
Maillard Reaction Induced Changes in Allergenicity of Food


Abstract
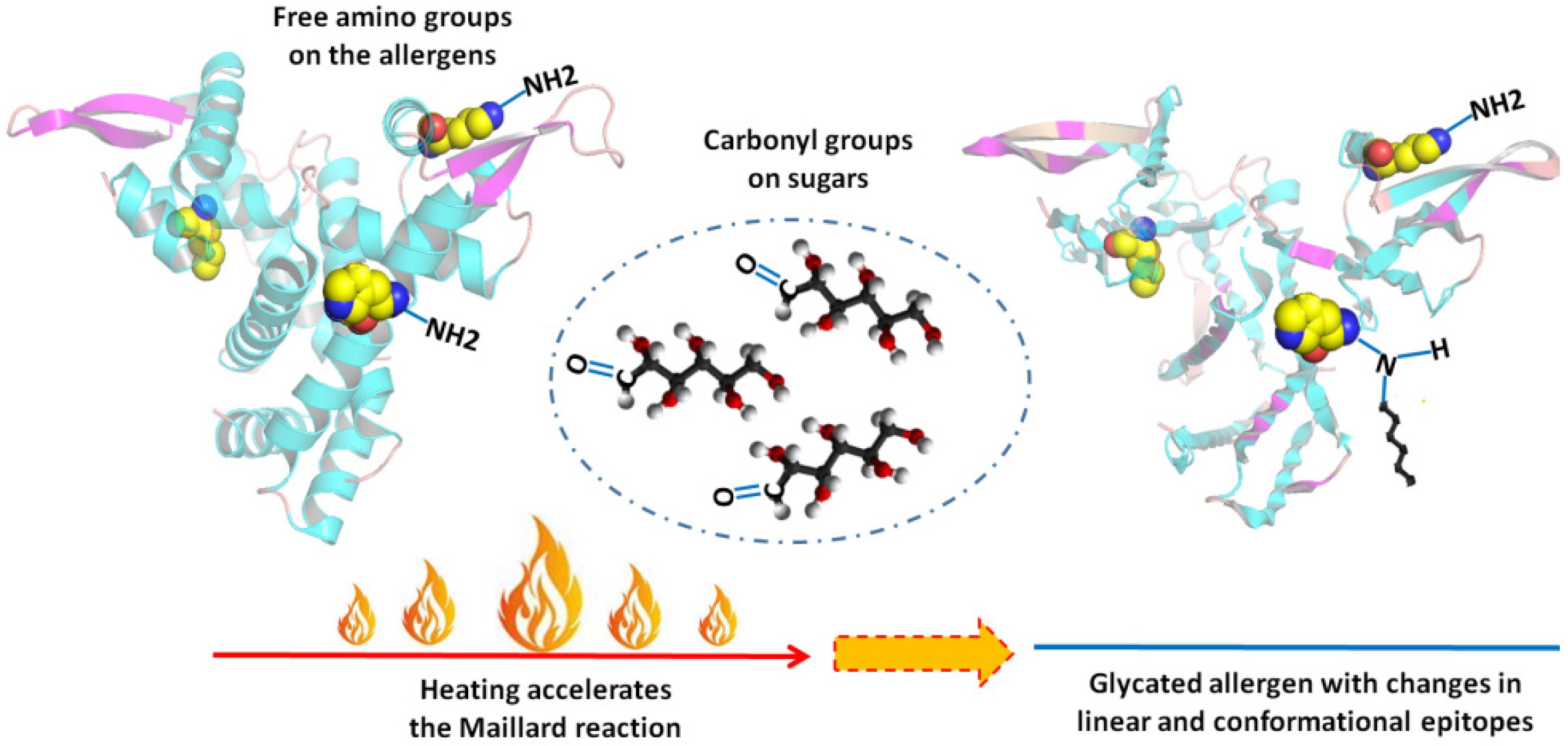
:1. Introduction
2. Definition of Indicators Usedto Determine Food Allergenicity
3. Maillard Reaction and Allergenicity of Foods
3.1. Milk
3.2. Egg
3.3. Peanut
3.4. Shellfish
3.5. Fish
3.6. Tree Nuts
3.7. Wheat
3.8. Sesame
3.9. Soy

{kind=link}
| Food | Protein | Glycation Condition | Allergenicity Change | Ref |
|---|---|---|---|---|
| Milk | β-lactoglobulin | Wet or dry heating with lactose | IgE reactivity similar to heating control, depending on sera used | [31] |
| Dry heating with fructo-, galacto-, and isomalto-oligosacharides | Reduced IgE reactivity | [32] | ||
| Dry heating with galactose. Sonication pretreatment assisted | Sonication assisted glycation to reduce its IgE reactivity | [34] | ||
| Dry heating with mannose. Ultrasound pretreatment assisted. | Ultrasound pretreatment assisted glycation to reduce its IgE reactivity | [35] | ||
| Dry heating with ribose. Ultrasound pretreatment assisted | Ultrasound assisted glycation to reduce its IgE reactivity | [36] | ||
| Dry heating with fructose | Glycation further reduced its IgE reactivity than heating | [30] | ||
| Dry heating with arabinose | Glycation but not heating reduced IgE reactivity. | [29] | ||
| Spray drying with lactose | Glycation further reduced its IgE reactivity than heating | [28] | ||
| α-Lactalbumin | Dry heating with galactose | Glycation further reduced its IgE reactivity than heating | [37] | |
| Dry heating with lactose. High-pressure microfluidiser pretreatment assisted | High-pressure microfluidization assisted glycation to reduce its IgE reactivity | [38] | ||
| Dry heating with galactose. Ultrasonic pretreatment assisted. | Ultrasonic pretreatment assisted. glycation to reduce its IgE reactivity | [39] | ||
| Whey protein isolate | Wet heating with dextran | Reduced IgE reactivity | [40] | |
| Egg | Ovalbumin | Dry heating with glucose, mannose, allose, galactose, and idose | Reduced IgE reactivity compared with heating control | [50] |
| Wet heating with glucose | Increased IgE reactivity | [51] | ||
| Dry heating with ribose. Preheating: 60 °C for 1 h. | Preheating assisted glycation to reduce IgE reactivity | [52] | ||
| Dry heating with mannose. Pretreating with ultrasound. | Ultrasound pretreatment assisted glycation to reduce IgE reactivity | [53] | ||
| Dry heating with mannose, glucose, fructose, and ribose | Glycation with mannose reduced allergenicity in vivo compared with heated control | [54] | ||
| Ovomucoid | Dry heating with glucose | Reduced IgE reactivity | [49] | |
| Egg white | Dry heating with mannose | Reduced allergenicity in vivo | [57] | |
| Peanut | Ara h 1, Ara h 2 or whole peanut extract | Wet heating with fructose, glucose, arabinose, mannose, xylose, galactose, or dextrose | IgE reactivity increased | [65] |
| rAra h 2 | Wet heating with maltose, glucose, fructose, or ribose | IgE reactivity increased | [66] | |
| Ara h 1; 2S albumins containing both Ara h 2 and Ara h 6 (Ara h 2/6) | Dry heating with glucose | Glycation reduced IgE reactivity of Ara h 1, but not Ara h 2/6 | [67] | |
| r-Ara h 1 | Dry heating with glucose | Reduced allergenicity in vivo | [71] | |
| Shellfish | Tropomyosin from Exopalaemon modestus | Dry heating with glucose | Reduced IgE reactivity | [81] |
| Tropomyosin from Exopalaemon modestus | Dry heating with glucose, maltose, maltotriose, maltopentaose, or maltoheptaose | Reductions in IgE reactivity except tropomyosin- maltose | [83] | |
| rTropomyosin | Dry heating with ribose, galacto-oligosaccharide, or chitosan-oligosaccharide | Reduced IgE binding capacity | [82] | |
