

The Impact of Ellagitannins and Their Metabolites through Gut Microbiome on the Gut Health and Brain Wellness within the Gut–Brain Axis


Abstract
:1. Introduction
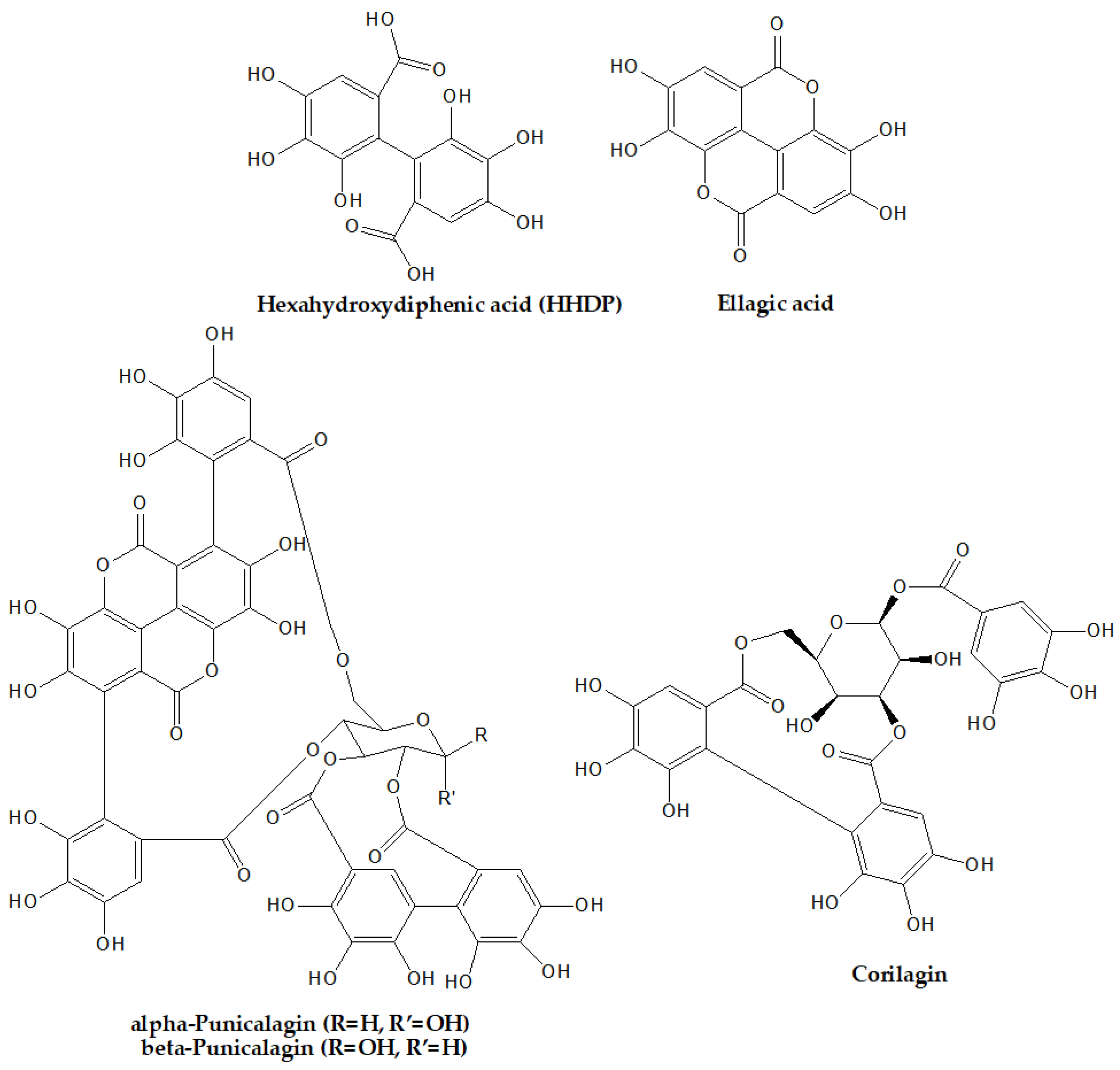
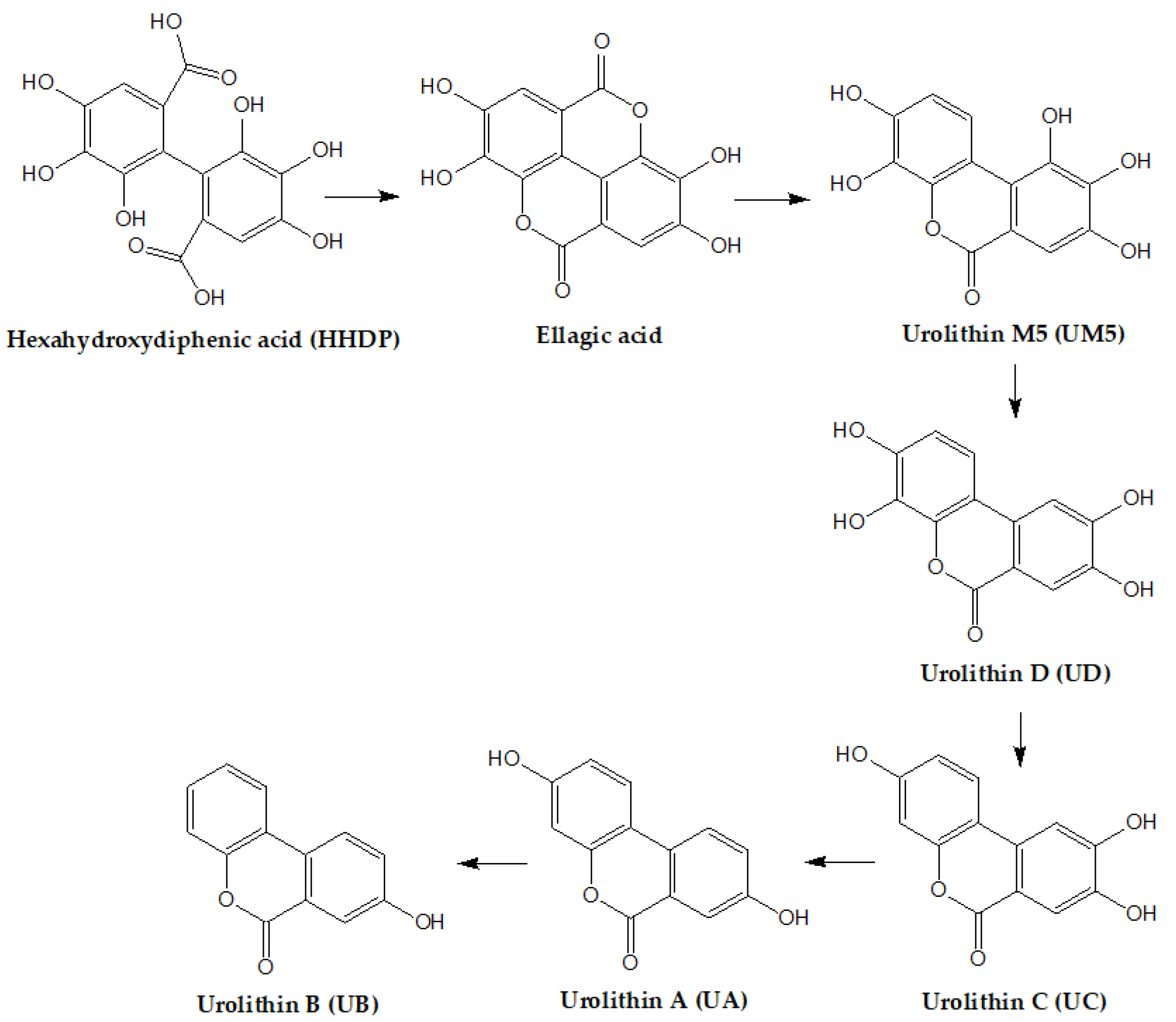
2. Chemistry of Ellagitannins
3. Sources of Ellagitannins
4. Metabolism and Bioavailability of ETs and EA
5. Biological Activities and Mechanisms of Action of ETs, EA, and Urolithins
5.1. Antioxidant Activity of ETs, EA, and Urolithins
5.2. In Vitro Studies Attesting to the Beneficial Influence of ETs and Their Metabolites on the Gut–Brain Axis
5.2.1. Antioxidant and Anti-Inflammatory Effects
5.2.2. Neuroprotective Effects
5.3. Other Mechanisms of Action of the Urolithins
6. In Vivo Preclinical and Clinical Evidence for Beneficial Effects of ETs, EA, and Urolithins on Gut–Brain Axis
6.1. In Vivo Studies
6.1.1. In Vivo Studies Performed at the Intestinal Level
6.1.2. In Vivo Studies Performed at the Cerebral Level
6.2. Clinical Studies
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Ramlagan, P.; Labib, R.M.; Farag, M.A.; Neergheen, V.S. Advances towards the Analysis, Metabolism and Health Benefits of Punicalagin, One of the Largest Ellagitannin from Plants, with Future Perspectives. Phytomed. Plus 2022, 2, 100313. [Google Scholar] [CrossRef]
- Moorthy, M.; Wie, C.C.; Mariño, E.; Palanisamy, U.D. The Prebiotic Potential of Geraniin and Geraniin-Enriched Extract against High-Fat-Diet-Induced Metabolic Syndrome in Sprague Dawley Rats. Antioxidants 2022, 11, 632. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Bastida, J.A.; Ávila-Gálvez, M.Á.; Espín, J.C.; González-Sarrías, A. Evidence for Health Properties of Pomegranate Juices and Extracts beyond Nutrition: A Critical Systematic Review of Human Studies. Trends Food Sci. Technol. 2021, 114, 410–423. [Google Scholar] [CrossRef]
- Fizeșan, I.; Rusu, M.E.; Georgiu, C.; Pop, A.; Ștefan, M.G.; Muntean, D.M.; Mirel, S.; Vostinaru, O.; Kiss, B.; Popa, D.S. Antitussive, Antioxidant, and Anti-Inflammatory Effects of a Walnut (Juglans Regia L.) Septum Extract Rich in Bioactive Compounds. Antioxidants 2021, 10, 119. [Google Scholar] [CrossRef]
- Banc, R.; Popa, D.S.; Cozma-Petruţ, A.; Filip, L.; Kiss, B.; Fărcaş, A.; Nagy, A.; Miere, D.; Loghin, F. Protective Effects of Wine Polyphenols on Oxidative Stress and Hepatotoxicity Induced by Acrylamide in Rats. Antioxidants 2022, 11, 1347. [Google Scholar] [CrossRef] [PubMed]
- Banc, R.; Loghin, F.; Miere, D.; Ranga, F.; Socaciu, C. Phenolic Composition and Antioxidant Activity of Red, Rosé and White Wines Originating from Romanian Grape Cultivars. Not. Bot. Horti Agrobot. Cluj-Napoca 2020, 48, 716–734. [Google Scholar] [CrossRef]
- Vrânceanu, M.; Galimberti, D.; Banc, R.; Dragoş, O.; Cozma-Petruţ, A.; Hegheş, S.-C.; Voştinaru, O.; Cuciureanu, M.; Stroia, C.M.; Miere, D.; et al. The Anticancer Potential of Plant-Derived Nutraceuticals via the Modulation of Gene Expression. Plants 2022, 11, 2524. [Google Scholar] [CrossRef] [PubMed]
- Banc, R.; Socaciu, C.; Miere, D.; Filip, L.; Cozma, A.; Stanciu, O.; Loghin, F. Benefits of Wine Polyphenols on Human Health: A Review. Bull. UASVM Food Sci. Technol. 2014, 71, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Das, A.K.; Islam, M.N.; Faruk, M.O.; Ashaduzzaman, M.; Dungani, R. Review on Tannins: Extraction Processes, Applications and Possibilities. S. Afr. J. Bot. 2020, 135, 58–70. [Google Scholar] [CrossRef]
- de Jesus, N.Z.T.; de Souza Falcão, H.; Gomes, I.F.; de Almeida Leite, T.J.; de Morais Lima, G.R.; Barbosa-Filho, J.M.; Tavares, J.F.; da Silva, M.S.; de Athayde-Filho, P.F.; Batista, L.M. Tannins, Peptic Ulcers and Related Mechanisms. Int. J. Mol. Sci. 2012, 13, 3203–3228. [Google Scholar] [CrossRef]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouységu, L. Plant Polyphenols: Chemical Properties, Biological Activities, and Synthesis. Angew. Chem. Int. Ed. 2011, 50, 586–621. [Google Scholar] [CrossRef] [PubMed]
- Aguilera-Carbo, A.; Augur, C.; Prado-Barragan, L.A.; Favela-Torres, E.; Aguilar, C.N. Microbial Production of Ellagic Acid and Biodegradation of Ellagitannins. Appl. Microbiol. Biotechnol. 2008, 78, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Bule, M.; Khan, F.; Nisar, M.F.; Niaz, K. Tannins (Hydrolysable Tannins, Condensed Tannins, Phlorotannins, Flavono-Ellagitannins). In Recent Advances in Natural Products Analysis; Silva, A.S., Nabavi, S.F., Saeedi, M., Nabavi, S.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Smeriglio, A.; Barreca, D.; Bellocco, E.; Trombetta, D. Proanthocyanidins and Hydrolysable Tannins: Occurrence, Dietary Intake and Pharmacological Effects. Br. J. Pharmacol. 2017, 174, 1244–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larrosa, M.; García-Conesa, M.T.; Espín, J.C.; Tomás-Barberán, F.A. Ellagitannins, Ellagic Acid and Vascular Health. Mol. Asp. Med. 2010, 31, 513–539. [Google Scholar] [CrossRef]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic Composition and Antioxidant Potential of Grain Legume Seeds: A Review. Food Res. Int. 2017, 101, 1–16. [Google Scholar] [CrossRef]
- González-Centeno, M.R.; Chira, K.; Teissedre, P.L. Ellagitannin Content, Volatile Composition and Sensory Profile of Wines from Different Countries Matured in Oak Barrels Subjected to Different Toasting Methods. Food Chem. 2016, 210, 500–511. [Google Scholar] [CrossRef]
- Gadrat, M.; Capello, Y.; Emo, C.; Lavergne, J.; Quideau, S.; Jourdes, M.; Teissèdre, P.L.; Chira, K. Identification, Quantitation and Sensory Contribution of New C-Glucosidic Ellagitannin-Derived Spirit Compounds. Food Chem. 2022, 384, 132307. [Google Scholar] [CrossRef]
- Chen, S.; Li, M.; Zheng, G.; Wang, T.; Lin, J.; Wang, S.; Wang, X.; Chao, Q.; Cao, S.; Yang, Z.; et al. Metabolite Profiling of 14 Wuyi Rock Tea Cultivars Using UPLC-QTOF MS and UPLC-QqQ MS Combined with Chemometrics. Molecules 2018, 23, 104. [Google Scholar] [CrossRef] [Green Version]
- Tomás-Barberán, F.A.; González-Sarrías, A.; García-Villalba, R.; Núñez-Sánchez, M.A.; Selma, M.V.; García-Conesa, M.T.; Espín, J.C. Urolithins, the Rescue of “Old” Metabolites to Understand a “New” Concept: Metabotypes as a Nexus among Phenolic Metabolism, Microbiota Dysbiosis, and Host Health Status. Mol. Nutr. Food Res. 2017, 61, 1500901. [Google Scholar] [CrossRef]
- Lopez-Corona, A.V.; Valencia-Espinosa, I.; González-Sánchez, F.A.; Sánchez-López, A.L.; Garcia-Amezquita, L.E.; Garcia-Varela, R. Antioxidant, Anti-Inflammatory and Cytotoxic Activity of Phenolic Compound Family Extracted from Raspberries (Rubus Idaeus): A General Review. Antioxidants 2022, 11, 1192. [Google Scholar] [CrossRef]
- Garrido, J.; Borges, F. Wine and Grape Polyphenols—A Chemical Perspective. Food Res. Int. 2013, 54, 1844–1858. [Google Scholar] [CrossRef] [Green Version]
- García-Villalba, R.; Tomás-Barberán, F.A.; Iglesias-Aguirre, C.E.; Giménez-Bastida, J.A.; González-Sarrías, A.; Selma, M.V.; Espín, J.C. Ellagitannins, Urolithins, and Neuroprotection: Human Evidence and the Possible Link to the Gut Microbiota. Mol. Asp. Med. 2022, 101109. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, D.; Andreux, P.A.; Valdés, P.; Singh, A.; Rinsch, C.; Auwerx, J. Impact of the Natural Compound Urolithin A on Health, Disease, and Aging. Trends Mol. Med. 2021, 27, 687–699. [Google Scholar] [CrossRef] [PubMed]
- García-Villalba, R.; Giménez-Bastida, J.A.; Cortés-Martín, A.; Ávila-Gálvez, M.Á.; Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C.; González-Sarrías, A. Urolithins: A Comprehensive Update on Their Metabolism, Bioactivity, and Associated Gut Microbiota. Mol. Nutr. Food Res. 2022, 66, 2101019. [Google Scholar] [CrossRef] [PubMed]
- Piechowiak, T.; Grzelak-Błaszczyk, K.; Sójka, M.; Balawejder, M. Changes in Phenolic Compounds Profile and Glutathione Status in Raspberry Fruit during Storage in Ozone-Enriched Atmosphere. Postharvest Biol. Technol. 2020, 168, 111277. [Google Scholar] [CrossRef]
- Tozzi, F.; Núñez-Gómez, D.; Legua, P.; del Bubba, M.; Giordani, E.; Melgarejo, P. Qualitative and Varietal Characterization of Pomegranate Peel: High-Value Co-Product or Waste of Production? Sci. Hortic. 2022, 291, 110601. [Google Scholar] [CrossRef]
- Rojas-Garbanzo, C.; Winter, J.; Montero, M.L.; Zimmermann, B.F.; Schieber, A. Characterization of Phytochemicals in Costa Rican Guava (Psidium Friedrichsthalianum-Nied.) Fruit and Stability of Main Compounds during Juice Processing—(U)HPLC-DAD-ESI-TQD-MSn. J. Food Compos. Anal. 2019, 75, 26–42. [Google Scholar] [CrossRef]
- Karlińska, E.; Masny, A.; Cieślak, M.; Macierzyński, J.; Pecio, Ł.; Stochmal, A.; Kosmala, M. Ellagitannins in Roots, Leaves, and Fruits of Strawberry (Fragaria × Ananassa Duch.) Vary with Developmental Stage and Cultivar. Sci. Hortic. 2021, 275, 109665. [Google Scholar] [CrossRef]
- van de Velde, F.; Pirovani, M.E.; Drago, S.R. Bioaccessibility Analysis of Anthocyanins and Ellagitannins from Blackberry at Simulated Gastrointestinal and Colonic Levels. J. Food Compos. Anal. 2018, 72, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Medic, A.; Jakopic, J.; Hudina, M.; Solar, A.; Veberic, R. Identification and Quantification of the Major Phenolic Constituents in Juglans Regia L. Peeled Kernels and Pellicles, Using HPLC–MS/MS. Food Chem. 2021, 352, 129404. [Google Scholar] [CrossRef]
- Aires, A.; Carvalho, R.; Saavedra, M.J. Valorization of Solid Wastes from Chestnut Industry Processing: Extraction and Optimization of Polyphenols, Tannins and Ellagitannins and Its Potential for Adhesives, Cosmetic and Pharmaceutical Industry. Waste Manag. 2016, 48, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M. Ellagitannins, Ellagic Acid and Their Derived Metabolites: A Review about Source, Metabolism, Functions and Health. Food Res. Int. 2011, 44, 1150–1160. [Google Scholar] [CrossRef]
- Roberts, K.M.; Grainger, E.M.; Thomas-Ahner, J.M.; Hinton, A.; Gu, J.; Riedl, K.M.; Vodovotz, Y.; Abaza, R.; Schwartz, S.J.; Clinton, S.K. Application of a Low Polyphenol or Low Ellagitannin Dietary Intervention and Its Impact on Ellagitannin Metabolism in Men. Mol. Nutr. Food Res. 2017, 61, 1600224. [Google Scholar] [CrossRef] [PubMed]
- Villalba, K.J.O.; Barka, F.V.; Pasos, C.V.; Rodríguez, P.E. Food Ellagitannins: Structure, Metabolomic Fate, and Biological Properties. In Tannins—Structural Properties, Biological Properties and Current Knowledge; Aires, A., Ed.; IntechOpen: London, UK, 2020. [Google Scholar]
- Cheng, H.S.; Goh, B.H.; Phang, S.C.W.; Amanullah, M.M.; Ton, S.H.; Palanisamy, U.D.; Kadir, K.A.; Tan, J.B.L. Pleiotropic Ameliorative Effects of Ellagitannin Geraniin against Metabolic Syndrome Induced by High-Fat Diet in Rats. Nutrition 2020, 79–80, 110973. [Google Scholar] [CrossRef] [PubMed]
- Alfei, S.; Marengo, B.; Zuccari, G. Oxidative Stress, Antioxidant Capabilities, and Bioavailability: Ellagic Acid or Urolithins? Antioxidants 2020, 9, 707. [Google Scholar] [CrossRef]
