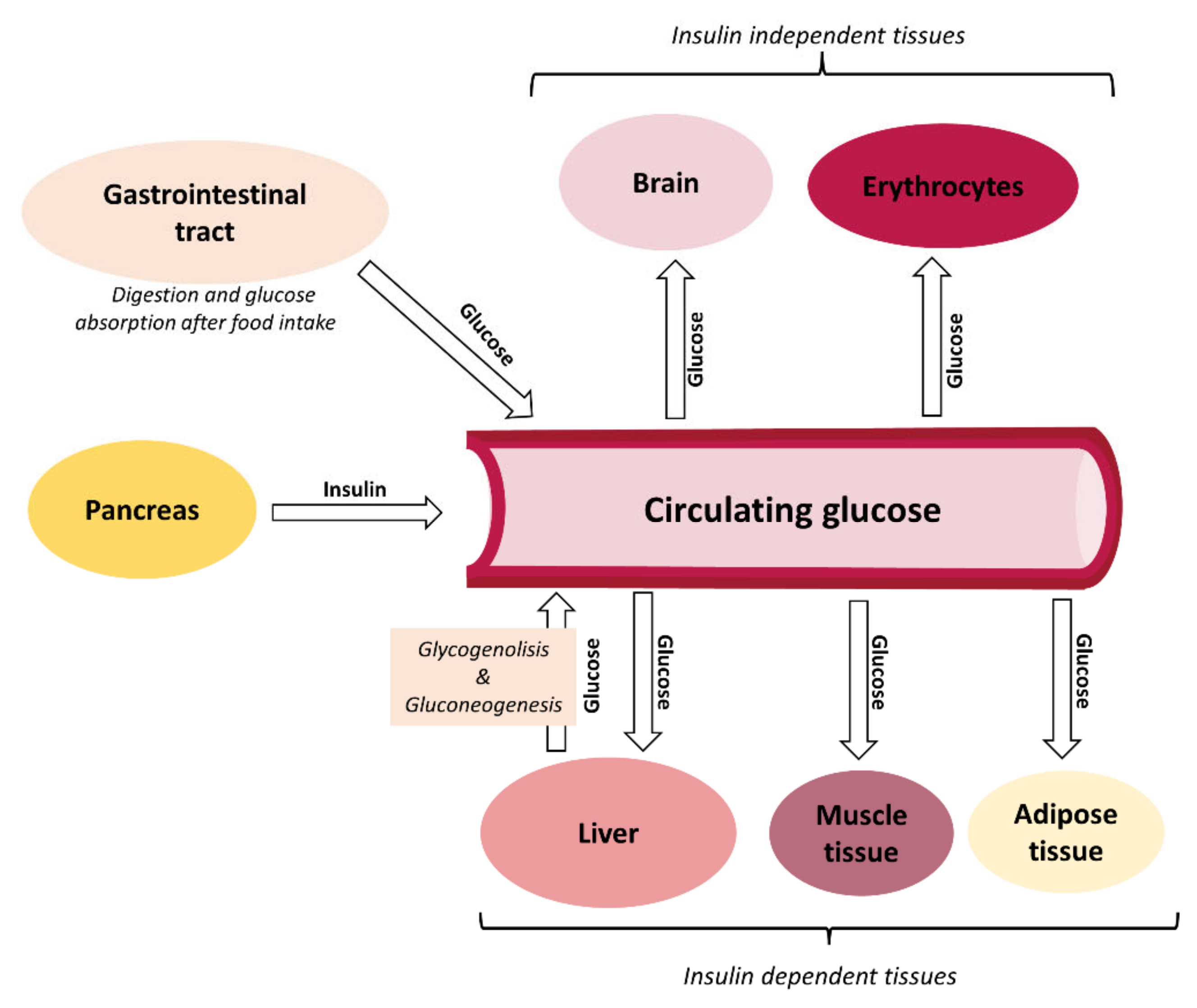
:1. Introduction
2. Postprandial Glycemic Responses
3. Postprandial Glycemic Responses: Food Matrix Effects
4. Glycemic Responses of Dairy Products
5. Glycemic Responses to Plant-Based Drinks Positioned as Dairy Alternatives
6. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- van Rossum, C.T.M.; Buurma-Rethans, E.J.M.; Dinnissen, C.S.; Beukers, M.H.; Brants, H.A.M.; Dekkers, A.L.M.; Ocké, M.C. The Diet of the Dutch: Results of the Dutch National Food Consumption Survey 2012–2016; National Institute for Public Health and the Environment RIVM: Bilthoven, The Netherlands, 2020. [Google Scholar]
- Huppertz, T. Lactose in Milk: Properties, Nutritional Characteristics and Role in Dairy Products. In Understanding and Improving the Functional and Nutritional Properties of Milk; Burleigh Dodds Science Publishing Limited: Cambridge, UK, 2022. [Google Scholar]
- Romero-Velarde, E.; Delgado-Franco, D.; García-Gutiérrez, M.; Gurrola-Díaz, C.; Larrosa-Haro, A.; Montijo-Barrios, E.; Muskiet, F.A.J.; Vargas-Guerrero, B.; Geurts, J. The Importance of Lactose in the Human Diet: Outcomes of a Mexican Consensus Meeting. Nutrients 2019, 11, 2737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClements, D.J.; Newman, E.; McClements, I.F. Plant-Based Milks: A Review of the Science Underpinning Their Design, Fabrication, and Performance. Compr. Rev. Food Sci. Food Saf. 2019, 18, 2047–2067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bocker, R.; Silva, E.K. Innovative Technologies for Manufacturing Plant-Based Non-Dairy Alternative Milk and Their Impact on Nutritional, Sensory and Safety Aspects. Future Foods 2022, 5, 100098. [Google Scholar] [CrossRef]
- Drewnowski, A.; Henry, C.J.; Dwyer, J.T. Proposed Nutrient Standards for Plant-Based Beverages Intended as Milk Alternatives. Front. Nutr. 2021, 8, 761442. [Google Scholar] [CrossRef] [PubMed]
- Jeske, S.; Zannini, E.; Arendt, E.K. Evaluation of Physicochemical and Glycaemic Properties of Commercial Plant-Based Milk Substitutes. Plant Foods Hum. Nutr. 2017, 72, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Smith, N.W.; Dave, A.C.; Hill, J.P.; McNabb, W.C. Nutritional Assessment of Plant-Based Beverages in Comparison to Bovine Milk. Front. Nutr. 2022, 9, 957486. [Google Scholar] [CrossRef]
- Augustin, L.S.A.; Kendall, C.W.C.; Jenkins, D.J.A.; Willett, W.C.; Astrup, A.; Barclay, A.W.; Björck, I.; Brand-Miller, J.C.; Brighenti, F.; Buyken, A.E.; et al. Glycemic Index, Glycemic Load and Glycemic Response: An International Scientific Consensus Summit from the International Carbohydrate Quality Consortium (ICQC). Nutr. Metab. Cardiovasc. Dis. 2015, 25, 795–815. [Google Scholar] [CrossRef] [Green Version]
- Blaak, E.E.; Antoine, J.M.; Benton, D.; Björck, I.; Bozzetto, L.; Brouns, F.; Diamant, M.; Dye, L.; Hulshof, T.; Holst, J.J.; et al. Impact of Postprandial Glycaemia on Health and Prevention of Disease. Obes. Rev. 2012, 13, 923–984. [Google Scholar] [CrossRef]
- Nakrani, M.N.; Wineland, R.H.; Anjum, F. Physiology, Glucose Metabolism; StatPearls Publishing: Tampa, FL, USA, 2021. [Google Scholar]
- American Diabetes Association. Diagnosis and Classification of Diabetes Mellitus. Diabetes Care 2006, 29, S43. [Google Scholar] [CrossRef]
- World Health Organization. Classification of Diabetes Mellitus; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Ceriello, A.; Colagiuri, S. International Diabetes Federation Guideline for Management of Postmeal Glucose: A Review of Recommendations. Diabet. Med. 2008, 25, 1151–1156. [Google Scholar] [CrossRef]
- Aronoff, S.L.; Berkowitz, K.; Shreiner, B.; Want, L. Glucose Metabolism and Regulation: Beyond Insulin and Glucagon. Diabetes Spectr. 2004, 17, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Maas, A.H.; Rozendaal, Y.J.W.; Van Pul, C.; Hilbers, P.A.J.; Cottaar, W.J.; Haak, H.R.; Van Riel, N.A.W. A Physiology-Based Model Describing Heterogeneity in Glucose Metabolism: The Core of the Eindhoven Diabetes Education Simulator (E-DES). J. Diabetes Sci. Technol. 2015, 9, 282–292. [Google Scholar] [CrossRef] [Green Version]
