
Tree Nuts and Peanuts as a Source of Beneficial Compounds and a Threat for Allergic Consumers: Overview on Methods for Their Detection in Complex Food Products

 , , ,
, , , 
Abstract
:1. Introduction
1.1. Nut/Peanut Allergy: Prevalence and Epidemiology
1.2. European Regulatory Framework
1.3. Analytical Methods for Protein Allergen Detection in Foods
1.4. Untargeted Versus Targeted Mass Spectrometry for Allergens Analysis
2. Overview of Nuts and Peanut Allergens Composition and Analytical Methods for Their Detection
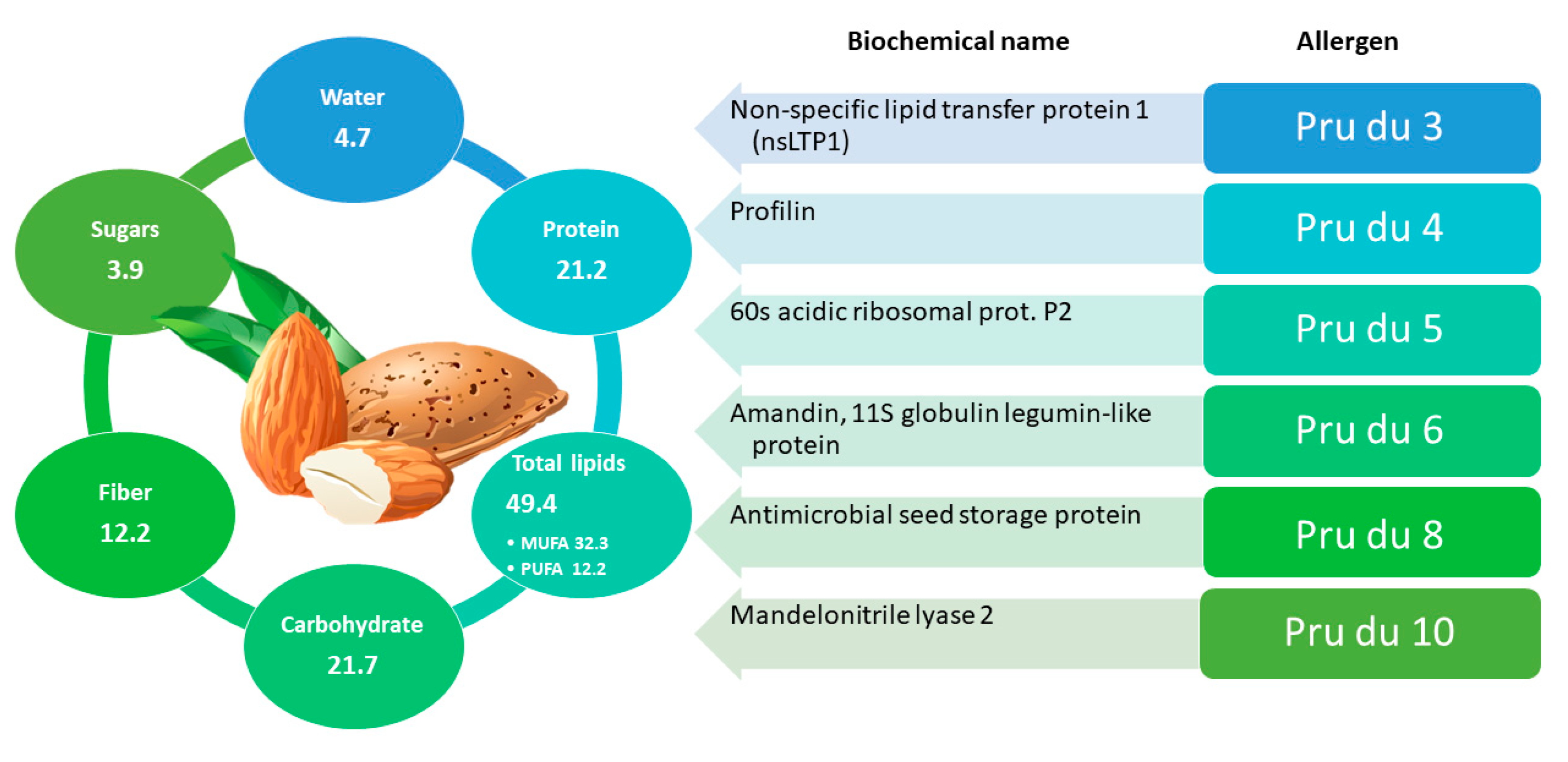
2.1. Almond
2.1.1. General Information and Main Allergens
2.1.2. Immunochemical/Molecular Biology Analytical Methods
2.1.3. Mass Spectrometry-Based Analytical Methods for Detecting Almond Allergens
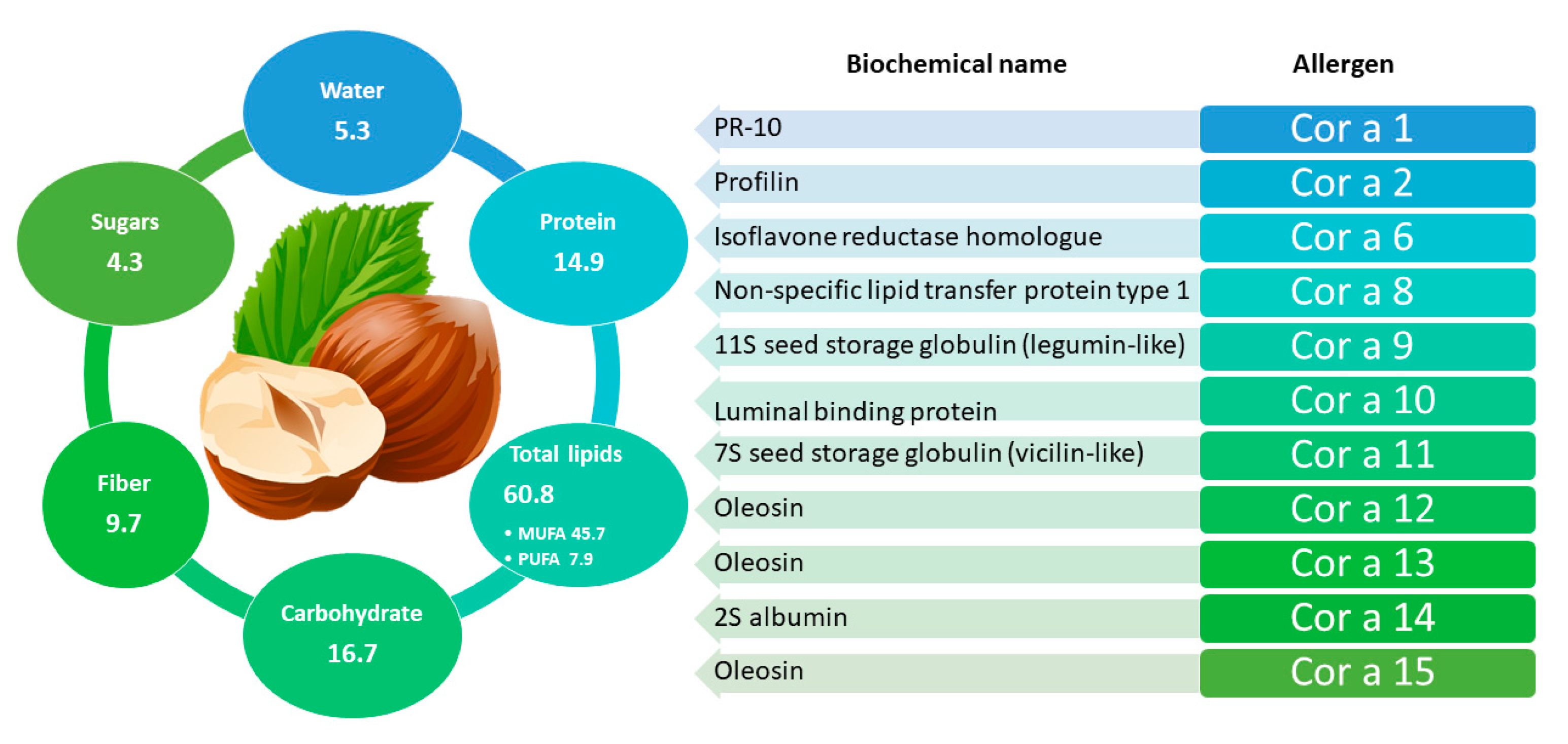
2.2. Hazelnut
2.2.1. General Information and Main Allergens
2.2.2. Immunochemical/Molecular Biology Analytical Methods for Hazelnut Detection
2.2.3. Mass Spectrometry-Based Analytical Methods for Hazelnut Analysis
2.3. Cashew
2.3.1. General Information and Main Allergens
2.3.2. Immunochemical/Molecular Biology Analytical Methods for Cashew Detection
2.3.3. Mass Spectrometry-Based Analytical Methods for Cashew Analysis
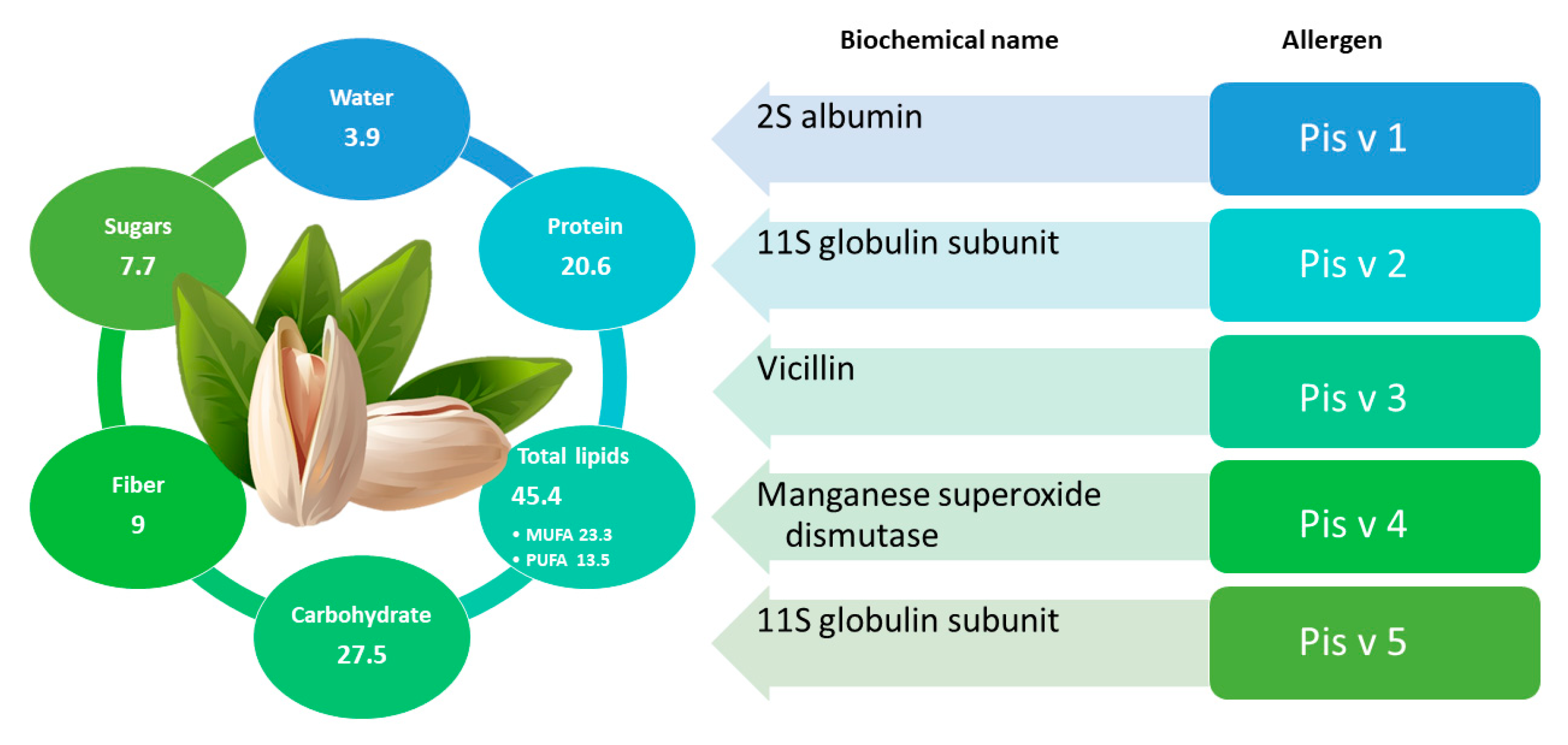
2.4. Pistachio
2.4.1. General Information and Main Allergens
2.4.2. Immunochemical/Molecular Biology Analytical Methods for Pistachio Detection
2.4.3. Mass Spectrometry-Based Analytical Methods for Pistachio Analysis
2.5. Walnut
2.5.1. General Information and Main Allergens
2.5.2. Immunochemical/Molecular Biology Analytical Methods for Walnut Detection
2.5.3. Mass Spectrometry-Based Analytical Methods for Walnut Analysis
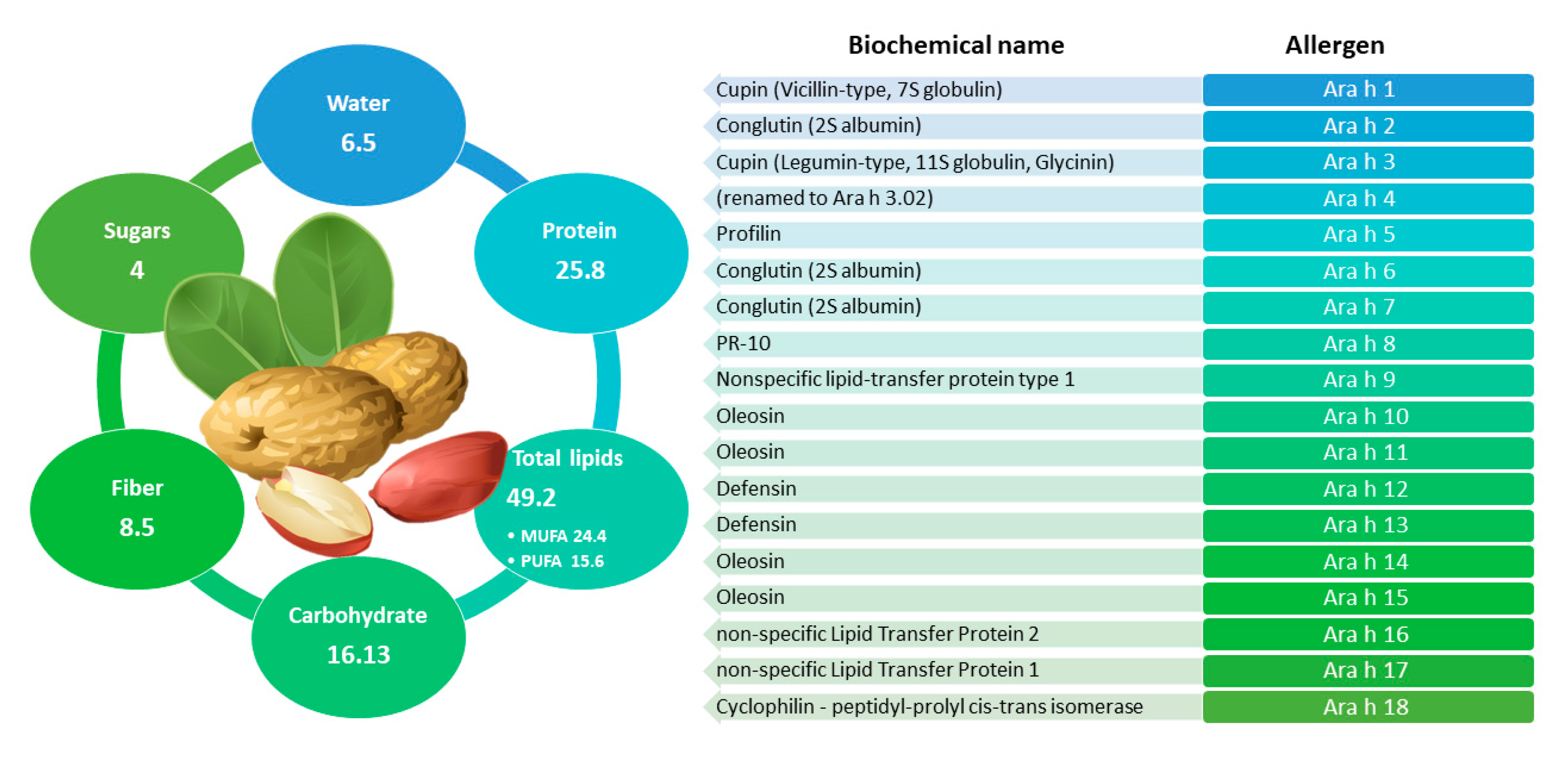
2.6. Peanut
2.6.1. General Information and Main Allergens
2.6.2. Immunochemical/Molecular Biology Analytical Methods for Peanuts Detection
2.6.3. Mass Spectrometry-Based Analytical Methods for Peanuts Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Allergen | Description | Target Protein | Matrix | LOD | LOQ | Cross-Reactivity | Reference |
|---|---|---|---|---|---|---|---|
| ALMOND | Sandwich 4C10 ELISA | Pru du 6 | Biscuit, Brittle, Granola bar with raw almond, Granola bar with roasted almond | 0.97 ± 0.32 mg/kg full fat almond | 3.24 ± 1.07 mg/kg full fat almond | Not reported | [87] |
| Sandwich MonoTrace ELISA | 0.30 ± 0.17 mg/kg full fat almond | 1.01 ± 0.58 mg/kg full fat almond | |||||
| Sandwich | Pru du 6 | Vanilla ice cream, cookies, pasta sauce, milk chocolate | 0.3 mg/kg | 2.5 mg/kg | No cross-reactivity | [88] | |
| Sandwich | Pru du 6 | Orange juice, Coffee liquor, chocolate soy drink, salad dressing, rice ice cream, goat cheese | 0.02 mg/kg | 0.12 mg/kg | Low cross reactivity to chestnut | [90] | |
| HAZELNUT | Sandwich | Hazelnut proteins | Cookies, chocolate | >0.03 mg/kg | 0.375 mg/kg | Not reported | [107] |
| Sandwich-Ridascreen Fast Hazelnut | Hazelnut proteins | 31 plant-derived foods and 2 animal-derived foods | 1.5 mg/kg | 2.5 mg/kg | No apparent cross-reactivity | [103] | |
| Sandwich-AgraQuant Hazelnut Assay | Hazelnut proteins | 30 plant-derived foods | 0.3 mg/kg | 1 mg/kg | No cross-reactivity | ||
| Sandwich-DIA hazelnut | Hazelnut proteins | 31 plant-derived foods | 0.33 mg/kg | 1 mg/kg | No cross-reactivity | ||
| Sandwich-Veratox for hazelnut | Hazelnut proteins | 2.5 mg/kg | 2.5 mg/kg | No available information about specificity | |||
| Sandwich | Protein from ground hazelnut & hazelnut chocolates | Dried hazelnuts and chocolate with 41% cocoa | 1 mg/kg | 50–100 mg/kg | Not reported | [37] | |
| CASHEW | Sandwich (CAS-ELISA-2) | Ana o 3 | Chocolate | 0.04 mg/kg | 0.4 mg/kg | Very low cross-reactivity with pistachio, pecan, almond, peanut, and hazelnut. | [140] |
| Cookie | 40 mg/kg | ||||||
| Sandwich (Ano3-ELISA-1) | Ana o 3 | Chocolate | 0.06 mg/kg | 0.4 mg/kg | |||
| Cookie | 40 mg/kg | ||||||
| Sandwich | Cashew proteins | Ice Cream, cookies, chocolate, nuts/ Mixed nuts, breakfast/granola or Nutritional bars, nut butter | 0.11 mg/g | 0.46 mgcashew g_1 | Not reported | [223] | |
| PISTACHIO | Sandwich enzyme-linked immunosorbent assay (ELISA) | Pistachio proteins | 156 commonly used foods and food ingredients | 0.09 mg/kg | 0.30 mg/kg | No cross-reactivity | [161] |
| Sandwich ELISA AgraQuant® Plus Pistachio | Pistachio proteins | 1 mg/kg | 1–25 mg/kg | Cashew (12%), hazelnut (0.17%), walnut (0.0008%), pecan nut (0.0005%), sunflower (0.0002%) | [145] | ||
| Sandwich-AgraQuant® ELISA Pistachio | Pistachio proteins | 0.13 mg/kg | 1–40 mg/kg | Cashew (12%), hazelnut (0.17%), walnut (0.0008%), pecan nut (0.0005%), sunflower (0.0002%) | |||
| Monoclonal antibody-based ELISA to pistachio allergen | Pistachio proteins | 0.12 mg/kg | 1–40 mg/kg | Pecan nut (0.001%) | |||
| WALNUT | Indirect competitive | Jug r 1 | Tree nuts, seeds, cereals, soy, milk, various animal products | 0.22 mg/kg walnut protein | 0.44 mg/kg walnut protein | 0.2% to pecan | [182] |
| Sandwich ELISA | Walnut proteins | Sauce, beverage, yoghurts, ice-cream, and sandwich | 2.2 mg/kg | 3.3 mg/kg | Cross-reactivity to almond and pecan | [183] | |
| Direct ELISA with Multimeric scFv | 1616 mg/kg | Cross-reactivity was found with pecan (2.25%) and almond (0.35%) | |||||
| Sandwich | Jug r 1 | chicken meatballs, rice porridge, bread, sponge cake, orange juice, jelly, biscuit | <0.16 mg/kg of walnut protein defined by the calibration curve | <0.31 mg/kg | Strong cross-reactivity with pecan nut, hazelnut, brazil nut, almond, pine nut, peanut, cashew nut, pistachio, macadamia and mustard | [180] | |
| PEANUT | Sandwich- | Ara h 1 Ara h 2 | Biscuits prepared with peanut butter | Ara h 1: 0.1 mg/kg Ara h 2: 0.13 mg/kg | 20–800 mg/mL | Slight interference nuts, seeds, cereals | [207] |
| Direct competitive | Ara h 1: 0.19 mg/kg Ara h 2: 0.06 mg/kg | 20 ng/mL–2 mg/mL | Slight with nuts, seeds, cereals | ||||
| Sandwich | Ara h 2 | Peanut kernels, peanut beans, peanut crispy rolls, chocolate-peanut beans | 0.7–1.7 μg/kg of peanut product | No information | Low cross-reactivity to cashews, walnuts, BSA, ovalbumin, soy & pea proteins | [224] | |
| Sandwich | Peanut proteins | Cookies, chocolate | >0.03 mg/kg | 0.555 mg/kg | Not reported | [107] | |
| Sandwich | Peanut proteins | Non-alcoholic beverages, confectionery, bread and bakery, mixed and/or processed foods | 0.13 mg/kg | 2.5 mg/kg | Not reported | [208] |
| ALMOND-Prunus dulcis | ||||
|---|---|---|---|---|
| Allergenic Protein | Molecular Weight | Food Processing Effects | Selected Peptide Sequences | References |
| Pru du 5 (Q8H2B9) (60s acidic ribosomal prot. P2) | 10 kDa | Unknown | DITELIASGR | [225] |
| Pru du 6 (E3SH28; E3SH29) (Amandin, 11S globulin legumin-like protein) | 360 kDa | Thermally stable to dry heat such as roasting but it can be denatured by boiling. | QETIALSSSQQR | [50,52,112,225,226] |
| GNLDFVQPPR | [48,49,50,92,112,144,164,225,226,227,228,229] | |||
| ALPDEVLANAYQISR | [48,49,92,112,225,226,227,228] | |||
| ISTLNSHNLPILR | [163,225,226,227] | |||
| NGLHLPSYSNAPQLIYIVQGR | [48,49,225,227] | |||
| QQEQLQQER | [91,92,143,225,229] | |||
| QQEQEQER | [229] | |||
| TEENAFINTLAGR | [163,225,226,227] | |||
| GVLGAVFSGCPETFEESQQSSQQGR | [48,225,227] | |||
| VQGQLDFVSPFSR | [50,144,164,225] | |||
| ALPDEVLQNAFR | [50,144,164,225,226,229] | |||
| ADIFSPR | [225,226,229] | |||
| LSQNIGDPSR | [225,229] | |||
| VQVVNENGDPILNDEVR | [50,229] | |||
| VQVVNENGDPILDDEVR | [225,229] | |||
| NGIYSPHWNVNAHSVVYVIR | [225] | |||
| NLQGQNDNR | [50,225] | |||
| FYLAGNPENEFNQQGQSQPR | [225,226] | |||
| QQGQQEQQQER | [91,226] | |||
| QQEEQQSQR | [229] | |||
| QQEQQQGQQGRPQQQQQFR | [225] | |||
| QEGGQGQQQFQGEDQQDR | [229] | |||
| FYLAGNPQDEFNPQQQGR | [225] | |||
| NHLPILR | [225] | |||
| ADFYNPQGGR | [92,225] | |||
| LLSATSPPR | [50,92,225] | |||
| QQQQQGEQGR | [229] | |||
| NQIIQVR | [226] | |||
| ENIGNPER | [226] | |||
| TDENGFTNTLAGR | [50] | |||
| HAZELNUT-Corylus avellana | ||||
| Cor a 8 (Q9ATH2) (Non-specific lipid transfer protein type 1) | 9 kDa | Resistant to the activity of gastric and intestinal enzymes, heat treatments, abrupt changes in pH and the inhospitable environment of the gastrointestinal tract (proteolysis). Less stable when subjected to temperatures above 90 °C | GIAGLNPNLAAGLPGK | [37,109] |
| AVNDASR | [110] | |||
| Cor a 9 (Q8W1C2) 11S seed storage globulin (legumin-like) | 40 kDa | Thermostable protein, sensitive to autoclaving processes (121 °C or 138 °C, 15 or 30 min) | LNALEPTNR | [50,110,144,163,164,221,226,228] |
| INTVNSNTLPVLR | [4,37,48,49,50,109,163,228,230] | |||
| WLQLSAER | [37,92,109,163,226,230] | |||
| ALPDDVLANAFQISR | [37,48,49,109,112,113,114,226,228,230] | |||
| VQVVDDNGNTVFDDELR | [144,164,230] | |||
| QGQQQFGQR | [230] | |||
| EGLYVPHWNLNAHSVVYAIR | [110] | |||
| ADIYTEQVGR | [4,47,48,49,66,90,91,111,112,141,214,220,225,226,229] | |||
| QEWER | [91] | |||
| AESEGFEWVAFK | [226] | |||
| QETTLVR | [226] | |||
| TNDNAQISPLAGR | [114,226,228] | |||
| HFYLAGNPDDEHQR | [110,230] | |||
| QGQVLTIPQNFAVAK | [37,48,49,50,109,112,114,144,164,228] | |||
| INTVNSNTLPVLR | [226] | |||
| QGQVLTIPQNFAVAK | [226] | |||
| Cor a 11 (Q8S4P9) 7S seed storage globulin (vicilin-like) | 48 kDa | Thermostable proteins with major thermal transition of around 70–75 °C; The physico-chemical properties of Cor at 11 were affected after heat treatment at 60 °C and at 145 °C in the presence of glucose; High pressure processing (300–600 Mba) of the hazelnuts did not affect the ability to bind the IgE of the allergens of Cor a 11. | LLSGIENFR | [4,37,109,110,163,226,230] |
| GNIVNEFER | [110] | |||
| VQVLENFTK | [226] | |||
| ALSQHEEGPPR | [230] | |||
| HPSQSNQFGR | [230] | |||
| ALSQHEEGPPR | [230] | |||
| GSMAGPYYNSR | [230] | |||
| IPAGTPVYMINR | [230] | |||
| ESFNVEHGDIIR | [230] | |||
| NQDQAFFFPGPNK | [230] | |||
| IWPFGGESSGPINLLHK | [110] | |||
| ILQPVSAPGHFEAFYGAGGEDPESFYR | [110,230] | |||
| AFSWEVLEAALK | [4,37,109,226,230] | |||
| ELAFNLPSR | [37,109,226,230] | |||
| Cor a 14 (D0PWG2) (2S albumin) | 10 kDa reducing | High thermal stability and in difficult conditions of the gastrointestinal tract due to their compact and rigid structure, thus preserving their allergenic activity. | QAVMQQQGEMR | [230] |
| QQNLNQCQR | [230] | |||
| PISTACHIO–Pistacia vera | ||||
| Pis v 1 (B7P072) 2S Albumin | 7 kDa | Unknown | LQELYETASELPR | [163,228] |
| TNGLSQTSQLAGR | [163] | |||
| Pis v 2 (B7P073; B7P074) 11S Globulin subunit | 32 kDa | Unknown | VTSINALNLPILR | [20,112] |
| ALPLDVIK | [112,228] | |||