- Biswas, S.K. Does the Interdependence between Oxidative Stress and Inflammation Explain the Antioxidant Paradox? Oxidative Med. Cell. Longev. 2016, 2016, 5698931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ríos, J.L.; Giner, R.M.; Marín, M.; Recio, M.C. A Pharmacological Update of Ellagic Acid. Planta Med. 2018, 84, 1068–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marín, M.; Giner, R.M.; Ríos, J.L.; Recio, M.C. Intestinal Anti-Inflammatory Activity of Ellagic Acid in the Acute and Chronic Dextrane Sulfate Sodium Models of Mice Colitis. J. Ethnopharmacol. 2013, 150, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Al-Sayed, E.; Korinek, M.; Esmat, A.; Chen, G.Y.; Cheng, Y.B.; Hsieh, P.W.; Chen, B.H.; Hwang, T.L. Anti-Inflammatory, Hepatoprotective and Antioxidant Activity of Ellagitannin Isolated from Melaleuca Styphelioides. Phytochemistry 2020, 177, 112429. [Google Scholar] [CrossRef]
- Senobari, Z.; Karimi, G.; Jamialahmadi, K. Ellagitannins, Promising Pharmacological Agents for the Treatment of Cancer Stem Cells. Phytother. Res. 2022, 36, 231–242. [Google Scholar] [CrossRef]
- Zhu, H.; Yan, Y.; Jiang, Y.; Meng, X. Ellagic Acid and Its Anti-Aging Effects on Central Nervous System. Int. J. Mol. Sci. 2022, 23, 937. [Google Scholar] [CrossRef] [PubMed]
- Garcia, G.; Pais, T.F.; Pinto, P.; Dobson, G.; McDougall, G.J.; Stewart, D.; Santos, C.N. Bioaccessible Raspberry Extracts Enriched in Ellagitannins and Ellagic Acid Derivatives Have Anti-Neuroinflammatory Properties. Antioxidants 2020, 9, 970. [Google Scholar] [CrossRef] [PubMed]
- Espín, J.C.; Larrosa, M.; García-Conesa, M.T.; Tomás-Barberán, F. Biological Significance of Urolithins, the Gut Microbial Ellagic Acid-Derived Metabolites: The Evidence so Far. Evid. Based Complement. Altern. Med. 2013, 2013, 270418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amarowicz, R.; Janiak, M. Hydrolysable Tannins. In Encyclopedia of Food Chemistry; Elsevier: Amsterdam, The Netherlands, 2018; pp. 337–343. [Google Scholar] [CrossRef]
- Jackson, R.S. Chemical Constituents of Grapes and Wine. In Wine Science; Elsevier: Amsterdam, The Netherlands, 2020; pp. 375–459. [Google Scholar] [CrossRef]
- Lipińska, L.; Klewicka, E.; Sójka, M. Structure, Occurrence and Biological Activity of Ellagitannins: A General Review. Acta Sci. Pol. Technol. Aliment. 2014, 13, 289–299. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Zhang, J.; Chen, N.G.; Shi, Z.; Qiu, J.; He, C.; Chen, M. Recent Advances in Anticancer Activities and Drug Delivery Systems of Tannins. Med. Res. Rev. 2017, 37, 665–701. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.; Wakamori, S.; Hirokane, T.; Ikeuchi, K.; Matsumoto, S. Structural Revisions in Natural Ellagitannins. Molecules 2018, 23, 1901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzo, J.M.; Munekata, P.E.; Putnik, P.; Kovačević, D.B.; Muchenje, V.; Barba, F.J. Sources, Chemistry, and Biological Potential of Ellagitannins and Ellagic Acid Derivatives. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2018; Volume 60, pp. 189–221. [Google Scholar] [CrossRef]
- Abe, L.T.; Lajolo, F.M.; Genovese, M.I. Potential Dietary Sources of Ellagic Acid and Other Antioxidants among Fruits Consumed in Brazil: Jabuticaba (Myrciaria Jaboticaba (Vell.) Berg). J. Sci. Food Agric. 2012, 92, 1679–1687. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, L.M.; Romanini, E.B.; Silva, E.; Pilau, E.J.; da Costa, S.C.; Madrona, G.S. Camu-Camu Bioactive Compounds Extraction by Ecofriendly Sequential Processes (Ultrasound Assisted Extraction and Reverse Osmosis). Ultrason. Sonochem. 2020, 64, 105017. [Google Scholar] [CrossRef] [PubMed]
- Torgbo, S.; Rugthaworn, P.; Sukatta, U.; Sukyai, P. Biological Characterization and Quantification of Rambutan (Nephelium Lappaceum L.) Peel Extract as a Potential Source of Valuable Minerals and Ellagitannins for Industrial Applications. ACS Omega 2022, 7, 34647–34656. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Gil, A.; del Alamo-Sanza, M.; Sánchez-Gómez, R.; Nevares, I. Alternative Woods in Enology: Characterization of Tannin and Low Molecular Weight Phenol Compounds with Respect to Traditional Oak Woods. A Review. Molecules 2020, 25, 1474. [Google Scholar] [CrossRef] [Green Version]
- Chira, K.; Teissedre, P.L. Extraction of Oak Volatiles and Ellagitannins Compounds and Sensory Profile of Wine Aged with French Winewoods Subjected to Different Toasting Methods: Behaviour during Storage. Food Chem. 2013, 140, 168–177. [Google Scholar] [CrossRef] [PubMed]
- García-Estévez, I.; Escribano-Bailón, M.T.; Rivas-Gonzalo, J.C.; Alcalde-Eon, C. Validation of a Mass Spectrometry Method to Quantify Oak Ellagitannins in Wine Samples. J. Agric. Food Chem. 2012, 60, 1373–1379. [Google Scholar] [CrossRef] [PubMed]
- Li, S.Y.; Duan, C.Q. Astringency, Bitterness and Color Changes in Dry Red Wines before and during Oak Barrel Aging: An Updated Phenolic Perspective Review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1840–1867. [Google Scholar] [CrossRef] [PubMed]
- Alañón, M.E.; Pimentel-Moral, S.; Arráez-Román, D.; Segura-Carretero, A. HPLC-DAD-Q-ToF-MS Profiling of Phenolic Compounds from Mango (Mangifera Indica L.) Seed Kernel of Different Cultivars and Maturation Stages as a Preliminary Approach to Determine Functional and Nutraceutical Value. Food Chem. 2021, 337, 127764. [Google Scholar] [CrossRef] [PubMed]
- Dorta, E.; González, M.; Lobo, M.G.; Sánchez-Moreno, C.; de Ancos, B. Screening of Phenolic Compounds in By-Product Extracts from Mangoes (Mangifera Indica L.) by HPLC-ESI-QTOF-MS and Multivariate Analysis for Use as a Food Ingredient. Food Res. Int. 2014, 57, 51–60. [Google Scholar] [CrossRef] [Green Version]
- You, Q.; Chen, F.; Sharp, J.L.; Wang, X.; You, Y.; Zhang, C. High-Performance Liquid Chromatography-Mass Spectrometry and Evaporative Light-Scattering Detector to Compare Phenolic Profiles of Muscadine Grapes. J. Chromatogr. A 2012, 1240, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.M.; Pereira, L.D.; Véras, J.H.; do Vale, C.R.; Chen-Chen, L.; da Santos, S.C. Protective Effect and Induction of DNA Repair by Myrciaria Cauliflora Seed Extract and Pedunculagin on Cyclophosphamide-Induced Genotoxicity. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2016, 810, 40–47. [Google Scholar] [CrossRef]
- Teixeira, L.D.L.; Bertoldi, F.C.; Lajolo, F.M.; Hassimotto, N.M.A. Identification of Ellagitannins and Flavonoids from Eugenia Brasilienses Lam. (Grumixama) by HPLC-ESI-MS/MS. J. Agric. Food Chem. 2015, 63, 5417–5427. [Google Scholar] [CrossRef]
- Lestario, L.N.; Howard, L.R.; Brownmiller, C.; Stebbins, N.B.; Liyanage, R.; Lay, J.O. Changes in Polyphenolics during Maturation of Java Plum (Syzygium Cumini Lam.). Food Res. Int. 2017, 100, 385–391. [Google Scholar] [CrossRef]
- Robbins, K.S.; Gong, Y.; Wells, M.L.; Greenspan, P.; Pegg, R.B. Investigation of the Antioxidant Capacity and Phenolic Constituents of U.S. Pecans. J. Funct. Foods 2015, 15, 11–22. [Google Scholar] [CrossRef]
- Sanz, M.; de Simón, B.F.; Esteruelas, E.; Muñoz, Á.M.; Cadahía, E.; Hernández, M.T.; Estrella, I.; Martinez, J. Polyphenols in Red Wine Aged in Acacia (Robinia Pseudoacacia) and Oak (Quercus Petraea) Wood Barrels. Anal. Chim. Acta 2012, 732, 83–90. [Google Scholar] [CrossRef]
- Winstel, D.; Capello, Y.; Quideau, S.; Marchal, A. Isolation of a New Taste-Active Brandy Tannin A: Structural Elucidation, Quantitation and Sensory Assessment. Food Chem. 2022, 377, 131963. [Google Scholar] [CrossRef]
- Elendran, S.; Muniyandy, S.; Lee, W.W.; Palanisamy, U.D. Permeability of the Ellagitannin Geraniin and Its Metabolites in a Human Colon Adenocarcinoma Caco-2 Cell Culture Model. Food Funct. 2019, 10, 602–615. [Google Scholar] [CrossRef]
- Sandhu, A.K.; Miller, M.G.; Thangthaeng, N.; Scott, T.M.; Shukitt-Hale, B.; Edirisinghe, I.; Burton-Freeman, B. Metabolic Fate of Strawberry Polyphenols after Chronic Intake in Healthy Older Adults. In Food and Function; Royal Society of Chemistry: London, UK, 2018; Volume 9, pp. 96–106. [Google Scholar] [CrossRef]
- Serrano, J.; Puupponen-Pimiä, R.; Dauer, A.; Aura, A.M.; Saura-Calixto, F. Tannins: Current Knowledge of Food Sources, Intake, Bioavailability and Biological Effects. Mol. Nutr. Food Res. 2009, 53, S310–S329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasprzak-Drozd, K.; Oniszczuk, T.; Stasiak, M.; Oniszczuk, A. Beneficial Effects of Phenolic Compounds on Gut Microbiota and Metabolic Syndrome. Int. J. Mol. Sci. 2021, 22, 3715. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhong, X.; Yan, J.; Sun, C.; Zhao, X.; Wang, X. Potential Roles of Gut Microbes in Biotransformation of Natural Products: An Overview. Front. Microbiol. 2022, 13, 956378. [Google Scholar] [CrossRef] [PubMed]
- Espín, J.C.; González-Barrio, R.; Cerdá, B.; López-Bote, C.; Rey, A.I.; Tomás-Barberán, F.A. Iberian Pig as a Model to Clarify Obscure Points in the Bioavailability and Metabolism of Ellagitannins in Humans. J. Agric. Food Chem. 2007, 55, 10476–10485. [Google Scholar] [CrossRef]
- Ludwig, I.A.; Mena, P.; Calani, L.; Borges, G.; Pereira-Caro, G.; Bresciani, L.; del Rio, D.; Lean, M.E.J.; Crozier, A. New Insights into the Bioavailability of Red Raspberry Anthocyanins and Ellagitannins. Free Radic. Biol. Med. 2015, 89, 758–769. [Google Scholar] [CrossRef] [Green Version]
- Cortés-Martín, A.; Selma, M.V.; Tomás-Barberán, F.A.; González-Sarrías, A.; Espín, J.C. Where to Look into the Puzzle of Polyphenols and Health? The Postbiotics and Gut Microbiota Associated with Human Metabotypes. Mol. Nutr. Food Res. 2020, 64, 1900952. [Google Scholar] [CrossRef]
- González-Sarrías, A.; García-Villalba, R.; Romo-Vaquero, M.; Alasalvar, C.; Örem, A.; Zafrilla, P.; Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C. Clustering According to Urolithin Metabotype Explains the Interindividual Variability in the Improvement of Cardiovascular Risk Biomarkers in Overweight-Obese Individuals Consuming Pomegranate: A Randomized Clinical Trial. Mol. Nutr. Food Res. 2017, 61, 1600830. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Martín, A.; García-Villalba, R.; González-Sarrías, A.; Romo-Vaquero, M.; Loria-Kohen, V.; Ramírez-De-Molina, A.; Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C. The Gut Microbiota Urolithin Metabotypes Revisited: The Human Metabolism of Ellagic Acid Is Mainly Determined by Aging. Food Funct. 2018, 9, 4100–4106. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; García-Villalba, R.; González-Sarrías, A.; Selma, M.V.; Espín, J.C. Ellagic Acid Metabolism by Human Gut Microbiota: Consistent Observation of Three Urolithin Phenotypes in Intervention Trials, Independent of Food Source, Age, and Health Status. J. Agric. Food Chem. 2014, 62, 6535–6538. [Google Scholar] [CrossRef]
- Selma, M.V.; Romo-Vaquero, M.; García-Villalba, R.; González-Sarrías, A.; Tomás-Barberán, F.A.; Espín, J.C. The Human Gut Microbial Ecology Associated with Overweight and Obesity Determines Ellagic Acid Metabolism. Food Funct. 2016, 7, 1769–1774. [Google Scholar] [CrossRef]
- Raimundo, A.F.; Ferreira, S.; Tomás-Barberán, F.A.; Santos, C.N.; Menezes, R. Urolithins: Diet-Derived Bioavailable Metabolites to Tackle Diabetes. Nutrients 2021, 13, 4285. [Google Scholar] [CrossRef] [PubMed]
