
Microbial Communities and Flavor Compounds during the Fermentation of Traditional Hong Qu Glutinous Rice Wine
by
 ,
,
Anqi Liu
1,2 ,
,
Xu Yang
1,
Quanyou Guo
1,*,
Baoguo Li
2,
Yao Zheng
1,
Yuzhuo Shi
1,2 and
Lin Zhu
1,2 1
East China Sea Fishery Research Institute, Chinese Academy of Fishery Sciences, Shanghai 200090, China
2
School of Health Science and Engineering, University of Shanghai for Science and Technology, Shanghai 200093, China
*
Author to whom correspondence should be addressed.
Foods 2022, 11(8), 1097; https://doi.org/10.3390/foods11081097
Submission received: 17 March 2022
/
Revised: 7 April 2022
/
Accepted: 8 April 2022
/
Published: 11 April 2022
(This article belongs to the Topic Alcoholic Beverage Research (Agriculture, Processing, Business and Circular Economy, Climate Effect))
Abstract
:As a traditional Chinese rice wine, Hong Qu glutinous rice wine (HQW) is popular among consumers due to its unique flavor. However, its quality changes during fermentation, and the potential relationships between flavor and microbes have not been systematically researched. In this work, physicochemical properties (pH, total sugar, alcohol, amino acid nitrogen), flavor compounds (organic acids, free amino acids, and volatile compounds), and microbial communities were investigated. The results revealed that Pantoea, Lactiplantibacillus, Lactobacillus, Leuconostoc, and Weissella predominated the bacterial genera, and Monascus was the predominant fungal genus. Organic acids, free amino acids, and key volatile compounds (esters and alcohols) significantly increased during fermentation. The correlations analysis showed that Lactiplantibacillus was closely associated with flavor compounds formation. This study deepens our understanding of the roles of microorganisms in flavor formation on traditional HQW fermentation.
1. Introduction
Hong Qu glutinous rice wine (HQW), a famous traditional Chinese rice wine invented in the Song Dynasty, has a red color, a special flavor, and a sourish taste, which is believed to improve digestion and blood circulation. Traditional HQW is brewed by open fermentation in pottery jars, and its process includes macerating and steaming rice, saccharification, fermentation, pressing, and storage. This traditional brewing process cannot meet the demand of consumers, so many companies expand the production of HQW through industrialized methods. However, there is a significant difference in flavor and quality between industrial and traditional HQW. Thus, it is necessary to conduct a comprehensive analysis of the flavor changes of traditional HQW during the fermentation process to provide a scientific reference for industrial production.
The complex flavor of HQW consists mainly of organic acids, free amino acids, and volatile flavor compounds, which are obtained from the raw materials and fermentation. More precisely, fermentation increases the concentration of organic acids, free amino acids, and volatile compounds, and is the main contributor to the formation of key flavors in rice wine [1]. During the saccharification stage, filamentous fungi promote the hydrolysis of starch to produce sugar, leading to a more rapid flavor formation in rice wine [2]. Microorganisms use fermentable sugars to produce large amounts of organic acids by carbohydrate metabolism [3]. Organic acids provide the main sour taste to the rice wine and improve the color of rice wine [4]. When some microorganisms undergo autolysis, proteases release to promote the production of free amino acids. The free amino acids contribute to the complex taste of the food including umami, sweet, bitter, and astringency [5]. As precursors to volatile compounds, organic acids and free amino acids contribute to the formation of aromas in rice wine due to the metabolic action of varieties bacteria, fungi, and yeasts [6]. In complex fermentation environments, different microflora may produce different flavors for rice wine. During the fermentation of traditional Guizhou black Huangjiu, Gluconobacter, Bacillus, Streptococcus, Lactobacillus, Lactococcus, Pediococcus, and Leuconostoc have been shown to influence the change in metabolites (esters, alcohols, acids, and alkanes) [7]. During the fermentation of traditional Wuyi Huangjiu, Lactobacillus, Pichia mississippiensis, and Saccharomyces cerevisiae were positively correlated with ethyl ester formation, while Bacillus myloliquefaciens and Bacillus subtilis were negatively correlated with volatile acids and esters [8]. Thus, it is necessary to analyze the flavor compounds and microbial communities of HQW during fermentation, which helps to elucidate which microorganisms play the different roles in the flavor formation of HQW.
As an advanced second-generation sequencing technology, high-throughput sequencing (HTS) overcomes the limitations of traditional molecular methods. Previous studies have reported that HTS could reveal the complex microbial community more precisely and comprehensively [9]. Thus, HTS has been widely used for comprehensive analysis of microorganisms in various fermented foods, such as black glutinous rice wine [10], Chinese rice wine [11], highland barley wine [12], and fermented fish [13]. The detection means for flavor compounds include high-performance liquid chromatography (HPLC), liquid chromatography–mass spectrometry (LC–MS), gas chromatography–mass spectrometry (GC–MS), and gas chromatography–ion mobility spectrometry (GC–IMS). As an emerging technique, GC–IMS has the advantages of a high sensitivity, a low detection limit for the ppbv level, no requirement for the pretreatment of samples, secondary separation, an improved separation effect, three-dimensional visual spectrum formation, and a direct comparison between spectra with plugins. Yang used GC–IMS to analyze volatile flavor compounds across different production stages of fermented soybean whey tofu (FSWT), and found that ketones, alcohols, and esters contribute to the unique flavor of the final FSWT product [14].
This study aims to detect the flavor compounds of HQW and analyze the microbial dynamics including fungal and bacterial communities during fermentation. It is hypothesized that physiochemical properties could reflect the degree of traditional HQW fermentation, and there is a clear link between the microbial community and various flavor compounds. Furthermore, results from the current study would provide new insights for improving the flavor and quality of HQW by screening core microbial communities.
2. Materials and Methods
2.1. Sample Preparation and Collection
HQW was brewed by traditional brewing methods. An amount of 4 kg of glutinous rice was washed and soaked in water at room temperature (around 15 °C) for 10 h, drained, and then steamed in 100 °C for 30 min. After, the steamed rice was cooled to 35 °C in a ventilated and cool environment. The cooled steamed glutinous rice was mixed with 6 L of cool sterile water and 400 g of Hong Qu fermentation starters in a clay vat and then fermented at 20 °C for 30 days.
At 0, 6, 12, 18, 24, and 30 days of fermentation, 10 g of HQW was randomly sampled for microbial diversity testing. At the same time, 500 mL of fermentation mash was collected for analysis of physicochemical properties, organic acids, free amino acids, and volatile flavor compounds (VFCs).
2.2. Physiochemical Properties Determination
The pH of the sample was tested by a pH meter. The alcohol (percentage of alcohol by volume), total sugar, and amino-acid nitrogen were measured by an alcohol meter and titration method according to Chinese national standard GB/T13662-2018.
2.3. Total DNA Extraction and PCR Amplification
Total genomic DNA samples were extracted using the OMEGA Soil DNA Kit (D5625-01) (Omega Bio-Tek, Norcross, GA, USA). The quantity and quality of extracted DNAs were measured using a NanoDrop ND-1000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) and agarose gel electrophoresis, respectively. The V3-V4 region of the bacteria 16S rRNA gene was amplified with primers 338F (5′-ACTCCTACGGGAGGCAGCA-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′), and the ITS1 region of the fungi was amplified with the forward primer ITS1F (5′-CTTGGTCATTTAGAGGAAGTAA-3′) and the reverse primer ITS1R (5′-GCTGCGTTCTTCATCGATGC-3′).
Sample-specific 7-bp barcodes were incorporated into the primers for multiplex sequencing. The PCR components contained 5 μL of buffer (5×), 0.25 μL of Fast pfu DNA Polymerase (5 U/μL), 2 μL (2.5 mM) of dNTPs, 1 μL (10 μM) of each Forward and Reverse primer, 1 μL of DNA Template, and 14.75 μL of ddH2O. Thermal cycling consisted of initial denaturation at 98 °C for 5 min, followed by 25 cycles consisting of denaturation at 98 °C for 30 s, annealing at 53 °C for 30 s, and extension at 72 °C for 45 s, with a final extension of 5 min at 72 °C. PCR amplicons were purified with Vazyme VAHTSTM DNA Clean Beads (Vazyme, Nanjing, China) and quantified using the Quant-iT PicoGreen dsDNA Assay Kit (Invitrogen, Carlsbad, CA, USA). After the individual quantification step, amplicons were pooled in equal amounts, and pair-end 2 × 250 bp sequencing was performed using the Illlumina NovaSeq platform with the NovaSeq 6000 SP Reagent Kit at Shanghai Personal Biotechnology Co., Ltd. (Shanghai, China)
2.4. Sequence Analysis
Microbiome bioinformatics were performed with QIIME2 2022.2 with slight modification according to the official tutorials (https://docs.qiime2.org/2022.2/tutorials/ (accessed on 5 April 2022)). Briefly, raw sequence data were demultiplexed using the demux plugin following by primers cutting with the cutadapt plugin. Sequences were then quality-filtered, denoised, merged, and the chimera removed using the DADA2 plugin. Nonsingleton amplicon sequence variants (ASVs) were aligned with mafft and used to construct a phylogeny with fasttree2. Taxonomy was assigned to ASVs using the classify-sklearn naїve Bayes taxonomy classifier in the feature-classifier plugin against the SILVA v138.1 (bacteria) and UNITE v8.0 (fungi) Database.