| Tropomyosin from Scylla paramamosain | Wet heating with ribose, arabinose, galactose, glucose, or maltose | Reaction with galactose, glucose, and arabinose showed reduced IgE reactivity. Tropomyosin-arabinose showed reduced allergenicity in vivo | [85] | |
| Tropomyosin from Penaeus aztecus | Dry heating with glucose, maltose, maltotriose, maltopentaose, or maltoheptaose | Reduced allergenicity in vivo excepted when glycated with maltose | [84] | |
| Tropomyosin from Exopalaemon modestus | Dry heating with glucose | Reduced allergenicity in vivo | [86] | |
| Tropomyosin from Exopalaemon modestus | Dry heating with Fructo-, galacto-, mannan-oligosaccharides, or Maltopentaose | Reduced allergenicity in vivo except reaction with Fructo-oligosaccharide | [73] | |
| Tropomyosin from Exopalaemon modestus | Dry heating with glucose, maltose, or maltotriose | Reduced allergenicity in vivo except reaction with maltose | [74] | |
| Tropomyosin from Patinopecten yessoensis | Dry heating with ribose, glucose, maltose, or maltotriose | No change or increased IgE reactivity | [87] | |
| Arginine Kinase from Scylla paramamosain | Wet heating with ribose, arabinose, galactose, glucose, and maltose | Only arabinose reduced the IgE reactivity and allergenicity in vivo | [85] | |
| rSarcoplasmic-calcium-binding protein | Wet heating with xylose | Reduced IgE reactivity | [89] | |
| Fish | Parvalbumin from Decapterus maruadsi | Wet heating with glucose in the autoclave sterilizer | Pressure assisted glycation to reduce IgE reactivity and allergenicity in vivo | [97] |
| rParvalbumin | Dry heating with glucose | Reduced IgE reactivity | [95] | |
| Parvalbumin from Alaska Pollock | Dry heating with glucose, fructose, ribose, lactose, and galactose | Reduced IgE reactivity when reacted with ribose/galactose | [96] | |
| Parvalbumin from Gadus morhua | Wet heating with glucose | stronger IgE reactivity | [98] | |
| β’-component from Pseudosciaena crocea | Dry heating with glucose | Reduced IgE reactivity | [99] | |
| Tree Nuts | Cor a 11 | Dry heating with glucose | IgE reactivity not reduced more than heating | [106] |
| Soy | Gly m 6 | Dry heating with lactose | Reduced IgE reactivity compared with heating | [127] |
| Acid-precipitated soy protein | Dry heating with galactomannan | Reduced IgE reactivity, especially for Gly m Bd 30 K | [129] | |
| Acid-precipitated soy protein | Dry heating with galactomannan or chitosan | Reduced IgE reactivity, especially for Gly m Bd 30 K | [130] |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Tsuang, A.; Chan, E.S.; Wang, J. Food-induced anaphylaxis in infants: Cannew evidence assist with implementation of food allergy prevention and treatment? J. Allergy Clin. Immunol. Pract. 2021, 9, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Bellinger, S.V.; Stateman, A.J.; Srour, H.H.; Baguley, J.K.; Rivera-Mariani, F.E. Evaluating differences in prevalence of food allergies between two geographic regions: Australia and US. J. Allergy Clin. Immunol. 2019, 143, AB268. [Google Scholar] [CrossRef]
- Clarke, A.E.; Elliott, S.J.; Pierre, Y.S.; Soller, L.; La Vieille, S.; Ben-Shoshan, M. Temporal trends in prevalence of food allergy in Canada. J. Allergy Clin. Immunol. Pract. 2020, 8, 1428–1430.e5. [Google Scholar] [CrossRef] [PubMed]
- Aksoy, A.G.; Boran, P.; Karakoc-Aydiner, E.; Gokcay, G.; Tamay, Z.U.; Devecioglu, E.; Baris, S.; Ozen, A. Prevalence of allergic disorders and risk factors associated with food allergy in Turkish preschoolers. Allergol. Immunopathol. 2021, 49, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, J.; Li, H. Comparison of food allergy prevalence among Chinese infants in Chongqing, 2009versus1999. Pediatr. Int. 2010, 52, 820–824. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.U.; Stahlman, S. Increasing incidence and prevalence of food allergies in the US military, 2000–2017. J. Allergy Clin. Immunol. Pract. 2020, 8, 361–363. [Google Scholar] [CrossRef] [PubMed]