- Cerdá, B.; Tomás-Barberán, F.A.; Espín, J.C. Metabolism of Antioxidant and Chemopreventive Ellagitannins from Strawberries, Raspberries, Walnuts, and Oak-Aged Wine in Humans: Identification of Biomarkers and Individual Variability. J. Agric. Food Chem. 2005, 53, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, J.; Quispe, C.; Castillo, C.M.S.; Caroca, R.; Lazo-Vélez, M.A.; Antonyak, H.; Polishchuk, A.; Lysiuk, R.; Oliinyk, P.; de Masi, L.; et al. Ellagic Acid: A Review on Its Natural Sources, Chemical Stability, and Therapeutic Potential. Oxidative Med. Cell. Longev. 2022, 2022, 3848084. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Singh, A.K.; Kumar, R.; Jamieson, S.; Pandey, A.K.; Bishayee, A. Neuroprotective Potential of Ellagic Acid: A Critical Review. Adv. Nutr. 2021, 12, 1211–1238. [Google Scholar] [CrossRef]
- Galano, A.; Francisco Marquez, M.; Pérez-González, A. Ellagic Acid: An Unusually Versatile Protector against Oxidative Stress. Chem. Res. Toxicol. 2014, 27, 904–918. [Google Scholar] [CrossRef]
- Galano, A.; Mazzone, G.; Alvarez-Diduk, R.; Marino, T.; Alvarez-Idaboy, J.R.; Russo, N. Food Antioxidants: Chemical Insights at the Molecular Level. Annu. Rev. Food Sci. Technol. 2016, 7, 335–352. [Google Scholar] [CrossRef]
- Marković, Z.; Milenković, D.; Crossed, D.; Signorović, J.; Marković, J.M.D.; Lučić, B.; Amić, D. A DFT and PM6 Study of Free Radical Scavenging Activity of Ellagic Acid. Mon. Für Chem. Chem. Mon. 2013, 144, 803–812. [Google Scholar] [CrossRef]
- Rusu, M.E.; Gheldiu, A.M.; Mocan, A.; Vlase, L.; Popa, D.S. Anti-Aging Potential of Tree Nuts with a Focus on the Phytochemical Composition, Molecular Mechanisms and Thermal Stability of Major Bioactive Compounds. Food Funct. 2018, 9, 2554–2575. [Google Scholar] [CrossRef] [PubMed]
- Rusu, M.E.; Simedrea, R.; Gheldiu, A.M.; Mocan, A.; Vlase, L.; Popa, D.S.; Ferreira, I.C.F.R. Benefits of Tree Nut Consumption on Aging and Age-Related Diseases: Mechanisms of Actions. Trends Food Sci. Technol. 2019, 8, 104–120. [Google Scholar] [CrossRef]
- Baek, B.; Lee, S.H.; Kim, K.; Lim, H.W.; Lim, C.J. Ellagic Acid Plays a Protective Role against UV-B-Induced Oxidative Stress by up-Regulating Antioxidant Components in Human Dermal Fibroblasts. Korean J. Physiol. Pharmacol. 2016, 20, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Pereira, D.M.; Valentão, P.; Pereira, J.A.; Andrade, P.B. Phenolics: From Chemistry to Biology. Molecules 2009, 14, 2202–2211. [Google Scholar] [CrossRef]
- Salem, A.M.; Mohammaden, T.F.; Ali, M.A.M.; Mohamed, E.A.; Hassan, H.F. Ellagic and Ferulic Acids Alleviate Gamma Radiation and Aluminium Chloride-Induced Oxidative Damage. Life Sci. 2016, 160, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Eghbaliferiz, S.; Iranshahi, M. Prooxidant Activity of Polyphenols, Flavonoids, Anthocyanins and Carotenoids: Updated Review of Mechanisms and Catalyzing Metals. Phytother. Res. 2016, 30, 1379–1391. [Google Scholar] [CrossRef]
- Rudrapal, M.; Khairnar, S.J.; Khan, J.; Dukhyil, A.B.; Ansari, M.A.; Alomary, M.N.; Alshabrmi, F.M.; Palai, S.; Deb, P.K.; Devi, R. Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights into Protective Effects, Antioxidant Potentials and Mechanism(s) of Action. Front. Pharmacol. 2022, 13, 806470. [Google Scholar] [CrossRef]
- Olas, B. Berry Phenolic Antioxidants—Implications for Human Health? Front. Pharmacol. 2018, 9, 78. [Google Scholar] [CrossRef]
- Kallio, T.; Kallio, J.; Jaakkola, M.; Mäki, M.; Kilpeläinen, P.; Virtanen, V. Urolithins Display Both Antioxidant and Pro-Oxidant Activities Depending on Assay System and Conditions. J. Agric. Food Chem. 2013, 61, 10720–10729. [Google Scholar] [CrossRef]
- Moilanen, J.; Karonen, M.; Tähtinen, P.; Jacquet, R.; Quideau, S.; Salminen, J.P. Biological Activity of Ellagitannins: Effects as Anti-Oxidants, pro-Oxidants and Metal Chelators. Phytochemistry 2016, 125, 65–72. [Google Scholar] [CrossRef]
- Seeram, N.P.; Adams, L.S.; Henning, S.M.; Niu, Y.; Zhang, Y.; Nair, M.G.; Heber, D. In Vitro Antiproliferative, Apoptotic and Antioxidant Activities of Punicalagin, Ellagic Acid and a Total Pomegranate Tannin Extract Are Enhanced in Combination with Other Polyphenols as Found in Pomegranate Juice. J. Nutr. Biochem. 2005, 16, 360–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfundstein, B.; el Desouky, S.K.; Hull, W.E.; Haubner, R.; Erben, G.; Owen, R.W. Polyphenolic Compounds in the Fruits of Egyptian Medicinal Plants (Terminalia Bellerica, Terminalia Chebula and Terminalia Horrida): Characterization, Quantitation and Determination of Antioxidant Capacities. Phytochemistry 2010, 71, 1132–1148. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Tao, X.; Men, X.; Xu, Z.; Wang, T. In Vitro and In Vivo Antioxidant Activities of Three Major Polyphenolic Compounds in Pomegranate Peel: Ellagic Acid, Punicalin, and Punicalagin. J. Integr. Agric. 2017, 16, 1808–1818. [Google Scholar] [CrossRef] [Green Version]
- Gülçin, I. Antioxidant Activity of Food Constituents: An Overview. Arch. Toxicol. 2012, 86, 345–391. [Google Scholar] [CrossRef] [PubMed]
- del Rio, D.; Costa, L.G.; Lean, M.E.J.; Crozier, A. Polyphenols and Health: What Compounds Are Involved? Nutr. Metab. Cardiovasc. Dis. 2010, 20, 1–6. [Google Scholar] [CrossRef]
- Panchal, S.K.; Brown, L. Cardioprotective and Hepatoprotective Effects of Ellagitannins from European Oak Bark (Quercus Petraea L.) Extract in Rats. Eur. J. Nutr. 2013, 52, 397–408. [Google Scholar] [CrossRef]
- Cerdá, B.; Espín, J.C.; Parra, S.; Martínez, P.; Tomás-Barberán, F.A. The Potent in Vitro Antioxidant Ellagitannins from Pomegranate Juice Are Metabolised into Bioavailable but Poor Antioxidant Hydroxy-6H-Dibenzopyran-6-One Derivatives by the Colonic Microflora of Healthy Humans. Eur. J. Nutr. 2004, 43, 205–220. [Google Scholar] [CrossRef]
- Cásedas, G.; Les, F.; López, V.; Choya-Foces, C.; Hugo, M. The Metabolite Urolithin-a Ameliorates Oxidative Stress in Neuro-2a Cells, Becoming a Potential Neuroprotective Agent. Antioxidants 2020, 9, 177. [Google Scholar] [CrossRef] [Green Version]
- Alfei, S.; Turrini, F.; Catena, S.; Zunin, P.; Parodi, B.; Zuccari, G.; Pittaluga, A.M.; Boggia, R. Preparation of Ellagic Acid Micro and Nano Formulations with Amazingly Increased Water Solubility by Its Entrapment in Pectin or Non-PAMAM Dendrimers Suitable for Clinical Applications. New J. Chem. 2019, 43, 2438–2448. [Google Scholar] [CrossRef]
- Ito, H. Metabolites of the Ellagitannin Geraniin and Their Antioxidant Activities. Planta Med. 2011, 77, 1110–1115. [Google Scholar] [CrossRef] [Green Version]
- Ishimoto, H.; Tai, A.; Yoshimura, M.; Amakura, Y.; Yoshida, T.; Hatano, T.; Ito, H. Antioxidative Properties of Functional Polyphenols and Their Metabolites Assessed by an ORAC Assay. Biosci. Biotechnol. Biochem. 2012, 76, 395–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized Methods for the Determination of Antioxidant Capacity and Phenolics in Foods and Dietary Supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A.; Nance, K.; Chen, S. The Gut–Brain Axis. Annu. Rev. Med. 2022, 73, 439–453. [Google Scholar] [CrossRef]
- Long-Smith, C.; O’riordan, K.J.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Microbiota-Gut-Brain Axis: New Therapeutic Opportunities. Annu. Rev. Pharmacol. Toxicol. 2019, 60, 477–502. [Google Scholar] [CrossRef] [Green Version]
- Battaglin, F.; Jayachandran, P.; Strelez, C.; Lenz, A.; Algaze, S.; Soni, S.; Lo, J.H.; Yang, Y.; Millstein, J.; Zhang, W.; et al. Neurotransmitter Signaling: A New Frontier in Colorectal Cancer Biology and Treatment. Oncogene 2022, 41, 4769–4778. [Google Scholar] [CrossRef]
- Yoshina, S.; Izuhara, L.; Kamatani, N.; Mitani, S. Regulation of Aging by Balancing Mitochondrial Function and Antioxidant Levels. J. Physiol. Sci. 2022, 72, 28. [Google Scholar] [CrossRef] [PubMed]
- Mateș, L.; Popa, D.S.; Rusu, M.E.; Fizeșan, I.; Leucuța, D. Walnut Intake Interventions Targeting Biomarkers of Metabolic Syndrome and Inflammation in Middle-Aged and Older Adults: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Antioxidants 2022, 11, 1412. [Google Scholar] [CrossRef]
- Liu, C.F.; Li, X.L.; Zhang, Z.L.; Qiu, L.; Ding, S.X.; Xue, J.X.; Zhao, G.P.; Li, J. Antiaging Effects of Urolithin a on Replicative Senescent Human Skin Fibroblasts. Rejuvenation Res. 2019, 22, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.B.; Lee, S.; Kim, J.H. Neuroprotective Effects of Urolithin a on H2O2-Induced Oxidative Stress-Mediated Apoptosis in SK-N-MC Cells. Nutr. Res. Pract. 2020, 14, 3–11. [Google Scholar] [CrossRef]
- Shi, P.-Z.; Wang, J.-W.; Wang, P.-C.; Han, B.; Lu, X.-H.; Ren, Y.-X.; Feng, X.-M.; Cheng, X.-F.; Zhang, L. Urolithin a Alleviates Oxidative Stress-Induced Senescence in Nucleus Pulposus-Derived Mesenchymal Stem Cells through SIRT1/PGC-1α Pathway. World J. Stem Cells 2021, 13, 1928–1946. [Google Scholar] [CrossRef]
- Albasher, G.; Alkahtani, S.; Al-Harbi, L.N. Urolithin A Prevents Streptozotocin-Induced Diabetic Cardiomyopathy in Rats by Activating SIRT1. Saudi J. Biol. Sci. 2022, 29, 1210–1220. [Google Scholar] [CrossRef]
- Muku, G.E.; Murray, I.A.; Espín, J.C.; Perdew, G.H. Urolithin a Is a Dietary Microbiota-Derived Human Aryl Hydrocarbon Receptor Antagonist. Metabolites 2018, 8, 86. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Chandrashekharappa, S.; Bodduluri, S.R.; Baby, B.V.; Hegde, B.; Kotla, N.G.; Hiwale, A.A.; Saiyed, T.; Patel, P.; Vijay-Kumar, M.; et al. Enhancement of the Gut Barrier Integrity by a Microbial Metabolite through the Nrf2 Pathway. Nat. Commun. 2019, 10, 89. [Google Scholar] [CrossRef] [Green Version]
- Rebeaud, J.; Peter, B.; Pot, C. How Microbiota-Derived Metabolites Link the Gut to the Brain during Neuroinflammation. Int. J. Mol. Sci. 2022, 23, 10128. [Google Scholar] [CrossRef]
- Rothhammer, V.; Mascanfroni, I.D.; Bunse, L.; Takenaka, M.C.; Kenison, J.E.; Mayo, L.; Chao, C.C.; Patel, B.; Yan, R.; Blain, M.; et al. Type I Interferons and Microbial Metabolites of Tryptophan Modulate Astrocyte Activity and Central Nervous System Inflammation via the Aryl Hydrocarbon Receptor. Nat. Med. 2016, 22, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Duarte-Silva, E.; Meuth, S.G.; Peixoto, C.A. Microbial Metabolites in Multiple Sclerosis: Implications for Pathogenesis and Treatment. Front. Neurosci. 2022, 16, 885031. [Google Scholar] [CrossRef]
- Hering, N.A.; Luettig, J.; Jebautzke, B.; Schulzke, J.D.; Rosenthal, R. The Punicalagin Metabolites Ellagic Acid and Urolithin A Exert Different Strengthening and Anti-Inflammatory Effects on Tight Junction-Mediated Intestinal Barrier Function In Vitro. Front. Pharmacol. 2021, 12, 610164. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, D.E.; Cremonini, E.; Fraga, C.G.; Oteiza, P.I. Ellagic Acid Protects Caco-2 Cell Monolayers against Inflammation-Induced Permeabilization. Free Radic. Biol. Med. 2020, 152, 776–786. [Google Scholar] [CrossRef]
