
Agaricus bisporus Polysaccharides Ameliorates Behavioural Deficits in D-Galactose-Induced Aging Mice: Mediated by Gut Microbiota

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction of the WMP
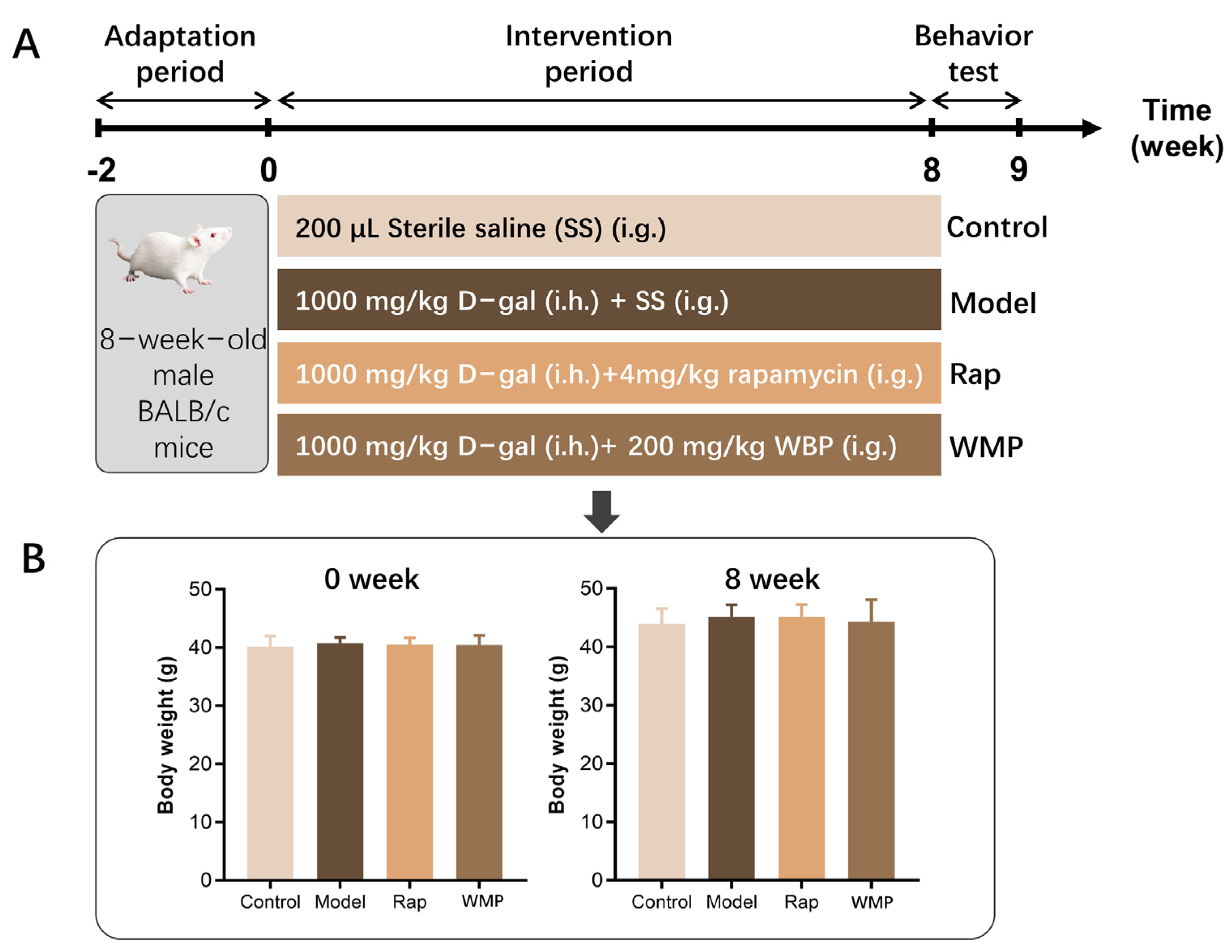
2.2. Animal and Experimental Design
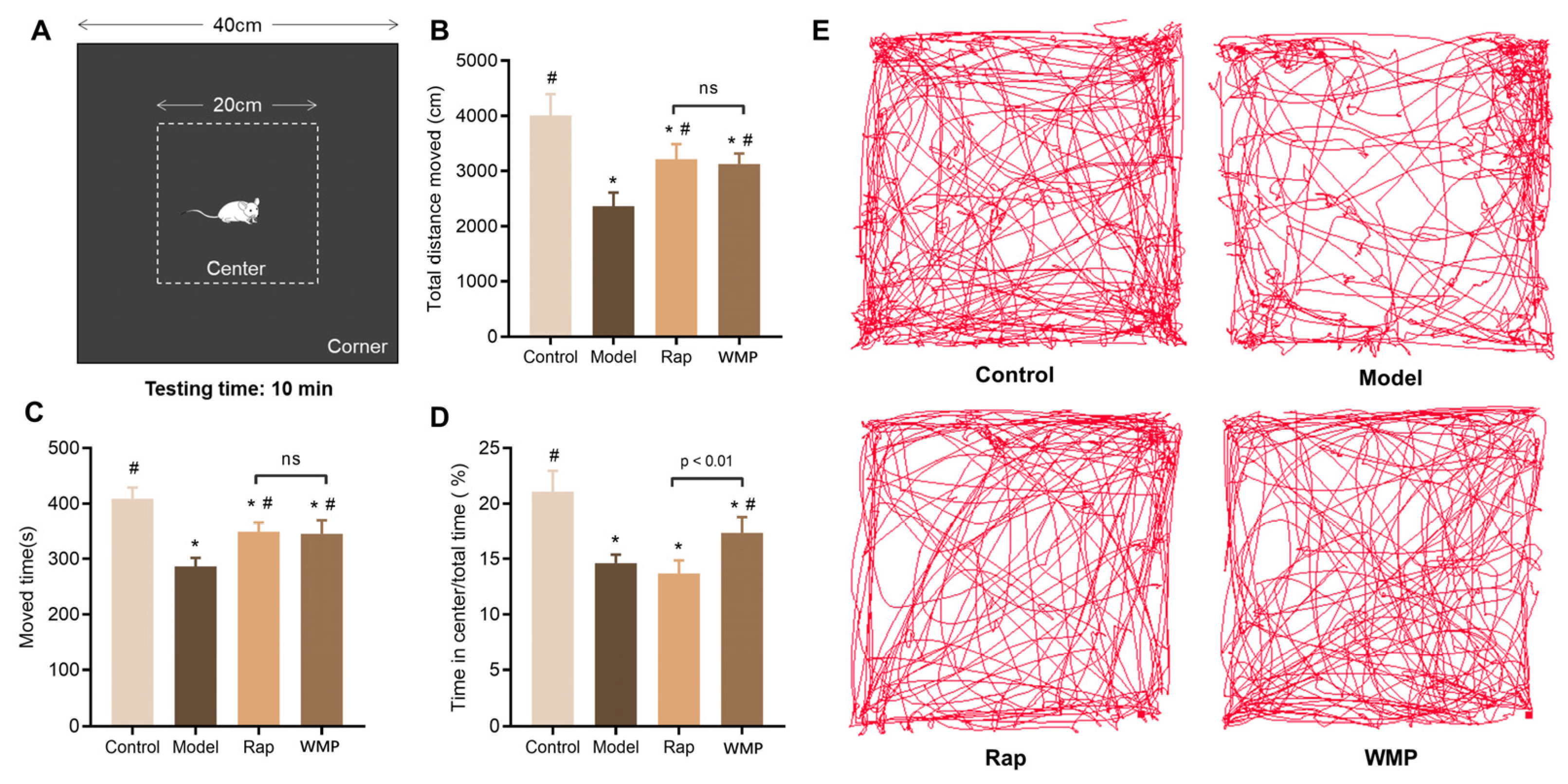
2.3. Behavioral Tests
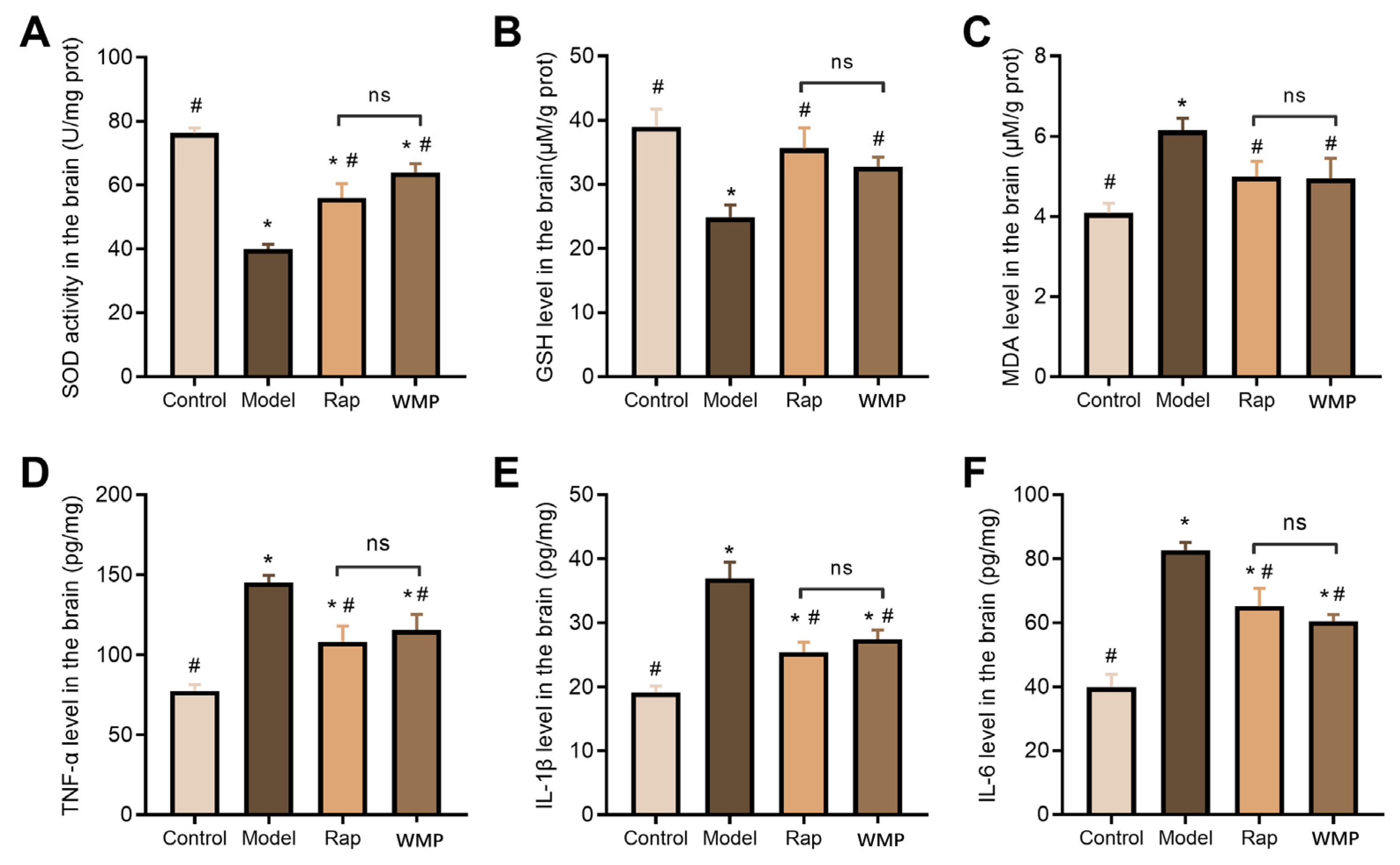
2.4. Determination of Biochemical Markers of Oxidative Stress in the Brain
2.5. Determination of Pro-Inflammatory Cytokine in the Brain
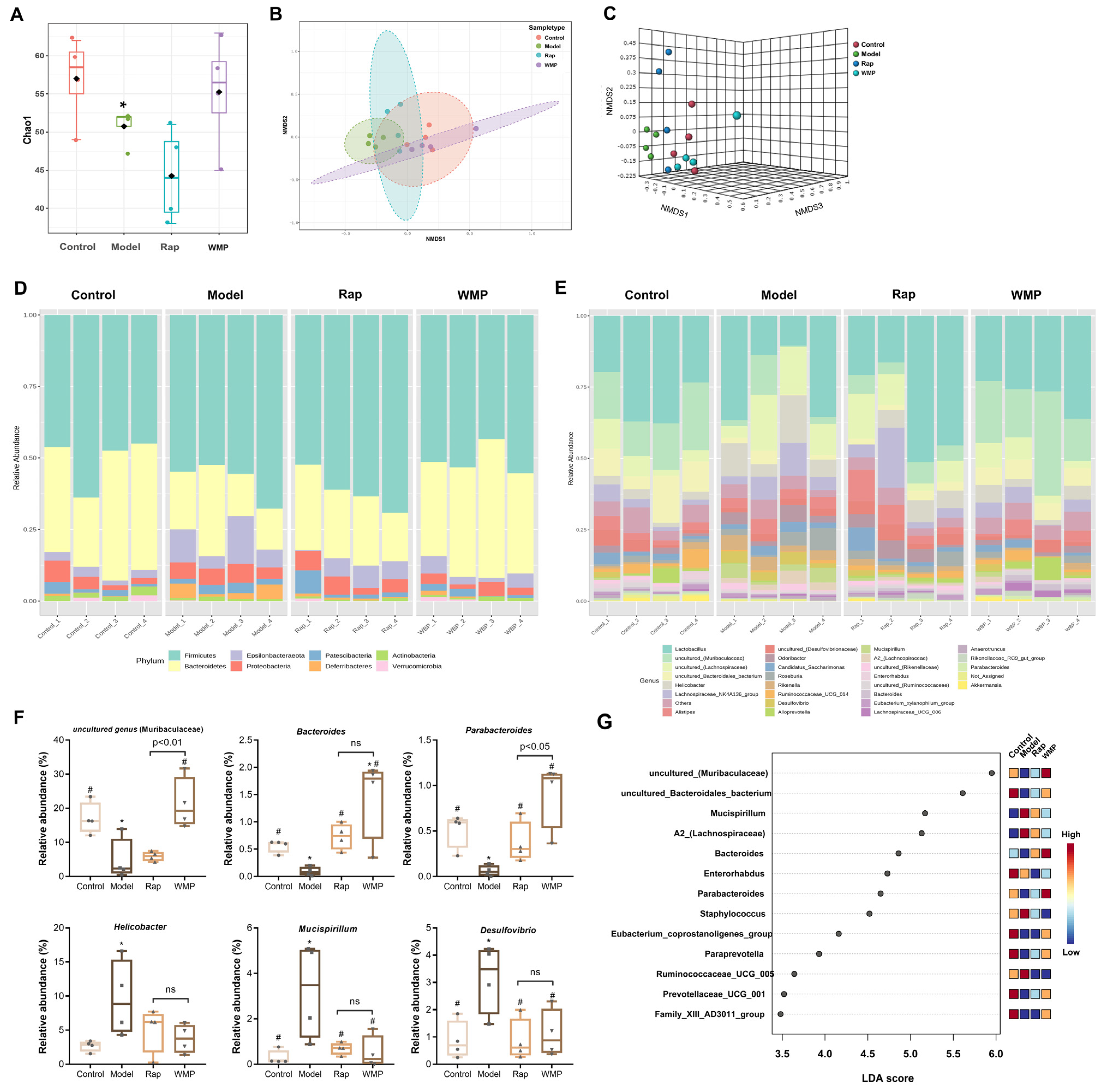
2.6. 16S rRNA Sequencing Analysis of Gut Microbiota
2.7. Determination of SCFAs Level
2.8. Statistical Analysis
3. Results
3.1. The Effects of WMP on the Body Weight of Mice
3.2. The Effects of WMP on Locomotor Activity and Anxiety-Like Behavior of Mice
3.3. The Effects of WMP on the Short-Term Recognition Memory of Mice
3.4. The Effects of WMP on Short-Term Spatial Memory of Mice
3.5. The Effects of WMP on the Antioxidant Index
3.6. The Effects of WMP on the Inflammatory Cytokine
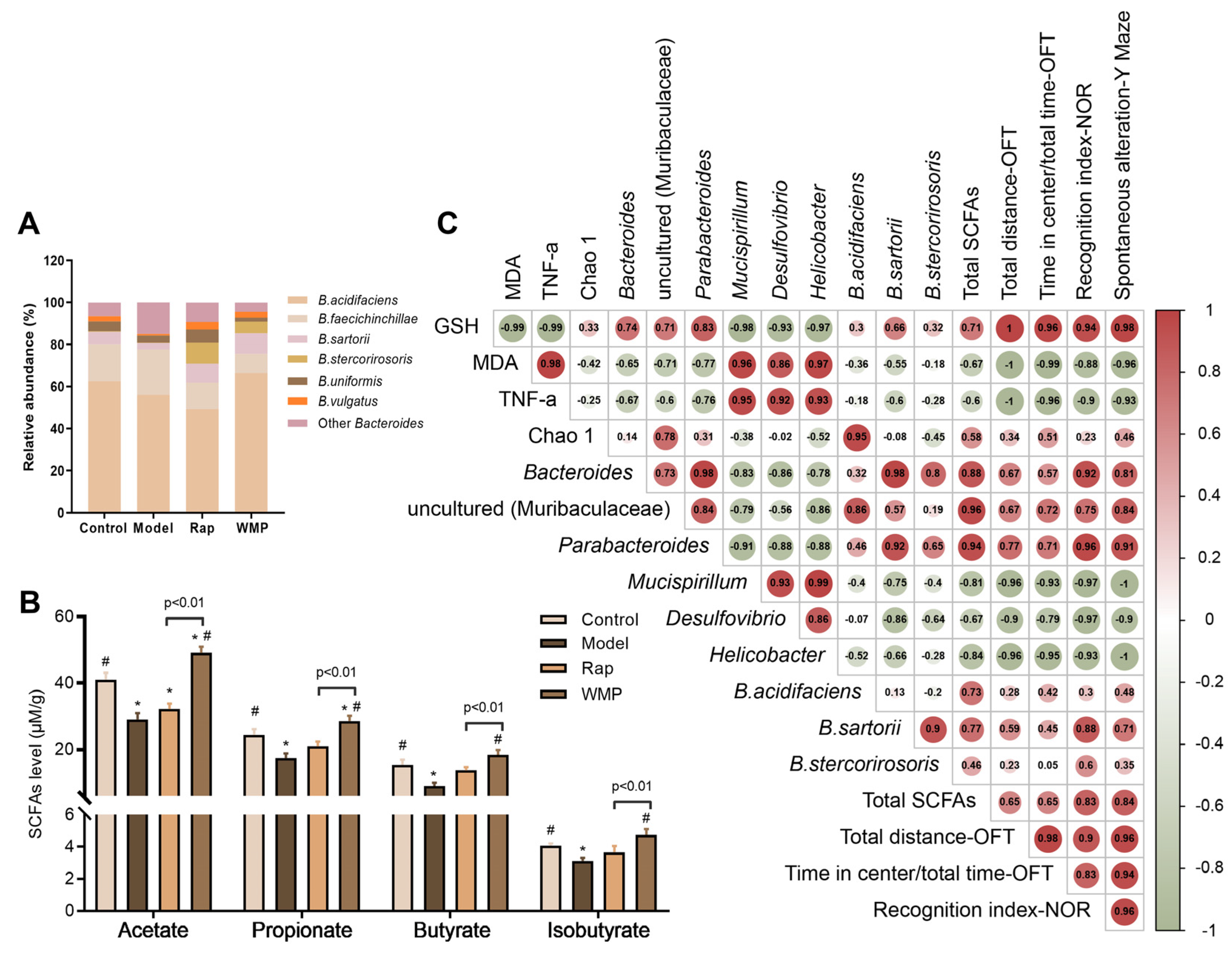
3.7. The Effects of WMP on Gut Microbiota
3.8. The Effects of WMP on SCFA Levels in Mice Faeces
3.9. Spearman Correlation Analysis
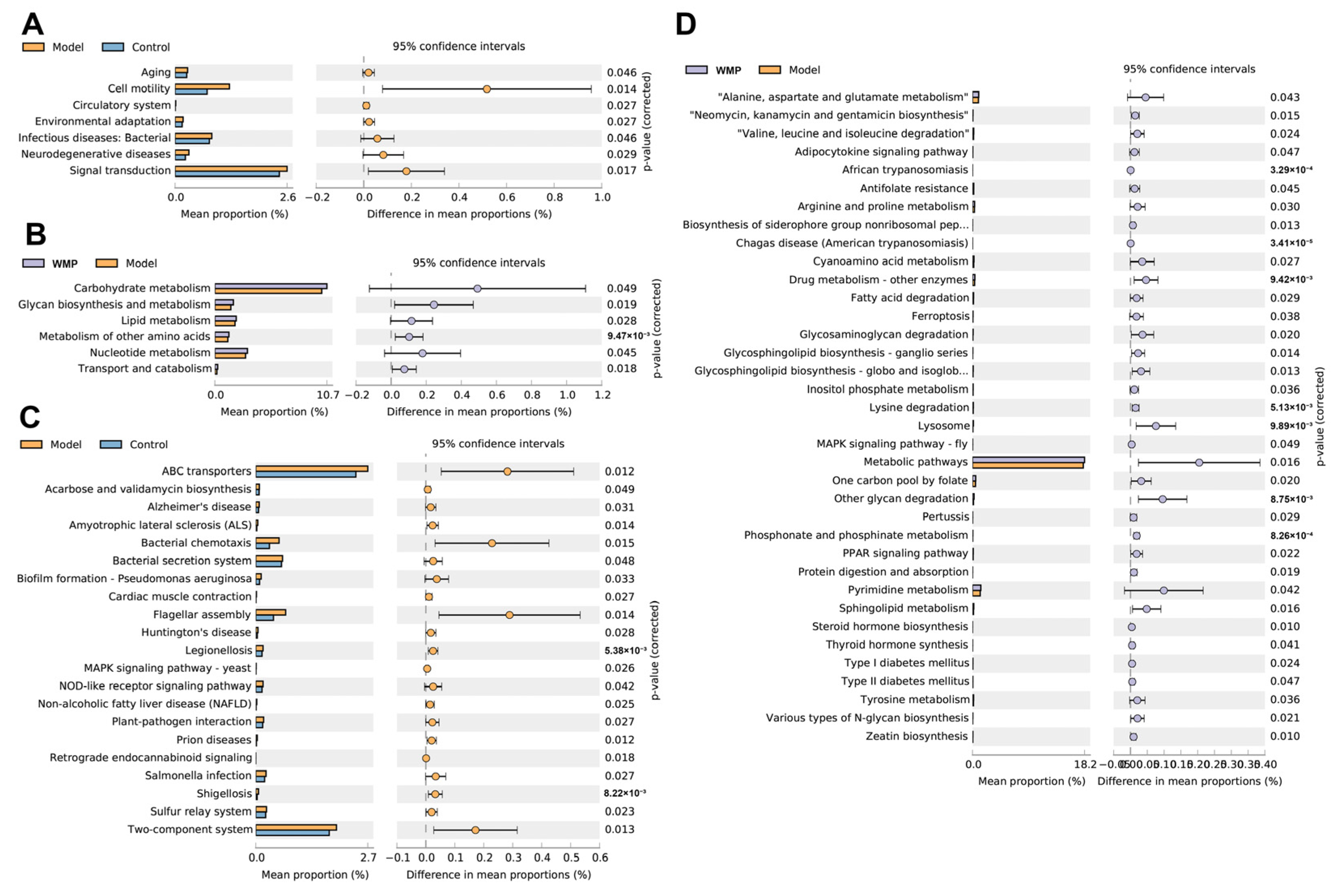
3.10. Effects of WMP on the Functional Profile of the Gut Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hui, D.; Li, J.; Yu, L.; Fan, L. The Road Ahead of Dietary Restriction on Anti-Aging: Focusing on Personalized Nutrition. Crit. Rev. Food Sci. Nutr. 2022, 1–18. [Google Scholar] [CrossRef]
- Most, J.; Mey, J.T.; Redman, L.M. Calorie Restriction and Aging in Humans. Annu. Rev. Nutr. 2020, 40, 105–133. [Google Scholar]
- Boehme, M.; Guzzetta, K.E.; Bastiaanssen, T.F.S.; van de Wouw, M.; Moloney, G.M.; Gual-Grau, A.; Spichak, S.; Olavarría-Ramírez, L.; Fitzgerald, P.; Morillas, E.; et al. Microbiota from Young Mice Counteracts Selective Age-Associated Behavioral Deficits. Nat. Aging 2021, 1, 666–676. [Google Scholar] [CrossRef]
- Honarpisheh, P.; Bryan, R.M.; McCullough, L.D. Aging Microbiota-Gut-Brain Axis in Stroke Risk and Outcome. Circ. Res. 2022, 130, 1112–1144. [Google Scholar] [CrossRef]
- Margolis, K.G.; Cryan, J.F.; Mayer, E.A. The Microbiota-Gut-Brain Axis: From Motility to Mood. Gastroenterology 2021, 160, 1486–1501. [Google Scholar] [CrossRef]