- Rozendaal, Y.J.; Maas, A.H.; van Pul, C.; Cottaar, E.J.; Haak, H.R.; Hilbers, P.A.; van Riel, N.A. Model-Based Analysis of Postprandial Glycemic Response Dynamics for Different Types of Food. Clin. Nutr. Exp. 2018, 19, 32–45. [Google Scholar] [CrossRef] [Green Version]
- Maggs, D.; Macdonald, I.; Nauck, M.A. Glucose Homeostasis and the Gastrointestinal Tract: Insights into the Treatment of Diabetes. Diabetes Obes. Metab. 2007, 10, 18–33. [Google Scholar] [CrossRef]
- Goyal, R.K.; Cristofaro, V.; Sullivan, M.P. Rapid Gastric Emptying in Diabetes Mellitus: Pathophysiology and Clinical Importance. J. Diabetes Complicat. 2019, 33, 107414. [Google Scholar] [CrossRef]
- Marroquí, L.; Alonso-Magdalena, P.; Merino, B.; Fuentes, E.; Nadal, A.; Quesada, I. Nutrient Regulation of Glucagon Secretion: Involvement in Metabolism and Diabetes. Nutr. Res. Rev. 2014, 27, 48–62. [Google Scholar] [CrossRef] [Green Version]
- Quesada, I.; Tudurí, E.; Ripoll, C.; Nadal, Á. Physiology of the Pancreatic α-Cell and Glucagon Secretion: Role in Glucose Homeostasis and Diabetes. J. Endocrinol. 2008, 199, 5–19. [Google Scholar] [CrossRef] [Green Version]
- Ramnanan, C.J.; Edgerton, D.S.; Kraft, G.; Cherrington, A.D. Physiologic Action of Glucagon on Liver Glucose Metabolism. Diabetes Obes. Metab. 2011, 13, 118–125. [Google Scholar] [CrossRef]
- Rui, L. Energy Metabolism in the Liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar] [CrossRef] [Green Version]
- Pessin, J.E.; Saltiel, A.R. Signaling Pathways in Insulin Action: Molecular Targets of Insulin Resistance. J. Clin. Investig. 2000, 106, 165–169. [Google Scholar] [CrossRef]
- Gautier, J.F.; Choukem, S.P.; Girard, J. Physiology of Incretins (GIP and GLP-1) and Abnormalities in Type 2 Diabetes. Diabetes Metab. 2008, 34, S65–S72. [Google Scholar] [CrossRef] [PubMed]
- Seino, Y.; Fukushima, M.; Yabe, D. GIP and GLP-1, the Two Incretin Hormones: Similarities and Differences. J. Diabetes Investig. 2010, 1, 8–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudaliar, S.; Henry, R.R. The Incretin Hormones: From Scientific Discovery to Practical Therapeutics. Diabetologia 2012, 55, 1865–1868. [Google Scholar] [CrossRef] [PubMed]
- van Ginneken, V. Are There Any Biomarkers of Aging? Biomarkers of the Brain. Biomed. J. Sci. Tech. Res. 2017, 1, 2017. [Google Scholar] [CrossRef]
- Wolever, T.M.S.; Bolognesi, C. Source and Amount of Carbohydrate Affect Postprandial Glucose and Insulin in Normal Subjects. J. Nutr. 1996, 126, 2798–2806. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.J.A.; Wolever, T.M.S.; Taylor, R.H.; Barker, H.; Fielden, H.; Baldwin, J.M.; Bowling, A.C.; Newman, H.C.; Goff, D.V. Glycemic Index of Foods: A Physiological Basis for Carbohydrate Exchange. Am. J. Clin. Nutr. 1981, 34, 362–366. [Google Scholar] [CrossRef] [Green Version]
- Carbohydrates in Human Nutrition. Report of a Joint FAO/WHO Expert Consultation. In FAO Food and Nutrition Paper; FAO: Rome, Italy, 1998; Volume 66, pp. 1–140. [Google Scholar]
- Brouns, F.; Bjorck, I.; Frayn, K.N.; Gibbs, A.L.; Lang, V.; Slama, G.; Wolever, T.M.S. Glycaemic Index Methodology. Nutr. Res. Rev. 2005, 18, 145–171. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, S.; Colagiuri, S.; Faramus, E.; Petocz, P.; Brand-Miller, J.C. Postprandial Hyperglycemia and Insulin Sensitivity Differ among Lean Young Adults of Different Ethnicities. J. Nutr. 2002, 132, 2574–2579. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, M.; Venn, B.J.; Williams, S.M.; Te Morenga, L.A.; Heemels, I.M.; Mann, J.I. Glycaemic Responses to Glucose and Rice in People of Chinese and European Ethnicity. Diabet. Med. 2013, 30, e101–e107. [Google Scholar] [CrossRef]
- Wolever, T.M.S.; Jenkins, A.L.; Vuksan, V.; Campbell, J. The Glycaemic Index Values of Foods Containing Fructose Are Affected by Metabolic Differences between Subjects. Eur. J. Clin. Nutr. 2009, 63, 1106–1114. [Google Scholar] [CrossRef]
- Venn, B.J.; Williams, S.M.; Mann, J.I. Comparison of Postprandial Glycaemia in Asians and Caucasians. Diabet. Med. 2010, 27, 1205–1208. [Google Scholar] [CrossRef]