2.5. Analysis of VFCs
The VFCs of traditional fermented HQW were detected by using Yang’s modified method [14]. Headspace-solid-phase microextraction (HS-SPME) coupled with gas chromatography–ion mobility spectrometry (GC–IMS) was used to identify VFCs, and an Agilent 490 gas chromatograph (Agilent Technologies, Palo Alto, CA, USA) and IMS instrument (FlavourSpec®, Gesellschaft für Analytische Sensorsysteme mbH, Dortmund, Germany) equipped with an automatic sampling device were used to detect the flavor substances in samples collected from different production stages.
A sample of 1 g was added to the 20 mL headspace vial of the automatic sampler. The incubation temperature of the automatic sampler was set at 60 °C, and the incubation time was 10 min. The headspace injection method was selected, with oscillation heating at 500 rpm, an injection temperature of 85 °C, and an injection volume of 100 μL. The sample entered the MXT-WAX capillary column (30 m × 0.53 mm, 1 μm) as a gas. The chromatographic column temperature was set at 60 °C, the carrier gas was N2 (purity ≥ 99.999%), and the carrier gas flow rate was programmed as follows: the initial flow rate reached 2.0 mL/min within 2 min; the internal linearity was increased to 10.0 mL/min within 8 min, followed by 100.0 mL/min within 10 min and then to 150.0 mL/min within 10 min; the total running time was 30 min.
2.6. Analysis of Free Amino Acids
Free amino acids in standards and samples were detected by high-performance liquid chromatography (HPLC) (Agilent 1260, Agilent Technologies, Palo Alto, CA, USA) equipped with a C18 column (Zorbax Eclipse-AAA, 150 mm × 4.6 mm, 3.5 μm) and a variable-wavelength detector (VWD). Samples of 40 mL were centrifuged at 10,000 rpm for 10 min at 4 °C and were then filtered with a 0.45 μm PTFE membrane. An amount of 1 μL was injected from the filtered samples and the flow rate was maintained at 2.0 mL/min. Two amino acid derivatives were used, including o-phthalaldehyde (OPA) and 9-fluorenylmethyl chloroformate (FMOC). The column temperature was maintained at 40 °C with the detection wavelengths of 262 nm and 338 nm. Eluent A: 0.04 mol/L sodium dihydrogen phosphate (pH is adjusted to 7.8 ± 0.01 by 10% NaOH). Eluent B: methanol, acetonitrile, and water (45:45:10, v/v/v). Amino acid species was identified by retention times and reference standards, and the content was determined by standard curves.
2.7. Analysis of Organic Acids
The organic acids of traditional fermented HQW were detected by using Huang’s modified method [8]. Organic acids in standards and samples were detected by HPLC (Agilent 1260, Agilent Technologies, Palo Alto, CA, USA) equipped with a C18 column (Zorbax SB-AQ, 250 mm × 4.6 mm, 5 μm) and a variable-wavelength detector (VWD). Samples of 40 mL were centrifuged at 10,000 rpm for 10 min at 4 °C and were then filtered with a 0.22 μm PTFE membrane. An amount of 1 μL was injected from the filtered samples and the flow rate was maintained at 0.8 mL/min. The column temperature was 25 °C, and the detector wavelength was 210 nm. Only one eluent was used as the mobile phase: eluent A: 0.02 mol/L sodium dihydrogen phosphate and acetonitrile (99:1, v/v, pH is adjusted to 2 ± 0.01 by 85% H3PO4).
2.8. Statistical Analysis
All the analyses were conducted in triplicate, and then averages and standard deviations were calculated using SPSS 17.0 software (SPSS Inc., Chicago, IL, USA). Analysis of variance (ANOVA) and t-tests were used to evaluate the differences between glutinous rice wines at different fermentation stages. The VFCs were analyzed by principal component analysis (PCA) using SIMCA-14.1 software (UMETRICS, Malmö, Sweden). Correlation coefficients between microbial genera and flavor compounds were calculated using R software (v4.0.4, R Foundation for Statistical Computing, Vienna, Austria), and p values were multiply corrected by fdr.
3. Results and Discussion
3.1. Analysis of the Composition of Microbial Communities
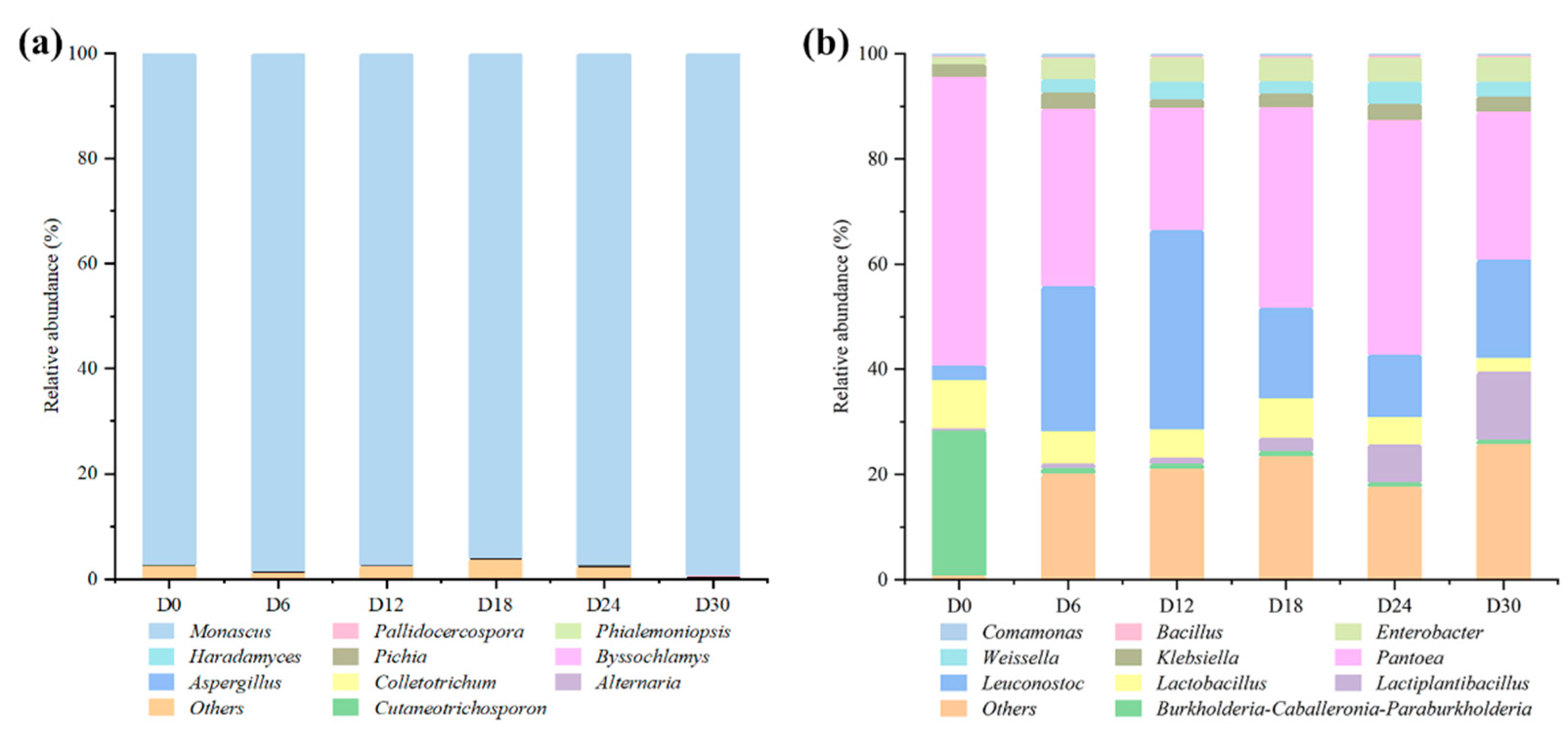
The microbial genera dynamics of HQW was analyzed during fermentation. As shown in Figure 1a, Monascus was the dominant fungi genus, with a relative abundance greater than 97% during the fermentation, and as high as 99.5% at the end. Previous studies have shown similar results that Monascus was detected as the dominant fungi genus in both Wuyi and Gutian HQW [15,16]. Monascus is a type of filamentous fungi and was first screened in red mold rice and characterized by van Tieghem (1884). Monascus can secrete a variety of hydrolytic enzymes including α-amylase, glucoamylase, lipase, and protease, playing an extremely important role in the glycation phase of the fermentation process [2]. As a characteristic species of East Asia, Monascus has received worldwide attention for its abundant beneficial flavor compounds, such as monacolin and pigments [15]. Besides Monascus, the relative abundance of the other fungi genera was below 0.1% during the fermentation, such as Pallidocercospora, Phialemoniopsis, Haradamyces, and Pichia. The alcohol and low pH in rice wine will inhibit the growth of non-acid and alcohol-tolerant microorganisms [1].