- Fang, X.; Yue, M.; Wei, J.; Wang, Y.; Hong, D.; Wang, B.; Zhou, X.; Chen, T. Evaluation of the Anti-Aging Effects of a Probiotic Combination Isolated from Centenarians in a Samp8 Mouse Model. Front. Immunol. 2021, 12, 792746. [Google Scholar] [CrossRef]
- Shi, H.; Ge, X.; Ma, X.; Zheng, M.; Cui, X.; Pan, W.; Zheng, P.; Yang, X.; Hu, M.; Hu, T.; et al. A Fiber-Deprived Diet Causes Cognitive Impairment and Hippocampal Microglia-Mediated Synaptic Loss through the Gut Microbiota and Metabolites. Microbiome 2021, 9, 223. [Google Scholar] [CrossRef]
- Ghosh, T.S.; Rampelli, S.; Jeffery, I.; Santoro, A.; Neto, M.; Capri, M.; Giampieri, E.; Jennings, A.; Candela, M.; Turroni, S.; et al. Mediterranean diet intervention alters the gut microbiome in older people reducing frailty and improving health status: The NU-AGE 1-year dietary intervention across five European countries. Gut 2020, 1218–1228. [Google Scholar] [CrossRef] [Green Version]
- Ramos, M.; Burgos, N.; Barnard, A.; Evans, G.; Preece, J.; Graz, M.; Ruthes, A.C.; Jiménez-Quero, A.; Martínez-Abad, A.; Vilaplana, F.; et al. Agaricus Bisporus and Its by-Products as a Source of Valuable Extracts and Bioactive Compounds. Food Chem. 2019, 292, 176–187. [Google Scholar] [CrossRef] [Green Version]
- Morin, E.; Kohler, A.; Baker, A.R.; Foulongne-Oriol, M.; Lombard, V.; Nagye, L.G.; Ohm, R.A.; Patyshakuliyeva, A.; Brun, A.; Aerts, A.L.; et al. Genome Sequence of the Button Mushroom Agaricus Bisporus Reveals Mechanisms Governing Adaptation to a Humic-Rich Ecological Niche. Proc. Natl. Acad. Sci. USA 2012, 109, 17501–17506. [Google Scholar] [CrossRef] [Green Version]
- Meng, D.-M.; Zhang, Y.-X.; Yang, R.; Wang, J.; Zhang, X.-H.; Sheng, J.-P.; Wang, J.-P.; Fan, Z.-C. Arginase Participates in the Methyl Jasmonate-Regulated Quality Maintenance of Postharvest Agaricus Bisporus Fruit Bodies. Postharvest Biol. Technol. 2017, 132, 7–14. [Google Scholar] [CrossRef]
- Yang, K.-X.; Xi, Z.-A.; Zhang, Y.-X.; Sheng, J.-P.; Meng, D.-M. Polyamine Biosynthesis and Distribution in Different Tissues of Agaricus Bisporus During Postharvest Storage. Sci. Hortic. 2020, 270, 109457. [Google Scholar] [CrossRef]
