
Comparison of Inhibitory Effects of Cinnamic Acid, β-Cyclodextrin, L-Cysteine, and Ascorbic Acid on Soluble and Membrane-Bound Polyphenol Oxidase in Peach Fruit

Abstract
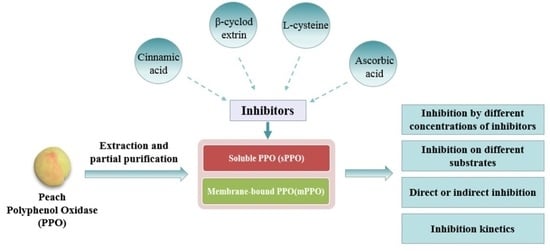
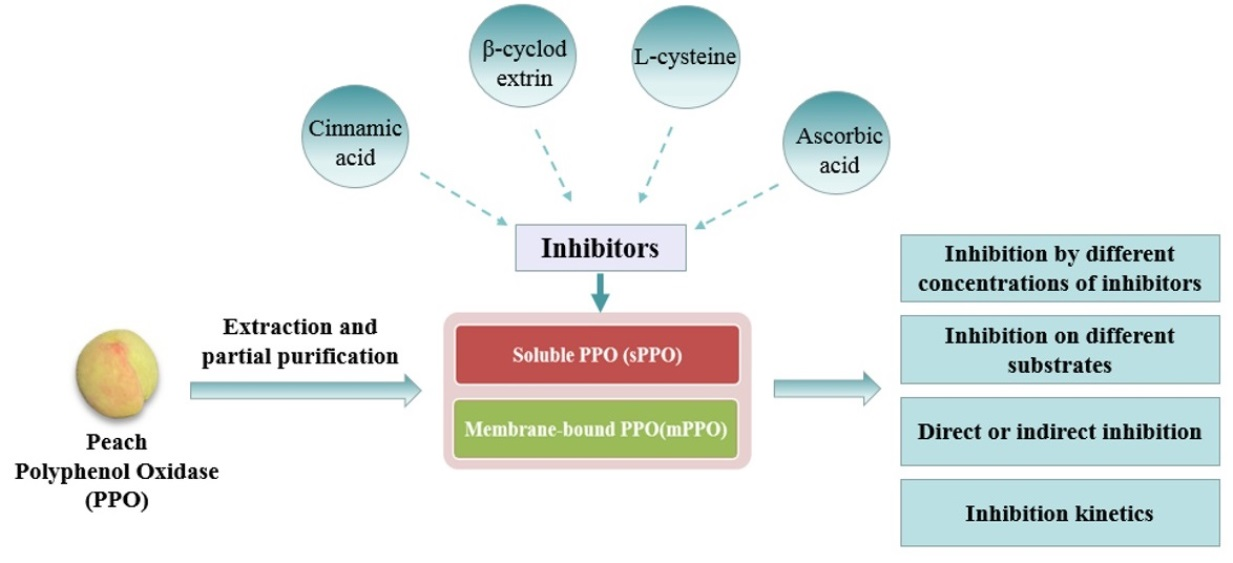
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extraction of sPPO and mPPO
2.3. Partial Purification of sPPO and mPPO
2.4. Quantification of PPO Activity
2.5. Determination of Protein Content
2.6. Effects of Inhibitors on sPPO and mPPO Activities
2.7. Effects of Inhibitors on sPPO- and mPPO-Catalyzed Reactions with Different Substrates
2.8. Direct Effects of Inhibitors on sPPO and mPPO Activities
2.9. Comparison of Inhibition Types of sPPO and mPPO
2.10. Statistical Analysis
3. Results and Discussion
3.1. Comparison of sPPO and mPPO Activities
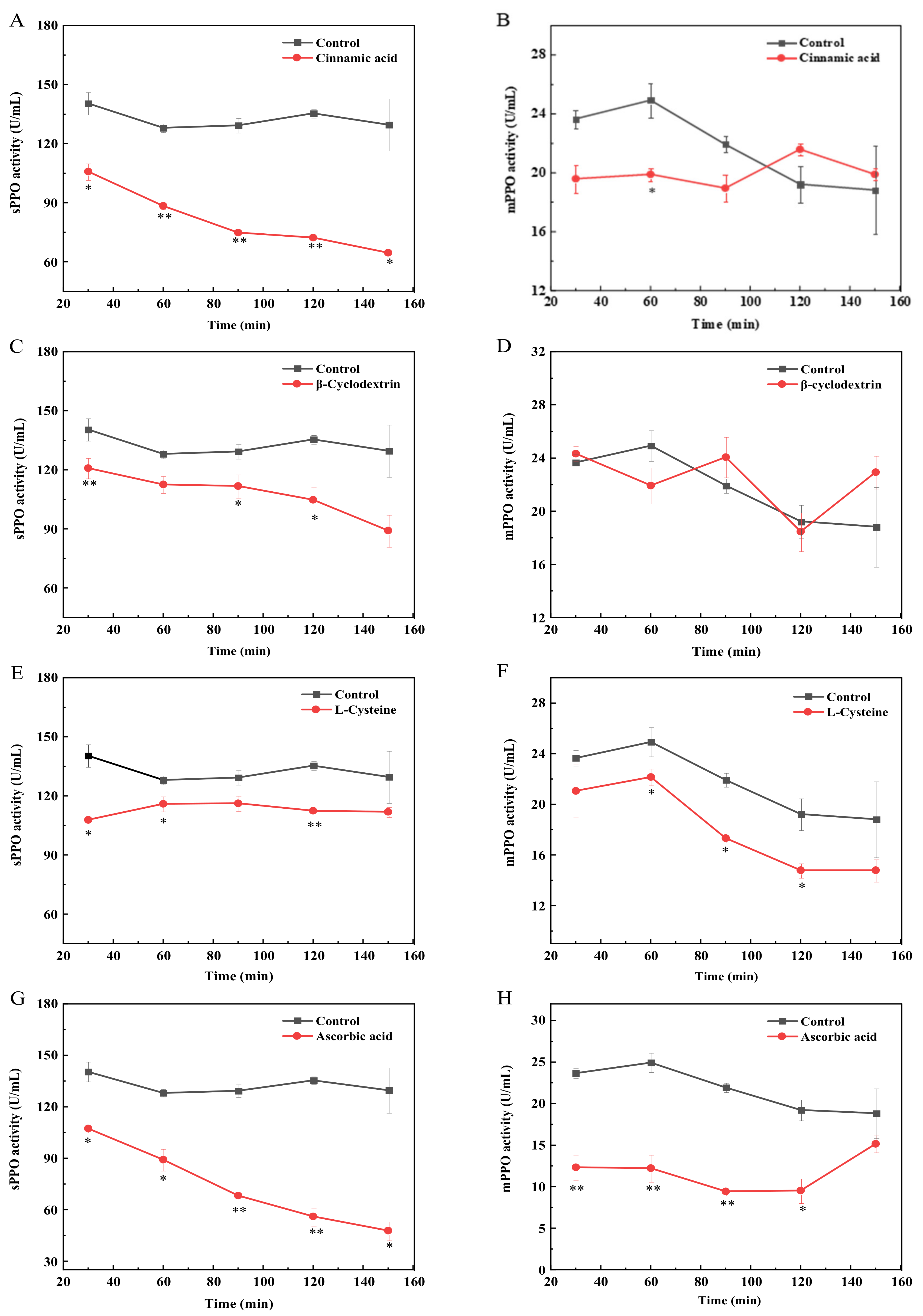
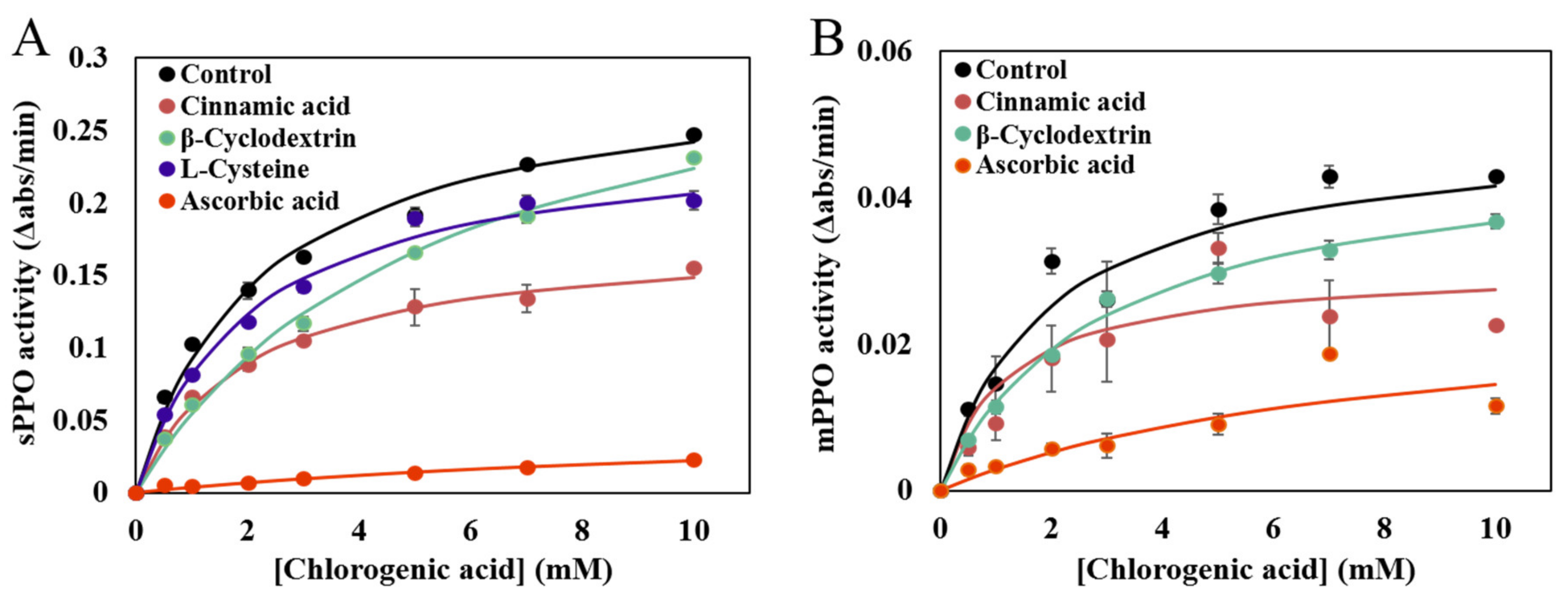
3.2. Effects of Inhibitors on sPPO and mPPO Activities
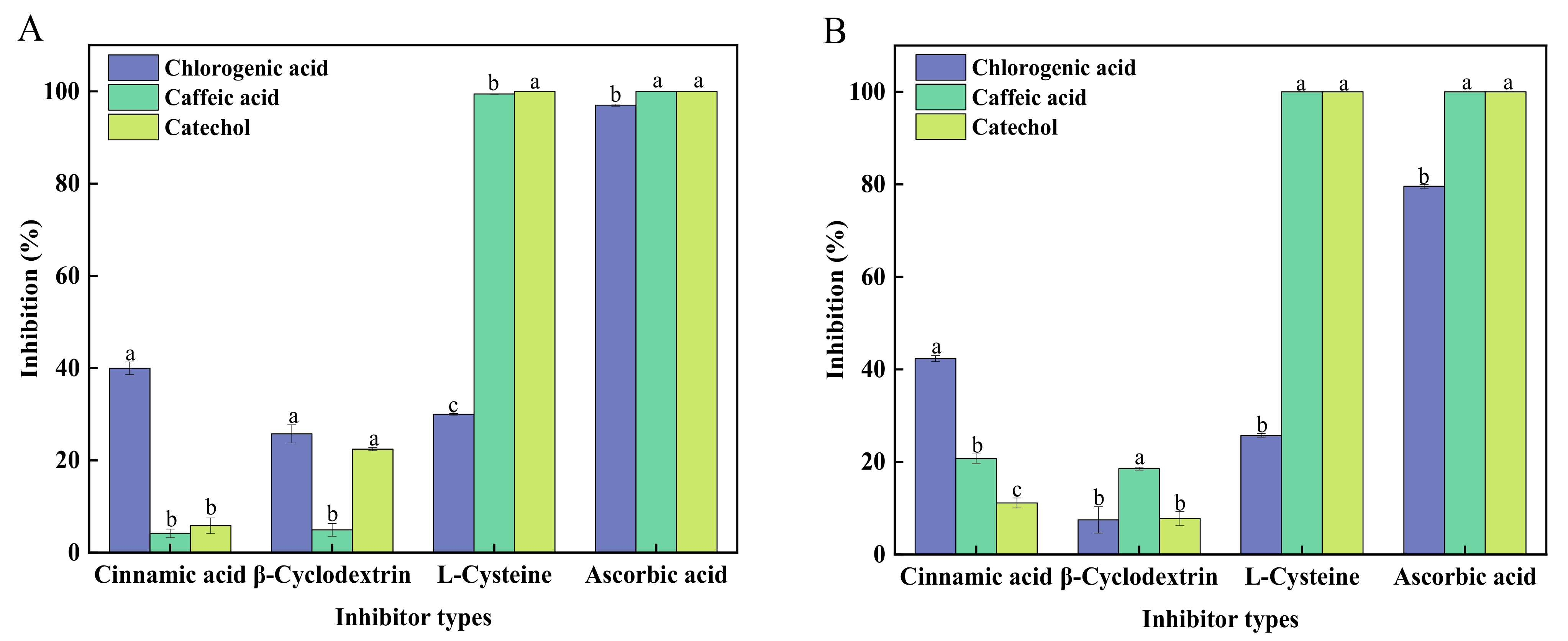
3.3. Effects of Inhibitors on sPPO and mPPO-Catalyzed Reactions with Different Substrates
3.4. Direct Effects of Inhibitors on sPPO and mPPO Activities
3.5. Comparison of Inhibition Types of sPPO and mPPO
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Budak, N.H.; Ozdemir, N.; Gokirmakli, C. The changes of physicochemical properties, antioxidants, organic, and key volatile compounds associated with the flavor of peach (Prunus cerasus L. Batsch) vinegar during the fermentation process. J. Food Biochem. 2021, 46, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bleoanca, I.; Neagu, C.; Turtoi, M.; Borda, D. Mild-thermal and high pressure processing inactivation kinetics of polyphenol oxidase from peach puree. J. Food Process Eng. 2018, 41, e12871. [Google Scholar] [CrossRef]
- Meng, X.H.; Han, J.; Wang, Q.; Tian, S.P. Changes in physiology and quality of peach fruits treated by methyl jasmonate under low temperature stress. Food Chem. 2009, 114, 1028–1035. [Google Scholar] [CrossRef]