The bacterial genera dynamics during fermentation is shown in Figure 1b. The predominant bacteria include Pantoea, Leuconostoc, Lactiplantibacillus, Lactobacillus, and Weissella. At the beginning, Pantoea was the dominant bacterium with an abundance of 54.8%; then, it significantly decreased to 23.2% on day 12, and finally reached 28.1% after a brief fluctuation. Pantoea is derived from HQ, which is a common endophyte found in rice [10]. Meanwhile, Leuconostoc increased from an initial abundance of 2.8% to 38.0% on day 12, and then fluctuated to a final abundance of 18.7%. Lactiplantibacillus continued to increase from an initial abundance of 0.1% to 12.9%, and the abundance of Lactobacillus increased from 0.02% to 2.7% during the fermentation. In recent studies, the genus Lactobacillus has been reclassified into 25 genera including Lactiplantibacillus and the emended genus Lactobacillus [17]. Due to the fact that Lactobacillus can tolerate low pH and inhibit the growth of other microorganisms, they are usually detected as the dominant genus in most fermented foods [8]. Weissella was not identified at the beginning of the fermentation, and its abundance was 2.5% on day 6, indicating that it might be brought in from the environment. In Liu’s study, Weissella was shown to promote the formation of acetic acid during the brewing process of soy sauce [18]. During the fermentation process, the proliferation of lactic acid bacteria (LAB) can produce organic acids and bacteriocins, reducing the diversity of microorganisms and inhibiting the growth of some pathogenic bacteria, which make a great contribution to the flavor formation and food safety of wine [13]. Furthermore, their parthenogenic anaerobic growth characteristics and bacteriocin production make lactic acid bacteria dominant in most fermented foods [3]. In addition, Klebsiella, Enterobacter, and Bacillus were also identified with low abundance during the fermentation of HQW. As a ubiquitous Gram-positive bacteria in the environment, Bacillus of rice wine is obtained from fermentation starters or the environment, and contribute to the formation of the rice wine flavor by secreting hydrolytic enzymes [19]. However, the growth of Bacillus is inhibited due to the increase in alcohol concentrations during the fermentation of rice wine [20]. Klebsiella is a common endophyte found in cereals. Along with Enterobacter, they were identified in corn silage, where they can carry out carbohydrate fermentation to promote lactic and acetic acid formation and pH reduction [21]. They were also identified with low abundance during the brewing of some grain wines [22].
3.2. Physicochemical Properties and Flavor Compounds
3.2.1. Physicochemical Properties Analysis
The physicochemical properties (pH, total sugars, alcohol, amino acid nitrogen) during HQW fermentation are shown in Table 1. These indicators are typically used to determine the fermentation status of rice wine. After 6 days of fermentation, the pH decreased significantly from an initial 5.83 ± 0.01 to 3.86 ± 0.02 (p < 0.05), and stabilized during the subsequent fermentation stages, with a final pH of 3.84 ± 0.01. On the other hand, total sugars significantly increased (p < 0.05) on day 6 and then significantly decreased (p < 0.05) on day 12, gradually decreasing to a final concentration of 1.56 ± 0.08 g/L. Meanwhile, alcohol and amino acid nitrogen kept increasing during the whole fermentation process, similar to the results of another study on the fermentation of black glutinous rice wine [10]. This indicates that the first 6 days were the saccharification stage of fermentation, in which amylase secreted by some filamentous fungi hydrolyzed the starch in glutinous rice to produce large amounts of glucose [23]. Ethanol, organic acids, and other metabolites such as amino acids were synthesized, and the decrease in pH value was obtained.
3.2.2. Changes in the Free Amino Acid Fraction of HQW during Fermentation
As shown in Table 2, the 17 amino acids were identified during fermentation and classified into umami, sweet, bitter, and astringent amino acids according to their taste presentation characteristics. Among them, Trp, Arg, Ile, Glu, and Pro were the main components of free amino acids. During the fermentation process, free amino acids are produced mainly through the enzymatic degradation of proteins in the raw material by microorganisms secreting proteases, as well as by intracellular proteases after autolysis of microorganisms [5]. The most significant increase in four groups of amino acids was observed on day 12 compared to the other stages, with almost 3 times more than the previous stage. The total concentration of four groups of amino acids all continued to increase during fermentation, while the bitter amino acids were the most abundant. Previous studies have indicated that high temperatures increase the content of bitter amino acids during the fermentation of Chinese rice wine [16]. Six of the eight bitter amino acids are essential to the human diet, except for Arg and His. Free amino acids bring umami, sweet, bitter, and astringent flavors to the food, and are also important nitrogen sources for microbial growth and proliferation and can produce many secondary metabolites such as higher alcohols [24]. In this biotransformation process, there is a corresponding relationship between amino acids and higher alcohols, such as isoleucine to 2-methylbutanol, valine to 2-methylpropanol, and leucine to 3-methylbutanol [25].
3.2.3. Changes in Organic Acids
The acidity of HQW results from organic acids, which affect the organoleptic quality of the wine dramatically, and can inhibit the growth of microbes as well as promote the production of esters. They mainly come from chemical synthesis by microbial metabolism such as alcoholic fermentation, malolactic fermentation, and oxidation of the ethanol [26].
Table 3 shows the changes in six organic acids identified during the fermentation of HQW, including lactic acid, succinic acid, citric acid, malic acid, tartaric acid, and oxalic acid. During the fermentation process, the total concentration of organic acids increased continuously. The most abundant organic acid was lactic acid, followed by succinic acid, citric acid, and malic acid. After 6 days of fermentation, lactic acid increased significantly from an initial level of 27.1 ± 0.6 mg/L to 4027.6 ± 6.5 mg/L (p < 0.05), then gradually increasing in the following days to a final concentration of 8073.9 ± 46.9 mg/L, accounting for 55.29% of the total organic acids. Rice and grains are the raw materials for HQW, and the rich carbohydrates make lactic acid the main metabolite of LAB [3]. This also involves malolactic fermentation (MLF) under the action of LAB, and MLF is carried out just after the inoculation with a starter culture, the decarboxylation of malic acid into lactic acid and CO2 [27]. Thus, the concentration of lactic acid maintains a continuous increase during the fermentation of traditional rice wine. Tartaric acid had the highest abundance at the beginning of fermentation, increased on day 12, and then gradually decreased to a final concentration of 953.9 ± 14.1 mg/L. The initial concentration of malic acid was lower than that of tartaric acid, and gradually increased after 6 days to a final concentration of 1223.2 ± 38.4 mg/L. As an important substrate for MLF, malic acid is always detected in rice wine with a moderate level [11]. The lowest organic acid content was oxalic acid, which fluctuated and remained below 70 mg/L during fermentation. Generally, organic acids bring different acidity perceptions to form the harmonious sour taste: lactic acid gives a soft sour taste, slightly with frankincense to rice wine, citric acid tastes fresh and cool, succinic acid tastes salty and bitter, malic acid represent a bit of sharp sour, and tartaric acid is bitter and tough sour [12].