- Smiderle, F.R.; Alquini, G.; Tadra-Sfeir, M.Z.; Iacomini, M.; Wichers, H.J.; Van Griensven, L.J. Agaricus Bisporus and Agaricus Brasiliensis (1 -> 6)-Beta-D-Glucans Show Immunostimulatory Activity on Human Thp-1 Derived Macrophages. Carbohydr. Polym. 2013, 94, 91–99. [Google Scholar] [CrossRef]
- Li, S.; Liu, M.; Zhang, C.; Tian, C.; Wang, X.; Song, X.; Jing, H.; Gao, Z.; Ren, Z.; Liu, W.; et al. Purification, in Vitro Antioxidant and in Vivo Anti-Aging Activities of Soluble Polysaccharides by Enzyme-Assisted Extraction from Agaricus Bisporus. Int. J. Biol. Macromol. 2017, 109, 457–466. [Google Scholar] [CrossRef]
- Li, S.; Li, J.; Zhang, J.; Wang, W.; Wang, X.; Jing, H.; Ren, Z.; Gao, Z.; Song, X.; Gong, Z.; et al. The Antioxidative, Antiaging, and Hepatoprotective Effects of Alkali-Extractable Polysaccharides by Agaricus Bisporus. Evid.-Based Complement. Altern. Med. 2017, 2017, 7298683. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Zheng, Y.-L.; Luo, L.; Wu, D.-M.; Sun, D.-X.; Feng, Y.-J. Quercetin Reverses D-Galactose Induced Neurotoxicity in Mouse Brain. Behav. Brain Res. 2006, 171, 251–260. [Google Scholar] [CrossRef]
- Wang, B.; Ntim, M.; Xia, M.; Wang, Y.; Lu, J.-C.; Yang, J.-Y.; Li, S. Long-Term Social Isolation-Induced Autophagy Inhibition and Cell Senescence Aggravate Cognitive Impairment in D(+)Galactose-Treated Male Mice. Front. Aging Neurosci. 2022, 14, 777700. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Yang, X.; Zheng, L.; Wang, Z.; Wu, L.; Jiang, J.; Yang, T.; Ma, L.; Fu, Z. Lactobacillus and Bifidobacterium Improves Physiological Function and Cognitive Ability in Aged Mice by the Regulation of Gut Microbiota. Mol. Nutr. Food Res. 2019, 63, 1900603. [Google Scholar] [CrossRef]
- Gervasi, S.S.; Opiekun, M.; Martin, T.; Beauchamp, G.K.; Kimball, B.A. Sharing an Environment with Sick Conspecifics Alters Odors of Healthy Animals. Sci. Rep. 2018, 8, 14255. [Google Scholar] [CrossRef] [Green Version]
- Ali, T.; Badshah, H.; Kim, T.H.; Kim, M.O. Melatonin Attenuates D-Galactose-Induced Memory Impairment, Neuroinflammation and Neurodegeneration Via Rage/Nf-Kb/Jnk Signaling Pathway in Aging Mouse Model. J. Pineal Res. 2015, 58, 71–85. [Google Scholar] [CrossRef]