- Queiroz, C.; Lopes, M.L.M.; Fialho, E.; Valente-Mesquita, V.L. Polyphenol oxidase: Characteristics and mechanisms of browning control. Food Rev. Int. 2008, 24, 361–375. [Google Scholar] [CrossRef]
- Han, Q.Y.; Liu, F.; Li, M.; Wang, K.L.; Ni, Y.Y. Comparison of biochemical properties of membrane-bound and soluble polyphenol oxidase from Granny Smith apple (Malus x domestica Borkh.). Food Chem. 2019, 289, 657–663. [Google Scholar] [CrossRef]
- Zhou, L.; Liu, W.; Terefe, N.S. The inactivation kinetics of soluble and membrane-bound polyphenol oxidase in pear during thermal and high-pressure processing. Food Bioprocess Technol. 2018, 11, 1039–1049. [Google Scholar] [CrossRef]
- Liu, F.; Zhao, J.H.; Gan, Z.L.; Ni, Y.Y. Comparison of membrane-bound and soluble polyphenol oxidase in Fuji apple (Malus domestica Borkh. cv. Red Fuji). Food Chem. 2015, 173, 86–91. [Google Scholar] [CrossRef]
- Tian, Y.X.; Yan, W.X.; Tang, Y.L.; Yang, R.J.; Zhao, W. Inactivation of membrane-bound and soluble polyphenol oxidases in apple (Malus domestica Borkh) by radio frequency processing for improved juice quality. J. Food Process Eng. 2018, 41, e12923. [Google Scholar] [CrossRef]
- Ma, Y.; Chen, Y.-Y.; Liu, P.; Meng, A.; Deng, L.; Xue, W.; Chen, F.; Che, Z. Comparative study of the biochemical properties of membrane-bound and soluble polyphenol oxidase from Prunus mume. LWT 2022, 171, 114156. [Google Scholar] [CrossRef]
- Sener, A.; Unal, M.U. Purification and characterization of polyphenol oxidase from Akko XIII Loquat (Eriobotrya japonica cv Akko XIII). Food Biotechnol. 2011, 25, 30–42. [Google Scholar] [CrossRef]