- Henry, C.J.K.; Lightowler, H.J.; Newens, K.; Sudha, V.; Radhika, G.; Sathya, R.M.; Mohan, V. Glycaemic Index of Common Foods Tested in the UK and India. British. J. Nutr. 2008, 99, 840–845. [Google Scholar] [CrossRef] [Green Version]
- Chan, H.M.S.; Brand-Miller, J.C.; Holt, S.H.A.; Wilson, D.; Rozman, M.; Petocz, P. The Glycaemic Index Values of Vietnamese Foods. Eur. J. Clin. Nutr. 2001, 55, 1076–1083. [Google Scholar] [CrossRef] [Green Version]
- Venn, B.J.; Green, T.J. Glycemic Index and Glycemic Load: Measurement Issues and Their Effect on Diet–Disease Relationships. Eur. J. Clin. Nutr. 2007, 61, S122–S131. [Google Scholar] [CrossRef]
- Atkinson, F.S.; Foster-Powell, K.; Brand-Miller, J.C. International Tables of Glycemic Index and Glycemic Load Values: 2008. Diabetes Care 2008, 31, 2281–2283. [Google Scholar] [CrossRef] [Green Version]
- Eleazu, C.O. The Concept of Low Glycemic Index and Glycemic Load Foods as Panacea for Type 2 Diabetes Mellitus; Prospects, Challenges and Solutions. Afr. Health. Sci. 2016, 16, 468–479. [Google Scholar] [CrossRef] [Green Version]
- Wolever, T.M.S. Yogurt Is a Low-Glycemic Index Food. J. Nutr. 2017, 147, 1462S–1467S. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.T.; Lampe, J.W.; Schwarz, Y.; Breymeyer, K.L.; Noar, K.A.; Song, X.; Neuhouser, M.L. Low Glycemic Load Experimental Diet More Satiating than High Glycemic Load Diet. Nutr. Cancer 2012, 64, 666–673. [Google Scholar] [CrossRef] [Green Version]
- Ojo, O.; Ojo, O.O.; Adebowale, F.; Wang, X.H. The Effect of Dietary Glycaemic Index on Glycaemia in Patients with Type 2 Diabetes: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2018, 10, 373. [Google Scholar] [CrossRef] [Green Version]
- Wolever, T.M.S. Is Glycaemic Index (GI) a Valid Measure of Carbohydrate Quality? Eur. J. Clin. Nutr. 2013, 67, 522–531. [Google Scholar] [CrossRef]
- McQuilken, S.A. Digestion and Absorption. Anaesth. Intensive Care Medic. 2021, 22, 336–338. [Google Scholar] [CrossRef]
- Hall, J.E. Guyton and Hall Textbook of Medical Physiology, 12th ed.; Saunders: Philadelphia, PA, USA, 2011. [Google Scholar]
- Dashty, M. A Quick Look at Biochemistry: Carbohydrate Metabolism. Clin. Biochem. 2013, 46, 1339–1352. [Google Scholar] [CrossRef] [PubMed]
- Goodman, B.E. Insights into Digestion and Absorption of Major Nutrients in Humans. Am. J. Physiol. 2010, 34, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Sitrin, M.D. Digestion and Absorption of Carbohydrates and Proteins. In The Gastrointestinal System: Gastrointestinal, Nutritional and Hepatobiliary Physiology; Springer: Dordrecht, The Netherlands, 2014; ISBN 9789401787710. [Google Scholar]
- Holmes, R. Carbohydrate Digestion and Absorption. J. Clin. Pathol. 1971, s3-5, 10–13. [Google Scholar] [CrossRef] [Green Version]
- MacFarlane, N.G. Digestion and Absorption. Anaesth. Intensive Care Medic. 2018, 19, 125–127. [Google Scholar] [CrossRef] [Green Version]
- Adeva-Andany, M.M.; Pérez-Felpete, N.; Fernández-Fernández, C.; Donapetry-García, C.; Pazos-García, C. Liver Glucose Metabolism in Humans. Biosci. Rep. 2016, 36, e00416. [Google Scholar] [CrossRef] [Green Version]
- Hantzidiamantis, P.J.; Lappin, S.L. Physiology, Glucose. StatPearls Publishing: Tampa, FL, USA, 2019. [Google Scholar]
- Aston, L.M.; Gambell, J.M.; Lee, D.M.; Bryant, S.P.; Jebb, S.A. Determination of the Glycaemic Index of Various Staple Carbohydrate-Rich Foods in the UK Diet. Eur. J. Clin. Nutr. 2008, 62, 279–285. [Google Scholar] [CrossRef] [Green Version]
- Mackie, A.R.; Bajka, B.H.; Rigby, N.M.; Wilde, P.J.; Alves-Pereira, F.; Mosleth, E.F.; Rieder, A.; Kirkhus, B.; Salt, L.J. Oatmeal Particle Size Alters Glycemic Index but Not as a Function of Gastric Emptying Rate. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G239–G246. [Google Scholar] [CrossRef] [Green Version]
- Collier, G.; McLean, A.; O’Dea, K. Effect of Co-Ingestion of Fat on the Metabolic Responses to Slowly and Rapidly Absorbed Carbohydrates. Diabetologia 1984, 26, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Moghaddam, E.; Vogt, J.A.; Wolever, T.M.S. The Effects of Fat and Protein on Glycemic Responses in Nondiabetic Humans Vary with Waist Circumference, Fasting Plasma Insulin, and Dietary Fiber Intake. J. Nutr. 2006, 136, 2506–2511. [Google Scholar] [CrossRef]