| TNGLSQTSQLAGR | [50] | |||
| GLPLDVIQNSFDISR | [50] | |||
| NSFDISR | [92] | |||
| EGQLVVVPQNFAVVK | [92] | |||
| IQIVSENGESVFDEEIR | [50] | |||
| Pis v 3 (B4X640) Vicilin | 55 kDa | Unknown | IAIVVSGEGR | [50] |
| STGTFNLFK | [50] | |||
| Pis v 5 (B7SLJ1) 11S Globulin subunit | 36 kDa (acidic subunit) | Unknown | ITSLNSLNLPILK | [112,144,164,228] |
| AMISPLAGSTSVLR | [50,112,144,164,228] | |||
| GFESEEESEYER | [50,144,164] | |||
| CASHEW-Anacardium occidentale | ||||
| Ana o 1 (Q8L5L5; Q8L5L6) Vicilin-like protein | 50 kDa | Resistant to pH and high temperatures. IgE reactivity was strongly reduced in cashew nut subjected to gamma irradiation at followed by autoclaving at 121 °C during 30 min. | AFSWEILEAALK | [50] |
| CAGVALVR | [112,228] | |||
| AMTSPLAGR | [92,112,143,228] | |||
| ADIYTPEVGR | [50,91,92,143,144,163,228] | |||
| Ana o 2 (Q8GZP6) Legumin-like protein | 55 kDa | Thermostable protein Ana or 2 immunoreactivity was markedly reduced with high sodium sulfite concentrations (≥50 mM) and high temperatures (≥100 °C). | LDALEPDNR | [92] |
| WLQLSVEK | [50,134] | |||
| TSVLGGMPEEVLANAFQISR | [50,134] | |||
| EGQMLVVPQNFAVVK | [50,134,144] | |||
| LTTLNSLNLPILK | [50,134,144] | |||
| VFDGEVR | [91] | |||
| Ana o 3 (Q8H2B8) 2S albumin | 14 kDa | High thermal stability Stable over a pH range of 1 to 11. | ELYETASELPR | [112,228] |
| QLQQQEQIK | [163] | |||
| WALNUT–Juglans regia | ||||
| Jug r 1 (P93198) 2S albumin seed storage protein | 15–16 kDa | High resistance to enzymatic activity (trypsin/chymotrypsin) at basic (pH 8.0), although they progressively lose allergenicity in acidic conditions (pH 1.3) in the presence of pepsin. Resistance to thermal denaturation. | DLPNECGISSQR | [48,49,50] |
| QCCQQLSQMDEQCQCEGLR | [48,49] | |||
| GEEMEEMVQSAR | [48,49,112,163,228] | |||
| Jug r 2 (Q9SEW4) Vicilin seed storage protein | 44 kDa | Remarkable thermal stability, which allows them to maintain their conformation at temperatures below 70–75 °C. However, when subjected to elevated temperatures, 7S globulins can undergo structural disruption and covalent modifications. | ATLTLVSQETR | [50,112,144,164,228] |
| HNPYYFHSQSIR | [50] | |||
| FFDQQEQR | [50,92,144,164] | |||
| DFLAGQNNIINQLER | [92] | |||
| VFSNDILVAALNTPR | [92] | |||
| QQQQQGLR | [163] | |||
| Jug r 4 (Q2TPW5) 11S globulin seed storage protein | 58.1 kDa | High thermal stability and great resistance to proteolysis, which allows them to maintain their allergenic properties along the gastrointestinal system. | ALPEEVLATAFQIPR | [50,144,164] |
| EGQLLTIPQNFAVVKR | [50] | |||
| LDALEPTNR | [91,143] | |||
| NFYLAGNPDDEFR | [50] | |||
| EFQQDR | [91] | |||
| ADIYTEEAGR | [143] | |||
| PEANUT-Arachis hypogaea | ||||
| Ara h 1 (P43238) Cupin (Vicillin-type, 7S globulin) | 64 kDa | Heat stable protein that undergoes irreversible denaturation at T > 80 °C. Roasting at T > 140 °C produces an increase in the IgE binding capacity of Ara h 1. Hydration before autoclaving increases the effectiveness of the heat treatment by significantly altering its immunoreactivity. | DLAFPGSGEQVEK | [4,41,47,48,212,215,216,225,226,227,230,231,232] |
| VLLEENAGGEQEER | [4,41,49,112,212,214,225,226,227,230,231,232] | |||
| GTGNLELVAVR | [24,47,48,91,112,161,218,220,227,229,230] | |||
| SFNLDEGHALR | [47,92,163,231] | |||
| NNPFYFPSR | [25,47,92,219,229,233] | |||
| NTLEAAFNAEFNEIR | [47,188,231] | |||
| EGALMLPHFNSK | [231] | |||
| ISMPVNTPGQFEDFFPASSR | [231] | |||
| EEEEDEDEEEEGSNR | [231] | |||
| EGEQEWGTPGSHVR | [216,231] | |||
| AMVIVVVNK | [114,231] | |||
| EHVEELTK | [231] | |||
| IVQIEAKPNTLVLPK | [231] | |||
| QFQNLQNHR | [231] | |||
| EGEPDLSNNFGK | [231] | |||
| DQSSYLQGFSR | [110,216,226,231] | |||
| SSENNEGVIVK | [229,231] | |||
| NNPFYFPSR | [221] | |||
| GSEEEGDITNPINLR | [110,221] | |||
| GSEEEDITNPINLR | [221,222] | |||
| IFLAGDKDNVIDQIEK | [217] | |||
| Ara h 2 (Q6PSU2-1) Conglutin (2S albumin) | 17 kDa | Stable to heat treatment and proteolysis, Ara h 2 allergenicity can increase with roasting and decrease with frying or boiling. Hydration before autoclaving increases the effectiveness of the heat treatment by significantly altering its immunoreactivity. | CCNELNEFENNQR | [25,215,219,228,229,232,233,234,235,236] |
| NLPQQCGLR | [25,112,216,219,228,229,232,234,236,237] | |||
| CMCEALQQIMENQSDR | [25,215,229,232,235,236,238] | |||
| CQSQLER | [236] | |||
| CDLEVESGGR | [216,229,232,236] | |||
| GAGSSQHQER | [236] | |||
| DEDSYGR | [236] | |||
| ANLRPCEQHLMQK | [236] | |||
| QQEQQFK | [216,236] | |||
| QQWELQGDR | [236] | |||
| Ara h 3 (O82580; Q9SQH7) Cupin (Legumin-type, 11S globulin, Glycinin) | 60 kDa, 37 kDa (fragment) | The allergenicity of Ara h 3 increases with roasting and decreases with frying or boiling. Hydration before autoclaving increases the effectiveness of the heat treatment by significantly altering its immunoreactivity. | SPDIYNPQAGSLK | [25,42,91,92,143,215,219,229,232,234,235,239] |
| RPFYSNAPQEIFIQQGR | [4,20,48,49,92,112,188,237,239] | |||
| FNLAGNHEQEFLR | [25,92,143,144,164,188,219,221,228,229,239] | |||
| LNAQRPDNR | [231,232,237] | |||
| WLGLSAEYGNLYR | [4,25,48,49,50,67,144,164,221,226,229,239] | |||
| SQSENFEYVAFK | [42,50,221,229,233,239] | |||
| AHVQVVDSNGNR | [163,232,239] | |||
| GETESEEEGAIVTVR | [231,239] | |||
| QQPEENACQFQR | [228,234,239] | |||
| TANDLNLLILR | [144,163,164,226,239] | |||
| FFVPPSQQSPR | [239] | |||
| FFVPPFQQSPR | [239] | |||
| NALFVPHYNTNAHSIIYALR | [221,239] | |||
| QIVQNLWGENESEEEGAIVTVR | [239] | |||
| GYFGLIFPGCPSTYEEPAQQGR | [239] | |||
| ADEEEEYDEDEYEYDEEDR | [239] | |||
| VYDEELQEGHVLVVPQNFAVAGK | [221,239] | |||
| TANELNLLILR | [20,112,226,228] | |||
| FFVPPSEQSLR | [226] | |||
| GENESDEQGAIVTVR | [226] | |||
| QIVQNLR | [226] | |||
| AQSENYEYLAFK | [221,226] | |||
| SQSDNFEYVAFK | [221,226] | |||
| TANDNLLLLILR | [221] | |||
| TANELLILILR | [110,221] | |||
| TVNELDLPILNR | [221] | |||
| VYDEELQEGHVLVVPQNFAVAAK | [221] | |||
| Ara h 6 (Q647G9) Conglutin (2S albumin) | 15 kDa | Stable to heat treatment and proteolysis. Hydration before autoclaving increases the effectiveness of the heat treatment by significantly altering its immunoreactivity. | CDLDVSGGR | [228,234] |
| Ara h 7 (Q9SQH1; B4XID4; Q647G8) Conglutin (2S albumin) | 15 kDa | Stable to heat treatment and proteolysis. | NLPQNCGFR | [228,234] |
| Allergen | MS Analyzer (Analysis Mode) | Target Protein | Matrix | LOD | LOQ | Reference |
|---|---|---|---|---|---|---|
| ALMOND | LIT (SRM/SRM3) | Pru du 1 | Cookies | SRM 17 mg/kg, SRM3 25 mg/kg | SRM 58 mg/kg, SRM3 80 mg/kg | [91] |
| QqQ/LIT (SRM) | Pru du 6 | Bread | 3 mg/kg | / | [49] | |
| LIT (SRM) | Pru du 1 | Biscuits | 0.9 mg/kg | 3.1 mg/kg | [143] | |
| Dark chocolate | 9 mg/kg | 30 mg/kg | ||||
| OrbitrapTM (Full MS) | Pru du 6 | Milk chocolate, vanilla ice cream, commercial bread, and breakfast cereals | 0.34–1.92 mgPROT/kg, 1.8–10.1 mgNUT/kg | / | [50] | |
| TQ-S (MRM) | Pru du 6 | Incurred chocolate, ice cream, cookies, and sauce | >3 mg/kg | 5–25 mg/kg | [52] | |
| QqQ/LIT (SRM) | Pru du 1 | Chocolates | 0.4 mg/kg | 1.3 mg/kg | [92] | |
| Pru du 2 | Chocolates | 0.8 mg/kg | 2.6 mg/kg | |||
| HAZELNUT | LIT (SRM/SRM3) | Cor a 9 | Cookies | SRM 30 mg/kg, SRM3 35 mg/kg | SRM 90 mg/kg, SRM3 110 mg/kg | [91] |
| QqQ/LIT (SRM) | Cor a 9 | Bread | 5 mg/kg | / | [49] | |
| LIT (SRM) | Cor a 9 | Biscuits | 1.3 mg/kg | 4.5 mg/kg | [143] | |
| Dark chocolate | 14 mg/kg | 49 mg/kg | ||||
| LIT (SRM) | Cor a 9 | Cookies | 1–5 mg/kg | 16–32 mg/kg | [218] | |
| TQ-S (MRM) | Cor a 9 | Incurred chocolate, ice cream, cookies, and sauce | >3 mg/kg | 2.5–5 mg/kg | [52] | |
| Q-Orbitrap (t-SIM/dd2) | Cor a 9 | Incurred cookie | 4 mg/kg | 12 mg/kg | [114] | |
| QTRAP 6500 (IDA-MS/MS) | Cor a 9 | Cookies | ≤2.25 mg/kg | ≤3 mg/kg | [4] | |
| Ice cream | ≤2 mg/kg | ≤10 mg/kg | ||||
| Breakfast cereal | ≤0.45 mg/kg | ≤3 mg/kg | ||||
| Milk chocolate | ≤10 mg/kg | ≤30 mg/kg | ||||
| QqQ/LIT (SRM) | Cor a 9 | Chocolates | 0.5 mg/kg | 1.7 mg/kg | [92] | |
| TripleTOF 6600 (IDA-MS/MS) | Cor a 9 | Cookies | 3.1 mg/kg | 5.3 mg/kg | [67] | |
| CASHEW | LIT (SRM/SRM3) | Ana o 2 | Cookies | SRM 14 mg/kg, SRM3 30 mg/kg | SRM 46 mg/kg, SRM3 98 mg/kg | [91] |
| LIT (SRM) | Ana o 2 | Biscuits | 0.5 mg/kg | 1.6 mg/kg | [143] | |
| Dark chocolate | 15 mg/kg | 50 mg/kg | ||||
| OrbitrapTM (Full MS) | Ana o 2 | Milk chocolate, vanilla ice cream, commercial bread, and breakfast cereals | 0.78–2.02 mgPROT/kg 4.3–11.2 mgNUT/kg | / | [50] | |
| QqQ/LIT (SRM) | Ana o 2 | Chocolates | 0.7 mg/kg | 2.3 mg/kg | [92] | |
| TQ-S (MRM) | Ana o 2 | Incurred chocolate, ice cream, cookies, and sauce | >3 mg/kg | 2.5 mg/kg | [52] | |
| Ana o 3 | >3 mg/kg | 2.5 mg/kg | ||||
| PISTACHIO | OrbitrapTM (Full MS) | Pis v 2 | Milk chocolate, vanilla ice cream, commercial bread, and breakfast cereals | 1.28–1.90 mgPROT/kg 7.10–10.6 mgNUT/kg | / | [50] |
| Pis v 5 | 0.91–1.38 mgPROT/kg 5.1–7.6 mgNUT/kg | / | ||||
| TQ-S (MRM) | Pis v 2, Pis v 5 | Incurred chocolate, ice cream, cookies, and sauce | >3 mg/kg | 2.5 mg/kg | [52] | |
| QqQ/LIT (SRM) | Pis v 2 | Chocolates | 0.4 mg/kg | 1.3 mg/kg | [92] | |
| WALNUT | LIT (SRM/SRM3) | Jug r 4 | Cookies | SRM 55 mg/kg, SRM3 50 mg/kg | SRM 180 mg/kg, SRM3 160 mg/kg | [91] |
| QqQ/LIT (SRM) | Jug r 1 | Bread | 70 mg/kg | / | [49] | |
| LIT (SRM) | Jug r 4 | Biscuits | 0.8 mg/kg | 2.6 mg/kg | [143] | |
| Dark chocolate | 5 mg/kg | 18 mg/kg | ||||
| OrbitrapTM (Full MS) | ND | Milk chocolate, vanilla ice cream, commercial bread, and breakfast cereals | 0.80–5 mgPROT/kg 5.7–35.7 mgNUT/kg | / | [50] | |
| TQ-S (MRM) | Vici lin-like protein | Incurred chocolate, ice cream, cookies, and sauce | >3 mg/kg | 12.5 mg/kg | [52] | |
| Jug r 1 | 5 mg/kg | |||||
| QqQ/LIT (SRM) | Jug r 2 | Chocolates | 0.6 mg/kg | 2.0 mg/kg | [92] | |
| PEANUT | LIT (SRM/SRM3) | Ara h 3/4 | Cookies | SRM 10 mg/kg, SRM3 27 mg/kg | SRM 37 mg/kg, SRM3 90 mg/kg | [91] |
| QqQ/LIT (SRM) | Ara h 1 | Bread | 11 mg/kg | / | [49] | |
| LIT (SRM) | Ara h 3/4 | Biscuits | 0.1 mg/kg | 0.3 mg/kg | [143] | |
| Dark chocolate | 7 mg/kg | 25 mg/kg | ||||
| LIT (SRM) | Ara h 1 | Cookies | 8–9 mg/kg | 30 mg/kg | [218] | |
| TQ-S (MRM) | Ara h 2, Ara h 3/4 | Incurred chocolate, ice cream, cookies, and sauce | >3 mg/kg | 5 mg/kg | [52] | |
| Q-Orbitrap (t-SIM/dd2) | Ara h 1 | Incurred cookie | 7 mgPROT/kg | 24 mgPROT/kg | [114] | |
| QTRAP 6500 (IDA-MS/MS) | Ara h 3 | Cookies | ≤0.3 mg/kg | ≤3 mg/kg | [4] | |
| Ice cream | ≤2 mg/kg | ≤10 mg/kg | ||||
| Breakfast cereal | ≤0.1 mg/kg | ≤1 mg/kg | ||||