- Tao, Y.M.; Yao, L.Y.; Qin, Q.Y.; Shen, W. Purification and characterization of polyphenol oxidase from jackfruit (Artocarpus heterophyllus) bulbs. J. Agric. Food Chem. 2013, 61, 12662–12669. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.; Aguilar-Osorio, G.; Camacho, M.; Basurto, F.; Navarro-Ocana, A. Characterization of polyphenol oxidase from purple sweet potato (Ipomoea batatas L. Lam) and its affinity towards acylated anthocyanins and caffeoylquinic acid derivatives. Food Chem. 2021, 356, 129709. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.Y.; Du, C.; Yu, H.Y.; Zhao, X.Y.; Zhang, X.Y.; Wang, X.Y. Purification and characterization of polyphenol oxidase (PPO) from water yam (Dioscorea alata). CyTA J. Food 2019, 17, 676–684. [Google Scholar] [CrossRef] [Green Version]
- Zaini, N.A.M.; Osman, A.; Hamid, A.A.; Ebrahimpour, A.; Saari, N. Purification and characterization of membrane-bound polyphenoloxidase (mPPO) from Snake fruit [Salacca zalacca (Gaertn.) Voss]. Food Chem. 2013, 136, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Li, F.M. Purification, kinetic parameters, and isoforms of polyphenol oxidase from "Xushu 22" sweet potato skin. J. Food Biochem. 2020, 44, e13452. [Google Scholar] [CrossRef]
- Liu, F.; Zhao, J.H.; Wen, X.; Ni, Y.Y. Purification and structural analysis of membrane-bound polyphenol oxidase from Fuji apple. Food Chem. 2015, 183, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Parrilla, E.; de la Rosa, L.A.; Rodrigo-García, J.; Escobedo-González, R.; Mercado-Mercado, G.; Moyers-Montoya, E.; Vázquez-Flores, A.; González-Aguilar, G.A. Dual effect of β-cyclodextrin (β-CD) on the inhibition of apple polyphenol oxidase by 4-hexylresorcinol (HR) and methyl jasmonate (MJ). Food Chem. 2007, 101, 1346–1356. [Google Scholar] [CrossRef]
- Farouk, B.; Aref, N.; Rachid, C.; Mourad, L.; Emna, K.; Fethi, B.; Rania, B.; Wafa, N.; Kenza, B.; Boumediene, M.; et al. Characterization of three polyphenol oxidase isoforms in royal dates and inhibition of its enzymatic browning reaction by indole-3-acetic acid. Int. J. Biol. Macromol. 2020, 145, 894–903. [Google Scholar] [CrossRef]