- Park, S.; Zhang, T.; Wu, X.; Qiu, J.Y. Ketone Production by Ketogenic Diet and by Intermittent Fasting Has Different Effects on the Gut Microbiota and Disease Progression in an Alzheimer’s Disease Rat Model. J. Clin. Biochem. Nutr. 2020, 67, 188–198. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Shi, L.; Qi, Y.; Wang, Q.; Zhao, J.; Zhang, H.; Wang, G.; Chen, W. Butylated Starch Alleviates Polycystic Ovary Syndrome by Stimulating the Secretion of Peptide Tyrosine-Tyrosine and Regulating Faecal Microbiota. Carbohydr. Polym. 2022, 287, 119304. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Feng, S.; Xiao, Y.; Pan, M.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. A New Illumina Miseq High-Throughput Sequencing-Based Method for Evaluating the Composition of the Bacteroides Community in the Intestine Using the Rpsd Gene Sequence. Microb. Biotechnol. 2020, 14, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Lu, W.; Wang, L.; Pan, M.; Zhang, H.; Zhao, J.; Chen, W. Assessment of Bifidobacterium Species Using Groel Gene on the Basis of Illumina Miseq High-Throughput Sequencing. Genes 2017, 8, 336. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Yu, L.; Zhai, Q.; Zhao, R.; Zhao, J.; Zhang, H.; Chen, W.; Tian, F. In Vitro Fermentation of Heparin by the Human Gut Microbiota: Changes in the Microbiota Community and Metabolic Functions. Food Chem. 2023, 406, 135010. [Google Scholar] [CrossRef]
- Muszyńska, B.; Grzywacz-Kisielewska, A.; Kała, K.; Gdula-Argasińska, J. Anti-Inflammatory Properties of Edible Mushrooms: A Review. Food Chem. 2018, 243, 373–381. [Google Scholar] [CrossRef]
- Harikrishnan, R.; Devi, G.; Van Doan, H.; Balasundaram, C.; Thamizharasan, S.; Hoseinifar, S.H.; Abdel-Tawwab, M. Effect of diet enriched with Agaricus bisporus polysaccharides (ABPs) on antioxidant property, innate-adaptive immune response and pro-anti inflammatory genes expression in Ctenopharyngodon idella against Aeromonas hydrophila. Fish Shellfish. Immunol. 2021, 114, 238–252. [Google Scholar] [CrossRef]
- Khan, I.; Huang, G.; Li, X.A.; Liao, W.; Leong, W.K.; Xia, W.; Bian, X.; Wu, J.; Hsiao, W.L.W. Mushroom Polysaccharides and Jiaogulan Saponins Exert Cancer Preventive Effects by Shaping the Gut Microbiota and Microenvironment in Apc(Min/+) Mice. Pharmacol. Res. 2019, 148, 104448. [Google Scholar] [CrossRef]