| Milk chocolate | ≤1.5 mg/kg | ≤10 mg/kg | ||||
| QqQ/LIT (SRM) | Ara h 1 | Chocolates | 0.8 mg/kg | 2.6 mg/kg | [92] | |
| Ara h 3/4 | Chocolates | 1.3 mg/kg | 4.3 mg/kg | |||
| Q-TOF (MS/MS) | Ara h 1 | Cookies | <2.5 mg/kg | 0.30 mg/kg | [216] | |
| Ara h 2 | 0.13 mg/kg | |||||
| TripleTOF 6600 (IDA-MS/MS) | Ara h 3 | Cookies | 2.2 mg/kg | 6.7 mg/kg | [67] | |
| Ara h 3 | Breakfast cereal | 1.2 mg/kg | 2.0 mg/kg | |||
| QqQ (MRM) | Ara h 1 | Wheat flour matrix (both raw and cooked) | 0.15 mg/kg | 0.31 mg/kg | [217] |
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Husain, Z.; Schwartz, R.A. Food allergy update: More than a peanut of a problem. Int. J. Dermatol. 2013, 52, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Sicherer, S.H.; Sampson, H.A. Food allergy: Epidemiology, pathogenesis, diagnosis, and treatment. J. Allergy Clin. Immunol. 2014, 133, 291–307.e5. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.T.; Ewan, P.W. The development and progression of allergy to multiple nuts at different ages. Pediatr. Allergy Immunol. 2005, 16, 507–511. [Google Scholar] [CrossRef] [PubMed]
- Sun New, L.; Schreiber, A.; Stahl-Zeng, J.; Liu, H.F. Simultaneous analysis of multiple allergens in food products by LC-MS/MS. J. AOAC Int. 2018, 101, 132–145. [Google Scholar] [CrossRef]
- McWilliam, V.; Koplin, J.; Lodge, C.; Tang, M.; Dharmage, S.; Allen, K. The Prevalence of Tree Nut Allergy: A Systematic Review. Curr. Allergy Asthma Rep. 2015, 15, 54. [Google Scholar] [CrossRef] [PubMed]
- Husain, Z.; Schwartz, R.A. Peanut allergy: An increasingly common life-threatening disorder. J. Am. Acad. Dermatol. 2012, 66, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Crespo, J.F.; James, J.M.; Fernandez-Rodriguez, C.; Rodriguez, J. Food allergy: Nuts and tree nuts. Br. J. Nutr. 2006, 96, 95–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, A.L.; Eigenmann, P.A.; Sicherer, S.H. Clinical relevance of cross-reactivity in food allergy. J. Allergy Clin. Immunol. Pract. 2021, 9, 82–99. [Google Scholar] [CrossRef]
- Bolling, B.W.; Mckay, D.L.; Blumberg, J.B. The phytochemical composition and antioxidant actions of tree nuts. Asia Pac. J. Clin. Nutr. Author Manuscr. 2016, 19, 117–123. [Google Scholar]
- Sicherer, S.H.; Sampson, H.A. Food allergy: A review and update on epidemiology, pathogenesis, diagnosis, prevention, and management. J. Allergy Clin. Immunol. 2018, 141, 41–58. [Google Scholar] [CrossRef] [Green Version]
- Bégin, P.; Winterroth, L.C.; Dominguez, T.; Wilson, S.P.; Bacal, L.; Mehrotra, A.; Kausch, B.; Trela, A.; Hoyte, E.; O’Riordan, G.; et al. Erratum to: Safety and feasibility of oral immunotherapy to multiple allergens for food allergy [Allergy, Asthma & Clinical Immunology 2014, 10:1]. Allergy Asthma Clin. Immunol. 2016, 12, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Croote, D.; Quake, S.R. Food allergen detection by mass spectrometry: The role of systems biology. NPJ Syst. Biol. Appl. 2016, 2, 16022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manohar, M.; Nadeau, K.C. The Potential of Anti-IgE in Food Allergy Therapy. Curr. Treat. Options Allergy 2014, 1, 145–156. [Google Scholar] [CrossRef] [PubMed]
- European Union Commission. Notice C 428/1 of 13 July 2017 Commission Notice of 13 July 2017 relating to the provision of information on substances or products causing allergies or intolerances as listed in Annex II to Regulation (EU) No 1169/2011 of the European Parliament and of the Council on the provision of. Off. J. Eur. Union 2017, II, 11–15. [Google Scholar]
- Clare Mills, E.N.; Adel-Patient, K.; Bernard, H.; De Loose, M.; Gillard, N.; Huet, A.C.; Larré, C.; Nitride, C.; Pilolli, R.; Tranquet, O.; et al. Detection and quantification of allergens in foods and minimum eliciting doses in food-Allergic individuals (ThRAll). J. AOAC Int. 2019, 102, 1346–1353. [Google Scholar] [CrossRef] [PubMed]
- Muraro, A.; Werfel, T.; Hoffmann-Sommergruber, K.; Roberts, G.; Beyer, K.; Bindslev-Jensen, C.; Cardona, V.; Dubois, A.; Dutoit, G.; Eigenmann, P.; et al. EAACI Food Allergy and Anaphylaxis Guidelines: Diagnosis and management of food allergy. Allergy Eur. J. Allergy Clin. Immunol. 2014, 69, 1008–1025. [Google Scholar] [CrossRef]
- Allen, K.J.; Turner, P.J.; Pawankar, R.; Taylor, S.; Sicherer, S.; Lack, G.; Rosario, N.; Ebisawa, M.; Wong, G.; Mills, E.N.C.; et al. Precautionary labelling of foods for allergen content: Are we ready for a global framework? World Allergy Organ. J. 2014, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- DunnGalvin, A.; Chan, C.H.; Crevel, R.; Grimshaw, K.; Poms, R.; Schnadt, S.; Taylor, S.L.; Turner, P.; Allen, K.J.; Austin, M.; et al. Precautionary allergen labelling: Perspectives from key stakeholder groups. Allergy Eur. J. Allergy Clin. Immunol. 2015, 70, 1039–1051. [Google Scholar] [CrossRef]
- Ben-Shoshan, M.; Sheth, S.; Harrington, D.; Soller, L.; Fragapane, J.; Joseph, L.; St Pierre, Y.; La Vieille, S.; Elliott, S.; Waserman, S.; et al. Effect of precautionary statements on the purchasing practices of Canadians directly and indirectly affected by food allergies. J. Allergy Clin. Immunol. 2012, 129, 1401–1404. [Google Scholar] [CrossRef]
- Planque, M.; Arnould, T.; Renard, P.; Delahaut, P.; Dieu, M.; Gillard, N. Highlight on bottlenecks in food allergen analysis: Detection and quantification by mass spectrometry. J. AOAC Int. 2017, 100, 1126–1130. [Google Scholar] [CrossRef] [Green Version]
- Allen, K.J.; Remington, B.C.; Baumert, J.L.; Crevel, R.W.R.; Houben, G.F.; Brooke-Taylor, S.; Kruizinga, A.G.; Taylor, S.L. Allergen reference doses for precautionary labeling (VITAL 2.0): Clinical implications. J. Allergy Clin. Immunol. 2014, 133, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Buhler, S.; Tedeschi, T.; Faccini, A.; Garino, C.; Arlorio, M.; Dossena, A.; Sforza, S. Isolation and full characterisation of a potentially allergenic lipid transfer protein (LTP) in almond. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2015, 32, 648–656. [Google Scholar] [CrossRef]
- Remington, B.C.; Westerhout, J.; Meima, M.Y.; Blom, W.M.; Kruizinga, A.G.; Wheeler, M.W.; Taylor, S.L.; Houben, G.F.; Baumert, J.L. Updated population minimal eliciting dose distributions for use in risk assessment of 14 priority food allergens. Food Chem. Toxicol. 2020, 139, 111259. [Google Scholar] [CrossRef] [PubMed]
- Prado, M.; Ortea, I.; Vial, S.; Rivas, J.; Calo-Mata, P.; Barros-Velázquez, J. Advanced DNA- and protein-based methods for the detection and investigation of food allergens. Crit. Rev. Food Sci. Nutr. 2016, 56, 2511–2542. [Google Scholar] [CrossRef]
- Parker, C.H.; Khuda, S.E.; Pereira, M.; Ross, M.M.; Fu, T.J.; Fan, X.; Wu, Y.; Williams, K.M.; DeVries, J.; Pulvermacher, B.; et al. Multi-allergen Quantitation and the Impact of Thermal Treatment in Industry-Processed Baked Goods by ELISA and Liquid Chromatography-Tandem Mass Spectrometry. J. Agric. Food Chem. 2015, 63, 10669–10680. [Google Scholar] [CrossRef] [PubMed]
- Khuda, S.; Slate, A.; Pereira, M.; Al-Taher, F.; Jackson, L.; Diaz-Amigo, C.; Bigley, E.C.; Whitaker, T.; Williams, K.M. Effect of processing on recovery and variability associated with immunochemical analytical methods for multiple allergens in a single matrix: Sugar cookies. J. Agric. Food Chem. 2012, 60, 4195–4203. [Google Scholar] [CrossRef] [PubMed]
- Clare Mills, E.N.; Sancho, A.I.; Rigby, N.M.; Jenkins, J.A.; Mackie, A.R. Impact of food processing on the structural and allergenic properties of food allergens. Mol. Nutr. Food Res. 2009, 53, 963–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albillos, S.M.; Al-Taher, F.; Maks, N. Increasing extractability of protein for allergen detection after food processing. Food Chem. 2011, 127, 1831–1834. [Google Scholar] [CrossRef]
- Wei, Y.; Sathe, S.K.; Teuber, S.S.; Roux, K.H. A Sensitive Sandwich EUSA for the Detection of Trace Amount of Cashew Nut in Foods. J. Allergy Clin. Immunol. 2002, 109, 8303. [Google Scholar] [CrossRef]
- Monaci, L.; Visconti, A. Immunochemical and DNA-based methods in food allergen analysis and quality assurance perspectives. Trends Food Sci. Technol. 2010, 21, 272–283. [Google Scholar] [CrossRef]
- Arlorio, M.; Cereti, E.; Coïsson, J.D.; Travaglia, F.; Martelli, A. Detection of hazelnut (Corylus spp.) in processed foods using real-time PCR. Food Control 2007, 18, 140–148. [Google Scholar] [CrossRef]
- Costa, J.; Oliveira, M.B.P.P.; Mafra, I. Effect of thermal processing on the performance of the novel single-tube nested real-time PCR for the detection of walnut allergens in sponge cakes. Food Res. Int. 2013, 54, 1722–1729. [Google Scholar] [CrossRef]
- Costa, J.; Oliveira, M.B.P.P.; Mafra, I. Novel approach based on single-tube nested real-time PCR to detect almond allergens in foods. Food Res. Int. 2013, 51, 228–235. [Google Scholar] [CrossRef]
- Köppel, R.; Dvorak, V.; Zimmerli, F.; Breitenmoser, A.; Eugster, A.; Waiblinger, H.U. Two tetraplex real-time PCR for the detection and quantification of DNA from eight allergens in food. Eur. Food Res. Technol. 2010, 230, 367–374. [Google Scholar] [CrossRef]
- Köppel, R.; van Velsen-Zimmerli, F.; Bucher, T. Two quantitative hexaplex real-time PCR systems for the detection and quantification of DNA from twelve allergens in food. Eur. Food Res. Technol. 2012, 235, 843–852. [Google Scholar] [CrossRef]
- Pafundo, S.; Gullì, M.; Marmiroli, N. Multiplex real-time PCR using SYBR® GreenERTM for the detection of DNA allergens in food. Anal. Bioanal. Chem. 2010, 396, 1831–1839. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.; Ansari, P.; Mafra, I.; Oliveira, M.B.P.P.; Baumgartner, S. Assessing hazelnut allergens by protein- and DNA-based approaches: LC-MS/MS, ELISA and real-time PCR. Anal. Bioanal. Chem. 2014, 406, 2581–2590. [Google Scholar] [CrossRef]
- Siegel, M.; Mutschler, A.; Boernsen, B.; Pietsch, K.; Waiblinger, H.U. Food matrix standards for the quantification of allergenic food ingredients using real-time PCR. Eur. Food Res. Technol. 2013, 237, 185–197. [Google Scholar] [CrossRef]
- Poms, R.E.; Klein, C.L.; Anklam, E. Methods for allergen analysis in food: A review. In Food Additives and Contaminants; European Commission, DG Joint Research Centre, Institute for Reference Materials and Measurements, Retieseweg: Geel, Belgium, 2004; Volume 21, pp. 1–31. ISBN 0265203031. [Google Scholar]
- Pilolli, R.; Chaudhari, R.; Palmisano, F.; Monaci, L. Development of a mass spectrometry immunoassay for unambiguous detection of egg allergen traces in wines. Anal. Bioanal. Chem. 2017, 409, 1581–1589. [Google Scholar] [CrossRef]