- Gacche, R.N.; Shete, A.M.; Dhole, N.A.; Ghole, V.S. Reversible inhibition of polyphenol oxidase from apple using L-cysteine. Indian J. Chem. Technol. 2006, 13, 459–463. [Google Scholar]
- Jiang, S.; Penner, M.H. The nature of β-cyclodextrin inhibition of potato polyphenol oxidase-catalyzed reactions. Food Chem. 2019, 298, 125004. [Google Scholar] [CrossRef]
- Cerit, I.; Pfaff, A.; Ercal, N.; Demirkol, O. Postharvest application of thiol compounds affects surface browning and antioxidant activity of fresh-cut potatoes. J. Food Biochem. 2020, 44, e13378. [Google Scholar] [CrossRef] [PubMed]
- Sikora, M.; Swieca, M. Effect of ascorbic acid postharvest treatment on enzymatic browning, phenolics and antioxidant capacity of stored mung bean sprouts. Food Chem. 2018, 239, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Adeseko, C.J.; Sanni, D.M.; Salawu, S.O.; Kade, I.J.; Bamidele, S.O.; Lawal, O.T. Purification and biochemical characterization of polyphenol oxidase of African bush mango (Irvingia gabonensis) fruit peel. Biocatal. Agric. Biotechnol. 2021, 36, 102119. [Google Scholar] [CrossRef]
- Sharma, S.; Rao, T.V.R. Xanthan gum based edible coating enriched with cinnamic acid prevents browning and extends the shelf-life of fresh-cut pears. LWT Food Sci. Technol. 2015, 62, 791–800. [Google Scholar] [CrossRef]
- Campo, A.F.; Sancho, M.I.; Melo, G.; Davila, Y.A.; Gasull, E. In vitro and in vivo inhibition of Hass avocado polyphenol oxidase enzymatic browning by paeonol, β-cyclodextrin, and paeonol:beta-cyclodextrin inclusion complex. J. Biosci. Bioeng. 2019, 127, 703–709. [Google Scholar] [CrossRef]
- Cabanes, J.; Escribano, J.; Gandia-Herrero, F.; Garcia-Carmona, F.; Jimenez-Atienzar, M. Partial purification of latent polyphenol oxidase from peach (Prunus persica L. Cv. Catherina). Molecular properties and kinetic characterization of soluble and membrane-bound forms. J. Agric. Food Chem. 2007, 55, 10446–10451. [Google Scholar] [CrossRef]