- Han, Q.A.; Yan, C.; Wang, L.; Li, G.; Xu, Y.; Xia, X. Urolithin A Attenuates Ox-LDL-Induced Endothelial Dysfunction Partly by Modulating MicroRNA-27 and ERK/PPAR-γ Pathway. Mol. Nutr. Food Res. 2016, 60, 1933–1943. [Google Scholar] [CrossRef]
- Dirimanov, S.; Högger, P. Screening of Inhibitory Effects of Polyphenols on Akt-phosphorylation in Endothelial Cells and Determination of Structure-activity Features. Biomolecules 2019, 9, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotewicz, M.; Krauze-Baranowska, M.; Daca, A.; Płoska, A.; Godlewska, S.; Kalinowski, L.; Lewko, B. Urolithins Modulate the Viability, Autophagy, Apoptosis, and Nephrin Turnover in Podocytes Exposed to High Glucose. Cells 2022, 11, 2471. [Google Scholar] [CrossRef] [PubMed]
- Bobowska, A.; Granica, S.; Filipek, A.; Melzig, M.F.; Moeslinger, T.; Zentek, J.; Kruk, A.; Piwowarski, J.P. Comparative Studies of Urolithins and Their Phase II Metabolites on Macrophage and Neutrophil Functions. Eur. J. Nutr. 2021, 60, 1957–1972. [Google Scholar] [CrossRef]
- Komatsu, W.; Kishi, H.; Yagasaki, K.; Ohhira, S. Urolithin A Attenuates Pro-Inflammatory Mediator Production by Suppressing PI3-K/Akt/NF-ΚB and JNK/AP-1 Signaling Pathways in Lipopolysaccharide-Stimulated RAW264 Macrophages: Possible Involvement of NADPH Oxidase-Derived Reactive Oxygen Species. Eur. J. Pharmacol. 2018, 833, 411–424. [Google Scholar] [CrossRef]
- Piwowarski, J.P.; Kiss, A.K.; Granica, S.; Moeslinger, T. Urolithins, Gut Microbiota-Derived Metabolites of Ellagitannins, Inhibit LPS-Induced Inflammation in RAW 264.7 Murine Macrophages. Mol. Nutr. Food Res. 2015, 59, 2168–2177. [Google Scholar] [CrossRef]
- Kiss, A.K.; Piwowarski, J.P. Ellagitannins, Gallotannins and Their Metabolites- The Contribution to the Anti-Inflammatory Effect of Food Products and Medicinal Plants. Curr. Med. Chem. 2016, 25, 4946–4967. [Google Scholar] [CrossRef] [PubMed]
- Boakye, Y.D.; Groyer, L.; Heiss, E.H. An Increased Autophagic Flux Contributes to the Anti-Inflammatory Potential of Urolithin A in Macrophages. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 61–70. [Google Scholar] [CrossRef]
- Giménez-Bastida, J.A.; González-Sarrías, A.; Espín, J.C.; Schneider, C. Inhibition of 5-Lipoxygenase-Derived Leukotrienes and Hemiketals as a Novel Anti-Inflammatory Mechanism of Urolithins. Mol. Nutr. Food Res. 2020, 64, 2000129. [Google Scholar] [CrossRef]
- Zhang, S.; Al-Maghout, T.; Cao, H.; Pelzl, L.; Salker, M.S.; Veldhoen, M.; Cheng, A.; Lang, F.; Singh, Y. Gut Bacterial Metabolite Urolithin A (UA) Mitigates Ca2+ Entry in T Cells by Regulating MiR-10a-5p. Front. Immunol. 2019, 10, 1737. [Google Scholar] [CrossRef] [Green Version]
- Velagapudi, R.; Lepiarz, I.; El-Bakoush, A.; Katola, F.O.; Bhatia, H.; Fiebich, B.L.; Olajide, O.A. Induction of Autophagy and Activation of SIRT-1 Deacetylation Mechanisms Mediate Neuroprotection by the Pomegranate Metabolite Urolithin A in BV2 Microglia and Differentiated 3D Human Neural Progenitor Cells. Mol. Nutr. Food Res. 2019, 63, 1801237. [Google Scholar] [CrossRef] [Green Version]
- Toney, A.M.; Albusharif, M.; Works, D.; Polenz, L.; Schlange, S.; Chaidez, V.; Ramer-Tait, A.E.; Chung, S. Differential Effects of Whole Red Raspberry Polyphenols and Their Gut Metabolite Urolithin a on Neuroinflammation in BV-2 Microglia. Int. J. Environ. Res. Public Health 2021, 18, 68. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Park, J.S.; Lee, E.J.; Ahn, J.H.; Kim, H.S. Anti-Inflammatory and Antioxidant Mechanisms of Urolithin B in Activated Microglia. Phytomedicine 2019, 55, 50–57. [Google Scholar] [CrossRef]
- Xu, J.; Yuan, C.; Wang, G.; Luo, J.; Ma, H.; Xu, L.; Mu, Y.; Li, Y.; Seeram, N.P.; Huang, X.; et al. Urolithins Attenuate LPS-Induced Neuroinflammation in BV2Microglia via MAPK, Akt, and NF-ΚB Signaling Pathways. J. Agric. Food Chem. 2018, 66, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Busto, R.; Serna, J.; Perianes-Cachero, A.; Quintana-Portillo, R.; García-Seisdedos, D.; Canfrán-Duque, A.; Paino, C.L.; Lerma, M.; Casado, M.E.; Martín-Hidalgo, A.; et al. Ellagic Acid Protects from Myelin-Associated Sphingolipid Loss in Experimental Autoimmune Encephalomyelitis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 958–967. [Google Scholar] [CrossRef]
- Jayatunga, D.P.W.; Hone, E.; Khaira, H.; Lunelli, T.; Singh, H.; Guillemin, G.J.; Fernando, W.M.A.D.B.; Garg, M.L.; Verdile, G.; Martins, R.N. Therapeutic Potential of Mitophagy-Inducing Microflora Metabolite, Urolithin a for Alzheimer’s Disease. Nutrients 2021, 13, 3744. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, N.; Das, A.; Biswas, N.; Gnyawali, S.; Singh, K.; Gorain, M.; Polcyn, C.; Khanna, S.; Roy, S.; Sen, C.K. Urolithin A Augments Angiogenic Pathways in Skeletal Muscle by Bolstering NAD+ and SIRT1. Sci. Rep. 2020, 10, 20184. [Google Scholar] [CrossRef] [PubMed]
- Li, H.L.; Zhang, S.Y.; Ren, Y.S.; Zhou, J.C.; Zhou, Y.X.; Huang, W.Z.; Piao, X.H.; Yang, Z.Y.; Wang, S.M.; Ge, Y.W. Identification of Ellagic Acid and Urolithins as Natural Inhibitors of Aβ25–35-Induced Neurotoxicity and the Mechanism Predication Using Network Pharmacology Analysis and Molecular Docking. Front. Nutr. 2022, 9, 1761. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Jung, Y.H.; Choi, G.E.; Kim, J.S.; Chae, C.W.; Lim, J.R.; Kim, S.Y.; Yoon, J.H.; Cho, J.H.; Lee, S.J.; et al. Urolithin A Suppresses High Glucose-Induced Neuronal Amyloidogenesis by Modulating TGM2-Dependent ER-Mitochondria Contacts and Calcium Homeostasis. Cell Death Differ. 2021, 28, 184–202. [Google Scholar] [CrossRef]
- Esselun, C.; Theyssen, E.; Eckert, G.P. Effects of Urolithin a on Mitochondrial Parameters in a Cellular Model of Early Alzheimer Disease. Int. J. Mol. Sci. 2021, 22, 8333. [Google Scholar] [CrossRef]
- Chen, P.; Chen, F.; Lei, J.; Wang, G.; Zhou, B. The Gut Microbiota Metabolite Urolithin B Improves Cognitive Deficits by Inhibiting Cyt C-Mediated Apoptosis and Promoting the Survival of Neurons Through the PI3K Pathway in Aging Mice. Front. Pharmacol. 2021, 12, 768097. [Google Scholar] [CrossRef]
- Singh, R.; Chandrashekharappa, S.; Vemula, P.K.; Haribabu, B.; Jala, V.R. Microbial Metabolite Urolithin B Inhibits Recombinant Human Monoamine Oxidase A Enzyme. Metabolites 2020, 10, 258. [Google Scholar] [CrossRef]
- Zhao, W.; Shi, F.; Guo, Z.; Zhao, J.; Song, X.; Yang, H. Metabolite of Ellagitannins, Urolithin A Induces Autophagy and Inhibits Metastasis in Human Sw620 Colorectal Cancer Cells. Mol. Carcinog. 2018, 57, 193–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Wetidy, M.S.; Ahmad, R.; Rady, I.; Helal, H.; Rady, M.I.; Vaali-Mohammed, A.; Al-Khayal, K.; Traiki, T.B.; Abdulla, M.H. Urolithin A Induces Cell Cycle Arrest and Apoptosis by Inhibiting Bcl-2, Increasing P53-P21 Proteins and Reactive Oxygen Species Production in Colorectal Cancer Cells. Cell Stress Chaperones 2021, 26, 473–493. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Jung, H.; Lee, H.; Yi, H.C.; Kwak, H.K.; Hwang, K.T. Chemopreventive Activity of Ellagitannins and Their Derivatives from Black Raspberry Seeds on HT-29 Colon Cancer Cells. Food Funct. 2015, 6, 1675–1683. [Google Scholar] [CrossRef]
- Norden, E.; Heiss, E.H. Urolithin A Gains in Antiproliferative Capacity by Reducing the Glycolytic Potential via the P53/TIGAR Axis in Colon Cancer Cells. Carcinogenesis 2019, 40, 93–101. [Google Scholar] [CrossRef]
- Liu, F.; Cui, Y.; Yang, F.; Xu, Z.; Da, L.T.; Zhang, Y. Inhibition of Polypeptide N-Acetyl-α-Galactosaminyltransferases Is an Underlying Mechanism of Dietary Polyphenols Preventing Colorectal Tumorigenesis. Bioorg. Med. Chem. 2019, 27, 3372–3382. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Bastida, J.A.; Ávila-Gálvez, M.Á.; Espín, J.C.; González-Sarrías, A. The Gut Microbiota Metabolite Urolithin A, but Not Other Relevant Urolithins, Induces P53-Dependent Cellular Senescence in Human Colon Cancer Cells. Food Chem. Toxicol. 2020, 139, 111260. [Google Scholar] [CrossRef]
- Qiu, Z.; Zhou, J.; Zhang, C.; Cheng, Y.; Hu, J.; Zheng, G. Antiproliferative Effect of Urolithin a, the Ellagic Acid-Derived Colonic Metabolite, on Hepatocellular Carcinoma HepG2.2.15 Cells by Targeting Lin28a/Let-7a Axis. Braz. J. Med. Biol. Res. 2018, 51, e7220. [Google Scholar] [CrossRef] [Green Version]
- Lv, M.Y.; Shi, C.J.; Pan, F.F.; Shao, J.; Feng, L.; Chen, G.; Ou, C.; Zhang, J.F.; Fu, W.M. Urolithin B Suppresses Tumor Growth in Hepatocellular Carcinoma through Inducing the Inactivation of Wnt/β-Catenin Signaling. J. Cell. Biochem. 2019, 120, 17273–17282. [Google Scholar] [CrossRef]
- Mehra, S.; Deshpande, N.; Nagathihalli, N. Targeting Pi3k Pathway in Pancreatic Ductal Adenocarcinoma: Rationale and Progress. Cancers 2021, 13, 4434. [Google Scholar] [CrossRef]
- Totiger, T.M.; Srinivasan, S.; Jala, V.R.; Lamichhane, P.; Dosch, A.R.; Gaidarski, A.A.; Joshi, C.; Rangappa, S.; Castellanos, J.; Vemula, P.K.; et al. Urolithin A, a Novel Natural Compound to Target PI3K/AKT/MTOR Pathway in Pancreatic Cancer. Mol. Cancer Ther. 2019, 18, 301–311. [Google Scholar] [CrossRef] [Green Version]
- Wong, M.H.; Xue, A.; Baxter, R.C.; Pavlakis, N.; Smith, R.C. Upstream and Downstream Co-Inhibition of Mitogen-Activated Protein Kinase and PI3K/Akt/MTOR Pathways in Pancreatic Ductal Adenocarcinoma. Neoplasia 2016, 18, 425–435. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Lu, C.; Tang, R.; Pan, Y.; Bao, S.; Qiu, Y.; Xie, M. Ellagic Acid Inhibits the Proliferation of Human Pancreatic Carcinoma PANC-1 Cells in Vitro and in Vivo. Oncotarget 2017, 8, 12301. [Google Scholar] [CrossRef] [Green Version]
- Sahashi, H.; Kato, A.; Yoshida, M.; Hayashi, K.; Naitoh, I.; Hori, Y.; Natsume, M.; Jinno, N.; Kachi, K.; Asano, G.; et al. Urolithin A Targets the AKT/WNK1 Axis to Induce Autophagy and Exert Anti-Tumor Effects in Cholangiocarcinoma. Front. Oncol. 2022, 12, 963314. [Google Scholar] [CrossRef] [PubMed]
- Liberal, J.; Carmo, A.; Gomes, C.; Cruz, M.T.; Batista, M.T. Urolithins Impair Cell Proliferation, Arrest the Cell Cycle and Induce Apoptosis in UMUC3 Bladder Cancer Cells. Investig. New Drugs 2017, 35, 671–681. [Google Scholar] [CrossRef]
- Xu, Q.; Shen, M.; Han, Y.; Diao, H. Effects of Ellagic Acid Supplementation on Jejunal Morphology, Digestive Enzyme Activities, Antioxidant Capacity, and Microbiota in Mice. Front. Microbiol. 2021, 12, 793576. [Google Scholar] [CrossRef]
- Chen, J.; Yang, H.; Sheng, Z. Ellagic Acid Activated PPAR Signaling Pathway to Protect Ileums against Castor Oil-Induced Diarrhea in Mice: Application of Transcriptome Analysis in Drug Screening. Front. Pharmacol. 2020, 10, 1681. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhao, M.; Mo, J.; Lan, G.; Liang, J. Dietary Supplementation Ellagic Acid on the Growth, Intestinal Immune Response, Microbiota, and Inflammation in Weaned Piglets. Front. Vet. Sci. 2022, 9, 980271. [Google Scholar] [CrossRef]
- Kusmardi, K.; Baihaqi, L.A.; Estuningtyas, A.; Sahar, N.; Sunaryo, H.; Tedjo, A. Ethanol Extract of Pomegranate (Punica Granatum) Peel in Increasing the Expression of Caspase-3 in DSS-Induced Mice. Int. J. Inflamm. 2021, 2021, 4919410. [Google Scholar] [CrossRef] [PubMed]