- Picariello, G.; Mamone, G.; Addeo, F.; Ferranti, P. The frontiers of mass spectrometry-based techniques in food allergenomics. J. Chromatogr. A 2011, 1218, 7386–7398. [Google Scholar] [CrossRef]
- Chassaigne, H.; Nørgaard, J.V.; Van Hengel, A.J. Proteomics-based approach to detect and identify major allergens in processed peanuts by capillary LC-Q-TOF (MS/MS). J. Agric. Food Chem. 2007, 55, 4461–4473. [Google Scholar] [CrossRef] [PubMed]
- Eidhammer, I.; Flikka, K.; Martens, L.; Mikalsen, S.O. Computational Methods for Mass Spectrometry Proteomics; John Wiley and Sons: Chichester, England, 2007; pp. 1–284. ISBN 9780470512975. [Google Scholar]
- Monaci, L.; De Angelis, E.; Montemurro, N.; Pilolli, R. Comprehensive overview and recent advances in proteomics MS based methods for food allergens analysis. TrAC-Trends Anal. Chem. 2018, 106, 21–36. [Google Scholar] [CrossRef]
- Pilolli, R.; Nitride, C.; Gillard, N.; Huet, A.C.; van Poucke, C.; de Loose, M.; Tranquet, O.; Larré, C.; Adel-Patient, K.; Bernard, H.; et al. Critical review on proteotypic peptide marker tracing for six allergenic ingredients in incurred foods by mass spectrometry. Food Res. Int. 2020, 128, 108747. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.; Raymond, P.; Ben-Rejeb, S.; Lau, B. Development of a liquid chromatography-tandem mass spectrometry method using capillary liquid chromatography and nanoelectrospray ionization-quadrupole time-of-flight hybrid mass spectrometer for the detection of milk allergens. J. Agric. Food Chem. 2006, 54, 1604–1610. [Google Scholar] [CrossRef] [PubMed]
- Shefcheck, K.J.; Musser, S.M. Confirmation of the Allergenic Peanut Protein, Ara h 1, in a Model Food Matrix Using Liquid Chromatography/Tandem Mass Spectrometry (LC/MS/MS). J. Agric. Food Chem. 2004, 52, 2785–2790. [Google Scholar] [CrossRef] [PubMed]
- Heick, J.; Fischer, M.; Kerbach, S.; Tamm, U.; Popping, B. Application of a liquid chromatography tandem mass spectrometry method for the simultaneous detection of seven allergenic foods in flour and bread and comparison of the method with commercially available ELISA test kits. J. AOAC Int. 2011, 94, 1060–1068. [Google Scholar] [CrossRef] [Green Version]
- Heick, J.; Fischer, M.; Pöpping, B. First screening method for the simultaneous detection of seven allergens by liquid chromatography mass spectrometry. J. Chromatogr. A 2011, 1218, 938–943. [Google Scholar] [CrossRef] [PubMed]
- Korte, R.; Lepski, S.; Brockmeyer, J. Comprehensive peptide marker identification for the detection of multiple nut allergens using a non-targeted LC-HRMS multi-method. Anal. Bioanal. Chem. 2016, 408, 3059–3069. [Google Scholar] [CrossRef]
- Mattarozzi, M.; Bignardi, C.; Elviri, L.; Careri, M. Rapid shotgun proteomic liquid chromatography-electrospray ionization-tandem mass spectrometry-based method for the lupin (Lupinus albus L.) multi-allergen determination in foods. J. Agric. Food Chem. 2012, 60, 5841–5846. [Google Scholar] [CrossRef]
- Planque, M.; Arnould, T.; Dieu, M.; Delahaut, P.; Renard, P.; Gillard, N. Liquid chromatography coupled to tandem mass spectrometry for detecting ten allergens in complex and incurred foodstuffs. J. Chromatogr. A 2017, 1530, 138–151. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.B.; Turko, I.V.; Phinney, K.W. Quantitative performance of internal standard platforms for absolute protein quantification using multiple reaction monitoring-mass spectrometry. Anal. Chem. 2015, 87, 4429–4435. [Google Scholar] [CrossRef] [PubMed]
- Nouri-nigjeh, E.; Zhang, M.; Ji, T.; Yu, H.; An, B.; Duan, X. Effects of Calibration Approaches on the Accuracy for LC−MS Targeted Quantification of Therapeutic Protein. Anal. Chem. 2014, 86, 3575–3584. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.M.; Khuda, S.E.; Parker, C.H.; Eischeid, A.C.; Pereira, M. Detection of Allergen Markers in Food: Analytical Methods. In Food Safety; Spizzirri, U.G., Cirillo, G., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; pp. 65–121. ISBN 978111916058. [Google Scholar]
- Monaci, L.; van Hengel, A.J. Development of a method for the quantification of whey allergen traces in mixed-fruit juices based on liquid chromatography with mass spectrometric detection. J. Chromatogr. A 2008, 1192, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Monaci, L.; Pilolli, R.; De Angelis, E.; Godula, M.; Visconti, A. Multi-allergen detection in food by micro high-performance liquid chromatography coupled to a dual cell linear ion trap mass spectrometry. J. Chromatogr. A 2014, 1358, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lai, S.; Zhang, Y.; Huang, B.; Li, D.; Ren, Y. Multiple reaction monitoring-based determination of bovine α-lactalbumin in infant formulas and whey protein concentrates by ultra-high performance liquid chromatography-tandem mass spectrometry using tryptic signature peptides and synthetic peptide stand. Anal. Chim. Acta 2012, 727, 47–53. [Google Scholar] [CrossRef]
- Posada-Ayala, M.; Alvarez-Llamas, G.; Maroto, A.S.; Maes, X.; Muñoz-Garcia, E.; Villalba, M.; Rodríguez, R.; Perez-Gordo, M.; Vivanco, F.; Pastor-Vargas, C.; et al. Novel liquid chromatography-mass spectrometry method for sensitive determination of the mustard allergen Sin a 1 in food. Food Chem. 2015, 183, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Monaci, L.; Losito, I.; De Angelis, E.; Pilolli, R.; Visconti, A. Multi-Allergen quantification of fining-related egg and milk proteins in white wines by high-resolution mass spectrometry. Rapid Commun. Mass Spectrom. 2013, 27, 2009–2018. [Google Scholar] [CrossRef]
- Makarov, A.; Scigelova, M. Coupling liquid chromatography to Orbitrap mass spectrometry. J. Chromatogr. A 2010, 1217, 3938–3945. [Google Scholar] [CrossRef] [Green Version]
- De Angelis, E.; Pilolli, R.; Guagnano, R.; Monaci, L. Proteomics Applied to Food Allergen Research. In Comprehensive Foodomics; Elsevier, Ed.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 1, pp. 688–698. ISBN 9780128163955. [Google Scholar]
- Martinez-Esteso, M.J.; O’Connor, G.; Nørgaard, J.; Breidbach, A.; Brohée, M.; Cubero-Leon, E.; Nitride, C.; Robouch, P.; Emons, H. A reference method for determining the total allergenic protein content in a processed food: The case of milk in cookies as proof of concept. Anal. Bioanal. Chem. 2020, 412, 8249–8267. [Google Scholar] [CrossRef]
- Taylor, S.L.; Baumert, J.L.; Kruizinga, A.G.; Remington, B.C.; Crevel, R.W.R.; Brooke-Taylor, S.; Allen, K.J.; Houben, G. Establishment of Reference Doses for residues of allergenic foods: Report of the VITAL Expert Panel. Food Chem. Toxicol. 2014, 63, 9–17. [Google Scholar] [CrossRef]
- Turner, P.J.; Gowland, M.H.; Sharma, V.; Ierodiakonou, D.; Harper, N.; Garcez, T.; Pumphrey, R.; Boyle, R.J. Increase in anaphylaxis-related hospitalizations but no increase in fatalities: An analysis of United Kingdom national anaphylaxis data, 1992–2012. J. Allergy Clin. Immunol. 2015, 135, 956–963.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worm, M.; Moneret-Vautrin, A.; Scherer, K.; Lang, R.; Fernandez-Rivas, M.; Cardona, V.; Kowalski, M.L.; Jutel, M.; Poziomkowska-Gesicka, I.; Papadopoulos, N.G.; et al. First European data from the network of severe allergic reactions (NORA). Allergy Eur. J. Allergy Clin. Immunol. 2014, 69, 1397–1404. [Google Scholar] [CrossRef] [PubMed]
- New, L.S.; Stahl-Zeng, J.; Schreiber, A.; Cafazzo, M.; Liu, A.; Brunelle, S.; Liu, H.F. Detection and quantitation of selected food allergens by liquid chromatography with tandem mass spectrometry: First action 2017.17. J. AOAC Int. 2020, 103, 570–583. [Google Scholar] [CrossRef] [PubMed]
- Barreca, D.; Nabavi, S.M.; Sureda, A.; Rasekhian, M.; Raciti, R.; Sanches Silva, A.; Annunziata, G.; Arnone, A.; Tenore, G.C.; Süntar, I.; et al. Health Research Institute of the Balearic Islands (IdISBa), and CIBEROBN (Physiopathology of Obesity and Nutrition. Nutrients 2020, 12, 672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.Y.O.; Blumberg, J.B. In vitro activity of almond skin polyphenols for scavenging free radicals and inducing quinone reductase. J. Agric. Food Chem. 2008, 56, 4427–4434. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.J.A.; Kendall, C.W.C.; Marchie, A.; Josse, A.R.; Nguyen, T.H.; Faulkner, D.A.; Lapsley, K.G.; Singer, W. Effect of almonds on insulin secretion and insulin resistance in nondiabetic hyperlipidemic subjects: A randomized controlled crossover trial. Metabolism 2008, 57, 882–887. [Google Scholar] [CrossRef]
- Li, S.C.; Liu, Y.H.; Liu, J.F.; Chang, W.H.; Chen, C.M.; Chen, C.Y.O. Almond consumption improved glycemic control and lipid profiles in patients with type 2 diabetes mellitus. Metabolism 2011, 60, 474–479. [Google Scholar] [CrossRef]
- Hollis, J.; Mattes, R. Effect of chronic consumption of almonds on body weight in healthy humans. Br. J. Nutr. 2007, 98, 651–656. [Google Scholar] [CrossRef]
- Weigle, D.S.; Breen, P.A.; Matthys, C.C.; Callahan, H.S.; Meeuws, K.E.; Burden, V.R.; Purnell, J.Q. A high-protein diet induces sustained reductions in appetite, ad libitum caloric intake, and body weight despite compensatory changes in diurnal plasma leptin and ghrelin concentrations. Am. J. Clin. Nutr. 2005, 82, 41–48. [Google Scholar] [CrossRef]
- Fitzgerald, E.; Lambert, K.; Stanford, J.; Neale, E.P. The effect of nut consumption (tree nuts and peanuts) on the gut microbiota of humans: A systematic review. Br. J. Nutr. 2021, 125, 508–520. [Google Scholar] [CrossRef]
- Cetinkaya, P.G.; Buyuktiryaki, B.; Soyer, O.; Sahiner, U.M.; Sackesen, C.; Sekerel, B.E. Phenotypical characterization of tree nuts and peanut allergies in east Mediterranean children. Allergol. Immunopathol. 2020, 48, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jin, T. Almond allergens: Update and perspective on identification and characterization. J. Sci. Food Agric. 2020, 100, 4657–4663. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.; Mafra, I.; Carrapatoso, I.; Oliveira, M.B.P.P. Almond Allergens: Molecular Characterization, Detection, and Clinical Relevance. J. Agric. Food Chem. 2012, 60, 1337–1349. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Du, W.X.; Fregevu, C.; Kothary, M.H.; Harden, L.; McHugh, T.H. Expression, purification, and characterization of almond (Prunus dulcis) allergen Pru du 4. J. Agric. Food Chem. 2014, 62, 12695–12700. [Google Scholar] [CrossRef] [PubMed]