3.2.4. Volatile Flavor Compounds Analysis
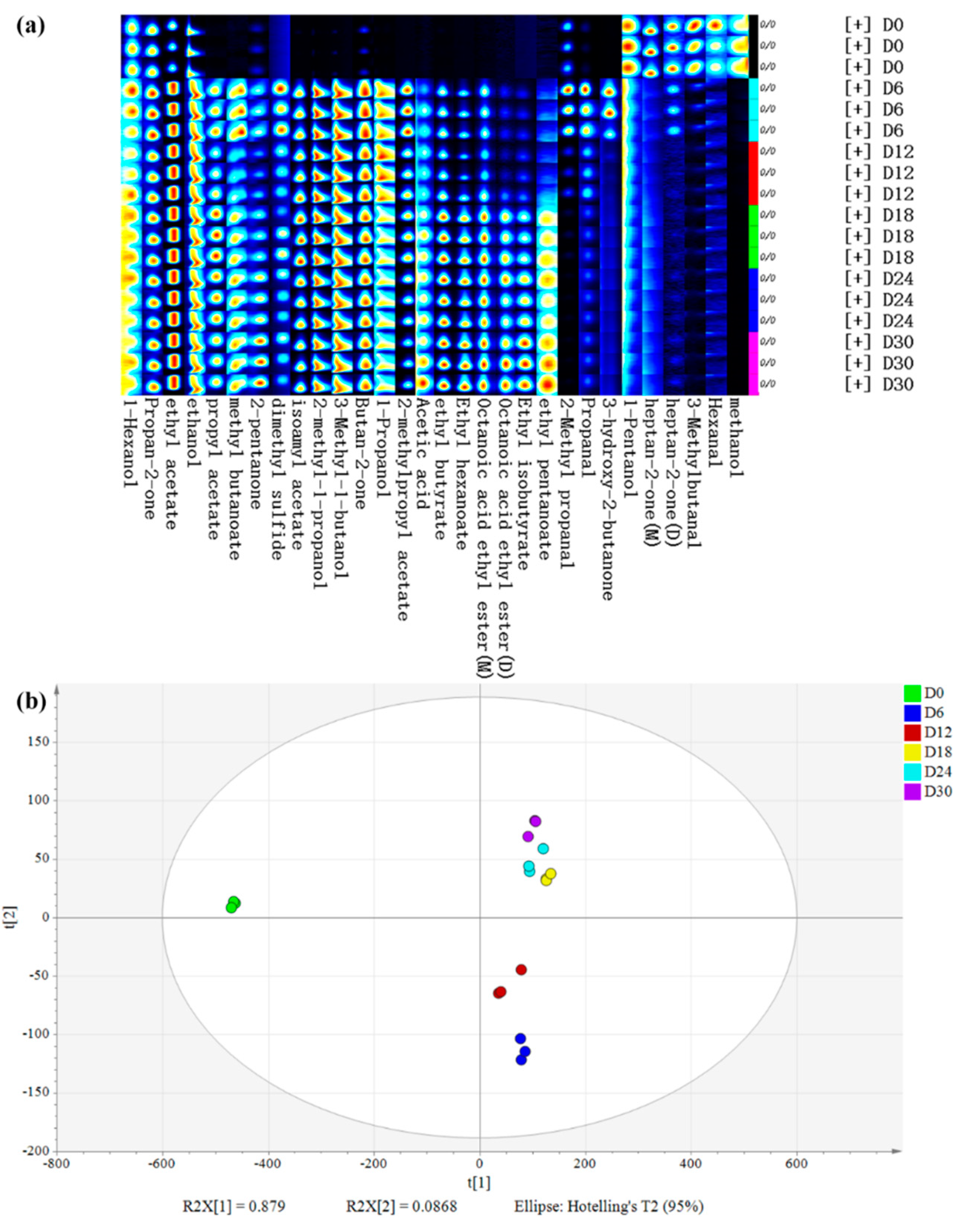
The volatile flavor compounds (VFCs) were analyzed during the fermentation of HQW using headspace–gas chromatography–ion mobility spectrometry (HS–GC–IMS). A total of 29 VFCs were identified (Figure 2a), including 11 esters, 7 alcohols, 5 ketones, 4 aldehydes, 1 acid, and 1 other substances. For most VFCs, their concentrations were low at the beginning of fermentation, increased significantly at day 6 and 18, and then after 18 days, the concentration of VFCs became relatively stable. Among them, esters, alcohols, and ketones were the main aromas during HQW fermentation.
Principal component analysis (PCA) was performed to deeply understand the differences in VFCs of samples at different stages of fermentation. PC 1 and PC 2 accounted for 87.9% and 8.68% of the variance in the population, respectively. Figure 2b suggests that the flavors were considerably different between the initial and other fermentation stages, and the difference between day 6 and day 12 decreased, further decreasing from day 18 to day 30. In other words, the differences in VFCs of samples were smaller as the fermentation time increased. The PCA results indicated that 13 compounds were selected because they were more likely to cause differences between samples, including 3 alcohols (3-methyl-1-butanol, 2-methyl-1-propanol and 1-propanol), 7 esters (isoamyl acetate, ethyl acetate, ethyl butyrate, ethyl hexanoate, octanoic acid ethyl ester, 2-methylpropyl acetate, and propanoic acid ethyl ester), and 1 ketone (butan-2-one). This suggests that esters, alcohols, and ketones are more likely to cause differences in volatile flavors in HQW than other volatile compounds, and these compounds were particularly noted in further analyses.
Esters are the most abundant VFCs in HQW, which can provide fruity, sweet, and floral aromas [10]. A total of 11 esters were identified, including ethyl acetate, isoamyl acetate, ethyl hexanoate, octanoic acid ethyl ester, ethyl butyrate, 2-methylpropyl acetate, propanoic acid ethyl ester, ethyl isobutyrate, methyl butanoate, propyl acetate, and ethyl pentanoate, all of which increased significantly on day 6, and then maintained a slight increase or decrease during the remainder of the fermentation phase. Some of esters come from raw materials, but most of esters are produced by the esterase-catalyzed reaction of alcohols and acids produced from glucose and amino acids during microbial metabolism [28]. Generally, the latter is the primary pathway in the formation of esters [29]. Ethyl acetate and isoamyl acetate are the most abundant ester components, which could render good organoleptic characteristics (fruity, sweet, pineapple) to a fermentative process [30]. Ethyl acetate is regarded as the most important ester component influencing the flavor of rice wine, which was also identified as the dominant VFCs in the final stage of highland barley wine brewing, due to its steady formation from the predominant ethanol and acetic acid [12]. All esters increased most significantly on day 6 than in the other brewing stages. During the final brewing stage (12 to 30 days), the ester components whose level increased mostly were ethyl hexanoate, ethyl butyrate, and octanoic acid ethyl ester. Ethyl hexanoate had a fruity aroma, while ethyl butyrate and octanoic acid ethyl ester brought apple-like and pineapple flavors.
Alcohols are the major contributors for the flavor and aroma of HQW. During fermentation, seven alcohols were identified, including: 3-methyl-1-butanol, ethanol, 2-methyl-1-propanol, 1-propanol,1-hexanol, 1-pentanol, and methanol, in which ethanol, 3-methyl-1 butanol, and 2-methyl-1-propanol accounted for more than 95% of the total alcohols. The concentrations of these three alcohols increased significantly on day 6, then remained stable. Alcohols are produced through the metabolism of sugars and the decarboxylation and dehydrogenation of amino acids [6]. In Yang’s study, 3-methyl-1-butanol and 2-methyl-1-propanol accounted for more than 60% of the total alcohols during the fermentation of Hong Qu Huangjiu [31]. Higher alcohols contain more than two carbons and have a higher boiling point, higher cetane number, and higher energy density than those of ethanol [32]. Higher alcohols can usually bring specific aromas to wines and enhance their aroma complexity [10]. For example, 3-methyl-1-butanol is an additive with a banana flavor, which can improve the taste of wine by reducing the bitter-tasting amino acids (leucine) [33]. 1-hexanol presents herbaceous, woody, and sweet aromas, and 1-pentanol and 2-methyl-1-propanol contribute a light wine aroma to rice wine [34].
During fermentation, the concentration of all five identified ketones tended to increase, including: 3-hydroxy-2-butanone, heptan-2-one, 2-pentanone, butan-2-one, and propan-2-one. Among them, the concentrations of butan-2-one, 2-pentanone, 3-hydroxy-2-butanone, and propan-2-one increased significantly on day 6 and then remained stable, except 3-hydroxy-2-butanone, which decreased significantly on day 12. Meanwhile, heptan-2-one decreased significantly on day 6 and then remained stable. Ketones can also bring a specific aroma to the wine; for example, 3-hydroxy-2 butanone presents a milk-like aroma and heptan-2-one presents a special floral aroma [35]. In addition, four aldehydes were identified during fermentation. They are mainly derived from the oxidation and degradation of lipids and have great influence on the flavor of wine due to their fruity, nutty aroma and low odor threshold [36]. During fermentation, 3-methylbutanal and hexanal continued to decrease to a very low final concentration. 3-methylbutanal is naturally present in lemon essential oil and has an apple and peach flavor when highly diluted [37]. The levels of 2-methyl propanal and propanal were highest on day 6, and they presented a pungent odor and were mainly used in the configuration of fruit flavors [32]. During fermentation, only one volatile acid (acetic acid) was identified. Acetic acid is produced during alcoholic and lactic fermentation and presents a pungent odor, and it is an important precursor to ethyl acetate [10].