- Guo, S.W.; Zhang, L.; Zhang, L.; Zhao, M.N.; Meng, X.H. Inhibition kinetics of oligochitosan as an uncompetitive inhibitor on fuji apple polyphenol oxidase. J. Food Biochem. 2018, 42, e12585. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Khettal, B.; Kadri, N.; Tighilet, K.; Adjebli, A.; Dahmoune, F.; Maiza-Benabdeslam, F. Phenolic compounds from Citrus leaves: Antioxidant activity and enzymatic browning inhibition. J. Complement. Integr. Med. 2017, 14, 20160030. [Google Scholar] [CrossRef]
- Wang, F.; Zhou, H.; Cheng, F.; Niu, H.; Yuan, L.; Yi, J.; Zhou, L. Comparison of the characterization and the temperature/pressure stability of soluble and membrane-bound polyphenol oxidase from ’Lijiang’ snow peach. LWT Food Sci. Technol. 2021, 146, 111401. [Google Scholar] [CrossRef]
- Xiao, Y.H.; Zhang, J.L.; Jiang, Y.Y.; Yuan, Y.; Xie, J.; He, J.M.; Wang, B. Cinnamic acid treatment reduces the surface browning of fresh-cut taro. Sci. Hortic. 2022, 291, 110613. [Google Scholar] [CrossRef]
- GeÇEr, M.K. Biochemical content in fruits of peach and nectarine cultivars. Turk. J. Agric. For. 2020, 44, 500–505. [Google Scholar] [CrossRef]
- Bento, C.; Gonçalves, A.C.; Silva, B.; Silva, L.R. Peach (Prunus Persica): Phytochemicals and health benefits. Food Rev. Int. 2020, 38, 1703–1734. [Google Scholar] [CrossRef]
- Ionita, E.; Gurgu, L.; Aprodu, I.; Stanciuc, N.; Dalmadi, I.; Bahrim, G.; Rapeanu, G. Characterization, purification, and temperature/pressure stability of polyphenol oxidase extracted from plums (Prunus domestica). Process Biochem. 2017, 56, 177–185. [Google Scholar] [CrossRef]
- Bravo, K.; Osorio, E. Characterization of polyphenol oxidase from Cape gooseberry (Physalis peruviana L.) fruit. Food Chem. 2016, 197, 185–190. [Google Scholar] [CrossRef]
- Jang, J.-H.; Moon, K.-D. Inhibition of polyphenol oxidase and peroxidase activities on fresh-cut apple by simultaneous treatment of ultrasound and ascorbic acid. Food Chem. 2011, 124, 444–449. [Google Scholar] [CrossRef]