- Wensing, M.; Akkerdaas, J.H.; Van Leeuwen, W.A.; Stapel, S.O.; Bruijnzeel-Koomen, C.A.F.M.; Aalberse, R.C.; Bast, B.J.E.G.; Knulst, A.C.; Van Ree, R. IgE to Bet v 1 and profilin: Cross-reactivity patterns and clinical relevance. J. Allergy Clin. Immunol. 2002, 110, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.N.C.; Jenkins, J.; Marigheto, N.; Belton, P.S.; Gunning, A.P.; Morris, V.J. Allergens of the cupin superfamily. Biochem. Soc. Trans. 2002, 30, 925–929. [Google Scholar] [CrossRef] [Green Version]
- Che, H.; Zhang, Y.; Jiang, S.; Jin, T.; Lyu, S.C.; Nadeau, K.C.; McHugh, T. Almond (Prunus dulcis) Allergen Pru du 8, the First Member of a New Family of Food Allergens. J. Agric. Food Chem. 2019, 67, 8626–8631. [Google Scholar] [CrossRef]
- Sánchez-Pérez, R.; Pavan, S.; Mazzeo, R.; Moldovan, C.; Aiese Cigliano, R.; Del Cueto, J.; Ricciardi, F.; Lotti, C.; Ricciardi, L.; Dicenta, F.; et al. Mutation of a bHLH transcription factor allowed almond domestication. Science 2019, 364, 1095–1098. [Google Scholar] [CrossRef]
- Alioto, T.; Alexiou, K.G.; Bardil, A.; Barteri, F.; Castanera, R.; Cruz, F.; Dhingra, A.; Duval, H.; Fernández i Martí, Á.; Frias, L.; et al. Transposons played a major role in the diversification between the closely related almond and peach genomes: Results from the almond genome sequence. Plant J. 2020, 101, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Blais, B.W.; Gaudreault, M.; Philippe, L.M. Multiplex enzyme immunoassay system for the simulataneous detection of multiple allergens in foods. Food Control 2003, 14, 43–47. [Google Scholar] [CrossRef]
- Schubert-Ullrich, P.; Rudolf, J.; Ansari, P.; Galler, B.; Führer, M.; Molinelli, A.; Baumgartner, S. Commercialized rapid immunoanalytical tests for determination of allergenic food proteins: An overview. Anal. Bioanal. Chem. 2009, 395, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Garber, E.A.E.; Perry, J. Detection of hazelnuts and almonds using commercial ELISA test kits. Anal. Bioanal. Chem. 2010, 396, 1939–1945. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chhabra, G.S.; Zhao, J.; Zaffran, V.D.; Gupta, S.; Roux, K.H.; Gradziel, T.M.; Sathe, S.K. Comparison of Laboratory-Developed and Commercial Monoclonal Antibody-Based Sandwich Enzyme-Linked Immunosorbent Assays for Almond (Prunus dulcis) Detection and Quantification. J. Food Sci. 2017, 82, 2504–2515. [Google Scholar] [CrossRef] [PubMed]
- Slotwinski, E.; Almy, D.; Viator, R.; Abouzed, M.; Klein, F.; Rice, J. Development and Validation of a Quantitative ELISA for the Detection of Almond Residues in Foods. J. AOAC Int. 2018, 101, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Senyuva, H.Z.; Jones, I.B.; Sykes, M.; Baumgartner, S. A critical review of the specifications and performance of antibody and DNA-based methods for detection and quantification of allergens in foods. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2019, 36, 507–547. [Google Scholar] [CrossRef] [PubMed]
- Civera, A.; Galan-Malo, P.; Segura-Gil, I.; Mata, L.; Tobajas, A.P.; Sánchez, L.; Pérez, M.D. Development of sandwich ELISA and lateral flow immunoassay to detect almond in processed food. Food Chem. 2022, 371, 131338. [Google Scholar] [CrossRef]
- Bignardi, C.; Elviri, L.; Penna, A.; Careri, M.; Mangia, A. Particle-packed column versus silica-based monolithic column for liquid chromatography-electrospray-linear ion trap-tandem mass spectrometry multiallergen trace analysis in foods. J. Chromatogr. A 2010, 1217, 7579–7585. [Google Scholar] [CrossRef]
- Gu, S.; Chen, N.; Zhou, Y.; Zhao, C.; Zhan, L.; Qu, L.; Cao, C.; Han, L.; Deng, X.; Ding, T.; et al. A rapid solid-phase extraction combined with liquid chromatography-tandem mass spectrometry for simultaneous screening of multiple allergens in chocolates. Food Control 2018, 84, 89–96. [Google Scholar] [CrossRef]
- Alasalvar, C.; Shahidi, F.; Liyanapathirana, C.M.; Ohshima, T. Turkish Tombul hazelnut (Corylus avellana L.). 1. Compositional characteristics. J. Agric. Food Chem. 2003, 51, 3790–3796. [Google Scholar] [CrossRef]
- Di Nunzio, M. Hazelnuts as source of bioactive compounds and health value underestimated food. Curr. Res. Nutr. Food Sci. 2019, 7, 17–28. [Google Scholar] [CrossRef]
- Ortolani, C.; Ballmer-Weber, B.K.; Hansen, K.S.; Ispano, M.; Wüthrich, B.; Bindslev-Jensen, C.; Ansaloni, R.; Vannucci, L.; Pravettoni, V.; Scibilia, J.; et al. Hazelnut allergy: A double-blind, placebo-controlled food challenge multicenter study. J. Allergy Clin. Immunol. 2000, 105, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Waserman, S.; Bégin, P.; Watson, W. IgE-mediated food allergy. Allergy Asthma Clin. Immunol. 2018, 14, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masthoff, L.J.; van Hoffen, E.; de Reus, A.; Boonacker, C.W.; Bruijnzeel-Koomen, C.A.; Pasmans, S.G.; Knulst, A.C. Hazelnut allergy differs between children and adults in frequency of severity, aetiology and relevance of diagnostic parameters. Clin. Exp. Allergy 2014, 44, 1539–1545. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, C.; Berthold, M.; Mascialino, B.; Orme, M.; Sjölander, S.; Hamilton, R. Allergen components in diagnosing childhood hazelnut allergy: Systematic literature review and meta-analysis. Pediatr. Allergy Immunol. 2020, 31, 186–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burney, P.; Summers, C.; Chinn, S.; Hooper, R.; Van Ree, R.; Lidholm, J. Prevalence and distribution of sensitization to foods in the European Community Respiratory Health Survey: A EuroPrevall analysis. Allergy Eur. J. Allergy Clin. Immunol. 2010, 65, 1182–1188. [Google Scholar] [CrossRef] [PubMed]
- Datema, M.R.; van Ree, R.; Asero, R.; Barreales, L.; Belohlavkova, S.; de Blay, F.; Clausen, M.; Dubakiene, R.; Fernández-Perez, C.; Fritsche, P.; et al. Component-resolved diagnosis and beyond: Multivariable regression models to predict severity of hazelnut allergy. Allergy Eur. J. Allergy Clin. Immunol. 2018, 73, 549–559. [Google Scholar] [CrossRef]
- Ribeiro, M.; Costa, J.; Mafra, I.; Cabo, S.; Silva, A.P.; Gonçalves, B.; Hillion, M.; Hébraud, M.; Igrejas, G. Natural variation of hazelnut allergenicity: Is there any potential for selecting hypoallergenic varieties? Nutrients 2020, 12, 2100. [Google Scholar] [CrossRef]
- Pfeifer, S.; Bublin, M.; Dubiela, P.; Hummel, K.; Wortmann, J.; Hofer, G.; Keller, W.; Radauer, C.; Hoffmann-Sommergruber, K. Cor a 14, the allergenic 2S albumin from hazelnut, is highly thermostable and resistant to gastrointestinal digestion. Mol. Nutr. Food Res. 2015, 59, 2077–2086. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.; Mafra, I.; Carrapatoso, I.; Oliveira, M.B.P.P. Hazelnut allergens: Molecular characterization, detection, and clinical relevance. Crit. Rev. Food Sci. Nutr. 2016, 56, 2579–2605. [Google Scholar] [CrossRef] [Green Version]
- Cucu, T.; Platteau, C.; Taverniers, I.; Devreese, B.; de Loose, M.; de Meulenaer, B. ELISA detection of hazelnut proteins: Effect of protein glycation in the presence or absence of wheat proteins. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2011, 28, 1–10. [Google Scholar] [CrossRef]
- Costa, J.; Ansari, P.; Mafra, I.; Oliveira, M.B.P.P.; Baumgartner, S. Development of a sandwich ELISA-type system for the detection and quantification of hazelnut in model chocolates. Food Chem. 2015, 173, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Ehlert, A.; Demmel, A.; Hupfer, C.; Busch, U.; Engel, K.H. Simultaneous detection of DNA from 10 food allergens by ligation-dependent probe amplification. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2009, 26, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Manny, E.; La Vieille, S.; Barrere, V.; Théolier, J.; Godefroy, S.B. Peanut and hazelnut occurrence as allergens in foodstuffs with precautionary allergen labeling in Canada. NPJ Sci. Food 2021, 5, 11. [Google Scholar] [CrossRef] [PubMed]
- Arlorio, M.; Coisson, J.D.; Bordiga, M.; Travaglia, F.; Garino, C.; Zuidmeer, L.; van Ree, R.; Giuffrida, M.G.; Conti, A.; Martelli, A. Olive oil adulterated with hazelnut oils: Simulation to identify possible risks to allergic consumers. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2010, 27, 11–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansari, P.; Stoppacher, N.; Baumgartner, S. Marker peptide selection for the determination of hazelnut by LC-MS/MS and occurrence in other nuts. Anal. Bioanal. Chem. 2012, 402, 2607–2615. [Google Scholar] [CrossRef] [PubMed]
- Van Vlierberghe, K.; Gavage, M.; Dieu, M.; Renard, P.; Arnould, T.; Gillard, N.; Coudijzer, K.; De Loose, M.; Gevaert, K.; Van Poucke, C. Selection of universal peptide biomarkers for the detection of the allergen hazelnut in food trough a comprehensive, high resolution mass spectrometric (HRMS) based approach. Food Chem. 2019, 309, 125679. [Google Scholar] [CrossRef]
- Gomaa, A.; Boye, J. Simultaneous detection of multi-allergens in an incurred food matrix using ELISA, multiplex flow cytometry and liquid chromatography mass spectrometry (LC-MS). Food Chem. 2015, 175, 585–592. [Google Scholar] [CrossRef]
- Planque, M.; Arnould, T.; Delahaut, P.; Renard, P.; Dieu, M.; Gillard, N. Development of a strategy for the quantification of food allergens in several food products by mass spectrometry in a routine laboratory. Food Chem. 2019, 274, 35–45. [Google Scholar] [CrossRef]
- Pilolli, R.; De Angelis, E.; Monaci, L. Streamlining the analytical workflow for multiplex MS/MS allergen detection in processed foods. Food Chem. 2017, 221, 1747–1753. [Google Scholar] [CrossRef]
- Pilolli, R.; De Angelis, E.; Monaci, L. In house validation of a high resolution mass spectrometry Orbitrap-based method for multiple allergen detection in a processed model food. Anal. Bioanal. Chem. 2018, 410, 5653–5662. [Google Scholar] [CrossRef]
- Asogwa, E.U.; Hammed, L.A.; Ndubuaku, T.C.N. Integrated production and protection practices of cashew (Anacardium occidentale) in Nigeria. Afr. J. Biotechnol. 2008, 7, 4868–4873. [Google Scholar] [CrossRef]
- Mah, E.; Schulz, J.A.; Kaden, V.N.; Lawless, A.L.; Rotor, J.; Mantilla, L.B.; Liska, D.J. Cashew consumption reduces total and LDL cholesterol: A randomized, crossover, controlled-feeding trial. Am. J. Clin. Nutr. 2017, 105, 1070–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ras, R.T.; Geleijnse, J.M.; Trautwein, E.A. LDL-cholesterol-lowering effect of plant sterols and stanols across different dose ranges: A meta-analysis of randomised controlled studies. Br. J. Nutr. 2014, 112, 214–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trox, J.; Vadivel, V.; Vetter, W.; Stuetz, W.; Kammerer, D.R.; Carle, R.; Scherbaum, V.; Gola, U.; Nohr, D.; Biesalski, H.K. Catechin and epicatechin in testa and their association with bioactive compounds in kernels of cashew nut (Anacardium occidentale L.). Food Chem. 2011, 128, 1094–1099. [Google Scholar] [CrossRef]