3.3. Correlation Analysis of Microbial Communities and Flavor Compounds
The top 10 abundant fungi and bacterial genera were selected to analyze their correlations with flavor compounds by Spearman’s correlation coefficient. To reduce the mistakes due to the increasing number of correlation analyses, correlation p-values were calculated and multi-corrected. As shown in Figure 3, each genus had complex correlations with flavor compounds, and significant correlations were marked.
As shown in Figure 3a, Lactiplantibacillus, Lactobacillus, Leuconostoc, Klebsiella, Weissella, Enterobacter, Bacillus, Cutaneotrichosporon, Alternaria, Colletotrichum, Byssochlamys, Pichia, Haradamyces, Pallidocercospora, and Monascus had positive correlations with the formation of most organic acids and free amino acids. Among them, Lactiplantibacillus had significant positive correlations with lactic acid, citric acid, succinic acid (p < 0.01), and malic acid (p < 0.05). During the fermentation of traditional rice wine, the dominant lactic acid bacteria (LAB) produce most of the organic acids due to multiple carbohydrate metabolic pathways. Lactic acid is the main metabolite of LAB in both homo- and hetero-lactic fermentation [3]. In homolactic fermentation, glucose is converted to pyruvate via the EMP pathway, and to lactic acid catalyzed by lactate dehydrogenase [38]. In heterolactic fermentation, the EMP pathway produces pyruvate, and which is metabolized to ethanol and acetic acid by pyruvate formate lyase and pyruvate oxidase [39]. In the carbohydrate metabolism of LAB, pyruvate plays a central intermediate in the branch of the metabolic pathway. Lactiplantibacillus promotes the formation of pyruvate to secure the output of metabolic downstream products through the EMP pathway [40]. Thus, Lactiplantibacillus promotes the production of citric acid, succinic acid, and malic acid through the tricarboxylic acid cycle (TCA) [41]. Enterobacter had significant positive correlations with lactic acid, citric acid, and succinic acid (p < 0.05), while Burkholderia-Caballeronia-Paraburkholderia was negatively correlated with them (p < 0.05). As parthenogenic anaerobic bacteria, Enterobacter ferments sugar to produce various organic acids, including lactic acids, succinic acids, acetic acids, and formic acids [21].
Free amino acids are generated through proteolysis of the raw material by proteases from endogenous enzymes or microorganisms. The dynamics of the microbial genera leads to changes in the free amino acid components. As shown in Figure 3a, 16 microbial genera are potentially related to the formation of free amino acids. Lactiplantibacillus had significant positive correlations with Ile, Phe, Trp, Arg, Tyr, His, Asp, Thr, Met, Ala, and Pro (p < 0.01), and Leu, Val, Glu, and Ser (p < 0.05). During food fermentation, the glutaminase activity of LAB promotes the accumulation of glutamate (Glu), which contributes to the freshness and umami of fermented foods [42]. When LAB apoptosis occurs, various free amino acids are released from the cells [5]. Previous studies have described that the total bitter amino acid and total umami amino acid content of HQW were strongly and positively correlated with Lactobacillus [16]. Shang used correlation analysis to point out that L. alimentarius was highly correlated with the formation of 10 free amino acids during the natural fermentation of pickled chayote [43]. In addition, Enterobacter was also observed to have a significant positive correlation with 11 free amino acids (p < 0.05). In Spanish fermented meat products, Enterobacter was isolated and, when cultured in the medium, produced large amounts of free amino acids [44]. As a filamentous fungus, Monascus can secrete a large number of proteases to promote the release of free amino acids [15]. Compared to unfermented soybeans, Monascus fermented soybeans achieved a 4.5-fold increase in free amino acids and a significant increase in γ-aminobutyric acid (GABA) [45]. Bacillus is the dominant bacteria in most traditional fermented soy foods, and some of them exhibit strong amylase, protease, and lipase activities [46]. Jang used strains of four Bacillus spp. to ferment soybeans and analyzed changes in free amino acid content, showing that B. licheniformis increased serine, threonine, and glutamate; B. subtilis increased alanine, asparagine, glycine, leucine, proline, tryptophan, and lysine; B. velezensis increased the GABA concentration to >200% of that in the control samples [47]. On the other hand, some free amino acid contents were negatively correlated with microbial genera, such as Burkholderia-Caballeronia-Paraburkholderia and Phialemoniopsis. As an important source of nitrogen for microbial growth, free amino acids are consumed due to the growth of miscellaneous bacteria.
As shown in Figure 3b, microbial genera were divided into two parts by cluster analysis due to different correlations with flavor compounds. Among them, Lactobacillus, Lactiplantibacillus, Cutaneotrichosporon, Colletotrichum, Enterobacter, Haradamyces, Pallidocercospora, Klebsiella, Alternaria, Weissella, Monascus, Comamonas, Leuconostoc, and Bacillus had positive correlations with most of the volatile flavor compounds. Among them, Lactiplantibacillus had significant positive correlations with ethyl acetate, acetic acid, and methyl isobutyrate (p < 0.01). Carbohydrate metabolism and amino acid metabolism are important pathways for the production of volatile flavor substances [11]. In heterolactic fermentation, Lactobacillus produces volatile compounds such as acetic acid and ethanol by consuming sugars. Organic acids and alcohols will produce esters through esterification reactions [27]. As the core flavor-contributing microbiota of traditional Chinese fermented vegetables, Lactobacillus was highly correlated with the formation of several key esters (benzoic acid ethyl ester, isopentyl formate, and ethyl octanoate) [48]. In the traditional brewing process of Chinese Huangjiu, LAB produce large amounts of organic acids that react with alcohols to produce ester aroma components [11]. On the other hand, Lactiplantibacillus was negatively correlated with methanol and 1-pentanol (p < 0.01). Methanol has been historically considered an exogenous product that leads to damage to the human nervous systems when consumed [49]. Previous studies have shown that Lactobacillus can reduce the levels of bad compounds in foods through complex metabolism; for example, the inoculation of Lactobacillus-fermented soybeans effectively reduces the levels of methanol, acetaldehyde, and hexanal [50]. Cutaneotrichosporon had a significant positive correlation with the formation of ethyl pentanoate (p < 0.05). Burkholderia-Caballeronia-Paraburkholderia had a significant positive correlation with ethyl pentanoate (p < 0.01). However, Enterobacter had a negative positive correlation with ethyl pentanoate (p < 0.01). This suggests that different microbial genera may lead to the increase or decrease in the same volatile flavor compounds during HQW fermentation.
Although some dominant microbial genera have no significant correlation with flavor compounds, their positive correlation is equally noteworthy, such as Leuconostoc, Weissella, and Monascus. Monascus is used for saccharification due to its amylase- and protease-producing properties, which contribute to the rapid formation of the flavor of HQW [15]. As shown in Figure 3b, Monascus had positive correlations with the formation of esters, alcohols, acids, aldehydes, and ketones. Leuconostoc and Weissella preferred the formation of esters and ketones. As a fermentation starter, Leuconostoc was shown to promote the formation of 2-butanone during milk fermentation [51]. Xiang found that the combined inoculation of Weissella cibaria and Lactobacillus plantarum significantly increased the content of organic acids (lactic acid, acetic acid), esters (butyl butyrate), alcohols (terpinen-4-ol, cineole, linalool, 4-(1-methylethyl)-cyclohexanol), and olefins (alpha-phellandrene, 1,3-p-menthadiene, myrcene, chrithmene) in traditional Sichuan Pickle [52]. During the fermentation of traditional Korean soybean paste, Weissella had a significant positive correlation with the formation of lactic acid and ethyl acetate [46].
4. Conclusions
The work presented the physicochemical properties, flavor compounds changes, and bacterial and fungal genera succession and their relationships during the fermentation of Hong Qu glutinous rice wine (HQW). Physicochemical properties were used to show the fermentation status of HWQ; the pH decreased and AAN and alcohol increased with prolonged fermentation time. The total sugar increased significantly at day 6 and then significantly decreased thereafter. Flavor compounds, especially Bitter-tasting BAAs (e.g., Arg and Trp), organic acids (e.g., lactic acid, citric acid, and succinic acid), and volatile compounds (e.g., esters and alcohol), accumulated significantly with prolonged fermentation. As for the microbial communities, Pantoea, Lactiplantibacillus, Lactobacillus, Leuconostoc, and Weissella predominated the bacterial genera during the fermentation, and Monascus was the most dominant fungal genus. By correlation analysis and multiple correction, Lactiplantibacillus was identified as a microbial genus due to its significant positive correlation with 4 organic acids, 15 free amino acids, and 3 volatile flavor compounds. This study contributes to the in-depth understanding of the different roles of microorganisms in the formation of flavor compounds during the fermentation of traditional HQW, and provides a theoretical reference for the industrial production of traditional HQW.