- Liang, X.; Fu, Y.; Cao, W.; Wang, Z.; Zhang, K.; Jiang, Z.; Jia, X.; Liu, C.; Lin, H.; Zhong, H.; et al. Gut Microbiome, Cognitive Function and Brain Structure: A Multi-Omics Integration Analysis. Transl. Neurodegener. 2022, 11, 49. [Google Scholar] [CrossRef]
- Shwe, T.; Pratchayasakul, W.; Chattipakorn, N.; Chattipakorn, S.C. Role of D-Galactose-Induced Brain Aging and Its Potential Used for Therapeutic Interventions. Exp. Gerontol. 2018, 101, 13–36. [Google Scholar] [CrossRef]
- Kou, X.J.; Liu, X.; Chen, X.; Li, J.; Yang, X.; Fan, J.; Yang, Y.; Chen, N. Ampelopsin Attenuates Brain Aging of D-Gal- Induced Rats through Mir-34a-Mediated Sirt1/Mtor Signal Pathway. Oncotarget 2016, 7, 74484–74495. [Google Scholar] [CrossRef] [PubMed]
- Ghaisas, S.; Maher, J.; Kanthasamy, A. Gut Microbiome in Health and Disease: Linking the Microbiome-Gut-Brain Axis and Environmental Factors in the Pathogenesis of Systemic and Neurodegenerative Diseases. Pharmacol. Ther. 2016, 158, 52–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bharwani, A.; Mian, M.F.; Surette, M.G.; Bienenstock, J.; Forsythe, P. Oral Treatment with Lactobacillus Rhamnosus Attenuates Behavioural Deficits and Immune Changes in Chronic Social Stress. BMC Med. 2017, 15, 7. [Google Scholar] [CrossRef] [Green Version]
- Tillisch, K.; Mayer, E.A.; Gupta, A.; Gill, Z.; Brazeilles, R.; Le Nevé, B.; Vlieg, J.E.V.H.; Guyonnet, D.; Derrien, M.; Labus, J.S. Brain Structure and Response to Emotional Stimuli as Related to Gut Microbial Profiles in Healthy Women. Psychosom. Med. 2017, 79, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Strandwitz, P.; Kim, K.H.; Terekhova, D.; Liu, J.K.; Sharma, A.; Levering, J.; McDonald, D.; Dietrich, D.; Ramadhar, T.R.; Lekbua, A.; et al. Gaba-Modulating Bacteria of the Human Gut Microbiota. Nat. Microbiol. 2019, 4, 396–403. [Google Scholar] [CrossRef]
- Wang, K.; Liao, M.; Zhou, N.; Bao, L.; Ma, K.; Zheng, Z.; Wang, Y.; Liu, C.; Wang, W.; Wang, J.; et al. Parabacteroides Distasonis Alleviates Obesity and Metabolic Dysfunctions Via Production of Succinate and Secondary Bile Acids. Cell Rep. 2019, 26, 222–235. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Zhou, M.; Wang, J.; Yao, J.; Yu, J.; Liu, W.; Wu, L.; Wang, J.; Gao, R. Involvement of the Microbiota-Gut-Brain Axis in Chronic Restraint Stress: Disturbances of the Kynurenine Metabolic Pathway in Both the Gut and Brain. Gut Microbes 2021, 13, 1–16. [Google Scholar] [CrossRef]