- Medeiros-Linard, C.F.B.; Andrade-da-Costa, B.L.D.S.; Augusto, R.L.; Sereniki, A.; Trevisan, M.T.S.; Perreira, R.D.C.R.; de Souza, F.T.C.; Braz, G.R.F.; Lagranha, C.J.; de Souza, I.A.; et al. Anacardic Acids from Cashew Nuts Prevent Behavioral Changes and Oxidative Stress Induced by Rotenone in a Rat Model of Parkinson’s Disease. Neurotox. Res. 2018, 34, 250–262. [Google Scholar] [CrossRef] [PubMed]
- Rico, R.; Bulló, M.; Salas-Salvadó, J. Nutritional composition of raw fresh cashew (Anacardium occidentale L.) kernels from different origin. Food Sci. Nutr. 2016, 4, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Alasalvar, C.; Shahidi, F. Tree Nuts: Composition, Phytochemicals, and Health Effects, 1st ed.; CRC Press: Boca Raton, FL, USA, 2008; ISBN 9780849337352. [Google Scholar]
- Ros, E. Health benefits of nut consumption. Nutrients 2010, 2, 652–682. [Google Scholar] [CrossRef] [Green Version]
- Rancé, F.; Bidat, E.; Bourrier, T.; Sabouraud, D. Cashew allergy: Observations of 42 children without associated peanut allergy. Allergy Eur. J. Allergy Clin. Immunol. 2003, 58, 1311–1314. [Google Scholar] [CrossRef]
- Van Der Valk, J.P.M.; Dubois, A.E.J.; Gerth Van Wijk, R.; Wichers, H.J.; De Jong, N.W. Systematic review on cashew nut allergy. Allergy Eur. J. Allergy Clin. Immunol. 2014, 69, 692–698. [Google Scholar] [CrossRef]
- Inoue, T.; Ogura, K.; Takahashi, K.; Nishino, M.; Asaumi, T.; Yanagida, N.; Sato, S.; Ebisawa, M. Risk factors and clinical features in cashew nut oral food challenges. Int. Arch. Allergy Immunol. 2018, 175, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Chitta, S.; Lian, B.X.; Rao, R.; Loh, W.; Goh, A.; Chong, K.W. Cashew nut allergy in Singaporean children. Asia Pac. Allergy 2018, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Davoren, M.; Peake, J. Cashew nut allergy is associated with a high risk of anaphylaxis. Arch. Dis. Child. 2005, 90, 1084–1085. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.; Inomata, N.; Yamazaki, H.; Morita, A.; Kirino, M.; Ikezawa, Z. Clinical features of four cases with cashew nut allergy and cross-reactivity between cashew nut and pistachio. Allergol. Int. 2009, 58, 209–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robotham, J.M.; Wang, F.; Seamon, V.; Teuber, S.S.; Sathe, S.K.; Sampson, H.A.; Beyer, K.; Seavy, M.; Roux, K.H. Ana o 3, an important cashew nut (Anacardium occidentale L.) allergen of the 2S albumin family. J. Allergy Clin. Immunol. 2005, 115, 1284–1290. [Google Scholar] [CrossRef] [PubMed]
- De Leon, M.P.; Glaspole, I.N.; Drew, A.C.; Rolland, J.M.; O’Hehir, R.E.; Suphioglu, C. Immunological analysis of allergenic cross-reactivity between peanut and tree nuts. Clin. Exp. Allergy 2003, 33, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Barre, A.; Sordet, C.; Culerrier, R.; Rancé, F.; Didier, A.; Rougé, P. Vicilin allergens of peanut and tree nuts (walnut, hazelnut and cashew nut) share structurally related IgE-binding epitopes. Mol. Immunol. 2008, 45, 1231–1240. [Google Scholar] [CrossRef]
- Rougé, P.; Thibau, F.; Bourrier, T.; Saggio, B.; Culerrier, R.; Rancé, F.; Barre, A. Signification clinique des allergènes croisants de la noix de Cajou (Anacardium occidentale). Rev. Fr. Allergol. 2011, 51, 31–35. [Google Scholar] [CrossRef]
- Willison, L.N.; Tawde, P.; Robotham, J.M.; Penney, R.M.; Teuber, S.S.; Sathe, S.K.; Roux, K.H. Pistachio vicilin, Pis v 3, is immunoglobulin E-reactive and cross-reacts with the homologous cashew allergen, Ana o 1. Clin. Exp. Allergy 2008, 38, 1229–1238. [Google Scholar] [CrossRef]
- Wang, F.; Robotham, J.M.; Teuber, S.S.; Sathe, S.K.; Roux, K.H. Ana o 2, a major cashew (Anacardium occidentale L.) nut allergen of the legumin family. Int. Arch. Allergy Immunol. 2003, 132, 27–39. [Google Scholar] [CrossRef]
- Robotham, J.M.; Hoffman, G.G.; Teuber, S.S.; Beyer, K.; Sampson, H.A.; Sathe, S.K.; Roux, K.H. Linear IgE-epitope mapping and comparative structural homology modeling of hazelnut and English walnut 11S globulins. Mol. Immunol. 2009, 46, 2975–2984. [Google Scholar] [CrossRef]
- Teuber, S.S.; Sathe, S.K.; Peterson, W.R.; Roux, K.H. Characterization of the soluble allergenic proteins of cashew nut (Anacardium occidentale L.). J. Agric. Food Chem. 2002, 50, 6543–6549. [Google Scholar] [CrossRef] [PubMed]
- Ahn, K.; Bardina, L.; Grishina, G.; Beyer, K.; Sampson, H.A. Identification of two pistachio allergens, Pis v 1 and Pis v 2, belonging to the 2S albumin and 11S globulin family. Clin. Exp. Allergy 2009, 39, 926–934. [Google Scholar] [CrossRef] [PubMed]
- Mendes, C.; Costa, J.; Vicente, A.A.; Oliveira, M.B.P.P.; Mafra, I. Cashew Nut Allergy: Clinical Relevance and Allergen Characterisation. Clin. Rev. Allergy Immunol. 2019, 57, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Rejeb, S.; Abbott, M.; Davies, D.; Cléroux, C.; Delahaut, P. Multi-allergen screening immunoassay for the detection of protein markers of peanut and four tree nuts in chocolate. Food Addit. Contam. 2005, 22, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Sun, X.; Marquis, C.P.; Lee, N.A. Development of a sensitive sandwich ELISA specific to 2S albumin (Ana o 3) as a stable protein marker for cashew nut residue detection in pre-packaged food products. Food Control 2019, 96, 432–440. [Google Scholar] [CrossRef]
- Brzezinski, J.L. Detection of cashew Nut DNA in spiked baked goods using a real-time polymerase chain reaction method. J. AOAC Int. 2006, 89, 1035–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Calleja, I.M.; de la Cruz, S.; González, I.; García, T.; Martín, R. Development of real-time PCR assays to detect cashew (Anacardium occidentale) and macadamia (Macadamia intergrifolia) residues in market analysis of processed food products. LWT 2015, 62, 233–241. [Google Scholar] [CrossRef]
- Bignardi, C.; Mattarozzi, M.; Penna, A.; Sidoli, S.; Elviri, L.; Careri, M.; Mangia, A. A Rapid Size-Exclusion Solid-Phase Extraction Step for Enhanced Sensitivity in Multi-Allergen Determination in Dark Chocolate and Biscuits by Liquid Chromatography-Tandem Mass Spectrometry. Food Anal. Methods 2013, 6, 1144–1152. [Google Scholar] [CrossRef]
- Korte, R.; Oberleitner, D.; Brockmeyer, J. Determination of food allergens by LC-MS: Impacts of sample preparation, food matrix, and thermal processing on peptide detectability and quantification. J. Proteom. 2019, 196, 131–140. [Google Scholar] [CrossRef]
- Costa, J.; Silva, I.; Vicente, A.A.; Oliveira, M.B.P.P.; Mafra, I. Pistachio nut allergy: An updated overview. Crit. Rev. Food Sci. Nutr. 2019, 59, 546–562. [Google Scholar] [CrossRef] [Green Version]
- Mao, R.; Xie, K.; Zhao, M.; Li, M.; Lu, L.; Liu, Y.; Wu, Q.; Chen, Y.; Zhang, T.; Diao, E. Development and Evaluation of a Loop-Mediated Isothermal Amplification (LAMP) Assay for Rapid Detection of Pistachio (Pistacia vera) in Food Samples. Food Anal. Methods 2020, 13, 658–666. [Google Scholar] [CrossRef]
- Vadivel, V.; Kunyanga, C.N.; Biesalski, H.K. Health benefits of nut consumption with special reference to body weight control. Nutrition 2012, 28, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Dreher, M.L. Pistachio nuts: Composition and potential health benefits. Nutr. Rev. 2012, 70, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Kashaninejad, M.; Tabil, L.G. Pistachio (Pistacia vera L.). In Postharvest Biology and Technology of Tropical and Subtropical Fruits; Yahia, E.M., Ed.; Woodhead Publishing Limited: Sawston, UK, 2011; Volume 4, pp. 218–247. ISBN 9780857090904. [Google Scholar]
- Sheridan, M.J.; Cooper, J.N.; Erario, M.; Cheifetz, C.E. Pistachio nut consumption and serum lipid levels. J. Am. Coll. Nutr. 2007, 26, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Kaur, A. Plantation Crops and Tree Nuts: Composition, Phytochemicals and Health Benefits. In Handbook of Plant Food Phytochemicals: Sources, Stability and Extraction; Tiwari, B.K., Brunton, N.P., Brennan, C.S., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 163–179. ISBN 9781444338102. [Google Scholar]
- Tomaino, A.; Martorana, M.; Arcoraci, T.; Monteleone, D.; Giovinazzo, C.; Saija, A. Antioxidant activity and phenolic profile of pistachio (Pistacia vera L., variety Bronte) seeds and skins. Biochimie 2010, 92, 1115–1122. [Google Scholar] [CrossRef] [PubMed]
- Ando, K.; Watanabe, D.; Tamada, Y.; Matsumoto, Y. Oral allergy syndrome with severe anaphylaxis induced by pistachio. Int. J. Dermatol. 2011, 50, 632–633. [Google Scholar] [CrossRef]
- Bock, S.A.; Muoz-Furlong, A.; Sampson, H.A. Fatalities due to anaphylactic reactions to foods. J. Allergy Clin. Immunol. 2001, 107, 191–193. [Google Scholar] [CrossRef]
- Geiselhart, S.; Hoffmann-Sommergruber, K.; Bublin, M. Tree nut allergens. Mol. Immunol. 2018, 100, 71–81. [Google Scholar] [CrossRef]
- Andorf, S.; Borres, M.P.; Block, W.; Tupa, D.; Bollyky, J.B.; Sampath, V.; Elizur, A.; Lidholm, J.; Jones, J.E.; Galli, S.J.; et al. Association of Clinical Reactivity with Sensitization to Allergen Components in Multifood-Allergic Children. J. Allergy Clin. Immunol. Pract. 2017, 5, 1325–1334.e4. [Google Scholar] [CrossRef]
- Elizur, A.; Appel, M.Y.; Nachshon, L.; Levy, M.B.; Epstein-Rigbi, N.; Golobov, K.; Goldberg, M.R. NUT Co Reactivity-ACquiring Knowledge for Elimination Recommendations (NUT CRACKER) study. Allergy Eur. J. Allergy Clin. Immunol. 2018, 73, 593–601. [Google Scholar] [CrossRef]
- Noorbakhsh, R.; Mortazavi, S.A.; Sankian, M.; Shahidi, F.; Assarehzadegan, M.A.; Varasteh, A.R. Cloning, expression, characterization, and computational approach for cross-reactivity prediction of manganese superoxide dismutase allergen from pistachio nut. Allergol. Int. 2010, 59, 295–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maloney, J.M.; Rudengren, M.; Ahlstedt, S.; Bock, S.A.; Sampson, H.A. The use of serum-specific IgE measurements for the diagnosis of peanut, tree nut, and seed allergy. J. Allergy Clin. Immunol. 2008, 122, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Ayuso, R.; Grishina, G.; Ahn, K.; Bardina, L.; Beyer, K.; Sampson, H. Identification of a MnSOD-like Protein as a New Major Pistachio Allergen. J. Allergy Clin. Immunol. 2007, 119, S115. [Google Scholar] [CrossRef]