- Parisio, C.; Lucarini, E.; Micheli, L.; Toti, A.; Khatib, M.; Mulinacci, N.; Calosi, L.; Bani, D.; Mannelli, L.D.C.; Ghelardini, C. Pomegranate Mesocarp against Colitis-Induced Visceral Pain in Rats: Effects of a Decoction and Its Fractions. Int. J. Mol. Sci. 2020, 21, 4304. [Google Scholar] [CrossRef]
- Fotschki, B.; Juśkiewicz, J.; Kołodziejczyk, K.; Jurgoński, A.; Kosmala, M.; Milala, J.; Ognik, K.; Zduńczyk, Z. Protective Effects of Ellagitannin-Rich Strawberry Extracts on Biochemical and Metabolic Disturbances in Rats Fed a Diet High in Fructose. Nutrients 2018, 10, 445. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Yang, Q.; Shi, Z.H.; Zhou, M.; Yan, L.; Li, H.; Xie, Y.H.; Wang, S.W. The Anti-Inflammatory Effect of Feiyangchangweiyan Capsule and Its Main Components on Pelvic Inflammatory Disease in Rats via the Regulation of the NF-ΚB and BAX/BCL-2 Pathway. Evid. Based Complement. Altern. Med. 2019, 2019, 9585727. [Google Scholar] [CrossRef] [Green Version]
- Budriesi, R.; Vivarelli, F.; Canistro, D.; Aldini, R.; Marquillas, C.B.; Corazza, I.; Fato, R.; Cirillo, S.; Bergamini, C.; D’Errico, A.; et al. Liver and Intestinal Protective Effects of Castanea Sativa Mill. Bark Extract in High-Fat Diet Rats. PLoS ONE 2018, 13, e0201540. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Dang, H.; Wu, J.; Yuan, L.; Chen, X.; Yao, J. Weizmannia Coagulans BC2000 Plus Ellagic Acid Inhibits High-Fat-Induced Insulin Resistance by Remodeling the Gut Microbiota and Activating the Hepatic Autophagy Pathway in Mice. Nutrients 2022, 14, 4206. [Google Scholar] [CrossRef]
- Lo, Y.L.; Wang, C.S.; Chen, Y.C.; Wang, T.Y.; Chang, Y.H.; Chen, C.J.; Yang, C.P. Mitochondrion-Directed Nanoparticles Loaded with a Natural Compound and a Microrna for Promoting Cancer Cell Death via the Modulation of Tumor Metabolism and Mitochondrial Dynamics. Pharmaceutics 2020, 12, 756. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, S.; Wu, Y.; Liu, X.; Wang, J.; Han, D. Ellagic Acid Alleviates Diquat-Induced Jejunum Oxidative Stress in C57BL/6 Mice through Activating Nrf2 Mediated Signaling Pathway. Nutrients 2022, 14, 1103. [Google Scholar] [CrossRef]
- Mousavi, S.; Weschka, D.; Bereswill, S.; Heimesaat, M.M. Preclinical Evaluation of Oral Urolithin-a for the Treatment of Acute Campylobacteriosis in Campylobacter Jejuni Infected Microbiota-Depleted Il-10−/− Mice. Pathogens 2021, 10, 7. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Li, Y.; Yang, F.; Zeng, A.; Yang, S.; Luo, Y.; Zhang, Y.; Xie, Y.; Ye, T.; Xia, Y.; et al. The Extract from Punica Granatum (Pomegranate) Peel Induces Apoptosis and Impairs Metastasis in Prostate Cancer Cells. Biomed. Pharmacother. 2017, 93, 976–984. [Google Scholar] [CrossRef] [PubMed]
- Ghani, R.A.; Nik, N.; Malek, A.; Abdullah, N. Pomegranate Juice Induced Cell Cycle Arrest and Apoptosis via Mitochondrial Pathway in Human Lung Adenocarcinoma A549 Cells. Int. J. Eng. Technol. 2018, 7, 287–292. [Google Scholar] [CrossRef]
- Chen, P.; Chen, F.; Zhou, B. Antioxidative, Anti-Inflammatory and Anti-Apoptotic Effects of Ellagic Acid in Liver and Brain of Rats Treated by D-Galactose. Sci. Rep. 2018, 8, 1465. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Chen, F.; Lei, J.; Li, Q.; Zhou, B. Activation of the MiR-34a-Mediated SIRT1/MTOR Signaling Pathway by Urolithin A Attenuates d-Galactose-Induced Brain Aging in Mice. Neurotherapeutics 2019, 16, 1269–1282. [Google Scholar] [CrossRef]
- Zhong, L.; Liu, H.; Zhang, W.; Liu, X.; Jiang, B.; Fei, H.; Sun, Z. Ellagic Acid Ameliorates Learning and Memory Impairment in APP/PS1 Transgenic Mice via Inhibition of β-Amyloid Production and Tau Hyperphosphorylation. Exp. Ther. Med. 2018, 16, 4951–4958. [Google Scholar] [CrossRef] [Green Version]
- Ramadan, W.S.; Alkarim, S. Ellagic Acid Modulates the Amyloid Precursor Protein Gene via Superoxide Dismutase Regulation in the Entorhinal Cortex in an Experimental Alzheimer’s Model. Cells 2021, 10, 3511. [Google Scholar] [CrossRef]
- Wang, W.; Yang, L.; Liu, T.; Wang, J.; Wen, A.; Ding, Y. Ellagic Acid Protects Mice against Sleep Deprivation-Induced Memory Impairment and Anxiety by Inhibiting TLR4 and Activating Nrf2. Aging 2020, 12, 10457–10472. [Google Scholar] [CrossRef] [PubMed]
- Falahieh, K.H.; Falahieh, K.H.; Sarkaki, A.; Sarkaki, A.; Edalatmanesh, M.; Naseri, M.K.G.; Farbood, Y. Ellagic Acid Attenuates Post-Cerebral Ischemia and Reperfusion Behavioral Deficits by Decreasing Brain Tissue Inflammation in Rats. Iran. J. Basic Med. Sci. 2020, 23, 645–653. [Google Scholar] [CrossRef]
- Gong, Z.; Huang, J.; Xu, B.; Ou, Z.; Zhang, L.; Lin, X.; Ye, X.; Kong, X.; Long, D.; Sun, X.; et al. Urolithin A Attenuates Memory Impairment and Neuroinflammation in APP/PS1 Mice. J. Neuroinflamm. 2019, 16, 62. [Google Scholar] [CrossRef] [Green Version]
- Kshirsagar, S.; Alvir, R.V.; Pradeepkiran, J.A.; Hindle, A.; Vijayan, M.; Ramasubramaniam, B.; Kumar, S.; Reddy, A.P.; Reddy, P.H. A Combination Therapy of Urolithin A+EGCG Has Stronger Protective Effects than Single Drug Urolithin A in a Humanized Amyloid Beta Knockin Mice for Late-Onset Alzheimer’s Disease. Cells 2022, 11, 2660. [Google Scholar] [CrossRef]
- Moon, J.H.; Kim, J.M.; Lee, U.; Kang, J.Y.; Kim, M.J.; Lee, H.L.; Jeong, H.R.; Go, M.J.; Kim, H.J.; Park, H.W.; et al. Walnut Prevents Cognitive Impairment by Regulating the Synaptic and Mitochondrial Dysfunction via JNK Signaling and Apoptosis Pathway in High-Fat Diet-Induced C57BL/6 Mice. Molecules 2022, 27, 5316. [Google Scholar] [CrossRef]
- Kujawska, M.; Jourdes, M.; Kurpik, M.; Szulc, M.; Szaefer, H.; Chmielarz, P.; Kreiner, G.; Krajka-Kuźniak, V.; Mikołajczak, P.Ł.; Teissedre, P.L.; et al. Neuroprotective Effects of Pomegranate Juice against Parkinson’s Disease and Presence of Ellagitannins-Derived Metabolite—Urolithin A—In the Brain. Int. J. Mol. Sci. 2020, 21, 202. [Google Scholar] [CrossRef] [Green Version]
- Kujawska, M.; Jourdes, M.; Witucki, Ł.; Karaźniewicz-łada, M.; Szulc, M.; Górska, A.; Mikołajczak, P.; Teissedre, P.L.; Jodynis-liebert, J. Pomegranate Juice Ameliorates Dopamine Release and Behavioral Deficits in a Rat Model of Parkinson’s Disease. Brain Sci. 2021, 11, 1127. [Google Scholar] [CrossRef]
- Tancheva, L.P.; Lazarova, M.I.; Alexandrova, A.V.; Dragomanova, S.T.; Nicoletti, F.; Tzvetanova, E.R.; Hodzhev, Y.K.; Kalfin, R.E.; Miteva, S.A.; Mazzon, E.; et al. Neuroprotective Mechanisms of Three Natural Antioxidants on a Rat Model of Parkinson’s Disease: A Comparative Study. Antioxidants 2020, 9, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.M.; Zhou, Y.Z.; Sheng, S.; Li, J.J.; Wang, G.Q.; Zhang, F. Ellagic Acid Protects Dopamine Neurons via Inhibition of NLRP3 Inflammasome Activation in Microglia. Oxidative Med. Cell. Longev. 2020, 2020, 2963540. [Google Scholar] [CrossRef] [PubMed]
- Pieróg, M.; Socała, K.; Wyska, E.; Poleszak, E.; Wlaź, P. Effect of Ellagic Acid on Seizure Threshold in Two Acute Seizure Tests in Mice. Molecules 2021, 26, 4841. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhou, T.; Yu, H.; Chang, L.Y.; Wei, L.L. Corilagin Reduces the Frequency of Seizures and Improves Cognitive Function in a Rat Model of Chronic Epilepsy. Med. Sci. Monit. 2018, 24, 2832–2840. [Google Scholar] [CrossRef] [Green Version]
- Shen, P.X.; Li, X.; Deng, S.Y.; Zhao, L.; Zhang, Y.Y.; Deng, X.; Han, B.; Yu, J.; Li, Y.; Wang, Z.Z.; et al. Urolithin A Ameliorates Experimental Autoimmune Encephalomyelitis by Targeting Aryl Hydrocarbon Receptor. EBioMedicine 2021, 64, 103227. [Google Scholar] [CrossRef] [PubMed]
- Vallarino, G.; Salis, A.; Lucarini, E.; Turrini, F.; Olivero, G.; Roggeri, A.; Damonte, G.; Boggia, R.; Mannelli, L.D.C.; Ghelardini, C.; et al. Healthy Properties of a New Formulation of Pomegranate-Peel Extract in Mice Suffering from Experimental Autoimmune Encephalomyelitis. Molecules 2022, 27, 914. [Google Scholar] [CrossRef]
- Gong, Q.Y.; Cai, L.; Jing, Y.; Wang, W.; Yang, D.X.; Chen, S.W.; Tian, H.L. Urolithin A Alleviates Blood-Brain Barrier Disruption and Attenuates Neuronal Apoptosis Following Traumatic Brain Injury in Mice. Neural Regen. Res. 2022, 17, 2007–2013. [Google Scholar] [CrossRef]
- Okuyama, S.; Furukawa, Y.; Yoshimura, M.; Amakura, Y.; Nakajima, M.; Yoshida, T. Oenothein b, a Bioactive Ellagitannin, Activates the Extracellular Signal-Regulated Kinase 2 Signaling Pathway in the Mouse Brain. Plants 2021, 10, 1030. [Google Scholar] [CrossRef]
- Tan, S.; Tong, W.H.; Vyas, A. Urolithin-A Attenuates Neurotoxoplasmosis and Alters Innate Response towards Predator Odor. Brain Behav. Immun. Health 2020, 8, 100128. [Google Scholar] [CrossRef]
- Rusu, M.E.; Georgiu, C.; Pop, A.; Mocan, A.; Kiss, B.; Vostinaru, O.; Fizesan, I.; Stefan, M.G.; Gheldiu, A.M.; Mates, L.; et al. Antioxidant Effects of Walnut (Juglans Regia L.) Kernel and Walnut Septum Extract in a D-Galactose-Induced Aging Model and in Naturally Aged Rats. Antioxidants 2020, 9, 424. [Google Scholar] [CrossRef]
- Mirzaie, Z.; Bastani, A.; Hesami, S.; Pouryousefi, E.; Kavianpour, M.; Haghighian, H.K. Improving Effect of Ellagic Acid on Sleep Quality and Gastrointestinal Symptoms in Patient with Irritable Bowel Syndrome: Randomized Double-Blind Clinical Trial. Turk. J. Gastroenterol. 2021, 32, 937–944. [Google Scholar] [CrossRef]
- Mirzaie, Z.; Bastani, A.; Haji-Aghamohammadi, A.A.; Rashidi Nooshabadi, M.; Ahadinezhad, B.; Khadem Haghighian, H. Effects of Ellagic Acid on Oxidative Stress Index, Inflammatory Markers and Quality of Life in Patients with Irritable Bowel Syndrome: Randomized Double-Blind Clinical Trial. Clin. Nutr. Res. 2022, 11, 98. [Google Scholar] [CrossRef] [PubMed]
- Siddarth, P.; Li, Z.; Miller, K.J.; Ercoli, L.M.; Merril, D.A.; Henning, S.M.; Heber, D.; Small, G.W. Randomized Placebo-Controlled Study of the Memory Effects of Pomegranate Juice in Middle-Aged and Older Adults. Am. J. Clin. Nutr. 2020, 111, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Guo, Y.; Lee, R.; Henning, S.M.; Wang, J.; Pan, Y.; Qing, T.; Hsu, M.; Nguyen, A.; Prabha, S.; et al. Pomegranate Metabolites Impact Tryptophan Metabolism in Humans and Mice. Curr. Dev. Nutr. 2020, 4, nzaa165. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; D’Amico, D.; Andreux, P.A.; Fouassier, A.M.; Blanco-Bose, W.; Evans, M.; Aebischer, P.; Auwerx, J.; Rinsch, C. Urolithin A Improves Muscle Strength, Exercise Performance, and Biomarkers of Mitochondrial Health in a Randomized Trial in Middle-Aged Adults. Cell Rep. Med. 2022, 3, 100633. [Google Scholar] [CrossRef]
- Liu, S.; D’Amico, D.; Shankland, E.; Bhayana, S.; Garcia, J.M.; Aebischer, P.; Rinsch, C.; Singh, A.; Marcinek, D.J. Effect of Urolithin A Supplementation on Muscle Endurance and Mitochondrial Health in Older Adults: A Randomized Clinical Trial. JAMA Netw. Open 2022, 5, e2144279. [Google Scholar] [CrossRef]
- Andreux, P.A.; Blanco-Bose, W.; Ryu, D.; Burdet, F.; Ibberson, M.; Aebischer, P.; Auwerx, J.; Singh, A.; Rinsch, C. The Mitophagy Activator Urolithin A Is Safe and Induces a Molecular Signature of Improved Mitochondrial and Cellular Health in Humans. Nat. Metab. 2019, 1, 595–603. [Google Scholar] [CrossRef]