- Olson, C.A.; Vuong, H.; Yano, J.; Liang, Q.; Nusbaum, D.; Hsiao, E.Y. The Gut Microbiota Mediates the Anti-Seizure Effects of the Ketogenic Diet. Cell 2018, 173, 1728–1741. [Google Scholar] [CrossRef] [Green Version]
- David, L.A.; Maurice, C.; Carmody, R.; Gootenberg, D.; Button, J.; Wolfe, B.; Ling, A.; Devlin, A.; Varma, Y.; Fischbach, M.; et al. Diet Rapidly and Reproducibly Alters the Human Gut Microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Newman, J.C.; Covarrubias, A.J.; Zhao, M.; Yu, X.; Gut, P.; Ng, C.-P.; Huang, Y.; Haldar, S.; Verdin, E. Ketogenic Diet Reduces Midlife Mortality and Improves Memory in Aging Mice. Cell Metab. 2017, 26, 547–557. [Google Scholar] [CrossRef] [Green Version]
- Roberts, M.N.; Wallace, M.A.; Tomilov, A.A.; Zhou, Z.; Marcotte, G.R.; Tran, D.; Perez, G.; Gutierrez-Casado, E.; Koike, S.; Knotts, T.A.; et al. A Ketogenic Diet Extends Longevity and Healthspan in Adult Mice. Cell Metab. 2017, 27, 1156. [Google Scholar] [CrossRef] [PubMed]
- Yudkoff, M.; Daikhin, Y.; Nissim, I.; Lazarow, A.; Nissim, I. Ketogenic Diet, Amino Acid Metabolism, and Seizure Control. J. Neurosci. Res. 2001, 66, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.M.; Mancano, G.; Kashofer, K.; Frohlich, E.; Matak, A.; Mayerhofer, R.; Reichmann, F.; Olivares, M.; Neyrinck, A.; Delzenne, N.; et al. High-Fat Diet Induces Depression-Like Behaviour in Mice Associated with Changes in Microbiome, Neuropeptide Y, and Brain Metabolome. Nutr. Neurosci. 2019, 12, 877–893. [Google Scholar] [CrossRef] [Green Version]
- Marques, F.Z.; Nelson, E.; Chu, P.-Y.; Horlock, D.; Fiedler, A.; Ziemann, M.; Tan, J.K.; Kuruppu, S.; Rajapakse, N.W.; El-Osta, A.; et al. High-Fiber Diet and Acetate Supplementation Change the Gut Microbiota and Prevent the Development of Hypertension and Heart Failure in Hypertensive Mice. Circulation 2017, 135, 964–977. [Google Scholar] [CrossRef]
- Then, C.K.; Paillas, S.; Wang, X.; Hampson, A.; Kiltie, A.E. Association of Bacteroides Acidifaciens Relative Abundance with High-Fibre Diet-Associated Radiosensitisation. BMC Biol. 2020, 18, 102. [Google Scholar] [CrossRef]