- Nuttall, F.Q.; Mooradian, A.D.; Gannon, M.C.; Billington, C.J.; Krezowski, P. Effect of Protein Ingestion on the Glucose and Insulin Response to a Standardized Oral Glucose Load. Diabetes Care 1984, 7, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Jin, Y.; Wilde, P.J.; Hou, Y.; Wang, Y.; Han, J. Mechanisms, Physiology, and Recent Research Progress of Gastric Emptying. Crit. Rev. Food Sci. Nutr. 2021, 61, 2742–2755. [Google Scholar] [CrossRef] [PubMed]
- Thorning, T.K.; Bertram, H.C.; Bonjour, J.P.; De Groot, L.; Dupont, D.; Feeney, E.; Ipsen, R.; Lecerf, J.M.; Mackie, A.; McKinley, M.C.; et al. Whole Dairy Matrix or Single Nutrients in Assessment of Health Effects: Current Evidence and Knowledge Gaps. Am. J. Clin. Nutr. 2017, 105, 1033–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilera, J.M. The Food Matrix: Implications in Processing, Nutrition and Health. Crit. Rev. Food Sci. Nutr. 2019, 59, 3612–3629. [Google Scholar] [CrossRef] [PubMed]
- Capuano, E.; Janssen, A.E.M. Food Matrix and Macronutrient Digestion. Annu. Rev. Food Sci. Technol. 2021, 12, 193–212. [Google Scholar] [CrossRef]
- Kjølbæk, L.; Schmidt, J.M.; Rouy, E.; Jensen, K.J.; Astrup, A.; Bertram, H.C.; Hammershøj, M.; Raben, A. Matrix Structure of Dairy Products Results in Different Postprandial Lipid Responses: A Randomized Crossover Trial. Am. J. Clin. Nutr. 2021, 114, 1729–1742. [Google Scholar] [CrossRef]
- Feeney, E.L.; Barron, R.; Dible, V.; Hamilton, Z.; Power, Y.; Tanner, L.; Flynn, C.; Bouchier, P.; Beresford, T.; Noronha, N.; et al. Dairy Matrix Effects: Response to Consumption of Dairy Fat Differs When Eaten within the Cheese Matrix—A Randomized Controlled Trial. Am. J. Clin. Nutr. 2018, 108, 667–674. [Google Scholar] [CrossRef] [Green Version]
- Pennings, B.; Groen, B.B.L.; Van Dijk, J.W.; De Lange, A.; Kiskini, A.; Kuklinski, M.; Senden, J.M.G.; Van Loon, L.J.C. Minced Beef Is More Rapidly Digested and Absorbed than Beef Steak, Resulting in Greater Postprandial Protein Retention in Older Men. Am. J. Clin. Nutr. 2013, 98, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Horstman, A.M.H.; Ganzevles, R.A.; Kudla, U.; Kardinaal, A.F.M.; van den Borne, J.J.G.C.; Huppertz, T. Postprandial Blood Amino Acid Concentrations in Older Adults after Consumption of Dairy Products: The Role of the Dairy Matrix. Int. Dairy. J. 2021, 113, 104890. [Google Scholar] [CrossRef]
- Shkembi, B.; Huppertz, T. Calcium Absorption from Food Products: Food Matrix Effects. Nutrients 2021, 14, 180. [Google Scholar] [CrossRef]
- Shkembi, B.; Huppertz, T. Influence of Dairy Products on Bioavailability of Zinc from Other Food Products: A Review of Complementarity at a Meal Level. Nutrients 2021, 13, 4253. [Google Scholar] [CrossRef]
- Haber, G.B.; Heaton, K.W.; Murphy, D.; Burroughs, L.F. Depletion and Disruption of Dietary Fibre. Lancet 1977, 310, 679–682. [Google Scholar] [CrossRef]
- Bolton, R.P.; Heaton, K.W.; Burroughs, L.F. The Role of Dietary Fiber in Satiety, Glucose, and Insulin: Studies with Fruit and Fruit Juice. Am. J. Clin. Nutr. 1981, 34, 211–217. [Google Scholar] [CrossRef] [Green Version]
- Tey, S.L.; Lee, D.E.M.; Henry, C.J. Fruit Form Influences Postprandial Glycemic Response in Elderly and Young Adults. J. Nutr. Health Aging 2017, 21, 887–891. [Google Scholar] [CrossRef]
- Jenkins, D.J.; Wolever, T.M.; Taylor, R.H.; Griffiths, C.; Krzeminska, K.; Lawrie, J.A.; Bennett, C.M.; Goff, D.V.; Sarson, D.L.; Bloom, S.R. Slow Release Dietary Carbohydrate Improves Second Meal Tolerance. Am. J. Clin. Nutr. 1982, 35, 1339–1346. [Google Scholar] [CrossRef]
- Reynolds, A.N.; Mann, J.; Elbalshy, M.; Mete, E.; Robinson, C.; Oey, I.; Silcock, P.; Downes, N.; Perry, T.; Te Morenga, L. Wholegrain Particle Size Influences Postprandial Glycemia in Type 2 Diabetes: A Randomized Crossover Study Comparing Four Wholegrain Breads. Diabetes Care 2020, 43, 476–479. [Google Scholar] [CrossRef]