- Warren, C.M.; Turner, P.J.; Chinthrajah, R.S.; Gupta, R.S. Advancing food allergy through epidemiology: Understanding andaddressing disparities in food allergy management and outcomes. J. Allergy Clin. Immunol. Pract. 2021, 9, 110–118. [Google Scholar] [CrossRef]
- Toda, M.; Hellwig, M.; Henle, T.; Vieths, S. Influence of the Maillardreaction on the allergenicity of food proteins and the development of allergic inflammation. Curr. Allergy Asthma Rep. 2019, 19, 4. [Google Scholar] [CrossRef]
- Quan, W.; Li, Y.; Jiao, Y.; Xue, C.; Liu, G.; Wang, Z.; He, Z.; Qin, F.; Zeng, M.; Chen, J. Simultaneous generation of acrylamide, β-carboline heterocyclic amines and advanced glycation ends products in an aqueous Maillard reaction model system. Food Chem. 2020, 332, 127387. [Google Scholar] [CrossRef]
- Costa, J.; Villa, C.; Verhoeckx, K.; Cirkovic-Velickovic, T.; Schrama, D.; Roncada, P.; Rodrigues, P.M.; Piras, C.; Martín-Pedraza, L.; Monaci, L.; et al. Are physicochemical properties shaping the allergenic potency of animal allergens? Clin. Rev. Allergy Immunol. 2022, 62, 1–36. [Google Scholar] [CrossRef]
- Costa, J.; Bavaro, S.L.; Benedé, S.; Diaz-Perales, A.; Bueno-Diaz, C.; Gelencser, E.; Klueber, J.; Larré, C.; Lozano-Ojalvo, D.; Lupi, R.; et al. Are physicochemical properties shaping the allergenic potency of plant allergens? Clin. Rev. Allergy Immunol. 2020. Advance online publication. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Gupta, K.; Sharma, A.; Das, M.; Ansari, I.A.; Dwivedi, P.D. Maillard reaction in food allergy: Pros and Cons. Crit. Rev. Food Sci. Nutr. 2018, 58, 208–226. [Google Scholar] [CrossRef] [PubMed]
- Malgorzata, T.; Joost, V.N.; Huub, S. Food processing: The influence of the Maillard Reaction on immunogenicity and allergenicity of food proteins. Nutrients 2017, 9, 835. [Google Scholar] [CrossRef]
- Anvari, S.; Miller, J.; Yeh, C.Y.; Davis, C.M. IgE-mediated food allergy. Clin. Rev. Allergy Immunol. 2019, 57, 244–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, N.; Adelman, D.C.; Anagnostou, K.; Baumert, J.L.; Blom, W.M.; Campbell, D.E.; Chinthrajah, R.S.; Mills, E.; Javed, B.; Purington, N.; et al. Using data from food challenges to inform management of consumers with food allergy: Asystematic review with individual participant data meta-analysis. J. Allergy Clin. Immunol. 2021, 147, 2249–2262.e7. [Google Scholar] [CrossRef]
- Ahmed, I.; Lin, H.; Li, Z.; Xu, L.; Qazi, I.M.; Luo, C.; Gao, X.; Khan, M.U.; Iqbal, A.; Guo, Y.; et al. Tyrosinase/caffeic acid cross-linking alleviated shrimp (Metapenaeusensis) tropomyosin-induced allergic responses by modulating the Th1/Th2 immunobalance. Food Chem. 2021, 340, 127948. [Google Scholar] [CrossRef]
- El Mecherfi, K.E.; Todorov, S.D.; Cavalcanti de Albuquerque, M.A.; Denery-Papini, S.; Lupi, R.; Haertlé, T.; Dora Gombossy de Melo Franco, B.; Larré, C. Allergenicity of fermented foods: Emphasis on seeds protein-based products. Foods 2020, 9, 792. [Google Scholar] [CrossRef]
- Farjami, T.; Babaei, J.; Nau, F.; Dupont, D.; Madadlou, A. Effects of thermal, non-thermal and emulsification processes on the gastrointestinal digestibility of egg white proteins. Trends Food Sci. Technol. 2021, 107, 45–46. [Google Scholar] [CrossRef]
- Seth, D.; Poowutikul, P.; Pansare, M.; Kamat, D. Food allergy: A review. Pediatr. Ann. 2020, 49, e50–e58. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/food-safety/en/ (accessed on 24 May 2021).