Author Contributions
Conceptualization, experiment, software, writing—original draft, writing—review and editing, A.L.; methodology, data curation, writing—review and editing, validation, X.Y.; supervision, project administration, writing—review and editing, validation, Q.G.; supervision, writing—review and editing, validation, B.L.; writing—review and editing, Y.Z.; data curation, software, Y.S.; validation, L.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (31871872); the Central Public-interest Scientific Institution Basal Research Fund, CAFS (NO.2020TD68); and the Central Public-interest Scientific Institution Basal Research Fund, ECSFR, CAFS (NO.2021M01).
Data Availability Statement
Data is contained within the article.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Huang, Y.Y.; Liang, Z.C.; Lin, X.Z.; He, Z.G.; Ren, X.Y.; Li, W.X.; Molnar, I. Fungal community diversity and fermentation characteristics in regional varieties of traditional fermentation starters for Hong Qu glutinous rice wine. Food Res. Int. 2021, 141, 110146. [Google Scholar] [CrossRef]
- Yang, C.L.; Wu, X.P.; Chen, B.; Deng, S.S.; Chen, Z.E.; Huang, Y.Y.; Jin, S.S. Comparative analysis of genetic polymorphisms among Monascus strains by ISSR and RAPD markers. J. Sci. Food Agric. 2017, 97, 636–640. [Google Scholar] [CrossRef] [PubMed]
- Gänzle, M.G. Lactic metabolism revisited: Metabolism of lactic acid bacteria in food fermentations and food spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Ye, M.Q.; Yue, T.L.; Yuan, Y.H. Evolution of polyphenols and organic acids during the fermentation of apple cider. J. Sci. Food Agric. 2014, 94, 2951–2957. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Huang, J.; Xie, T.; Huang, L.; Zhuang, W.; Zheng, Y.; Zheng, B. Oenological characteristics, amino acids and volatile profiles of Hongqu rice wines during pottery storage: Effects of high hydrostatic pressure processing. Food Chem. 2016, 203, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Fan, W.L.; Xu, Y. Characterization of volatile and semi-volatile compounds in Chinese rice wines by headspace solid phase microextraction followed by gas chromatography-mass spectrometry. J. Inst. Brew. 2008, 114, 172–179. [Google Scholar] [CrossRef]
- Jiang, L.; Su, W.; Mu, Y.; Mu, Y. Major Metabolites and Microbial Community of Fermented Black Glutinous Rice Wine With Different Starters. Front. Microbiol. 2020, 11, 593. [Google Scholar] [CrossRef]
- Huang, Z.R.; Hong, J.L.; Xu, J.X.; Li, L.; Guo, W.L.; Pan, Y.Y.; Lv, X.C. Exploring core functional microbiota responsible for the production of volatile flavour during the traditional brewing of Wuyi Hong Qu glutinous rice wine. Food Microbiol. 2018, 76, 487–496. [Google Scholar] [CrossRef]
- Mayo, B.; Rachid, C.; Alegria, A.; Leite, A.M.O.; Peixoto, R.S.; Delgado, S. Impact of Next Generation Sequencing Techniques in Food Microbiology. Curr. Genom. 2014, 15, 293–309. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Su, W.; Mu, Y.; Jiang, L.; Mu, Y. Correlations between microbiota with physicochemical properties and volatile flavor components in black glutinous rice wine fermentation. Food Res. Int. 2020, 138, 109800. [Google Scholar] [CrossRef]
- Wang, P.; Mao, J.; Meng, X.; Li, X.; Liu, Y.; Feng, H. Changes in flavour characteristics and bacterial diversity during the traditional fermentation of Chinese rice wines from Shaoxing region. Food Control 2014, 44, 58–63. [Google Scholar] [CrossRef]
- Guo, L.; Luo, Y.; Zhou, Y.; Bianba, C.; Guo, H.; Zhao, Y.; Fu, H. Exploring microbial dynamics associated with flavours production during highland barley wine fermentation. Food Res. Int. 2020, 130, 108971. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lin, C.; Zhang, W.; Yang, Q.; Meng, J.; He, L.; Zeng, X. Exploring the bacterial community for starters in traditional high-salt fermented Chinese fish (Suanyu). Food Chem. 2021, 358, 129863. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, B.; Fu, Y.; Shi, Y.; Chen, F.; Guan, H.; Zhang, N. HS-GC-IMS with PCA to analyze volatile flavor compounds across different production stages of fermented soybean whey tofu. Food Chem. 2021, 346, 128880. [Google Scholar] [CrossRef]
- Chen, W.; He, Y.; Zhou, Y.; Shao, Y.; Feng, Y.; Li, M.; Chen, F. Edible Filamentous Fungi from the Species Monascus: Early Traditional Fermentations, Modern Molecular Biology, and Future Genomics. Compr. Rev. Food Sci. Food Saf. 2015, 14, 555–567. [Google Scholar] [CrossRef] [Green Version]
- Liang, Z.; Lin, X.; He, Z.; Su, H.; Li, W.; Ren, X. Amino acid and microbial community dynamics during the fermentation of Hong Qu glutinous rice wine. Food Microbiol. 2020, 90, 103467. [Google Scholar] [CrossRef]
- Zheng, J.S.; Wittouck, S.; Salvetti, E.; Franz, C.; Harris, H.M.B.; Mattarelli, P.; Lebeer, S. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Liu, X.; Qian, M.; Shen, Y.; Qin, X.; Huang, H.; Yang, H.; Bai, W. An high-throughput sequencing approach to the preliminary analysis of bacterial communities associated with changes in amino acid nitrogen, organic acid and reducing sugar contents during soy sauce fermentation. Food Chem. 2021, 349, 129131. [Google Scholar] [CrossRef]
- Li, W.W.; Fan, G.S.; Fu, Z.L.; Wang, W.H.; Xu, Y.Q.; Teng, C.; Li, X.T. Effects of fortification of Daqu with various yeasts on microbial community structure and flavor metabolism. Food Res. Int. 2020, 129, 10. [Google Scholar] [CrossRef]
- González, Á.; Hierro, N.; Poblet, M.; Mas, A.; Guillamón, J.M. Application of molecular methods to demonstrate species and strain evolution of acetic acid bacteria population during wine production. Int. J. Food Microbiol. 2005, 102, 295–304. [Google Scholar] [CrossRef]
- Sun, L.; Bai, C.; Xu, H.; Na, N.; Jiang, Y.; Yin, G.; Xue, Y. Succession of Bacterial Community During the Initial Aerobic, Intense Fermentation, and Stable Phases of Whole-Plant Corn Silages Treated with Lactic Acid Bacteria Suspensions Prepared From Other Silages. Front Microbiol. 2021, 12, 655095. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Zhang, T.; Zhang, Q.; Luo, J.; Cai, C.; Mao, J. Microbial diversity and chemical analysis of the starters used in traditional Chinese sweet rice wine. Food Microbiol. 2018, 73, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhou, Y.; Li, J.; Liu, S.; He, S.; Sun, H.; Xu, S. Effect of enzymes addition on the fermentation of Chinese rice wine using defined fungal starter. LWT 2021, 143, 111101. [Google Scholar] [CrossRef]
- Tian, T.; Sun, J.; Wu, D.; Xiao, J.; Lu, J. Objective measures of greengage wine quality: From taste-active compound and aroma-active compound to sensory profiles. Food Chem. 2021, 340, 128179. [Google Scholar] [CrossRef] [PubMed]
- El-Dalatony, M.M.; Saha, S.; Govindwar, S.P.; Abou-Shanab, R.A.I.; Jeon, B.H. Biological Conversion of Amino Acids to Higher Alcohols. Trends Biotechnol. 2019, 37, 855–869. [Google Scholar] [CrossRef]
- Mato, I.; Suárez-Luque, S.; Huidobro, J.F. A review of the analytical methods to determine organic acids in grape juices and wines. Food Res. Int. 2005, 38, 1175–1188. [Google Scholar] [CrossRef]