- Jenkins, D.J.A.; Thorne, M.J.; Wolever, T.M.S.; Rao, A.V.; Thompson, L.U. The Effect of Starch-Protein Interaction in Wheat on the Glycemic Response and Rate of in Vitro Digestion. Am. J. Clin. Nutr. 1987, 45, 946–951. [Google Scholar] [CrossRef] [Green Version]
- Damodaran, S.; Parkin, K.L.; Fennema, O.R. Fennema’s Food Chemistry, 4th ed.; CRC press: Boca Raton, FL, USA, 2007; ISBN 978-0-8247-2345-3. [Google Scholar]
- Touger-Decker, R.; van Loveren, C. Sugars and Dental Caries. Am. J. Clin. Nutr. 2003, 78, 881S–892S. [Google Scholar] [CrossRef] [Green Version]
- Giacaman, R.A. Sugars and beyond. The Role of Sugars and the Other Nutrients and Their Potential Impact on Caries. Oral Dis. 2017, 24, 1185–1197. [Google Scholar] [CrossRef]
- Aimutis, W.R. Lactose Cariogenicity with an Emphasis on Childhood Dental Caries. Int. Dairy J. 2012, 22, 152–158. [Google Scholar] [CrossRef]
- Chen, Y.Y.M.; Betzenhauser, M.J.; Snyder, J.A.; Burne, R.A. Pathways for Lactose/Galactose Catabolism by Streptococcus Salivarius. FEMS Microbiol. Lett. 2002, 209, 72–76. [Google Scholar] [CrossRef]
- Woodward, M.; Rugg-Gunn, A.J. Milk, Yoghurts and Dental Caries. Impact Nutr. Diet Oral Health 2020, 28, 77–90. [Google Scholar]
- Gille, D.; Walther, B.; Badertscher, R.; Bosshart, A.; Brügger, C.; Brühlhart, M.; Gauch, R.; Noth, P.; Vergères, G.; Egger, L. Detection of Lactose in Products with Low Lactose Content. Int. Dairy. J. 2018, 83, 17–19. [Google Scholar] [CrossRef]
- Gray, G.M.; Santiago, N.A. Disaccharide Absorption in Normal and Diseased Human Intestine. Gastroenterology 1966, 51, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Gray, G.M. Carbohydrate Digestion and Absorption. Gastroenterology 1970, 58, 96–107. [Google Scholar] [CrossRef]
- Henry, C.J.K.; Lightowler, H.J.; Strik, C.M.; Renton, H.; Hails, S. Glycaemic Index and Glycaemic Load Values of Commercially Available Products in the UK. Br. J. Nutr. 2005, 94, 922–930. [Google Scholar] [CrossRef] [Green Version]
- Hoyt, G.; Hickey, M.S.; Cordain, L. Dissociation of the Glycaemic and Insulinaemic Responses to Whole and Skimmed Milk. Br. J. Nutr. 2005, 93, 175–177. [Google Scholar] [CrossRef] [Green Version]
- Östman, E.M.; Liljeberg Elmståhl, H.G.M.; Björck, I.M.E. Inconsistency between Glycemic and Insulinemic Responses to Regular and Fermented Milk Products. Am. J. Clin. Nutr. 2001, 74, 96–100. [Google Scholar] [CrossRef] [Green Version]
- Anafy, A.; Moran-Lev, H.; Shapira, N.; Priel, M.; Oren, A.; Mangel, L.; Mandel, D.; Lubetzky, R. The Glycemic Response to Infant Formulas: A Randomized Clinical Trial. Nutrients 2022, 14, 1064. [Google Scholar] [CrossRef]
- Choi, H.K.; Willett, W.C.; Stampfer, M.J.; Rimm, E.; Hu, F.B. Dairy Consumption and Risk of Type 2 Diabetes Mellitus in Men: A Prospective Study. Arch. Intern. Med. 2005, 165, 997–1003. [Google Scholar] [CrossRef] [Green Version]
- Salas-Salvadó, J.; Guasch-Ferré, M.; Díaz-López, A.; Babio, N. Yogurt and Diabetes: Overview of Recent Observational Studies. J. Nutr. 2017, 147, 1452S–1461S. [Google Scholar] [CrossRef]
- Chen, M.; Sun, Q.; Giovannucci, E.; Mozaffarian, D.; Manson, J.A.E.; Willett, W.C.; Hu, F.B. Dairy Consumption and Risk of Type 2 Diabetes: 3 Cohorts of US Adults and an Updated Meta-Analysis. BMC Med. 2014, 12, 215. [Google Scholar] [CrossRef] [Green Version]
- Hidayat, K.; Du, X.; Shi, B.M. Milk in the Prevention and Management of Type 2 Diabetes: The Potential Role of Milk Proteins. Diabetes Metab. Res. Rev. 2019, 35, e3187. [Google Scholar] [CrossRef]
- Boirie, Y.; Dangin, M.; Gachon, P.; Vasson, M.P.; Maubois, J.L.; Beaufrère, B. Slow and Fast Dietary Proteins Differently Modulate Postprandial Protein Accretion. Proc. Natl. Acad. Sci. USA 1997, 94, 14930–14935. [Google Scholar] [CrossRef] [Green Version]
- Horstman, A.M.H.; Huppertz, T. Milk Proteins: Processing, Gastric Coagulation, Amino Acid Availability and Muscle Protein Synthesis. Crit. Rev. Food Sci. Nutr. 2022, 62, 1–16. [Google Scholar] [CrossRef]
- Nouri, M.; Pourghassem Gargari, B.; Tajfar, P.; Tarighat-Esfanjani, A. A Systematic Review of Whey Protein Supplementation Effects on Human Glycemic Control: A Mechanistic Insight. Diabetes Metab. Syndr. Clin. Res. Rev. 2022, 16, 102540. [Google Scholar] [CrossRef]