- Ebisawa, M.; Ito, K.; Fujisawa, T.; Committee for Japanese Pediatric Guideline for Food Allergy; The Japanese Society of Pediatric Allergy and Clinical Immunology; Japanese Society of Allergology. Japanese guidelines for food allergy 2020. Allergol. Int. 2020, 69, 370–386. [Google Scholar] [CrossRef]
- Arakali, S.R.; Green, T.D.; Dinakar, C. Prevalence of food allergies in South Asia. Ann. Allergy Asthma Immunol. 2017, 118, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Pham, M.N.; Wang, J. Mammalian milk allergy: Case presentation and review of prevalence, diagnosis, and treatment. Ann. Allergy Asthma Immunol. 2017, 118, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Tan, M.; Wu, J.; Chen, Z.; Long, X.; Zeng, Y.; Cai, H.; Zhang, Y.; Geng, L.; Xiao, Y.; et al. Prevalence, characteristics, and outcome of cow’s milk protein allergy in Chinese infants: A Population-Based Survey. J. Allergy Clin. Immunol. 2019, 43, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Munblit, D.; Perkin, M.R.; Palmer, D.J.; Allen, K.J.; Boyle, R.J. Assessment of evidence about common infant symptoms and cow’s milk allergy. JAMA Pediatr. 2020, 174, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Martorell-Aragonés, A.; Echeverría-Zudaire, L.; Alonso-Lebrero, E.; Boné-Calvo, J.; Martín-Muñoz, M.F.; Nevot-Falcó, S.; Piquer-Gibert, M.; Valdesoiro-Navarrete, L. Position document: IgE-mediated cow’s milk allergy. Allergol. Immunopathol. 2015, 43, 507–526. [Google Scholar] [CrossRef]
- Gasparini, A.; Buhler, S.; Faccini, A.; Sforza, S.; Tedeschi, T. Thermally-induced lactosylation of whey proteins: Identification and synthesis of lactosylated β-lactoglobulinepitope. Molecules 2020, 25, 1294. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Tu, Z.C.; Wang, H.; Huang, T. The reduction in the immunoglobulin G and immunoglobulin E binding capacity of β-lactoglobulin via spray-drying technology. J. Dairy Sci. 2020, 103, 2993–3001. [Google Scholar] [CrossRef]
- Wang, X.M.; Ye, Y.H.; Tu, Z.C.; Hu, Y.M.; Wang, H.; Huang, T. Mechanism of the reduced IgG/IgE binding abilities of glycated β-Lactoglobulin and its digests through high-resolution Mass Spectrometry. J. Agric. Food Chem. 2021, 69, 3741–3750. [Google Scholar] [CrossRef]
- Wang, X.M.; Ye, Y.H.; Tu, Z.C.; Hu, Y.M.; Wang, H.; He, C.Y. Investigation of the mechanism underlying the influence of mild glycation on the digestibility and IgG/IgE-binding abilities of β-lactoglobulin and its digests through LC orbitrap MS/MS. LWT 2021, 139, 110506. [Google Scholar] [CrossRef]
- Zenker, H.E.; Ewaz, A.; Deng, Y.; Savelkoul, H.; van Neerven, R.; De Jong, N.W.; Wichers, H.J.; Hettinga, K.A.; Teodorowicz, M. Differential effects of dry vs. wet heating of β-Lactoglobulin on formation of sRAGEbinding ligands and sIgEepitope recognition. Nutrients 2019, 11, 1432. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Liu, M.; Xia, L.; Wu, H.; Liu, Z.; Xu, X. Conjugation of functional oligosaccharides reduced in vitro allergenicity of β-lactoglobulin. Food Agric. Immunol. 2013, 24, 379–391. [Google Scholar] [CrossRef]
- Perusko, M.; van Roest, M.; Stanic-Vucinic, D.; Simons, P.J.; Pieters, R.; CirkovicVelickovic, T.; Smit, J.J. Glycation of the major milk allergen β-Lactoglobulinchanges its allergenicity by alterations in cellular uptake and degradation. Mol. Nutr. Food Res. 2018, 62, e1800341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, Y.H.; Zhang, Y.; Zhu, M.F.; Liu, J.; Tu, Z.C. Glycation of β-lactoglobulin combined by sonication pretreatment reduce its allergenic potential. Int. J. Biol. Macromol. 2020, 164, 1527–1535. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Tu, Z.; Wang, H.; Zhang, L.; Xu, S.; Niu, C.; Yao, H.; Kaltashov, I.A. Mechanism of Reduction in IgG and IgEbinding of β-Lactoglobulininduced by ultrasound pretreatment combined with dry-State glycation: A study using conventional spectrometry and high-resolution Mass Spectrometry. J. Agric. Food Chem. 2017, 65, 8018–8027. [Google Scholar] [CrossRef]
- Liu, G.X.; Tu, Z.C.; Yang, W.; Wang, H.; Zhang, L.; Ma, D.; Huang, T.; Liu, J.; Li, X. Investigation into allergenicity reduction and glycation sites of glycated β-lactoglobulin with ultrasound pretreatment by high-resolution mass spectrometry. Food Chem. 2018, 252, 99–107. [Google Scholar] [CrossRef]
- Liu, J.; Chen, W.M.; Shao, Y.H.; Zhang, J.L.; Tu, Z.C. The mechanism of the reduction in allergenic reactivity of bovine α-lactalbumin induced by glycation, phosphorylation and acetylation. Food Chem. 2020, 310, 125853. [Google Scholar] [CrossRef]