- Wei, B.; Zhang, B.; Du, A.-Q.; Zhou, Z.-Y.; Lu, D.-Z.; Zhu, Z.-H.; Ke, S.-Z.; Wang, S.-J.; Yu, Y.-L.; Chen, J.-W.; et al. Saccharina Japonica Fucan Suppresses High Fat Diet-Induced Obesity and Enriches Fucoidan-Degrading Gut Bacteria. Carbohydr. Polym. 2022, 290, 119411. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Lee, Y.; Kim, Y.; Lee, S.; Ryu, S.; Fukuda, S.; Hase, K.; Yang, C.; Lim, H.; Kim, M.; et al. Gut Commensal Bacteroides Acidifaciens Prevents Obesity and Improves Insulin Sensitivity in Mice. Mucosal Immunol. 2017, 10, 104–116. [Google Scholar] [CrossRef] [Green Version]
- Aoki, R.; Onuki, M.; Hattori, K.; Ito, M.; Yamada, T.; Kamikado, K.; Kim, Y.-G.; Nakamoto, N.; Kimura, I.; Clarke, J.M.; et al. Commensal Microbe-Derived Acetate Suppresses Nafld/Nash Development Via Hepatic Ffar2 Signalling in Mice. Microbiome 2021, 9, 188. [Google Scholar] [CrossRef]
- Kugadas, A.; Wright, Q.; Geddes-McAlister, J.; Gadjeva, M. Role of Microbiota in Strengthening Ocular Mucosal Barrier Function through Secretory Iga. Investig. Opthalmology Vis. Sci. 2017, 58, 4593–4600. [Google Scholar] [CrossRef]
- Wang, H.; Wang, Q.; Yang, C.; Guo, M.; Cui, X.; Jing, Z.; Liu, Y.; Qiao, W.; Qi, H.; Zhang, H.; et al. Bacteroides Acidifaciens in the Gut Plays a Protective Role against Cd95-Mediated Liver Injury. Gut Microbes 2022, 14, 2027853. [Google Scholar] [CrossRef]
- Lu, X.; Jing, Y.; Zhang, N.; Cao, Y. Eurotium Cristatum, a Probiotic Fungus from Fuzhuan Brick Tea, and Its Polysaccharides Ameliorated Dss-Induced Ulcerative Colitis in Mice by Modulating the Gut Microbiota. J. Agric. Food Chem. 2022, 70, 2957–2967. [Google Scholar] [CrossRef]
- Loy, A.; Pfann, C.; Steinberger, M.; Hanson, B.; Herp, S.; Brugiroux, S.; Neto, J.G.; Boekschoten, M.; Schwab, C.; Urich, T.; et al. Lifestyle and Horizontal Gene Transfer-Mediated Evolution of Mucispirillum Schaedleri, a Core Member of the Murine Gut Microbiota. Msystems 2017, 2, e00171-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Coker, O.O.; Chu, E.S.; Fu, K.; Lau, H.C.H.; Wang, Y.-X.; Chan, A.W.H.; Wei, H.; Yang, X.; Sung, J.J.Y.; et al. Dietary Cholesterol Drives Fatty Liver-Associated Liver Cancer by Modulating Gut Microbiota and Metabolites. Gut 2020, 70, 761–774. [Google Scholar] [CrossRef] [PubMed]