- Moraes-Neto, R.N.; Coutinho, G.G.; Ataíde, A.C.S.; de Oliveira Rezende, A.; Nascimento, C.E.C.; de Albuquerque, R.P.; da Rocha, C.Q.; Rêgo, A.S.; de Sousa Cartágenes, M.D.S.; Abreu-Silva, A.L.; et al. Ethyl Acetate Fraction of Bixa Orellana and Its Component Ellagic Acid Exert Antibacterial and Anti-Inflammatory Properties against Mycobacterium Abscessus Subsp. Massiliense. Antibiotics 2022, 11, 817. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Food Product | ETs/EA | Content | Reference |
|---|---|---|---|
| Fruits | |||
| Raspberry fruit (Rubus idaeus L.) | Sanguiin H-10 | 0.62 mg/g dw * | [26] |
| Sanguiin H-6 | 9.56 mg/g dw | [26] | |
| Lambertianin C | 9.79 mg/g dw | [26] | |
| Ellagic acid | 0.31 mg/g dw | [26] | |
| Blackberry fruit (Rubus fruticosus L.) | Lambertianin C | 11.0 mg/100 g rw ** | [30] |
| Lambertianin A | 39.8 mg/100 g rw | [30] | |
| Ellagic acid | 11.8 mg/100 g rw | [30] | |
| Strawberry (Fragaria × ananassa Duch.)—fruits of six cultivars | Pedunculagin | 0.24–1.38 mg/g dw | [29] |
| Potentillin | 0–1.69 mg/g dw | [29] | |
| Casuarictin | 0.12–1.30 mg/g dw | [29] | |
| Sanguiin H-6 | 0.12–1.55 mg/g dw | [29] | |
| Agrimoniin | 0.89–13.11 mg/g dw | [29] | |
| Fragariin A | 0.34–1.47 mg/g dw | [29] | |
| Pomegranate (Punica granatum L.)—peels of seven cultivars | Pedunculagin | 8.2–11.8 mg/g dw | [27] |
| α-punicalagin | 17.4–24.7 mg/g dw | [27] | |
| β-punicalagin | 25.3–32.9 mg/g dw | [27] | |
| β-punicalin | 3.1–7.1 mg/g dw | [27] | |
| Ellagic acid hexoside | 2.9–6.3 mg/g dw | [27] | |
| Galloyl-HHDP-gluconic acid | 1.1–1.7 mg/g dw | [27] | |
| Galloyl-HHDP-hexoside | 1.0–2.8 mg/g dw | [27] | |
| HHDP-hexoside | 1.7–3.2 mg/g dw | [27] | |
| Ellagic acid | 2.1–5.3 mg/g dw | [27] | |
| Mango peel and seed (Mangifera indica L.) | Valoneic acid dilactone | Not mentioned | [59] |
| Ellagic acid | 1.13–13.67 mg/100 g dw | [60] | |
| Costa Rican guava peel and flesh (Psidium friedrichsthalianum Nied.) | Geraniin isomer 1 | 120.6 mg/100 g dw (peel, flesh) | [28] |
| Geraniin isomer 2 | 9.3 mg/100 g dw (peel); 103.9 mg/100 g dw (flesh) | [28] | |
| Castalagin isomer 1 | 28.3 mg/100 g dw (peel); 39.6 mg/100 g dw (flesh) | [28] | |
| Castalagin isomer 2 | 84.6 mg/100 g dw (peel); 41.7 mg/100 g dw (flesh) | [28] | |
| Muscadine grapes (Vitis rotundifolia Michx.) | Sanguiin H-5 | 0.03–0.91 mg/g | [48] |
| Ellagic acid | 32.1 mg/100 g fw # (seed); 59.1–61.5 mg/100 g fw (skin); 14.63–17.70 mg/100 g fw (fruit) | [61] | |
| Jabuticaba (Myrciaria cauliflora Mart.) | Castalagin | 78.4 mg/100 g fruit fw | [62] |
| Vescalagin | 28.7 mg/100 g fruit fw | [62] | |
| Pedunculagin | 9.8 mg/100 g fruit fw | [62] | |
| Yellow grumixama fruit (Eugenia brasilienses Lam.—γ variety) | Castalagin/vescalagin, pedunculagin, strictinin, potentillin/casuarictin | 92 mg EAE ##/100 g fw | [63] |
| Purple grumixama fruit (Eugenia brasilienses Lam.—α variety) | Pedunculagin, strictinin, potentillin/casuarictin, tellimagrandin I | 82−243 mg EAE/100 g fw | [63] |
| Camu-camu (Myrciaria dubia (Kunth) McVaugh) | Alnusiin | Not mentioned | [53] |
| Java plum/black plum fruit (Syzygium cumini (L.) Skeels) | Vescalagin | 0–1.0 mg/g dw | [64] |
| Galloyl dihexahydroxydiphenoyl glucose | 1.0–7.5 mg/g dw | [64] | |
| Digalloyl dihexahydroxydiphenoyl glucose | 0.97–5.6 mg/g dw | [64] | |
| Ellagic acid | 1.1–11.9 mg/g dw | [64] | |
| Nuts | |||
| Walnuts (Juglans regia L.)—pellicle of six cultivars | Pedunculagin/casuariin (2 isomers) | 3.1–13.3 mg/g fw | [31] |
| Strictinin/isostrictinin (3 isomers) | 1.9–9.6 mg/g fw | [31] | |
| Trigalloyl-HHDP-glucose (4 isomers) | 2.9–27.5 mg/g fw | [31] | |
| Tellimagrandin 1 (3 isomers) | 3.1–27.9 mg/g fw | [31] | |
| Pterocarinin A | 0.5–2.8 mg/g fw | [31] | |
| Castalagin/vescalagin | 9.5–35.9 mg/g fw | [31] | |
| bis-HHDP-glucose derivative | 20.2–26.1 mg/g fw | [31] | |
| Casuarin/casuarictin | 8.6–39.8 mg/g fw | [31] | |
| Ellagic acid | 17.5–23.3 mg/g fw | [31] | |
| Pecans (Carya illinoinensis (Wangenh.) K.Koch) | Valoneic acid dilactone | 9.45 μg/g acetonic crude extract | [65] |
| Ellagic acid | 132.0 μg/g acetonic crude extract | [65] | |
| Chestnut (Castanea sativa Mill.)—peels of cultivar Longal | Vescalagin | 67.5–109.4 μg/g dw | [32] |
| Castalagin | 49.6–100.4 μg/g dw | [32] | |
| Ellagic acid | 47.6–3542.6 μg/g dw | [32] | |
| Beverages | |||
| Red oak-aged wines | Castalagin | 0.61–11.8 mg/L | [57] |
| 0.07–11.43 mg/L | [17] | ||
| Vescalagin | 0.194–6.4 mg/L | [57] | |
| 0.06–1.32 mg/L | [17] | ||
| Grandinin | 0.36–3.4 mg/L | [57] | |
| 0.02–0.63 mg/L | [17] | ||
| Roburin A | 0.01–0.24 mg/L | [17] | |
| Roburins B + C | 0.07–1.56 mg/L | [17] | |
| Roburin D | 0.04–0.91 mg/L | [17] | |
| Roburin E | 0.115–2.08 mg/L | [57] | |
| 0.02–0.79 mg/L | [17] | ||
| Ellagic acid | 7.88–11.61 mg/L | [66] | |
| White oak-aged wines | Castalagin | 0.328 mg/L | [57] |
| Vescalagin | 0.14 mg/L | [57] | |
| Grandinin | 0.061 mg/L | [57] | |
| Rosé oak-aged wines | Castalagin | 4.69 mg/L | [57] |
| Vescalagin | 2.3 mg/L | [57] | |
| Grandinin | 3.5 mg/L | [57] | |
| Roburin E | 1.5 mg/L | [57] | |
| Cognac eaux-de-vie | β-1-O-ethylvescalagin | 1.1 mg/L | [18] |
| β-1-O-ethylvescalin | 0.73–4.66 mg/L | [18] | |
| Brandy tannin A | 0.77–10.28 mg/L | [18] | |
| 0.4–4.2 mg/L | [67] | ||
| Brandy tannin B | 0.5–5.50 mg/L | [18] | |
| Castalagin | 1.34–4.90 mg/L | [18] | |
| Vescalagin | 0.06–0.64 mg/L | [18] | |
| Grandinin | 0.11–1.50 mg/L | [18] | |
| Roburin A | 0.01–0.07 mg/L | [18] | |
| Roburin B | 0.01–0.17 mg/L | [18] | |
| Roburin C | 0.03–0.61 mg/L | [18] | |
| Roburin D | 0.26–1.48 mg/L | [18] | |
| Roburin E | 0.03–0.61 mg/L | [18] | |
| Commercial cognac | β-1-O-ethylvescalin | 0.87–1.58 mg/L | [18] |
| Brandy tannin A | 1.7–3.38 mg/L | [18] | |
| 0.03–7.7 mg/L | [67] | ||
| Brandy tannin B | 0.37–1.21 mg/L | [18] |
| Substances Tested (ST)—Doses | Preclinical Model | Main Results | Reference |
|---|---|---|---|
| EA | C57BL/6J mice were divided into 3 groups: control, standard pellet diet supplemented with 0.1% EA, and with 0.3% EA, respectively, for 21 consecutive days. | EA treatment increased jejunal villus height (0.3% EA) and enhanced the enzymatic activities for the jejunal lactase and sucrase (both 0.1% and 0.3% EA), and the alkaline phosphatase (0.3% EA). EA (0.3%) showed significant antioxidant effects by increasing the mRNA expression of Nrf2 and HO-1, the enzymatic activities of the superoxide dismutase and catalase, and reducing the malondialdehyde level in the jejunum. EA (0.3%) proved a suitable ability to regulate intestinal microbiota: increased the count of Lactobacillus species and decreased the count of Escherichia coli. | [161] |
| EA—10 mg/mL | Castor oil-induced diarrhea in mice. BALB/c mice were divided into 4 groups: (1) control; (2) castor oil (orally, 0.2 mL); (3) castor oil (orally, 0.2 mL) + EA (orally, 0.3 mL) after 30 min; (4) 0.1 mL GW9662 (i.p., 1 mg/mL) and castor oil (orally, 0.2 mL) + EA (orally, 0.3 mL) after 30 min. | Transcriptome, histological assay, and qRT-PCR were performed on ileum tissues. EA protected the ileum of mice against castor oil-induced diarrhea, reducing inflammation. The pretreatment with GW9662, a PPAR-specific antagonist, inhibited the anti-inflammatory effect of EA. | [162] |
| EA-supplemented feed (500 mg/kg) | Healthy weaned piglets (30 days old) received feed supplemented with EA twice daily for 40 days vs. a control group fed with the same feed without EA. | EA reduced the diarrhea rate, significantly increased weight gain, and diminished the serum diamine oxidase (DAO) levels of weaned piglets. Transcriptome sequencing revealed the ability of EA to down-regulate the expression of some genes involved in seven pathways related to immune response. EA modulated the microbiota composition in the cecum and rectum. | [163] |
| Pomegranate peel extract (PPE) containing 11% EA | A colonic inflammation model induced by dextran sodium sulfate (DSS) in mice (7-day cycles with 2% DSS in drinking water and 7 days of drinking water without DSS, for 42 days). Swiss Webster mice receiving 2% DSS were divided into 5 groups: control, PPE-dose 1 (240 mg/kg/day), PPE-dose 2 (480 mg/kg/day), ASP (43 mg/kg/day), and ASP (43 mg/kg/day) + EA (26 mg/kg/day). | PPE (dose 2) significantly increased the caspase-3 expression in mice colon tissues collected three days after the last treatment of DSS. | [164] |
| Pomegranate mesocarp decoction (PMD)—300 mg/kg; Polysaccharide components of PMD (PCs)—300 mg/kg; Ellagitannin components of PMD (ECs)—45 mg/kg | Colitis-induced abdominal pain in male Sprague–Dawley rats by 2,4-dinitrobenzenesulfonic acid (DNBS). ST was orally administered for 14 days. | ECs have been more effective in reducing visceral pain than the equivalent dose of PMD, both at 7 and 14 days of repeated administration. All three treatments significantly diminished the inflammation degree of the colonic mucosa and the fibrosis state. However, ECs are the responsible bioactive compounds of PMD, and PCs support and enhance their effects. | [165] |
| Two dietary strawberry extracts rich in monomeric ellagitannins (ME) and dimeric ellagitannins (DE), respectively | Male Wistar rats fed with high-fructose diets for 6 weeks: 3 groups received a diet based on corn starch (C): C, C + ME, C + DE, in parallel with other 3 groups fed with a diet containing fructose (F): F, F + ME, F + DE. | ME was more effective in reducing oxidative stress (lipid peroxidation in some tissues) and serum inflammatory biomarkers (TNF-α, IL-6), as well as the serum and liver triglyceride levels, than DE. Higher levels of ET metabolites were determined in the cecum and urine of rats receiving ME than of those fed with DE. Both ME and DE regulated the biochemical disturbances induced by a high-fructose diet. The efficacity of the ME extract was associated with systemic parameters, while that of the DE extract was associated with local microbial activity. | [166] |
| Feiyangchangweiyan capsule (FYC) and its main components: EA, gallic acid (GA), and syringin (SY) | A pathogen-induced pelvic inflammatory disease (PID) model in female SD rats (E. coli and S. aureus induced infection in the upper genital tract) with 11 groups: control, PID, FYC (1.2 g/kg), GA (210 mg/kg), EA (30 mg/kg), SY (35 mg/kg), GA (105 mg/kg) + EA (15 mg/kg), GA (105 mg/kg) + SY (18 mg/kg), EA (15 mg/kg) + SY (18 mg/kg), GA (70 mg/kg) + EA (10 mg/kg) + SY (12 mg/kg), and Fuke Qianjin capsule (FKC) (2.4 g/kg) as a positive control. | Histological analysis, ELISA assays, and Western blot analysis were applied to detect the expression of NF-κB, BAX, BCL-2, and JNK. The expressions of IL-1β, TNF-α, MPC-1, and BAX induced by infection were significantly reduced, while the IL-10 level and the expression of Bcl-2 were increased by FYC, but also by its main components. The anti-inflammatory effects in the rats of the GA + EA + SY group were more intense than those observed in the rats treated with dimers or monomers. PID was associated with an elevated expression of BAX and a dramatic suppression of BCL-2 expression. FYC, and especially EA and SY, have shown an increased efficacy in reversing these effects. | [167] |