- Li, J.L.; Liu, J.; Ye, Y.H.; Yang, P.; Tu, Z.C. Reduced IgE/IgG binding capacities of bovine α-Lactalbumin by glycation after dynamic high-pressure microfluidization pretreatment evaluated by high resolution mass spectrometry. Food Chem. 2019, 299, 125166. [Google Scholar] [CrossRef]
- Liu, J.; Tu, Z.C.; Liu, G.X.; Niu, C.D.; Yao, H.L.; Wang, H.; Sha, X.M.; Shao, Y.H.; Kaltashov, I.A. Ultrasonic pretreatment combined with dry-state glycationreduced the Immunoglobulin E/Immunoglobulin G-binding ability of α-Lactalbuminrevealed by high-resolution Mass Spectrometry. J. Agric. Food Chem. 2018, 66, 5691–5698. [Google Scholar] [CrossRef]
- Xu, L.; Gong, Y.; Gern, J.E.; Ikeda, S.; Lucey, J.A. Glycation of whey protein with dextrans of different molar mass: Effect on immunoglobulin E-binding capacity with blood sera obtained from patients with cow milk protein allergy. J. Dairy Sci. 2018, 101, 6823–6834. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Gong, Y.; Gern, J.E.; Lucey, J.A. Influence of whey protein hydrolysis in combination with dextran glycation on immunoglobulin E binding capacity with blood sera obtained from patients with a cow milk protein allergy. J. Dairy Sci. 2020, 103, 1141–1150. [Google Scholar] [CrossRef]
- Zhao, D.; Sheng, B.; Li, H.; Wu, Y.; Xu, D.; Li, C. Glycation from α-dicarbonyl compounds has different effects on the heat-induced aggregation of bovine serum albumin and β-casein. Food Chem. 2021, 340, 128108. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, R.; Ferdowsi, L.; Tavaf, Z.; Sadeghian, T.; Tamaddon, A.M.; Moghtaderi, M.; Pourpak, Z. Evaluation of structure, chaperone-like activity and allergenicity of reduced glycatedadduct of bovine β-casein. Protein Pept. Lett. 2017, 24, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Peters, R.L.; Koplin, J.J.; Gurrin, L.C.; Dharmage, S.C.; Wake, M.; Ponsonby, A.L.; Tang, M.L.K.; Lowe, A.J.; Matheson, M.; Dwyer, T.; et al. The prevalence of food allergy and other allergic diseases in early childhood in a population-based study: HealthNuts age 4-year follow-up. J. Allergy Clin. Immunol. 2017, 140, 145–153.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samady, W.; Warren, C.; Wang, J.; Das, R.; Gupta, R.S. Egg allergy in US children. J. Allergy Clin. Immunol. Pract. 2020, 8, 3066–3073.e6. [Google Scholar] [CrossRef]
- Capucilli, P.; Kennedy, K.; Lee, J.; Grundmeier, R.W.; Spergel, J.M. Accidental versus new food allergy reactions in a pediatric emergency department. J. Allergy Clin. Immunol. Pract. 2019, 7, 1662–1664. [Google Scholar] [CrossRef]
- Claude, M.; Lupi, R.; Bouchaud, G.; Bodinier, M.; Brossard, C.; Denery-Papini, S. The thermal aggregation of ovalbumin as large particles decreases its allergenicity for egg allergic patients and in a murine model. Food Chem. 2016, 203, 136–144. [Google Scholar] [CrossRef]
- Bloom, K.A.; Huang, F.R.; Bencharitiwong, R.; Bardina, L.; Ross, A.; Sampson, H.A.; Nowak-Węgrzyn, A. Effect of heat treatment on milk and egg proteins allergenicity. Pediatr. Allergy Immunol. 2014, 25, 740–746. [Google Scholar] [CrossRef]
- Jiménez-Saiz, R.; Belloque, J.; Molina, E.; López-Fandiño, R. Human immunoglobulin E (IgE) binding to heated and glycated ovalbumin and ovomucoid before and after in vitro digestion. J. Agric. Food Chem. 2011, 59, 10044–10051. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, G.; Tu, Z.; Wang, H.; Hu, Y.; Mao, J.; Zhang, J. Insight into the mechanism of reduced IgG/IgE binding capacity in Ovalbumin as induced by glycation with monoseepimers through Liquid Chromatography and High-Resolution Mass Spectrometry. J. Agric. Food Chem. 2020, 68, 6065–6075. [Google Scholar] [CrossRef]
- Ma, X.J.; Gao, J.Y.; Chen, H.B. Combined effect of glycation and sodium carbonate-bicarbonate buffer concentration on IgG binding, IgE binding and conformation of ovalbumin. J. Sci. Food Agric. 2013, 93, 3209–3215. [Google Scholar] [CrossRef]
- Liao, Z.W.; Ye, Y.H.; Wang, H.; Chen, Y.; Sha, X.M.; Zhang, L.; Huang, T.; Hu, Y.M.; Tu, Z.C. The mechanism of decreased IgG/IgE-binding of Ovalbumin by preheating treatment combined with glycationidentified by Liquid Chromatography and High-Resolution Mass Spectrometry. J. Agric. Food Chem. 2018, 66, 10693–10702. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Tu, Z.; Wang, H.; Zhang, L.; Song, Q. Glycation of ovalbumin after high-intensity ultrasound pretreatment: Effects on conformation, immunoglobulin (Ig)G/IgE binding ability and antioxidant activity. J. Sci. Food Agric. 2018, 98, 3767–3773. [Google Scholar] [CrossRef] [PubMed]