- Chiang, S.-W.; Liu, H.-W.; Loh, E.-W.; Tam, K.-W.; Wang, J.-Y.; Huang, W.-L.; Kuan, Y.-C. Whey Protein Supplementation Improves Postprandial Glycemia in Persons with Type 2 Diabetes Mellitus: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutr. Res. Rev. 2022, 104, 44–54. [Google Scholar] [CrossRef]
- Smith, K.; Taylor, G.S.; Brunsgaard, L.H.; Walker, M.; Bowden Davies, K.A.; Stevenson, E.J.; West, D.J. Thrice Daily Consumption of a Novel, Premeal Shot Containing a Low Dose of Whey Protein Increases Time in Euglycemia during 7 Days of Free-Living in Individuals with Type 2 Diabetes. BMJ Open Diabetes Res. Care 2022, 104, 44–54. [Google Scholar] [CrossRef]
- Gao, K.P.; Mitsui, T.; Fujiki, K.; Ishiguro, H.; Kondo, T. Effect of Lactase Preparations in Asymptomatic Individuals with Lactase Deficiency--Gastric Digestion of Lactose and Breath Hydrogen Analysis. Nagoya J. Med. Sci. 2002, 65, 21–28. [Google Scholar]
- Huppertz, T. Lactose in Yogurt. In Yogurt in Health and Disease Prevention; Academic Press: Cambridge, MA, USA, 2017; pp. 387–394. [Google Scholar]
- Buchheim, W. Influences of Different Technological Treatments of Milk on the Digestion in the Stomach. IV. Electron Microscopical Characterization of the Coagulum and of Lipolytic Processes in the Stomach. Milchwissenschaft 1984, 19, 271–275. [Google Scholar]
- Pfeil, R. Influence of Different Technological Treatments of Milk on the Digestion in the Stomach. III. Proteolysis in the Stomach. Milchwissenschaft 1984, 39, 267–270. [Google Scholar]
- Walstra, P.; Walstra, P.; Wouters, J.T.M.; Geurts, T.J. Dairy Science and Technology; CRC press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Pedersen, M.G.B.; Søndergaard, E.; Nielsen, C.B.; Johannsen, M.; Gormsen, L.C.; Møller, N.; Jessen, N.; Rittig, N. Oral Lactate Slows Gastric Emptying and Suppresses Appetite in Young Males. Clin. Nutr. 2022, 41, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Chalupa-Krebzdak, S.; Long, C.J.; Bohrer, B.M. Nutrient Density and Nutritional Value of Milk and Plant-Based Milk Alternatives. Int. Dairy J. 2018, 87, 84–92. [Google Scholar] [CrossRef]
- McCarthy, K.S.; Parker, M.; Ameerally, A.; Drake, S.L.; Drake, M.A. Drivers of Choice for Fluid Milk versus Plant-Based Alternatives: What Are Consumer Perceptions of Fluid Milk? J. Dairy Sci. 2017, 100, 6125–6138. [Google Scholar] [CrossRef] [PubMed]
- Scholz-Ahrens, K.E.; Ahrens, F.; Barth, C.A. Nutritional and Health Attributes of Milk and Milk Imitations. Eur. J. Nutr. 2020, 59, 19–34. [Google Scholar] [CrossRef]
- Clegg, M.E.; Tarrado Ribes, A.; Reynolds, R.; Kliem, K.; Stergiadis, S. A Comparative Assessment of the Nutritional Composition of Dairy and Plant-Based Dairy Alternatives Available for Sale in the UK and the Implications for Consumers’ Dietary Intakes. Food Res. Int. 2021, 148, 110586. [Google Scholar] [CrossRef]
- Tangyu, M.; Muller, J.; Bolten, C.J.; Wittmann, C. Fermentation of Plant-Based Milk Alternatives for Improved Flavour and Nutritional Value. Appl. Microbiol. Biotechnol. 2019, 103, 9263–9275. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.R.A.; Silva, M.M.N.; Ribeiro, B.D. Health Issues and Technological Aspects of Plant-Based Alternative Milk. Food Res. Int. 2020, 131, 108972. [Google Scholar] [CrossRef]
- McClements, D.J. Development of Next-Generation Nutritionally Fortified Plant-Based Milk Substitutes: Structural Design Principles. Foods 2020, 9, 421. [Google Scholar] [CrossRef] [Green Version]
- Paul, A.A.; Kumar, S.; Kumar, V.; Sharma, R. Milk Analog: Plant Based Alternatives to Conventional Milk, Production, Potential and Health Concerns. Crit. Rev. Food Sci. Nutr. 2020, 60, 3005–3023. [Google Scholar] [CrossRef]
- Aydar, E.F.; Tutuncu, S.; Ozcelik, B. Plant-Based Milk Substitutes: Bioactive Compounds, Conventional and Novel Processes, Bioavailability Studies, and Health Effects. J. Funct. Foods 2020, 70, 103975. [Google Scholar] [CrossRef]