| Extract of Chestnut bark (ENC®) (20 mg/kg/day) rich in ellagitannins (administered by gavage) | A high-fat diet (HFD) model in rats Male Sprague–Dawley rats were divided into 4 groups: control (regular diet, RD), HFD, RD + ENC®, and HFD + ENC®, for 21 days. | In HFD-fed rats, ENC® improved lipidic profile (significantly reduced TC, LDL-C, TG, and increased HDL-C), exerted antioxidative and anti-inflammatory activities, and normalized intestinal contractility in ileal and colonic tissues. | [168] |
| EA—0.3 g/kg of HFD; Weizmannia coagulans BC2000 and BC77—0.1 g of lyophilized powder (containing 4 × 1011 CFU/g)/kg feed | Animal model of HFD-induced insulin resistance: C57BL/6J male mice were divided into 5 groups: (1) Low-fat diet (LFD); (2) HFD (providing 60% of fat energy); (3) HFD + EA; (4) HFD + EA + BC77; (5) HFD + EA + BC2000. | EA and W. coagulans BC2000 had a synergistic effect in reducing the insulin resistance index and HFD-induced endotoxemia. EA co-administered with BC200 activated the autophagy pathway in the mouse liver, a urolithin-like effect, but not with BC77. W. coagulans BC2000 promoted a favorable intestinal environment for the proliferation of EA-transformable bacteria. | [169] |
| EA (80 mg/kg) and miR-125 (1.25 mg/kg) nanoparticles carrying a mitochondrion-directed peptide (K) and a tumor-targeted ligand (L) | A SAS-tumor bearing mouse model. BALB/c nude mice were divided into 5 groups: saline (control), EA, EA/LPN (lipid-polymer nanoparticles), EA/LPN-KL, and EA/LPN-KL + miR-125/SLN (solid lipid nanoparticles)-KL. The formulations were administered every 2 days for 20 days. | All EA formulations have shown hypoglycemic and hypolipidemic effects, but the combined formula, EA/LPN-KL + miR-125/SLN-KL, was the most effective. This formula also showed great tumor-suppression ability in SAS-tumor-bearing BALB/c mice. | [170] |
| Urolithin A (UA)—20 mg/kg | Animal model of cholangiocarcinoma (CCA) in xenograft female nude mice (BALB/c Slc-nu/nu) injected with HuCCT-1 cells (5 × 106 cells/100 mL of media). UA treatment was initiated 2 weeks after tumor transplantation for a period of 35 days (orally, 3×/week). | The tumor volume was calculated twice a week, and immunohistochemical analysis was performed at the end of the experiment. UA inhibited tumor growth and increased LC3-II levels. On the other hand, the phospho-kinase array demonstrated the downregulation of the Akt/WNK1 pathway. LC3-II expression was elevated in WNK1 knocked-down cells, indicating that WNK1 is the key signal for regulating autophagy. Thus, UA exerted anti-tumor effects by suppressing the Akt/WNK1 signaling pathway and inducing autophagy. | [159] |
| EA—100 mg/kg bw | Animal model of oxidative stress induced in C57BL/6J mice by diquat (25 mg/kg bw, single dose). EA was administered orally for 5 days. | EA treatment significantly reduced diquat-induced weight loss and mitigated oxidative stress in jejunum: reduced ROS production, up- regulated the mRNA expression of Nrf2 and the antioxidant enzymes (GPx1 and HO-1). ML385, a specific Nrf2 inhibitor, counteracted the EA effects on jejunum oxidative stress. | [171] |
| UA (synthetic)—daily dose of 0.114 mg/kg bw in drinking water | Animal inflammation model: microbiota-depleted IL-10−/− mice perorally infected with C. jejuni (on days 0 and 1). UA treatment was initiated on day 2 post-infection (p.i.) for 5 consecutively days vs. a placebo group (water). | Versus the placebo group, UA improved the early (<24 h) clinical status of infected mice, reduced pathogen loads in the ileum, immune cell and apoptotic epithelial cell abundance in the colon (by histopathology and immunohistochemistry), and the secretion of proinflammatory biomarkers (IFN-γ, TNF-α, MCP-1, and IL-6) both in the intestinal tract and in the extraintestinal compartments (lung, liver, kidney, serum). | [172] |
| Substances Tested (ST)—Doses | Preclinical Model | Main Results | Reference |
|---|---|---|---|
| EA—p.o. administered in 3 doses: high (H), medium (M), and low (L) doses | Aging model obtained with D-gal (100 mg/kg/day, s.c., 8 weeks) in male Sprague–Dawley (SD) rats divided into 6 groups: (1) control, (2) D-gal, (3) positive control (vitamin E, 150 mg/kg, by gavage), (4) H-EA (D-gal + 150 mg EA/kg/day), (5) M-EA (D-gal + 100 mg EA/kg/day), (6) L-EA (D-gal + 50 mg EA/kg/day). | EA restored the antioxidant defense system (evaluated by SOD, CAT, GSH-Px, and T-AOC activities, and MDA levels, respectively) in the liver and brain of D-gal-induced aging rats, especially in H-dose. The treatment with M and H doses of EA for 8 weeks has significantly mitigated the D-gal-induced inflammation (TNF-α, IL-6, and IL-1β levels in serum) and the liver function decline (ALT and AST levels). Histopathological analysis showed that the H-dose of EA was more protective and kept the morphological structure in both the liver and brain. EA treatment significantly downregulated the expression of Bcl-2 and Bax proteins and showed anti-apoptotic effects in a concentration-dependent manner. | [175] |
| UA—p.o. administered in 3 doses: high (H), medium (M), and low (L) doses | I. Aging model obtained with D-gal (150 mg/kg/day, s.c., 8 weeks) in male Institute of Cancer Research (ICR) mice were divided into 5 groups: (1) control (Ctrl), (2) D-gal, (3) H-UA (D-gal + 150 mg UA/kg/day), (4) M-UA (D-gal + 100 mg UA/kg/day), (5) L-UA (D-gal + 50 mg UA/kg/day). II. Additional experiment with 4 groups:
| UA treatment significantly ameliorated D-gal-induced behavioral impairments (in Open field, Morris Water Maze, and Object–Place Recognition tests). UA significantly lowered the AChE and MAO levels and the oxidative stress (the activities of SOD, CAT, GSH-Px, T-AOC, and MDA levels, respectively) in the brain of D-gal-induced aging mice. UA showed neuroprotection against D-gal-induced aging downregulating miR-34a in the hippocampal tissue and activated autophagy by upregulating SIRT1 and downregulating the protein expression of p53/p21 and the mTOR signaling pathway. | [176] |
| Urolithin B (UB)—i.g. administered in 3 doses: high (H), medium (M), and low (L) doses | I. Aging model obtained with D-gal (150 mg/kg/day, s.c., 8 weeks) in male C57BL/6 mice were divided into 5 groups: (1) control, (2) D-gal, (3) H-UB (D-gal + 150 mg UB/kg/day), (4) M-UB (D-gal + 100 mg UB/kg/day), (5) L-UB (D-gal + 50 mg UB/kg/day). II. Additional experiment with 4 groups:
| Long-term UB treatment significantly ameliorated the behavioral features, learning, and memory function (in Open field, Morris Water Maze, and Y-maze tests) in D-gal-induced aging in mice. These outcomes were correlated with a significant reduction of AGE levels and elevation of Cu, Zn-SOD, and CAT expressions and activities in the brain. UB inhibited the apoptosis of hippocampal neurons induced by D-gal, downregulated the JNK signaling pathway, prevented the cytochrome c release from isolated mitochondria, increased the activation of Akt and p44/42 MAPK, and promoted the neuronal survival via the PI3K/Akt pathways. | [145] |
| EA—50 mg/kg/day, intragastric (i.g.) | Mouse model of AD: Male APP/PS1 double-transgenic and wild-type (WT) C57BL/6 mice were divided into 4 groups: (1) WT, (2) WT+EA, (3) APP/PS1, (4) APP/PS1 + EA, received EA or the same volume of 10% DMSO for 60 days. | EA treatment improved learning and memory abilities and ameliorated cognitive deficits in APP/PS1 mice, reduced neuronal cell apoptosis, the expression of caspase-3 level and the amyloid aggregates in hippocampus. EA also significantly inhibited tau hyperphosphorylation and decreased the expression of pSer199-tau and pSer396-tau in the hippocampus of APP/PS1 mice. Moreover, EA treatment significantly increased the expression of pSer473-AKT and decreased the pTyr216-GSK3β levels in APP/PS1 mice. | [177] |
| EA—50 mg/kg, p.o. | AD animal model induced with AlCl3 in male Wistar rats divided into 4 groups: (1) control, (2) EA (50 mg/kg, p.o., for 4 weeks), (3) AD model (50 mg AlCl3/kg, p.o., for 4 weeks), (4) AD + EA (50 mg AlCl3/kg, p.o., for 4 weeks, followed by 50 mg EA/kg/day, p.o., for 2 weeks). | The discrimination index for the novel object recognition test (NORT) was significantly increased by EA therapy in AD rats. EA treatment significantly increased SOD, GSH, and TAC levels and decreased MDA levels in the serum of AD rats. The neurofibrillary tangles and neuritic plaques in the entorhinal cortex (ERC) sections were reduced in the AD+EA group. Antioxidant activity of EA treatment (increased SOD mRNA expression and modulated the amyloid precursor protein toxicity and caspase-3-mediated apoptosis) was correlated with the restoration of ERC thickness in AD+EA rats vs. AD rats. | [178] |
| EA—50 mg/kg bw/day (EA50) and 100 mg/kg bw/day (EA100), i.p., for 21 days | Animal model of memory impairment and anxiety induced by sleep deprivation (SD). C57BL/6J mice were divided into 4 groups: (1) control, (2) SD, (3) SD + EA50, (4) SD + EA100. | EA ameliorated learning and memory deficits and alleviated anxiety-like behaviors in SD mice. EA treatment improved neuron survival, reversed dendritic spine density, and reduced shrinkage and loss of neurons in the hippocampus of SD mice. EA restored the SOD and GPx activities, decreased MDA levels, and activated the Nrf2/HO-1 pathway in the hippocampus of SD mice. EA also reduced the IL-1β, IL-6, and TNF-α hippocampal levels, normalized the expression levels of TLR4, MyD88, NF-κB p65, and p-IκBα, and inhibited the TLR4-mediated innate immune responses. Moreover, EA showed neuroprotective effects on glutamate-induced toxicity via both the Nrf2 and TLR4 signaling pathways. | [179] |
| EA—50, 75, and 100 EA mg/kg, by gavages, 3 times daily for one week | Animal model of brain inflammation induced by cerebral ischemia/reperfusion (I/R). Male Wistar rats were divided into 6 groups: (1) control (surgery without any I/R) + vehicle (Veh); (2) I/R + Veh; (3–5) I/R + EA; (6) positive control (intact rats received 100 mg EA/kg). | Only the higher dose of EA (100 mg/kg) improved post-ischemic complications: it significantly increased the neurological signs scores, significantly reversed all tested behaviors, restored the BBB permeability, and decreased brain edema and the brain tissue cytokine levels (TNF-α and IL-1β) vs. I/R + Veh rats. | [180] |
| UA—300 mg, p.o. | Mouse model of Alzheimer’s disease (AD): APPswe/PS1ΔE9 (APP/PS1) mice received 300 mg of UA dissolved in 0.5% carboxymethylcellulose each day for 14 days vs. 2 control groups: APP/PS1 transgenic mice and wild-type mice that received only the vehicle (0.5% carboxymethylcellulose). | UA ameliorated spatial learning and memory impairment, prevented neuronal apoptosis in the cortex and hippocampus, and enhanced hippocampal neurogenesis in APP/PS1 mice. UA also decreased Aβ plaque deposit levels in the cortex and hippocampus, attenuated reactive gliosis, and significantly reduced microglia and astrocyte activation in APP/PS1 mice. UA treatment increased the expression of p-AMPK and decreased the activation of P65NF-κB and P38MAPK. | [181] |