- Rupa, P.; Mine, Y. Comparison of glycatedovalbumin-monosaccharides in the attenuation of ovalbumin-induced allergic response in a BALB/C mouse model. J. Agric. Food Chem. 2019, 67, 8138–8148. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Pandey, N.K.; Singha Roy, A.; Tripathy, D.R.; Dinda, A.K.; Dasgupta, S. Prolonged glycation of hen egg white lysozyme generates non amyloidal structures. PLoS ONE 2013, 8, e74336. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Huang, Q. Modification of ovotransferrin by maillard reaction: Consequences for structure, fibrillation and emulsifying property of fibrils. Food Hydrocoll. 2019, 97, 105186. [Google Scholar] [CrossRef]
- Rupa, P.; Nakamura, S.; Katayama, S.; Mine, Y. Attenuation of allergic immune response phenotype by mannosylated egg white in orally induced allergy in BALB/c mice. J. Agric. Food Chem. 2014, 62, 9479–9487. [Google Scholar] [CrossRef]
- Mahr, T.A.; Lieberman, J.A.; Haselkorn, T.; Damle, V.; Ali, Y.; Chidambaram, A.; Griffin, N.M.; Sublett, J.W. Characteristics of Peanut Allergy Diagnosis in a US Health Care Claims Database (2011–2017). J. Allergy Clin. Immunol. Pract. 2021, 9, 1683–1694.e5. [Google Scholar] [CrossRef]
- Greenhawt, M.; Shaker, M.; Wang, J.; Oppenheimer, J.J.; Sicherer, S.; Keet, C.; Swaggart, K.; Rank, M.; Portnoy, J.M.; Bernstein, J.; et al. Peanut allergy diagnosis: A 2020 practice parameter update, systematic review, and GRADE analysis. J. Allergy Clin. Immunol. 2020, 146, 1302–1334. [Google Scholar] [CrossRef]
- Frischmeyer-Guerrerio, P.A.; Rasooly, M.; Gu, W.; Levin, S.; Jhamnani, R.D.; Milner, J.D.; Stone, K.; Guerrerio, A.L.; Jones, J.; Borres, M.P.; et al. IgE testing can predict food allergy status in patients with moderate to severe atopic dermatitis. Ann. Allergy Asthma Immunol. 2019, 122, 393–400.e2. [Google Scholar] [CrossRef]
- Guillon, B.; Bernard, H.; Drumare, M.F.; Hazebrouck, S.; Adel-Patient, K. Heat processing of peanut seed enhances the sensitization potential of the major peanut allergen Arah 6. Mol. Nutr. Food Res. 2016, 60, 2722–2735. [Google Scholar] [CrossRef] [Green Version]
- Comstock, S.S.; Maleki, S.J.; Teuber, S.S. Boiling and frying peanuts decreases soluble peanut (ArachisHypogaea) allergens Ara h 1 and Ara h 2 but does not generate hypoallergenic peanuts. PLoS ONE 2016, 11, e0157849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Shi, Y.; Zhao, Y.; Tang, G.; Niu, B.; Chen, Q. Boiling and roasting treatment affecting the peanut allergenicity. Ann. Transl. Med. 2018, 6, 357. [Google Scholar] [CrossRef] [PubMed]
- Cabanillas, B.; Novak, N. Effects of daily food processing on allergenicity. Crit. Rev. Food Sci. Nutr. 2019, 59, 31–42. [Google Scholar] [CrossRef]
- Maleki, S.J.; Chung, S.Y.; Champagne, E.T.; Raufman, J.P. The effects of roasting on the allergenic properties of peanut proteins. J. Allergy Clin. Immunol. 2000, 106, 763–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruber, P.; Becker, W.M.; Hofmann, T. Influence of the maillard reaction on the allergenicity of rAra h 2, a recombinant major allergen from peanut (Arachishypogaea), its major epitopes, and peanut agglutinin. J. Agric. Food Chem. 2005, 53, 2289–2296. [Google Scholar] [CrossRef] [PubMed]
- Vissers, Y.M.; Iwan, M.; Adel-Patient, K.; Stahl Skov, P.; Rigby, N.M.; Johnson, P.E.; Mandrup Müller, P.; Przybylski-Nicaise, L.; Schaap, M.; Ruinemans-Koerts, J.; et al. Effect of roasting on the allergenicity of major peanut allergens Ara h 1 and Ara h 2/6: The necessity of degranulation assays. Clin. Exp. Allergy 2011, 41, 1631–1642. [Google Scholar] [CrossRef]
- Blanc, F.; Vissers, Y.M.; Adel-Patient, K.; Rigby, N.M.; Mackie, A.R.; Gunning, A.P.; Wellner, N.K.; Skov, P.S.; Przybylski-Nicaise, L.; Ballmer-Weber, B.; et al. Boiling peanut Ara h 1 results in the formation of aggregates with reduced allergenicity. Mol. Nutr. Food Res. 2011, 55, 1887–1894. [Google Scholar] [CrossRef]
- Vissers, Y.M.; Blanc, F.; Skov, P.S.; Johnson, P.E.; Rigby, N.M.; Przybylski-Nicaise, L.; Bernard, H.; Wal, J.M.; Ballmer-Weber, B.; Zuidmeer-Jongejan, L.; et al. Effect of heating and glycation on the allergenicity of 2S albumins (Ara h 2/6) from peanut. PLoS ONE 2011, 6, e23998. [Google Scholar] [CrossRef]
- Mueller, G.A.; Maleki, S.J.; Johnson, K.; Hurlburt, B.K.; Cheng, H.; Ruan, S.; Nesbit, J.B.; Pomés, A.; Edwards, L.L.; Schorzman, A.; et al. Identification of Maillard reaction products on peanut allergens that influence binding to the receptor for advanced glycation end products. Allergy 2013, 68, 1546–1554. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Wang, M.; Ding, Y.; Chen, J.; Niu, B.; Chen, Q. Effects of Maillard reaction on structural modification and potential allergenicity of peanut 7S globulin (Ara h 1). J. Sci. Food Agric. 2020, 100, 5617–5626. [Google Scholar] [CrossRef]