- Liu, C.; Chhabra, G.S.; Sathe, S.K. Pistachio (Pistacia vera L.) Detection and Quantification Using a Murine Monoclonal Antibody-Based Direct Sandwich Enzyme-Linked Immunosorbent Assay. J. Agric. Food Chem. 2015, 63, 9139–9149. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Wu, J.; Zhang, J.; Pan, A.; Quan, S.; Zhang, D.; Kim, H.Y.; Li, X.; Zhou, S.; Yang, L. Development and inter-laboratory transfer of a decaplex polymerase chain reaction assay combined with capillary electrophoresis for the simultaneous detection of ten food allergens. Food Chem. 2016, 199, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Sealey-Voyksner, J.; Zweigenbaum, J.; Voyksner, R. Discovery of highly conserved unique peanut and tree nut peptides by LC-MS/MS for multi-allergen detection. Food Chem. 2016, 194, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Korte, R.; Brockmeyer, J. MRM3-based LC-MS multi-method for the detection and quantification of nut allergens. Anal. Bioanal. Chem. 2016, 408, 7845–7855. [Google Scholar] [CrossRef]
- Settaluri, V.S.; Kandala, C.V.K.; Puppala, N.; Sundaram, J. Peanuts and Their Nutritional Aspects—A Review. Food Nutr. Sci. 2012, 3, 1644–1650. [Google Scholar] [CrossRef] [Green Version]
- Davis, L.; Stonehouse, W.; Loots, D.T.; Mukuddem-Petersen, J.; Van Der Westhuizen, F.H.; Hanekom, S.M.; Jerling, J.C. The effects of high walnut and cashew nut diets on the antioxidant status of subjects with metabolic syndrome. Eur. J. Nutr. 2007, 46, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Lavedrine, F.; Zmirou, D.; Ravel, A.; Balducci, F.; Alary, J. Blood cholesterol and walnut consumption: A cross-sectional survey in France. Prev. Med. 1999, 28, 333–339. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Ros, E.; Núñez, I.; Pérez-Heras, A.; Serra, M.; Gilabert, R.; Casals, E.; Deulofeu, R. A Walnut Diet Improves Endothelial Function in Hypercholesterolemic Subjects: A Randomized Crossover Trial. Circulation 2004, 109, 1609–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, K.J.; Teuber, S.S.; Gobeille, A.; Cremin, P.; Waterhouse, A.L.; Steinberg, F.M. Walnut polyphenolics inhibit in vitro human plasma and LDL oxidation. J. Nutr. 2001, 131, 2837–2842. [Google Scholar] [CrossRef] [PubMed]
- Lyons, S.A.; Datema, M.R.; Le, T.M.; Asero, R.; Barreales, L.; Belohlavkova, S.; de Blay, F.; Clausen, M.; Dubakiene, R.; Fernández-Perez, C.; et al. Walnut Allergy across Europe: Distribution of Allergen Sensitization Patterns and Prediction of Severity. J. Allergy Clin. Immunol. Pract. 2021, 9, 225–235. [Google Scholar] [CrossRef]
- Costa, J.; Carrapatoso, I.; Oliveira, M.B.P.P.; Mafra, I. Walnut allergens: Molecular characterization, detection and clinical relevance. Clin. Exp. Allergy 2014, 44, 319–341.e10. [Google Scholar] [CrossRef] [PubMed]
- Teuber, S.S.; Dandekar, A.M.; Peterson, W.R.; Sellers, C.L. Cloning and sequencing of a gene encoding a 2S albumin seed storage protein precursor from English walnut (Juglans regia), a major food allergen. J. Allergy Clin. Immunol. 1998, 101, 807–814. [Google Scholar] [CrossRef]
- Sordet, C.; Culerrier, R.; Granier, C.; Rancé, F.; Didier, A.; Barre, A.; Rougé, P. Expression of Jug r 1, the 2S albumin allergen from walnut (Juglans regia), as a correctly folded and functional recombinant protein. Peptides 2009, 30, 1213–1221. [Google Scholar] [CrossRef]
- Hauser, M.; Roulias, A.; Ferreira, F.; Egger, M. Panallergens and their impact on the allergic patient. Allergy Asthma Clin. Immunol. 2010, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Pastorello, E.A.; Farioli, L.; Pravettoni, V.; Robino, A.M.; Scibilia, J.; Fortunato, D.; Conti, A.; Borgonovo, L.; Bengtsson, A.; Ortolani, C. Lipid transfer protein and vicilin are important walnut allergens in patients not allergic to pollen. J. Allergy Clin. Immunol. 2004, 114, 908–914. [Google Scholar] [CrossRef]
- López, E.; Cuadrado, C.; Burbano, C.; Jiménez, M.; Rodríguez, J.; Crespo, J.F. Effects of autoclaving and high pressure on allergenicity of hazelnut proteins. J. Clin. Bioinforma. 2012, 2, 12. [Google Scholar] [CrossRef] [Green Version]
- Su, M.; Venkatachalam, M.; Teuber, S.S.; Roux, K.H.; Sathe, S.K. Impact of γ-irradiation and thermal processing on the antigenicity of almond, cashew nut and walnut proteins. J. Sci. Food Agric. 2004, 84, 1119–1125. [Google Scholar] [CrossRef]
- Surojanametakul, V.; Doi, H.; Shibata, H.; Mizumura, T.; Takahashi, T.; Varanyanond, W.; Wannapinpong, S.; Shoji, M.; Ito, T.; Tamura, H. Reliable enzyme-linked immunosorbent assay for the determination of coconut milk proteins in processed foods. J. Agric. Food Chem. 2011, 59, 2131–2136. [Google Scholar] [CrossRef] [PubMed]
- Sakai, S.; Adachi, R.; Akiyama, H.; Teshima, R.; Doi, H.; Shibata, H.; Urisu, A. Determination of walnut protein in processed foods by enzyme-linked immunosorbent assay: Interlaboratory study. J. AOAC Int. 2010, 93, 1255–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niemann, L.; Taylor, S.L.; Hefle, S.L. Detection of walnut residues in foods using an enzyme-linked immunosorbent assay. J. Food Sci. 2009, 74, T51–T57. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, G.; Wu, Y.; Yuan, F.; Chen, Y. Development of an indirect competitive immunoassay for walnut protein component in food. Food Chem. 2014, 147, 106–110. [Google Scholar] [CrossRef]
- Madrid, R.; García-García, A.; Cabrera, P.; González, I.; Martín, R.; García, T. Survey of commercial food products for detection of walnut (Juglans regia) by two elisa methods and real time pcr. Foods 2021, 10, 440. [Google Scholar] [CrossRef] [PubMed]
- Haiyan, W.; Fei, Y.; Yajun, W.; Hairong, Y.; Baoliang, X.; Zhongxue, L.; Ying, C. Detection of allergen walnut component in food by an improved real-time PCR method. J. Food Prot. 2009, 72, 2433–2435. [Google Scholar] [CrossRef]
- Brežná, B.; Dudášová, H.; Kuchta, T. A novel real-time polymerase chain reaction method for the qualitative detection of pistachio in food. Eur. Food Res. Technol. 2008, 228, 197–203. [Google Scholar] [CrossRef]
- Janská, V.; Piknová, L.; Kuchta, T. Relative quantification of walnuts and hazelnuts in bakery products using real-time polymerase chain reaction. Eur. Food Res. Technol. 2011, 232, 1057–1060. [Google Scholar] [CrossRef]
- Yano, T.; Sakai, Y.; Uchida, K.; Nakao, Y.; Ishihata, K.; Nakano, S.; Yamada, T.; Sakai, S.; Urisu, A.; Akiyama, H.; et al. Detection of walnut residues in processed foods by polymerase chain reaction. Biosci. Biotechnol. Biochem. 2007, 71, 1793–1796. [Google Scholar] [CrossRef] [Green Version]
- Planque, M.; Arnould, T.; Dieu, M.; Delahaut, P.; Renard, P.; Gillard, N. Advances in ultra-high performance liquid chromatography coupled to tandem mass spectrometry for sensitive detection of several food allergens in complex and processed foodstuffs. J. Chromatogr. A 2016, 1464, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Toomer, O.T. Nutritional chemistry of the peanut (Arachis hypogaea). Crit. Rev. Food Sci. Nutr. 2018, 58, 3042–3053. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Chen, J.; Du, F. Potential use of peanut by-products in food processing: A review. J. Food Sci. Technol. 2012, 49, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.B.; Provost, M.; Keaver, L.; Breen, C.; Ludy, M.J.; Mattes, R.D. A randomized trial on the effects of flavorings on the health benefits of daily peanut consumption. Am. J. Clin. Nutr. 2014, 99, 490–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, C.A.; Salas-Salvadó, J. The potential of nuts in the prevention of cancer. Br. J. Nutr. 2006, 96, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.P.; Johnston, C.A.; El-Mubasher, A.A.; Papaioannou, M.A.; Tyler, C.; Gee, M.; Foreyt, J.P. Peanut consumption in adolescents is associated with improved weight status. Nutr. Res. 2013, 33, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Bouchenak, M.; Lamri-Senhadji, M. Nutritional quality of legumes, and their role in cardiometabolic risk prevention: A review. J. Med. Food 2013, 16, 185–198. [Google Scholar] [CrossRef]
- Burks, A.W.; Tang, M.; Sicherer, S.; Muraro, A.; Eigenmann, P.A.; Ebisawa, M.; Fiocchi, A.; Chiang, W.; Beyer, K.; Wood, R.; et al. ICON: Food allergy. J. Allergy Clin. Immunol. 2012, 129, 906–920. [Google Scholar] [CrossRef]
- Dyer, A.A.; Rivkina, V.; Perumal, D.; Smeltzer, B.M.; Smith, B.M.; Gupta, R.S. Epidemiology of childhood peanut allergy. Allergy Asthma Proc. 2015, 36, 58–64. [Google Scholar] [CrossRef]
- Sicherer, S.H.; Furlong, T.J.; Muñoz-Furlong, A.; Burks, A.W.; Sampson, H.A. A voluntary registry for peanut and tree nut allergy: Characteristics of the first 5149 registrants. J. Allergy Clin. Immunol. 2001, 108, 128–132. [Google Scholar] [CrossRef] [Green Version]
- Palladino, C.; Breiteneder, H. Peanut allergens. Mol. Immunol. 2018, 100, 58–70. [Google Scholar] [CrossRef] [PubMed]
- Porterfield, H.S.; Murray, K.S.; Schlichting, D.G.; Chen, X.; Hansen, K.C.; Duncan, M.W.; Dreskin, S.C. Effector activity of peanut allergens: A critical role for Ara h 2, Ara h 6, and their variants. Clin. Exp. Allergy 2009, 39, 1099–1108. [Google Scholar] [CrossRef] [Green Version]
- Nicolaou, N.; Murray, C.; Belgrave, D.; Poorafshar, M.; Simpson, A.; Custovic, A. Quantification of specific IgE to whole peanut extract and peanut components in prediction of peanut allergy. J. Allergy Clin. Immunol. 2011, 127, 684–685. [Google Scholar] [CrossRef] [PubMed]
- Suhr, M.; Wicklein, D.; Lepp, U.; Becker, W.M. Isolation and characterization of natural Ara h 6: Evidence for a further peanut allergen with putative clinical relevance based on resistance to pepsin digestion and heat. Mol. Nutr. Food Res. 2004, 48, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, K.; Schweimer, K.; Reese, G.; Randow, S.; Suhr, M.; Becker, W.M.; Vieths, S.; Rösch, P. Structure and stability of 2S albumin-type peanut allergens: Implications for the severity of peanut allergic reactions. Biochem. J. 2006, 395, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, A.; Shah, F.; Hamayun, M.; Ahmad, A.; Hussain, A.; Waqas, M.; Kang, S.M.; Lee, I.J. Allergens of Arachis hypogaeaand the effect of processing on their detection by ELISA. Food Nutr. Res. 2016, 60, 300–307. [Google Scholar] [CrossRef] [Green Version]
- Toomer, O.T.; Do, A.; Pereira, M.; Williams, K. Effect of simulated gastric and intestinal digestion on temporal stability and immunoreactivity of peanut, almond, and pine nut protein allergens. J. Agric. Food Chem. 2013, 61, 5903–5913. [Google Scholar] [CrossRef]
- Pele, M.; Brohée, M.; Anklam, E.; van Hengel, A.J. Peanut and hazelnut traces in cookies and chocolates: Relationship between analytical results and declaration of food allergens on product labels. Food Addit. Contam. 2007, 24, 1334–1344. [Google Scholar] [CrossRef] [Green Version]