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Microbial modulation of aromatic esters in wine: Current knowledge and future prospects. Food Chem. 2010, 121, 1–16. [Google Scholar] [CrossRef]
- Ye, Y.T.; Wang, L.X.; Zhan, P.; Tian, H.L.; Liu, J.S. Characterization of the aroma compounds of Millet Huangjiu at different fermentation stages. Food Chem. 2022, 366, 10. [Google Scholar] [CrossRef]
- Yang, Y.; Hu, W.; Xia, Y.; Mu, Z.; Tao, L.; Song, X.; Zhang, H.; Ni, B.; Ai, L. Flavor Formation in Chinese Rice Wine (Huangjiu): Impacts of the Flavor-Active Microorganisms, Raw Materials, and Fermentation Technology. Front. Microbiol. 2020, 11, 2541. [Google Scholar] [CrossRef]
- Larralde-Corona, C.P.; De la Torre-Gonzalez, F.J.; Vazquez-Landaverde, P.A.; Hahn, D.; Narvaez-Zapata, J.A. Rational Selection of Mixed Yeasts Starters for Agave Must Fermentation. Front. Sustain. Food Syst. 2021, 5, 13. [Google Scholar] [CrossRef]
- Yang, Y.J.; Xia, Y.J.; Song, X.; Mu, Z.Y.; Qiu, H.Z.; Tao, L.R.; Ai, L.Z. The Potential of Flos sophorae immaturus as a Pigment-Stabilizer to Improve the Monascus Pigments Preservation, Flavor Profiles, and Sensory Characteristic of Hong Qu Huangjiu. Front. Microbiol. 2021, 12, 678903. [Google Scholar] [CrossRef] [PubMed]
- El Hadi, M.A.; Zhang, F.J.; Wu, F.F.; Zhou, C.H.; Tao, J. Advances in fruit aroma volatile research. Molecules 2013, 18, 8200–8229. [Google Scholar] [CrossRef]
- Chen, C.; Liu, Z.; Zhou, W.Y.; Tian, H.X.; Huang, J.; Yuan, H.B.; Yu, H.Y. Comparison of the Fermentation Activities and Volatile Flavor Profiles of Chinese Rice Wine Fermented Using an Artificial Starter, a Traditional JIUYAO and a Commercial Starter. Front. Microbiol. 2021, 12, 716281. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.Y.; Xie, T.; Xie, J.R.; Ai, L.Z.; Tian, H.X. Characterization of key aroma compounds in Chinese rice wine using gas chromatography-mass spectrometry and gas chromatography-olfactometry. Food Chem. 2019, 293, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Contador, R.; Delgado, F.J.; Garcia-Parra, J.; Garrido, M.; Ramirez, R. Volatile profile of breast milk subjected to high-pressure processing or thermal treatment. Food Chem. 2015, 180, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Bueno, M.; Carrascon, V.; Ferreira, V. Release and Formation of Oxidation-Related Aldehydes during Wine Oxidation. J. Agric. Food Chem. 2016, 64, 608–617. [Google Scholar] [CrossRef] [Green Version]
- Yuan, F.; He, F.; Qian, Y.; Zheng, J.; Qian, M.C. Aroma Stability of Lemon-Flavored Hard Iced Tea Assessed by Chirality and Aroma Extract Dilution Analysis. J. Agric. Food Chem. 2016, 64, 5717–5723. [Google Scholar] [CrossRef]
- Gänzle, M.G.; Vermeulen, N.; Vogel, R.F. Carbohydrate, peptide and lipid metabolism of lactic acid bacteria in sourdough. Food Microbiol. 2007, 24, 128–138. [Google Scholar] [CrossRef]
- Cocaign-Bousquet, M.; Garrigues, C.; Loubiere, P.; Lindley, N.D. Physiology of pyruvate metabolism in Lactococcus lactis. Antonie Van Leeuwenhoek 1996, 70, 253–267. [Google Scholar] [CrossRef]
- Ganzle, M.G.; Follador, R. Metabolism of oligosaccharides and starch in lactobacilli: A review. Front. Microbiol. 2012, 3, 340. [Google Scholar] [CrossRef] [Green Version]
- Rabinowitz, J.D.; Enerback, S. Lactate: The ugly duckling of energy metabolism. Nat. Metab. 2020, 2, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, J.S.; Seeras, A.; Sanchez-Maldonado, A.F.; Zhang, C.; Su, M.S.-W.; Gänzle, M.G. Glutamine, glutamate, and argi-nine-based acid resistance in Lactobacillus reuteri. Food Microbiol. 2014, 42, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Shang, Z.; Ye, Z.; Li, M.; Ren, H.; Cai, S.; Hu, X.; Yi, J. Dynamics of microbial communities, flavor, and physicochemical properties of pickled chayote during an industrial-scale natural fermentation: Correlation between microorganisms and metabolites. Food Chem. 2022, 377, 132004. [Google Scholar] [CrossRef]
- Hortensia Silla Santos, M. Amino acid decarboxylase capability of microorganisms isolated in Spanish fermented meat products. Int. J. Food Microbiol. 1998, 39, 227–230. [Google Scholar] [CrossRef]
- Pyo, Y.-H. Effect of Monascus-fermentation on the Content of GABA and Free Amino Acids in Soybean. J. Korean Soc. Food Sci. Nutr. 2008, 37, 1208–1213. [Google Scholar] [CrossRef]
- Han, D.M.; Chun, B.H.; Kim, H.M.; Jeon, C.O. Characterization and correlation of microbial communities and metabolite and volatile compounds in doenjang fermentation. Food Res. Int. 2021, 148, 110645. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Jeong, D.W.; Heo, G.; Kong, H.; Kim, C.T.; Lee, J.H. Genetic Background Behind the Amino Acid Profiles of Fermented Soybeans Produced by Four Bacillus spp. J. Microbiol. Biotechnol. 2021, 31, 447–455. [Google Scholar] [CrossRef]
- Xiao, Y.; Huang, T.; Huang, C.; Hardie, J.; Peng, Z.; Xie, M.; Xiong, T. The microbial communities and flavour compounds of Jiangxi yancai, Sichuan paocai and Dongbei suancai: Three major types of traditional Chinese fermented vegetables. LWT 2020, 121, 108865. [Google Scholar] [CrossRef]
- Dorokhov, Y.L.; Shindyapina, A.V.; Sheshukova, E.V.; Komarova, T.V. Metabolic Methanol: Molecular Pathways and Physiological Roles. Physiol. Rev. 2015, 95, 603–644. [Google Scholar] [CrossRef]
- Blagden, T.D.; Gilliland, S.E. Reduction of levels of volatile components associated with the “Beany” flavor in soymilk by Lactobacilli and Streptococci. J. Food Sci. 2005, 70, M186–M189. [Google Scholar] [CrossRef]
- Alegría, Á.; Delgado, S.; Flórez, A.B.; Mayo, B. Identification, typing, and functional characterization of Leuconostoc spp. strains from traditional, starter-free cheeses. Dairy Sci. Technol. 2013, 93, 657–673. [Google Scholar] [CrossRef] [Green Version]
- Xiang, W.L.; Zhang, N.D.; Lu, Y.; Zhao, Q.H.; Xu, Q.; Rao, Y.; Zhang, Q. Effect of Weissella cibaria co-inoculation on the quality of Sichuan Pickle fermented by Lactobacillus plantarum. LWT 2020, 121, 7. [Google Scholar] [CrossRef]
Figure 1.
Changes in relative abundances of (a) fungi and (b) bacteria genera during Hong Qu glutinous rice wine (HQW) fermentation.
Figure 1.
Changes in relative abundances of (a) fungi and (b) bacteria genera during Hong Qu glutinous rice wine (HQW) fermentation.

Figure 2.
(a) Gallery plot and (b) PCA of volatile flavor compounds (VFCs) in HQW at different fermentation stages.
Figure 2.
(a) Gallery plot and (b) PCA of volatile flavor compounds (VFCs) in HQW at different fermentation stages.

Figure 3.
Correlation between the relative abundance of microbial genera and flavor compounds. The Spearman correlation coefficient reflects the correlation between the top 10 dominant bacterial/fungal species with (a) free amino acids and organic acids and (b) VFCs during the fermentation of HQW. fdr was used to correct the correlation p-values, “**” represents significance at p < 0.01, “*” represents significance at p < 0.05.
Figure 3.