- Chung, S.Y.; Champagne, E.T. Association of end-product adducts with increased IgE binding of roasted peanuts. J. Agric. Food Chem. 2001, 49, 3911–3916. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, X.M.; Xiao, H.; Nowak-Wegrzyn, A.; Zhou, P. Insight into the allergenicity of shrimp tropomyosin glycated by functional oligosaccharides containing advanced glycation end products. Food Chem. 2020, 302, 125348. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xiao, H.; Zhou, P. Glycation by saccharides of different molecular sizes affected the allergenicity of shrimp tropomyosin via epitope loss and the generation of advanced glycation end products. Food Funct. 2019, 10, 7042–7051. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Rao, H.; Zhang, K.; Tao, S.; Xue, W.T. Effects of different thermal processing methods on the structure and allergenicity of peanut allergen Ara h 1. Food Sci. Nutr. 2018, 6, 1706–1714. [Google Scholar] [CrossRef]
- Zhang, W.; Zhu, Q.; Zhang, T.; Cai, Q.; Chen, Q. Thermal processing effects on peanut allergen Ara h 2 allergenicity in mice and its antigenic epitope structure. Food Chem. 2016, 212, 657–662. [Google Scholar] [CrossRef]
- Khan, M.U.; Ahmed, I.; Lin, H.; Li, Z.; Costa, J.; Mafra, I.; Chen, Y.; Wu, Y.N. Potential efficacy of processing technologies for mitigating crustacean allergenicity. Crit. Rev. Food Sci. Nutr. 2019, 59, 2807–2830. [Google Scholar] [CrossRef]
- Ziyab, A.H. Prevalence of food allergy among schoolchildren in Kuwait and its association with the coexistence and severity of asthma, rhinitis, and eczema: A cross-sectional study. World Allergy Organ. J. 2019, 12, 100024. [Google Scholar] [CrossRef] [Green Version]
- Tham, E.H.; Shek, L.P.; Van Bever, H.P.; Vichyanond, P.; Ebisawa, M.; Wong, G.W.; Lee, B.W.; Asia Pacific Association of Pediatric Allergy, Respirology & Immunology (APAPARI). Early introduction of allergenic foods for the prevention of food allergy from an Asian perspective-An Asia Pacific Association of Pediatric Allergy, Respirology & Immunology (APAPARI) consensus statement. Pediatr. Allergy Immunol. 2018, 29, 18–27. [Google Scholar] [CrossRef]
- Ruethers, T.; Taki, A.C.; Johnston, E.B.; Nugraha, R.; Le, T.; Kalic, T.; McLean, T.R.; Kamath, S.D.; Lopata, A.L. Seafood allergy: A comprehensive review of fish and shellfish allergens. Mol. Immunol. 2018, 100, 28–57. [Google Scholar] [CrossRef]
- Zhang, Z.; Xiao, H.; Zhang, X.; Zhou, P. Conformation, allergenicity and human cell allergy sensitization of tropomyosin from Exopalaemonmodestus: Effects of deglycosylation and Maillard reaction. Food Chem. 2019, 276, 520–527. [Google Scholar] [CrossRef]
- Fu, L.; Wang, C.; Wang, J.; Ni, S.; Wang, Y. Maillard reaction with ribose, galacto-oligosaccharide or chitosan-oligosaccharide reduced the allergenicity of shrimp tropomyosin by inducing conformational changes. Food Chem. 2019, 274, 789–795. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xiao, H.; Zhou, P. Allergenicity suppression of tropomyosin from Exopalaemonmodestus by glycation with saccharides of different molecular sizes. Food Chem. 2019, 288, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, X.M.; Li, Z.; Lin, H. Investigation of glycated shrimp tropomyosin as a hypoallergen for potential immunotherapy. Food Funct. 2021, 12, 2750–2759. [Google Scholar] [CrossRef] [PubMed]
- Han, X.Y.; Yang, H.; Rao, S.T.; Liu, G.Y.; Hu, M.J.; Zeng, B.C.; Cao, M.J.; Liu, G.M. The Maillard reaction reduced the sensitization of Tropomyosin and Arginine Kinase from Scylla paramamosain, simultaneously. J. Agric. Food Chem. 2018, 66, 2934–2943. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xiao, H.; Zhang, X.; Zhou, P. Insight into the effects of deglycosylation and glycation of shrimp tropomyosin on in vivo allergenicity and mast cell function. Food Funct. 2019, 10, 3934–3941. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Watanabe, K.; Ojima, T.; Ahn, D.H.; Saeki, H. Effect of maillard reaction on allergenicity of scallop tropomyosin. J. Agric. Food Chem. 2005, 53, 7559–7564. [Google Scholar] [CrossRef]
- Yuan, F.; Lv, L.; Li, Z.; Mi, N.; Chen, H.; Lin, H. Effect of transglutaminase-catalyzed glycosylation on the allergenicity and conformational structure of shrimp (Metapenaeusensis) tropomyosin. Food Chem. 2017, 219, 215–222. [Google Scholar] [CrossRef]