- El Aidy, S.; Ramsteijn, A.; Dini-Andreote, F.; van Eijk, R.; Houwing, D.; Salles, J.; Olivier, J.D.A. Serotonin Transporter Genotype Modulates the Gut Microbiota Composition in Young Rats, an Effect Augmented by Early Life Stress. Front. Cell. Neurosci. 2017, 11, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caruso, R.; Mathes, T.; Martens, E.C.; Kamada, N.; Nusrat, A.; Inohara, N.; Núñez, G. A Specific Gene-Microbe Interaction Drives the Development of Crohn’s Disease-Like Colitis in Mice. Sci. Immunol. 2019, 4, eaaw4341. [Google Scholar] [CrossRef]
- Lin, C.H.; Chen, C.; Chiang, H.; Liou, J.; Chang, C.; Lu, T.; Chuang, E.; Tai, Y.; Cheng, C.; Lin, H.; et al. Altered Gut Microbiota and Inflammatory Cytokine Responses in Patients with Parkinson’s Disease. J. Neuroinflammation 2019, 16, 129. [Google Scholar] [CrossRef]
- Gao, L.; Li, J.; Zhou, Y.; Huang, X.; Qin, X.; Du, G. Effects of Baicalein on Cortical Proinflammatory Cytokines and the Intestinal Microbiome in Senescence Accelerated Mouse Prone 8. ACS Chem. Neurosci. 2018, 9, 1714–1724. [Google Scholar] [CrossRef]
- Couturier-Maillard, A.; Secher, T.; Rehman, A.; Normand, S.; De Arcangelis, A.; Häsler, R.; Huot, L.; Grandjean, T.; Bressenot, A.; Delanoye-Crespin, A.; et al. Nod2-Mediated Dysbiosis Predisposes Mice to Transmissible Colitis and Colorectal Cancer. J. Clin. Investig. 2013, 123, 700–711. [Google Scholar] [CrossRef] [Green Version]
- Ahn, E.H.; Liu, X.; Alam, A.M.; Kang, S.S.; Ye, K. Helicobacter Hepaticus Augmentation Triggers Dopaminergic Degeneration and Motor Disorders in Mice with Parkinson’s Disease. Mol. Psychiatry 2022. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, H.; Li, J.; Fan, L. Agaricus bisporus Polysaccharides Ameliorates Behavioural Deficits in D-Galactose-Induced Aging Mice: Mediated by Gut Microbiota. Foods 2023, 12, 424. https://doi.org/10.3390/foods12020424
Duan H, Li J, Fan L. Agaricus bisporus Polysaccharides Ameliorates Behavioural Deficits in D-Galactose-Induced Aging Mice: Mediated by Gut Microbiota. Foods. 2023; 12(2):424. https://doi.org/10.3390/foods12020424
Chicago/Turabian StyleDuan, Hui, Jinwei Li, and Liuping Fan. 2023. "Agaricus bisporus Polysaccharides Ameliorates Behavioural Deficits in D-Galactose-Induced Aging Mice: Mediated by Gut Microbiota" Foods 12, no. 2: 424. https://doi.org/10.3390/foods12020424