- Singh-Povel, C.M.; Van Gool, M.P.; Gual Rojas, A.P.; Bragt, M.C.E.; Kleinnijenhuis, A.J.; Hettinga, K.A. Nutritional Content, Protein Quantity, Protein Quality and Carbon Footprint of Plant-Based Drinks and Semi-Skimmed Milk in the Netherlands and Europe. Public Health Nutr. 2022, 25, 1416–1426. [Google Scholar] [CrossRef]
- Word Health Organization. Guideline: Sugars Intake for Adults and Children; WHO: Geneva, Switzerland, 2015. [Google Scholar]
- Foster-Powell, K.; Holt, S.H.A.; Brand-Miller, J.C. International Table of Gylcemic Index and Glycemic Load Values: 2002. Am. J. Clin. Nutr. 2002, 76, 5–56. [Google Scholar] [CrossRef] [Green Version]
- Vogelsang-O’Dwyer, M.; Sahin, A.W.; Zannini, E.; Arendt, E.K. Physicochemical and Nutritional Properties of High Protein Emulsion-Type Lupin-Based Model Milk Alternatives: Effect of Protein Source and Homogenization Pressure. J. Sci. Food Agric. 2021, 102, 5086–5097. [Google Scholar] [CrossRef]
- Pineli, L.L.O.; Botelho, R.B.A.; Zandonadi, R.P.; Solorzano, J.L.; de Oliveira, G.T.; Reis, C.E.G.; Teixeira, D.d.S. Low Glycemic Index and Increased Protein Content in a Novel Quinoa Milk. LWT-Food Sci. Technol. 2015, 63, 1261–1267. [Google Scholar] [CrossRef]
- Shin, H.S.; Ingram, J.R.; McGill, A.T.; Poppitt, S.D. Lipids, CHOs, Proteins: Can All Macronutrients Put a “brake” on Eating? Physiol. Behav. 2013, 120, 114–123. [Google Scholar] [CrossRef]
- Fardet, A.; Leenhardt, F.; Lioger, D.; Scalbert, A.; Rémésy, C. Parameters Controlling the Glycaemic Response to Breads. Nutr. Res. Rev. 2006, 19, 18–25. [Google Scholar] [CrossRef]
- Graf, B.L.; Poulev, A.; Kuhn, P.; Grace, M.H.; Lila, M.A.; Raskin, I. Quinoa Seeds Leach Phytoecdysteroids and Other Compounds with Anti-Diabetic Properties. Food Chem. 2014, 163, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Tan, K.W.J.; Siow, P.C.; Henry, C.J. Soya Milk Exerts Different Effects on Plasma Amino Acid Responses and Incretin Hormone Secretion Compared with Cows’ Milk in Healthy, Young Men. Br. J. Nutr. 2016, 116, 1216–1221. [Google Scholar] [CrossRef]


{kind=link}
| Foods | Experimental Portion § (g or mL) | GI | II | GL (Per Serving) | Standard Serving Size * (g) | Carbohydrate (g/100 g) | Available Carbohydrate (g) | Sugar | Protein (g) | Fat (g) | Subjects (n) | Subjects Type | References |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Milk, full fat | 39 | ||||||||||||
| Milk, skim | DNS | 37 | DNS | DNS | DNS | DNS | DNS | DNS | DNS | DNS | DNS | DNS | [40] |
| Ice cream | 51 | ||||||||||||
| Yoghurt, fruit | 41 | ||||||||||||
| Milk, skimmed, pasteurised, British | 500 g | 48 | 6 | 250 | 5.0 | 12.5 (g per serving) | 10 | ||||||
| Milk, semi-skimmed, pasteurised, British | 500 g | 25 | 3 | 250 | 5.0 | 12.5 (g per serving) | 8 | ||||||
| Milk, semi-skimmed, pasteurised, organic | 500 g | 34 | DNS | 4 | 250 | 5.0 | 12.5 (g per serving) | DNS | DNS | DNS | 9 | Healthy subjects | [85] |
| Milk, standardised homogenised, pasteurised, British | 531.9 g | 46 | 5 | 250 | 4.7 | 11.8 (g per serving) | 10 | ||||||
| Milk, whole, pasteurised, fresh, organic | 520.8 g | 34 | 4 | 250 | 4.8 | 12.0 (g per serving) | 9 | ||||||
| Yoghurt, low fat, natural | 431 g | 35 | 4 | 200 | 5.8 | 11.6 (g per serving) | 9 | ||||||
| Whole milk | DNS | 41 | 148 | DNS | DNS | DNS | 25 | DNS | DNS | DNS | 9 | Healthy subjects | [86] |
| Skimmed milk | 37 | 140 | 25 | 8 | |||||||||
| Regular milk | 30 | 90 | 25 | 17.3 | 15.3 | 10 | |||||||
| Fermented milk (Filmjölk) | DNS | 15 | 98 | DNS | DNS | DNS | 25 | DNS | 19.2 | 17.4 | 10 | Healthy subjects | [87] |
| Fermented milk (Ropy milk) | 15 | 97 | 25 | 19.3 | 17.0 | 10 | |||||||
| Lactose | 68 | 50 | 24 | DNS | DNS | 10 | |||||||
| Ice cream | 36 | 50 | 5 | ||||||||||
| Milk (skim) | 32 | 50 | 6 | ||||||||||
| Milk (whole) | DNS | 34 | DNS | DNS | DNS | DNS | 50 | DNS | DNS | DNS | 6 | Healthy subjects | [30] |
| Yoghurt | 36 | 50 | 5 | ||||||||||
| Milk, full-cream condensed, sweetened | 90.2 g | 61 | DNS | DNS | DNS | DNS | 55.4 (g/100g) | DNS | DNS | DNS | 11 | Healthy subjects | [38] |
| Bovine milk protein- based formula (whey 60%, casein 40%) | 200 mL | 21.5 | DNS | DNS | DNS | Lactose (100%) | 50 (g per experimental portion) | DNS | 9 (g per experimental portion) | DNS | 20 | Healthy subjects | [88] |