| UA—2.5 mg/kg/day, i.p., for 8 weeks | A streptozotocin (STZ)- induced diabetic mouse model. Male CrljOri:CD1(ICR) mice were divided into 2 groups: (1) STZ-control and (2) STZ-UA. | UA treatment improved cognitive impairment, APP and BACE1 expressions, Tau phosphorylation, and Aβ deposition in STZ-induced diabetic mice. UA decreased blood glucose levels, but only in control mice and not in the STZ-injected mice. | [143] |
| UA—2.5 mg/kg bw; egpigallocatechin gallate (EGCG)—25 mg/kg bw | Animal model of late-onset AD using humanized homozygous amyloid beta knockin (hAbKI) mice were divided into 3 groups: (1) control, (2) UA (i.p., 3×/week for 4 months), (3) UA+EGCG (i.p., 3×/week for 4 months). | Both UA and UA + EGCG have been effective in counteracting the onset of pathological features of AD in hAbKI mice: enhanced phenotypic behavior, significantly increased mitochondrial biogenesis proteins (Nrf2, TFAM), both mitophagy (PINK1, Parkin) and synaptic proteins (synaptophysin, PSD95), as well as autophagy proteins (Beclin, ATG5, LC3B1, LC3B2, BCL2). Neuroinflammatory biomarkers (microglial marker Iba1 and astrocytic marker GFAP) were reduced, and the neuronal marker NeuN was significantly increased by both UA and UA + EGCG treatments. Dendritic spines and lengths, mitochondrial length, and mitophagosomal formations were also increased by both treatments. However, combined treatment UA+EGCG was stronger and more effective than only UA for most of the determinations performed. | [182] |
| Juglans regia (Gimcheon 1ho cultivar, GC)—lyophilized ethanolic extracts (obtained with 80% ethanol at 40 °C for 2 h), containing EA and ellagitannins (pedunculagin/casuariin isomer, strictinin, tellimagrandin I, EA-O-pentoside, and EA)—20 mg/kg bw (GC20) and 50 mg/kg bw (GC50), p.o. | A diabetic animal model with cognitive dysfunction in mice induced by a high-fat diet (HFD) for 12 weeks. Male C57BL/6 mice were divided into 4 groups: (1) control, (2) HFD, (3) HFD+GC20, (4) HFD + GC50. The lyophilized extracts were administered for 4 weeks. | GC significantly restored the behavioral and memory dysfunction in HFD-induced diabetic mice for both doses (GC 20 and GC50). GC also improved serum lipid profile and reduced white adipose tissue (WAT) and liver fat mass and showed antioxidant effects (reduced levels of MDA—in hepatic and cerebral tissues—and serum AGEs, and increased serum level of FRAP). GC inhibited AChE activity in cerebral tissue, suppressed AChE expression, and upregulated choline acetyltransferase expression vs. HFD group. GC restored mitochondrial membrane potential, regulated the mitochondrial ROS production and the protein expressions associated with synaptic disorders and neuronal apoptosis: decreased p-JNK, p-tau, and Aβ levels, upregulated p-Akt (Ser 473) and insulin-degrading enzyme (IDE), and downregulated BAX and caspase-3 expression levels vs. HFD group. Moreover, GC ameliorated TNF-α, IL-1β, p-NFκB, and caspase-1 expression levels and upregulated heme oxygenase-1 (HO-1) expression levels vs. HFD group. | [183] |
| Pomegranate juice (PJ) rich in ellagitannins (galloyl-hexoside, EA-hexoside, pedunculagin, casuarinin) and EA, containing 18.90 ± 0.96 g/L (EA equivalents per L of juice) | Rat model of Parkinson’s disease (PD) induced with rotenone (1.3 mg/kg bw/day, s.c., for 35 days). Male albino Wistar rats were divided into 4 groups: (1) control (vehicle only), (2) PJ (500 mg/kg bw/day, i.g.), (3) rotenone only (1.3 mg/kg bw/day, s.c.) from the 11th day (ROT), (4) PJ (500 mg/kg bw/day, i.g.) + rotenone from the 11th day (PJ + ROT). Experiment included 10 days pretreatment with PJ and 35 days with PJ + ROT combined treatment. | PJ improved postural stability in rats affected by rotenone, enhanced neuronal survival, protected against oxidative damage (reduced MDA levels and increased CAT, GPx, GST activities in midbrain) and α-synuclein aggregation, increased the activity of mitochondrial aldehyde dehydrogenase, and normalized the expression of anti-apoptotic Bcl-xL protein. | [184] |
| PJ | Rat model of Parkinson’s disease (PD) induced with rotenone (1.3 mg/kg bw/day, s.c., for 35 days). Male albino Wistar rats were divided into 4 groups: (1) control (vehicle only), (2) PJ (500 mg/kg bw/day, i.g.), (3) rotenone only (1.3 mg/kg bw/day, s.c.) from the 11th day (ROT), (4) PJ (500 mg/kg bw/day, i.g.) + rotenone from the 11th day (PJ + ROT). Experiment included 10 days pretreatment with PJ and 35 days with PJ + ROT combined treatment. | PJ treatment proved neuroprotection against PD: improved vertical activity in rotenone-injected rats mitigated the reduction of the olfactory discrimination index to the level observed in control animals, attenuated the depletion of the dopamine (DA) and 3,4-dihydroxy-phenylacetic acid (DOPAC) in the midbrain. | [185] |
| EA, α-lipoic acid (LA), myrtenal (Myrt)—50 mg/kg (i.p.) | Rat model of Parkinson’s disease (PD) induced with 6-hydroxydopamine (6-OHDA) (intrastriatal injection). Male Wistar rats divided into 5 groups: (1) control, (2) striatal 6-OHDA-lesioned control, (3–5) striatal 6-OHDA-lesioned rats pre-treated for 5 days with EA, LA, and Myrt. | All three compounds (EA, LA, and Myrt) improved learning and memory performance as well as neuromuscular coordination in rats with 6-OHDA-induced PD. Moreover, all these compounds significantly decreased lipid peroxidation (LPO) levels and restored catalase (CAT) activity and DA levels that were impaired by the challenge with 6-OHDA. | [186] |
| EA—10 mg/kg and 50 mg/kg, p.o. | Animal model of dopamine (DA) neuronal damage induced by lipopolysaccharides (LPS). Male Wistar rats were divided into 5 groups: (1) control, (2) EA (50 mg/kg), (3) LPS, (4) LPS+EA (10 mg/kg), and (5) LPS +EA (50 mg/kg). | EA (50 mg/kg) attenuated LPS-induced DA neuronal loss and ameliorated the decrease in tyrosine hydroxylase expression in the substantia nigra neurons. EA also attenuated the activation of the NLRP3 inflammasome in microglia and inhibited the NLRP3 inflammasome signaling pathway activated by LPS. The protein expressions of Iba-1 and proinflammatory cytokines IL-1β, TNF-α, and IL-18 were significantly suppressed by EA treatment. | [187] |
| EA—100 mg/kg and 200 mg/kg, i.p. and p.o. | Male albino Swiss mice—i.v. pentylenetetrazole (PTZ) seizure threshold test, maximal electroshock seizure test (MEST), grip-strength test, and chimney test, vs. valproic acid as reference drug. | EA (100 mg/kg) significantly increased the threshold of mice injected with PTZ for the first myoclonic twitch and generalized clonic seizures associated with the loss of the righting reflex, but not the threshold for forelimb tonus. In the MEST test, EA affected the threshold for the tonic hindlimb extension only at the high dose (200 mg/kg). EA did not have any effects on neuromuscular strength and motor coordination in mice at any of the doses tested. | [188] |
| Corilagin—10 mg/kg and 20 mg/kg, i.p. | Male Wistar rats—i.v. PTZ seizure threshold test. | Corilagin significantly reduced the epileptic events and improved cognitive function (in the Morris water maze (MWM) navigation test), reduced TNF-α and increased IL-10 levels, reduced ROS production, mitochondrial swelling, and carbonic anhydrase activity in the brain tissues in a dose-dependent manner. Corilagin treatment prevented structural damage of neurons and maintained the number of surviving cells vs. control group. | [189] |
| UA—10, 25, or 50 mg/kg/day, p.o. | Animal model of an experimental autoimmune encephalomyelitis (EAE) in C57BL/6 female mice immunized with MOG35–55. | The dose of 25 mg/kg significantly suppressed the progress of EAE. UA treatment decreased demyelination, significantly inhibited inflammatory cell infiltration, reduced neuroinflammation (lowered numbers of M1-type microglia and inhibited the activation of dendritic cells). | [190] |
| Pomegranate peel extract (PEm) rich in EA and punicalagin, vs. EA—50 mg/kg/day of EA | Animal model of EAE in C57BL/6 female mice immunized with MOG35–55. | Both EA and PEm showed comparable efficiencies to reduce the progression of the EAE and to ameliorate the clinical symptoms in mice. Spinal microgliosis and astrocytosis were significantly reduced by the PEm treatment as well as a clear reduction of the CD45 staining in the spinal cord was observed. | [191] |
| UA—2.5 mg/kg, i.p. | Mouse model of traumatic brain injury (TBI). Male C57BL/6J mice were divided into 3 groups: (1) control, (2) TBI + vehicle, (3) TBI + UA. | UA attenuated neuronal apoptosis following TBI (significantly reduced the cleaved caspase-3 expression and increased the Bcl-2 expression vs. TBI+vehicle), reinforced neuronal autophagy (increased the immunopositivity of LC3 and p62, two neuronal autophagy markers), and downregulated (PI3K)/Akt/mTOR (decreased Akt and mTOR expression levels) and Akt/IKK/NFκB signaling pathways (decreased the phosphorylation levels of IκB, IKKα, and NFκB). | [192] |
| Oenothein B—100 or 500 mg/kg/day, p.o. | Male ddY mice received oenothein B once a day for 3 days (from days 1 to 3) or 7 days (from days 1 to 7). | Oenothein B (only 100 mg/kg/day) activated extracellular signal-regulated kinase 2 (ERK2) in the hippocampal region of healthy mice, the ratio pERK2/ERK2 being significantly increased after 7 days of daily treatment. The activation of cAMP response element-binding protein (CREB), reflected by pCREB/CREB ratio, was slightly increased, but without statistical significance (p = 0.0866). | [193] |
| UA (30 μg UA/100 μL PBS, i.p.) | BALB/cJInv female mice were chronically infected with Toxoplasma gondii (T. gondii). UA treatment started 2 days post-infection and continued daily for 5 weeks vs. a control group (injected with vehicle). | All UA-injected mice survived throughout the entire experiment vs. 60% in the control group (40% of the infected control mice died 10-days post-infection). UA treatment inhibited cyst formation in the brain and altered the response of infected mice toward cat odor. | [194] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banc, R.; Rusu, M.E.; Filip, L.; Popa, D.-S. The Impact of Ellagitannins and Their Metabolites through Gut Microbiome on the Gut Health and Brain Wellness within the Gut–Brain Axis. Foods 2023, 12, 270. https://doi.org/10.3390/foods12020270
Banc R, Rusu ME, Filip L, Popa D-S. The Impact of Ellagitannins and Their Metabolites through Gut Microbiome on the Gut Health and Brain Wellness within the Gut–Brain Axis. Foods. 2023; 12(2):270. https://doi.org/10.3390/foods12020270
Chicago/Turabian StyleBanc, Roxana, Marius Emil Rusu, Lorena Filip, and Daniela-Saveta Popa. 2023. "The Impact of Ellagitannins and Their Metabolites through Gut Microbiome on the Gut Health and Brain Wellness within the Gut–Brain Axis" Foods 12, no. 2: 270. https://doi.org/10.3390/foods12020270