- Poms, R.E.; Agazzi, M.E.; Bau, A.; Brohee, M.; Capelletti, C.; Nørgaard, J.V.; Anklam, E. Inter-laboratory validation study of five commercial ELISA test kits for the determination of peanut proteins in biscuits and dark chocolate. Food Addit. Contam. 2005, 22, 104–112. [Google Scholar] [CrossRef]
- Montserrat, M.; Sanz, D.; Juan, T.; Herrero, A.; Sánchez, L.; Calvo, M.; Pérez, M.D. Detection of peanut (Arachis hypogaea) allergens in processed foods by immunoassay: Influence of selected target protein and ELISA format applied. Food Control 2015, 54, 300–307. [Google Scholar] [CrossRef] [Green Version]
- Sheridan, M.J.; Koeberl, M.; Hedges, C.E.; Biros, E.; Ruethers, T.; Clarke, D.; Buddhadasa, S.; Kamath, S.; Lopata, A.L. Undeclared allergens in imported packaged food for retail in Australia. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2020, 37, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Scaravelli, E.; Brohée, M.; Marchelli, R.; Van Hengel, A.J. The effect of heat treatment on the detection of peanut allergens as determined by ELISA and real-time PCR. Anal. Bioanal. Chem. 2009, 395, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Hird, H.; Lloyd, J.; Goodier, R.; Brown, J.; Reece, P. Detection of peanut using real-time polymerase chain reaction. Eur. Food Res. Technol. 2003, 217, 265–268. [Google Scholar] [CrossRef]
- Stephan, O.; Vieths, S. Development of a real-time PCR and a sandwich ELISA for detection of potentially allergenic trace amounts of peanut (Arachis hypogaea) in processed foods. J. Agric. Food Chem. 2004, 52, 3754–3760. [Google Scholar] [CrossRef] [PubMed]
- Scaravelli, E.; Brohée, M.; Marchelli, R.; Van Hengel, A.J. Development of three real-time PCR assays to detect peanut allergen residue in processed food products. Eur. Food Res. Technol. 2008, 227, 857–869. [Google Scholar] [CrossRef]
- Watanabe, T.; Akiyama, H.; Maleki, S.; Yamakawa, H.; Iijima, K.; Yamazaki, F.; Matsumoto, T.; Futo, S.; Arakawa, F.; Watai, M.; et al. A specific qualitative detection method for peanut (arachis hypogaea) in foods using polymerase chain reaction. J. Food Biochem. 2006, 30, 215–233. [Google Scholar] [CrossRef]
- Shefcheck, K.J.; Callahan, J.H.; Musser, S.M. Confirmation of peanut protein using peptide markers in dark chocolate using liquid chromatography-tandem mass spectrometry (LC-MS/MS). J. Agric. Food Chem. 2006, 54, 7953–7959. [Google Scholar] [CrossRef]
- Careri, M.; Costa, A.; Elviri, L.; Lagos, J.B.; Mangia, A.; Terenghi, M.; Cereti, A.; Garoffo, L.P. Use of specific peptide biomarkers for quantitative confirmation of hidden allergenic peanut proteins Ara h 2 and Ara h 3/4 for food control by liquid chromatography-tandem mass spectrometry. Anal. Bioanal. Chem. 2007, 389, 1901–1907. [Google Scholar] [CrossRef]
- Zhang, J.; Hong, Y.; Cai, Z.; Huang, B.; Wang, J.; Ren, Y. Simultaneous determination of major peanut allergens Ara h 1 and Ara h 2 in baked foodstuffs based on their signature peptides using ultra-performance liquid chromatography coupled to tandem mass spectrometry. Anal. Methods 2019, 11, 1689–1696. [Google Scholar] [CrossRef]
- Chang, Y.; Peng, H.; Zhang, G. Development of a simplified, sensitive, and accurate LC-MS/MS method for peanut quantification in wheat flour-based dry matrices. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2021, 38, 1260–1272. [Google Scholar] [CrossRef]
- Monaci, L.; Pilolli, R.; De Angelis, E.; Carone, R.; Pascale, M. LC-tandem mass spectrometry as a screening tool for multiple detection of allergenic ingredients in complex foods. Acta IMEKO 2016, 5, 5–9. [Google Scholar] [CrossRef]
- Boo, C.C.; Parker, C.H.; Jackson, L.S. A targeted LC-MS/MS method for the simultaneous detection and quantitation of egg, milk, and peanut allergens in sugar cookies. J. AOAC Int. 2018, 101, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Monaci, L.; De Angelis, E.; Bavaro, S.L.; Pilolli, R. High-resolution OrbitrapTM-based mass spectrometry for rapid detection of peanuts in nuts. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2015, 32, 1607–1616. [Google Scholar] [CrossRef] [PubMed]
- Gavage, M.; Van Vlierberghe, K.; Van Poucke, C.; De Loose, M.; Gevaert, K.; Dieu, M.; Renard, P.; Arnould, T.; Gillard, N. High-resolution mass spectrometry-based selection of peanut peptide biomarkers considering food processing and market type variation. Food Chem. 2020, 304, 125428. [Google Scholar] [CrossRef] [PubMed]
- Gavage, M.; Van Vlierberghe, K.; Van Poucke, C.; De Loose, M.; Gevaert, K.; Dieu, M.; Renard, P.; Arnould, T.; Filee, P.; Gillard, N. Comparative study of concatemer efficiency as an isotope-labelled internal standard for allergen quantification. Food Chem. 2020, 332, 127413. [Google Scholar] [CrossRef]
- Gaskin, F.E.; Taylor, S.L. Sandwich Enzyme-Linked Immunosorbent Assay (ELISA) for Detection of Cashew Nut in Foods. J. Food Sci. 2011, 76, T218–T226. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Song, S.; Liu, L.; Kuang, H.; Xu, C. Development of Sandwich ELISA and Immunochromatographic Strip for the Detection of Peanut Allergen Ara h 2. Food Anal. Methods 2015, 8, 2605–2611. [Google Scholar] [CrossRef]
- Fasoli, E.; D’Amato, A.; Kravchuk, A.V.; Citterio, A.; Righetti, P.G. In-depth proteomic analysis of non-alcoholic beverages with peptide ligand libraries. I: Almond milk and orgeat syrup. J. Proteom. 2011, 74, 1080–1090. [Google Scholar] [CrossRef]
- Pilolli, R.; Van Poucke, C.; De Angelis, E.; Nitride, C.; de Loose, M.; Gillard, N.; Huet, A.C.; Tranquet, O.; Larré, C.; Adel-Patient, K.; et al. Discovery based high resolution MS/MS analysis for selection of allergen markers in chocolate and broth powder matrices. Food Chem. 2021, 343, 128533. [Google Scholar] [CrossRef]
- Inman, S.E.; Groves, K.; McCullough, B.; Quaglia, M.; Hopley, C. Development of a LC-MS method for the discrimination between trace level Prunus contaminants of spices. Food Chem. 2018, 245, 289–296. [Google Scholar] [CrossRef]
- Sayers, R.L.; Gethings, L.A.; Lee, V.; Balasundaram, A.; Johnson, P.E.; Marsh, J.A.; Wallace, A.; Brown, H.; Rogers, A.; Langridge, J.I.; et al. Microfluidic Separation Coupled to Mass Spectrometry for Quantification of Peanut Allergens in a Complex Food Matrix. J. Proteome Res. 2018, 17, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Garber, E.A.E.; Parker, C.H.; Handy, S.M.; Cho, C.Y.; Panda, R.; Samadpour, M.; Reynaud, D.H.; Ziobro, G.C. Presence of undeclared food allergens in cumin: The need for multiplex methods. J. Agric. Food Chem. 2016, 64, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Cucu, T.; de Meulenaer, B.; Devreese, B. MALDI-based identification of stable hazelnut protein derived tryptic marker peptides. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2012, 29, 1821–1831. [Google Scholar] [CrossRef] [PubMed]
- Chassaigne, H.; Trégoat, V.; Nørgaard, J.V.; Maleki, S.J.; van Hengel, A.J. Resolution and identification of major peanut allergens using a combination of fluorescence two-dimensional differential gel electrophoresis, Western blotting and Q-TOF mass spectrometry. J. Proteom. 2009, 72, 511–526. [Google Scholar] [CrossRef] [PubMed]
- Pedreschi, R.; Nørgaard, J.; Maquet, A. Current challenges in detecting food allergens by shotgun and targeted proteomic approaches: A case study on traces of peanut allergens in baked cookies. Nutrients 2012, 4, 132–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebling, C.M.; McFarland, M.A.; Callahan, J.H.; Ross, M.M. Global proteomic screening of protein allergens and advanced glycation endproducts in thermally processed peanuts. J. Agric. Food Chem. 2013, 61, 5638–5648. [Google Scholar] [CrossRef] [PubMed]
- Sayers, R.L.; Johnson, P.E.; Marsh, J.T.; Barran, P.; Brown, H.; Mills, E.N.C. The effect of thermal processing on the behaviour of peanut allergen peptide targets used in multiple reaction monitoring mass spectrometry experiments. Analyst 2016, 141, 4130–4141. [Google Scholar] [CrossRef] [Green Version]
- Careri, M.; Elviri, L.; Maffini, M.; Mangia, A.; Mucchino, C.; Terenghi, M. Determination of peanut allergens in cereal-chocolate-based snacks: Metal-tag inductively coupled plasma mass spectrometry immunoassay versus liquid chromatography/electrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2008, 22, 807–811. [Google Scholar] [CrossRef]
- Hebling, C.M.; Ross, M.M.; Callahan, J.H.; McFarland, M.A. Size-selective fractionation and visual mapping of allergen protein chemistry in arachis hypogaea. J. Proteome Res. 2012, 11, 5384–5395. [Google Scholar] [CrossRef]
- Vandekerckhove, M.; Van Droogenbroeck, B.; De Loose, M.; Taverniers, I.; Daeseleire, E.; Gevaert, P.; Lapeere, H.; Van Poucke, C. Development of an LC-MS/MS method for the detection of traces of peanut allergens in chili pepper. Anal. Bioanal. Chem. 2017, 409, 5201–5207. [Google Scholar] [CrossRef]
- Apostolovic, D.; Luykx, D.; Warmenhoven, H.; Verbart, D.; Stanic-Vucinic, D.; De Jong, G.A.H.; Velickovic, T.C.; Koppelman, S.J. Reduction and alkylation of peanut allergen isoforms Ara h 2 and Ara h 6; Characterization of intermediate- and end products. Biochim. Biophys. Acta-Proteins Proteom. 2013, 1834, 2832–2842. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, S.E.; Chu, Y.; Ozias-Akins, P.; Thelen, J.J. Validation of gel-free, label-free quantitative proteomics approaches: Applications for seed allergen profiling. J. Proteom. 2009, 72, 555–566. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luparelli, A.; Losito, I.; De Angelis, E.; Pilolli, R.; Lambertini, F.; Monaci, L. Tree Nuts and Peanuts as a Source of Beneficial Compounds and a Threat for Allergic Consumers: Overview on Methods for Their Detection in Complex Food Products. Foods 2022, 11, 728. https://doi.org/10.3390/foods11050728
Luparelli A, Losito I, De Angelis E, Pilolli R, Lambertini F, Monaci L. Tree Nuts and Peanuts as a Source of Beneficial Compounds and a Threat for Allergic Consumers: Overview on Methods for Their Detection in Complex Food Products. Foods. 2022; 11(5):728. https://doi.org/10.3390/foods11050728
Chicago/Turabian StyleLuparelli, Anna, Ilario Losito, Elisabetta De Angelis, Rosa Pilolli, Francesca Lambertini, and Linda Monaci. 2022. "Tree Nuts and Peanuts as a Source of Beneficial Compounds and a Threat for Allergic Consumers: Overview on Methods for Their Detection in Complex Food Products" Foods 11, no. 5: 728. https://doi.org/10.3390/foods11050728