| Lactose-free formula (whey 60%, casein 40%) | 200 mL | 21.5 | Glucose syrup solids (100%) | 50 (g per experimental portion) | 9 (g per experimental portion) | 20 |
| Foods | Experimental Portion § (mL) | GI | II | GL (Per Portion) | Portion Size * (g or mL) | Carbohydrate (g) | Available carbohydrate (g) | Sugar (g/100 g) | Protein (g/100 g) | Fat (g/100g) | Subjects (n) | Subjects Type | References |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bovine milk, full fat | 39 | ||||||||||||
| Bovine milk, skim | DNS | 37 | DNS | DNS | DNS | DNS | DNS | DNS | DNS | DNS | DNS | DNS | [40] |
| Soy milk | 34 | ||||||||||||
| Rice milk | 86 | ||||||||||||
| Bovine milk | DNS | 39 | 4.80 | DNS | 5.05 | 3.15 | 3.25 | DNS | |||||
| Quinoa milk | 312.5 | 52 | DNS | DNS | DNS | 14.7 | 50 (g per experimental portion) | 9.7 | 1.7 | 0.2 | 12 | Healthy subjects | [117] |
| Rice milk | DNS | 79 | 12.7 | DNS | 7.3 | 0.30 | 1.11 | DNS | |||||
| Bovine milk protein-based formula (whey 60%, casein 40%) | 200 | 21.5 | Lactose (100%) | 50 (g per experimental portion) | 9 g (g per experimental portion) | 20 | |||||||
| Soy protein-based formula (soy 100%) | 200 | 29.1 | DNS | DNS | DNS | Glucose syrup solids (100%) | 50 (g per experimental portion) | DNS | 9 g (g per experimental portion) | DNS | 20 | Healthy subjects | [88] |
| Lactose-free formula (whey 60%, casein 40%) | 200 | 21.5 | Glucose syrup solids (100%) | 50 (g per experimental portion) | 9 g (g per experimental portion) | 20 | |||||||
| Bovine milk | 44.66 | 5.08 | 250 mL | 0.5 | Lactose (4.55 g/100 mL) | 3.83% | 1.28% | ||||||
| Blue lupin milk alternative | 51.15 | 2.98 | 250 mL | 0.5 | Sucrose (2.33 g/100 mL) | 3.54% | 1.43% | ||||||
| White lupin milk alternative | DNS | 53.71 | DNS | 3.06 | 250 mL | DNS | 0.5 | Sucrose (2.28 g/100 mL) | 3.44% | 1.37% | In vitro | In vitro | [116] |
| Soy milk alternative | 57.73 | 3.76 | 250 mL | 0.5 | Sucrose (2.43 g/100 mL) and raffinose/stachyose (0.189 g/100 mL) | 3.20% | 1.69% | ||||||
| Almond MLK | 58.68 | 0.94 | 250 g | 0.5 | 0.58 | 2.11 | 4.40 | ||||||
| Almond original | 49.10 | 4.60 | 250 g | 0.5 | 3.69 | 0.41 | 1.18 | ||||||
| Organic almond drink | 64.21 | 0.37 | 250 g | 0.5 | 0.16 | 0.95 | 3.69 | ||||||
| Carob almond MLK | 54.33 | 6.32 | 250 g | 0.5 | 4.58 | 2.4 | 3.35 | ||||||
| Organic cashew drink | 52.82 | 4.76 | 250 g | 0.5 | 2.87 | 0.87 | 2.50 | ||||||
| Coconut original | 96.82 | 4.81 | 250 g | 0.5 | 1.86 | 0.08 | 0.84 | ||||||
| Hazelnut original | DNS | 55.76 | DNS | 4.37 | 250 g | DNS | 0.5 | 3.09 | 0.36 | 1.56 | In vitro | In vitro | [7] |
| Hemp milk unsweetened | 59.94 | 0.21 | 250 g | 0.5 | 0.09 | 0.08 | 2.44 | ||||||
| Organic macadamia drink | 49.47 | 3.71 | 250 g | 0.5 | 2.79 | 0.29 | 2.62 | ||||||
| Organic oat drink | 59.61 | 7.98 | 250 g | 0.5 | 3.35 | 0.70 | 0.38 | ||||||
| Quinoa drink | 53.28 | 4.51 | 250 g | 0.5 | 3.2 | 0.22 | 2.32 | ||||||
| Organic rice drink natural | 97.74 | 18.33 | 250 g | 0.5 | 7.02 | 0.32 | 0.85 | ||||||
| Organic brown rice drink | 99.96 | 16.85 | 250 g | 0.5 | 5.58 | 0.07 | 0.95 | ||||||
| Organic soya drink, calcium | 47.53 | 3.01 | 250 g | 0.5 | 2.43 | 2.72 | 2.11 | ||||||
| Plain UHT organic soya drink | 54.02 | 1.24 | 250 g | 0.5 | 0.88 | 3.70 | 2.04 | ||||||
| Soya organic, wholebean | 49.49 | 0.57 | 250 g | 0.5 | 0.36 | 3.16 | 1.77 | ||||||
| Soya original | 61.50 | 4.87 | 250 g | 0.5 | 3.09 | 2.61 | 1.48 | ||||||
| Fresh milk, pasteurised & homogenised | 46.93 | 4.03 | 250 g | 0.5 | 3.38 | 3.70 | 3.28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shkembi, B.; Huppertz, T. Glycemic Responses of Milk and Plant-Based Drinks: Food Matrix Effects. Foods 2023, 12, 453. https://doi.org/10.3390/foods12030453
Shkembi B, Huppertz T. Glycemic Responses of Milk and Plant-Based Drinks: Food Matrix Effects. Foods. 2023; 12(3):453. https://doi.org/10.3390/foods12030453
Chicago/Turabian StyleShkembi, Blerina, and Thom Huppertz. 2023. "Glycemic Responses of Milk and Plant-Based Drinks: Food Matrix Effects" Foods 12, no. 3: 453. https://doi.org/10.3390/foods12030453