Correlation between the relative abundance of microbial genera and flavor compounds. The Spearman correlation coefficient reflects the correlation between the top 10 dominant bacterial/fungal species with (a) free amino acids and organic acids and (b) VFCs during the fermentation of HQW. fdr was used to correct the correlation p-values, “**” represents significance at p < 0.01, “*” represents significance at p < 0.05.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Changes in physicochemical properties during HQW fermentation.
| Physicochemical Index | D0 | D6 | D12 | D18 | D24 | D30 |
|---|---|---|---|---|---|---|
| pH | 5.83 ± 0.01 a | 3.86 ± 0.02 c | 3.90 ± 0.01 b | 3.85 ± 0.01 c | 3.79 ± 0.02 d | 3.84 ± 0.01 c |
| Total sugar (g/L) | 14.36 ± 0.17 b | 48.70 ± 1.20 a | 3.47 ± 0.17 c | 3.21 ± 0.80 c | 2.29 ± 0.15 cd | 1.56 ± 0.08 d |
| Alcohol (v/v, %) | 0.18 ± 0.02 e | 7.33 ± 1.10 d | 14.00 ± 0.28 c | 17.20 ± 0.30 b | 18.80 ± 0.87 a | 19.77 ± 0.15 a |
| Amino acid nitrogen (g/L) | 0.10 ± 0.004 f | 0.14 ± 0.01 e | 0.33 ± 0.02 d | 0.43 ± 0.004 c | 0.54 ± 0.004 b | 0.65 ± 0.01 a |
Values are presented as mean ± standard error (n = 3), a–f values with different letters in the same row are significantly different (p < 0.05) from each other.
Table 2.
Changes in the free amino acids fraction of HQW during different fermentation stages.
| Amino Acid (mg/L) | D0 | D6 | D12 | D18 | D24 | D30 |
|---|---|---|---|---|---|---|
| Umami | ||||||
| Asp | 6.40 ± 0.12 f | 20.31 ± 0.15 e | 79.83 ± 2.06 d | 108.34 ± 2.25 c | 142.99 ± 1.42 b | 161.20 ± 2.11 a |
| Glu | 31.24 ± 0.53 e | 26.41 ± 0.12 e | 92.7 ± 1.92 d | 323.69 ± 9.55 c | 370.19 ± 5.43 b | 392.54 ± 7.64 a |
| TUAA | 37.64 | 46.72 | 172.53 | 432.03 | 513.18 | 553.74 |
| Sweet | ||||||
| Ser | 10.38 ± 0.16 e | 7.39 ± 0.09 f | 36.12 ± 0.93 d | 56.77 ± 1.04 c | 77.35 ± 1.29 b | 94.16 ± 2.64 a |
| Gly | 3.13 ± 0.07 c,d | 1.49 ± 0.01 e | 8.04 ± 0.25 b | 11.91 ± 0.25 a | 3.49 ± 0.64 c | 2.59 ± 0.08 d |
| Thr | 12.43 ± 0.27 f | 44.98 ± 0.25 e | 145.62 ± 3.23 d | 195.54 ± 3.98 c | 226.90 ± 3.98 b | 247.38 ± 5.50 a |
| Met | 7.21 ± 0.74 f | 15.97 ± 0.21 e | 22.71 ± 0.30 d | 53.72 ± 0.88 c | 81.95 ± 1.08 b | 85.64 ± 1.59 a |
| Ala | 1.43 ± 0.08 e | 3.26 ± 0.22 e | 7.49 ± 0.20 d | 26.98 ± 0.94 c | 67.06 ± 1.27 b | 75.28 ± 1.98 a |
| Pro | 31.05 ± 3.57 f | 113.86 ± 2.46 e | 228.6 ± 8.42 c | 247.15 ± 5.14 c | 274.8 ± 14.08 b | 306.4 ± 7.01 a |
| TSAA | 34.58 | 73.00 | 219.98 | 344.91 | 456.70 | 505.05 |
| Bitter | ||||||
| His | 13.08 ± 0.19 f | 35 ± 0.34 e | 95.93 ± 1.57 d | 139.9 ± 4.04 c | 185.57 ± 3.97 b | 218.86 ± 6.10 a |
| Arg | 46 ± 0.76 f | 127.77 ± 0.87 e | 261.12 ± 5.7 d | 377.87 ± 7.37 c | 481.23 ± 8.74 b | 518.08 ± 12.91 a |
| Lys | 1.54 ± 0.70 c | 1.71 ± 0.31 c | 1.29 ± 0.48 c | 2.59 ± 0.26 b | 3.9 ± 0.23 a | 4.63 ± 0.10 a |
| Val | 4.03 ± 0.07 e | 3.1 ± 0.09 e | 26.86 ± 0.64 d | 50.63 ± 0.98 c | 73.92 ± 1.07 b | 91.02 ± 2.43 a |
| Trp | 50.46 ± 1.51 f | 132.71 ± 0.53 e | 475.03 ± 10.21 d | 613.99 ± 13.01 c | 769.46 ± 9.59 b | 871.73 ± 14.36 a |
| Phe | 4.87 ± 0.15 f | 12.81 ± 0.05 e | 45.85 ± 0.99 d | 59.26 ± 1.26 c | 74.26 ± 0.93 b | 84.13 ± 1.39 a |
| Ile | 10.62 ± 0.17 f | 36.65 ± 0.63 e | 154.35 ± 4.67 d | 216.44 ± 4.41 c | 277.72 ± 4.31 b | 318.41 ± 6.75 a |
| Leu | 4.44 ± 1.03 e | 3.3 ± 0.06 e | 27.42 ± 0.23 d | 38.2 ± 1.55 c | 48.74 ± 0.42 b | 57.23 ± 1.05 a |
| TBAA | 135.04 | 353.05 | 1087.85 | 1498.88 | 1914.80 | 2164.09 |
| Astringent | ||||||
| Tyr | 16.46 ± 0.45 f | 38.79 ± 0.17 e | 89.47 ± 2.04 d | 113.22 ± 1.72 c | 130.5 ± 1.9 b | 140.08 ± 2.7 a |
| TAAA | 16.46 | 38.79 | 89.47 | 113.22 | 130.5 | 140.08 |
| TAA | 254.77 | 625.42 | 1798.43 | 2636.19 | 3289.98 | 3669.36 |
Values are presented as mean ± standard error (n = 3), a–f values with different letters in the same row are significantly different (p < 0.05) from each other.
Table 3.
Changes in the organic acids fraction of HQW during fermentation.
| Organic Acid (mg/L) | D0 | D6 | D12 | D18 | D24 | D30 |
|---|---|---|---|---|---|---|
| Oxalic acid | 54.3 ± 0.1 c | 37.1 ± 0.8 d | 59.4 ± 2.6 b | 61.9 ± 0.9 a | 57.4 ± 0.5 b | 53.0 ± 0.1 c |
| Tartaric acid | 785.0 ± 10.5 d | 784.7 ± 9.1 d | 1102.0 ± 4.5 a | 1067.5 ± 18.5 a,b | 1014.4 ± 17.4 b | 953.9 ± 14.1 c |
| Lactic acid | 27.1 ± 0.6 f | 4027.6 ± 6.5 e | 6486.9 ± 12.3 d | 7253.0 ± 11.7 c | 7654.2 ± 79.0 b | 8073.9 ± 46.9 a |
| Citric acid | 106.7 ± 2.2 d | 582.2 ± 17.0 c,d | 650.0 ± 11.5 c,d | 1086.0 ± 22.5b c | 1559.2 ± 23.2 a,b | 1877.4 ± 38.0 a |
| Succinic acid | 58.9 ± 0.6 f | 656.4 ± 18.6 e | 1056.3 ± 3.3 d | 1347.7 ± 6.7 c | 1808.5 ± 63.4 b | 2422.4 ± 13.0 a |
| Malic acid | 128.5 ± 2.8 e | 104.0 ± 5.4 c,d | 196.3 ± 9.7 d | 560.9 ± 13.7 c | 1123.6 ± 35.5 b | 1223.2 ± 38.4 a |
| Total | 1160.5 | 6192.0 | 9550.9 | 11,377.0 | 13,217.3 | 14,603.8 |
Values are presented as mean ± standard error (n = 3), a–f values with different letters in the same row are significantly different (p < 0.05) from each other.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, A.; Yang, X.; Guo, Q.; Li, B.; Zheng, Y.; Shi, Y.; Zhu, L. Microbial Communities and Flavor Compounds during the Fermentation of Traditional Hong Qu Glutinous Rice Wine. Foods 2022, 11, 1097. https://doi.org/10.3390/foods11081097
AMA Style
Liu A, Yang X, Guo Q, Li B, Zheng Y, Shi Y, Zhu L. Microbial Communities and Flavor Compounds during the Fermentation of Traditional Hong Qu Glutinous Rice Wine. Foods. 2022; 11(8):1097. https://doi.org/10.3390/foods11081097
Chicago/Turabian StyleLiu, Anqi, Xu Yang, Quanyou Guo, Baoguo Li, Yao Zheng, Yuzhuo Shi, and Lin Zhu. 2022. "Microbial Communities and Flavor Compounds during the Fermentation of Traditional Hong Qu Glutinous Rice Wine" Foods 11, no. 8: 1097. https://doi.org/10.3390/foods11081097
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.