- Derardja, A.E.; Pretzler, M.; Kampatsikas, I.; Barkat, M.; Rompel, A. Inhibition of apricot polyphenol oxidase by combinations of plant proteases and ascorbic acid. Food Chem. X 2019, 4, 100053. [Google Scholar] [CrossRef]
- Arias, E.; Gonzalez, J.; Peiro, J.M.; Oria, R.; Lopez-Buesa, P. Browning prevention by ascorbic acid and 4-hexylresorcinol: Different mechanisms of action on polyphenol oxidase in the presence and in the absence of substrates. J. Food Sci. 2007, 72, C464–C470. [Google Scholar] [CrossRef]
- Van Bree, I.; Baetens, J.M.; Samapundo, S.; Devlieghere, F.; Laleman, R.; Vandekinderen, I.; Noseda, B.; Xhaferi, R.; De Baets, B.; De Meulenaer, B. Modelling the degradation kinetics of vitamin C in fruit juice in relation to the initial headspace oxygen concentration. Food Chem. 2012, 134, 207–214. [Google Scholar] [CrossRef]
- Wu, X.T.; Guo, X.N.; Zhu, K.X. Inhibition of L-cysteine on the browning of fresh wet noodles. Foods 2021, 10, 1156. [Google Scholar] [CrossRef]
- Ding, C.K.; Chachin, K.; Ueda, Y.; Wang, C.Y. Inhibition of loquat enzymatic browning by sulfhydryl compounds. Food Chem. 2002, 76, 213–218. [Google Scholar] [CrossRef]
- Zhou, L.; Liu, W.; Xiong, Z.Q.; Zou, L.Q.; Chen, J.; Liu, J.P.; Zhong, J.Z. Different modes of inhibition for organic acids on polyphenoloxidase. Food Chem. 2016, 199, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.R.; Xiao, Y.J.; Meng, X.H.; Liu, B.J. Full inhibition of Whangkeumbae pear polyphenol oxidase enzymatic browning reaction by L-cysteine. Food Chem. 2018, 266, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yan, S.; Wang, Q.; Hussain, S.; Li, J. Inhibition of ascorbic acid on Lotus Rhizome polyphenol oxidase: Inhibition kinetics and computational simulation. Food Sci. Qual. Manag. 2014, 34, 103–112. Available online: https://www.researchgate.net/publication/270274443 (accessed on 6 May 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Stages | Volume (mL) | Total Activity (U) | Protein Content (mg) | Specific Activity (U/mg) | Yield (%) | Purification Fold |
|---|---|---|---|---|---|---|
| sPPO | ||||||
| Crude extract | 264 | 2640 | 36 | 74 | 100 | 1 |
| (NH4)2SO4 precipitation | 28 | 912 | 10 | 95 | 35 | 1.3 |
| mPPO | ||||||
| Crude extract | 20 | 179 | 7 | 24 | 100 | 1 |
| (NH4)2SO4 precipitation | 15 | 139 | 5 | 30 | 78 | 1.2 |
| Form | Inhibitors | Km (mM) | Vmax (Δabs/min) |
|---|---|---|---|
| sPPO | None | 2.04 ± 0.23 c | 0.29 ± 0.01 b |
| Cinnamic acid | 1.90 ± 0.10 c | 0.18 ± 0.00 d | |
| β-Cyclodextrin | 5.40 ± 0.31 b | 0.35 ± 0.00 a | |
| L-Cysteine | 2.12 ± 0.13 c | 0.25 ± 0.01 c | |
| Ascorbic acid | 11.18 ± 1.84 a | 0.05 ± 0.01 e | |
| mPPO | None | 2.14 ± 0.30 b | 0.05 ± 0.00 a |
| Cinnamic acid | 1.72 ± 0.76 b | 0.03 ± 0.01 b | |
| β-Cyclodextrin | 2.88 ± 0.07 b | 0.05 ± 0.00 a | |
| L-Cysteine | ** | ** | |
| Ascorbic acid | 5.99 ± 1.81 a | 0.02 ± 0.00 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, S.; Jiang, S.; Chen, Y.; Wei, Y.; Shao, X. Comparison of Inhibitory Effects of Cinnamic Acid, β-Cyclodextrin, L-Cysteine, and Ascorbic Acid on Soluble and Membrane-Bound Polyphenol Oxidase in Peach Fruit. Foods 2023, 12, 167. https://doi.org/10.3390/foods12010167
Jia S, Jiang S, Chen Y, Wei Y, Shao X. Comparison of Inhibitory Effects of Cinnamic Acid, β-Cyclodextrin, L-Cysteine, and Ascorbic Acid on Soluble and Membrane-Bound Polyphenol Oxidase in Peach Fruit. Foods. 2023; 12(1):167. https://doi.org/10.3390/foods12010167
Chicago/Turabian StyleJia, Shuang, Shu Jiang, Yi Chen, Yingying Wei, and Xingfeng Shao. 2023. "Comparison of Inhibitory Effects of Cinnamic Acid, β-Cyclodextrin, L-Cysteine, and Ascorbic Acid on Soluble and Membrane-Bound Polyphenol Oxidase in Peach Fruit" Foods 12, no. 1: 167. https://doi.org/10.3390/foods12010167