- Hu, M.J.; Liu, G.Y.; Yang, Y.; Pan, T.M.; Liu, Y.X.; Sun, L.C.; Cao, M.J.; Liu, G.M. Cloning, expression, and the effects of processing on Sarcoplasmic-Calcium-Binding protein: An important allergen in Mud Crab. J. Agric. Food Chem. 2017, 65, 6247–6257. [Google Scholar] [CrossRef]
- Liu, J.H.; Xu, Q.H.; Zhang, J.J. Characterization of silver carp (Hypophthalmichthysmolitrix) myosin protein glycated with konjacoligo-glucomannan. Food Hydrocoll. 2016, 57, 114–121. [Google Scholar] [CrossRef]
- Kourani, E.; Corazza, F.; Michel, O.; Doyen, V. What do we know about fish allergy at the end of the decade? J. Investig. Allergol. Clin. Immunol. 2019, 29, 414–421. [Google Scholar] [CrossRef]
- Mourad, A.A.; Bahna, S.L. Fish-allergic patients may be able to eat fish. Expert Rev. Clin. Immunol. 2015, 11, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Jeebhay, M.F.; Cartier, A. Seafood workers and respiratory disease: An update. Curr. Opin. Allergy Clin. Immunol. 2010, 10, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Hilger, C.; van Hage, M.; Kuehn, A. Diagnosis of Allergy to Mammals and Fish: Cross-Reactive vs. Specific Markers. Curr. Allergy Asthma Rep. 2017, 17, 64. [Google Scholar] [CrossRef]
- Zhao, Y.J.; Cai, Q.F.; Jin, T.C.; Zhang, L.J.; Fei, D.X.; Liu, G.M.; Cao, M.J. Effect of Maillard reaction on the structural and immunological properties of recombinant silver carp parvalbumin. LWT Food Sci. Technol. 2017, 75, 25–33. [Google Scholar] [CrossRef]
- Zhang, M.; Tu, Z.C.; Liu, J.; Hu, Y.M.; Wang, H.; Mao, J.H.; Li, J.L. The IgE/IgG binding capacity and structural changes of Alaska Pollock parvalbuminglycated with different reducing sugars. J. Food Biochem. 2021, 45, e13539. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Min, J.; Han, X.Y.; Li, X.Y.; Hu, J.W.; Liu, H.; Cao, M.J.; Liu, G.M. Reduction of the histamine content and immunoreactivity of parvalbumin in Decapterusmaruadsi by a Maillard reaction combined with pressure treatment. Food Funct. 2018, 9, 4897–4905. [Google Scholar] [CrossRef]
- De Jongh, H.H.; Robles, C.L.; Timmerman, E.; Nordlee, J.A.; Lee, P.W.; Baumert, J.L.; Hamilton, R.G.; Taylor, S.L.; Koppelman, S.J. Digestibility and IgE-binding of glycosylated codfish parvalbumin. Biomed Res. Int. 2013, 2013, 756789. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.Y.; Cao, M.J.; Zhang, M.L.; Hu, J.W.; Zhang, Y.X.; Zhang, L.J.; Liu, G.M. Purification, characterization and immunoreactivityof β’-component, a major allergen from the roe of large yellow croaker (Pseudosciaenacrocea). Food Chem. Toxicol. 2014, 72, 111–121. [Google Scholar] [CrossRef]
- Hong, P.K.; Gottardi, D.; Ndagijimana, M.; Betti, M. Glycation and transglutaminase mediated glycosylation of fish gelatin peptides with glucosamine enhance bioactivity. Food Chem. 2014, 142, 285–293. [Google Scholar] [CrossRef]
- Vanga, S.K.; Raghavan, V. Processing effects on tree nut allergens: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 3794–3806. [Google Scholar] [CrossRef]
- McWilliam, V.; Peters, R.; Tang, M.; Dharmage, S.; Ponsonby, A.L.; Gurrin, L.; Perrett, K.; Koplin, J.; Allen, K.J.; HealthNuts Investigators. Patterns of tree nut sensitization and allergy in the first 6 years of life in a population-based cohort. J. Allergy Clin. Immunol. 2019, 143, 644–650.e5. [Google Scholar] [CrossRef] [PubMed]
- Al-Tamemi, S.; Naseem, S.U.; Tufail-Alrahman, M.; Al-Kindi, M.; Alshekaili, J. Food allergen sensitisation patterns in Omani patients with allergic manifestations. Sultan Qaboos Univ. Med. J. 2018, 18, e483–e488. [Google Scholar] [CrossRef] [PubMed]
- Pouessel, G.; Turner, P.J.; Worm, M.; Cardona, V.; Deschildre, A.; Beaudouin, E.; Renaudin, J.M.; Demoly, P.; Tanno, L.K. Food-induced fatal anaphylaxis: From epidemiological data to general prevention strategies. Clin. Exp. Allergy 2018, 48, 1584–1593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smeekens, J.M.; Bagley, K.; Kulis, M. Tree nut allergies: Allergen homology, cross-reactivity, and implications for therapy. Clin. Exp. Allergy 2018, 48, 762–772. [Google Scholar] [CrossRef] [PubMed]
- Iwan, M.; Vissers, Y.M.; Fiedorowicz, E.; Kostyra, H.; Kostyra, E.; Savelkoul, H.F.; Wichers, H.J. Impact of Maillard reaction on immunoreactivity and allergenicity of the hazelnut allergen Cor a 11. J. Agric. Food Chem. 2011, 59, 7163–7171. [Google Scholar] [CrossRef] [PubMed]
- Cucu, T.; De Meulenaer, B.; Bridts, C.; Devreese, B.; Ebo, D. Impact of thermal processing and the Maillard reaction on the basophil activation of hazelnut allergic patients. Food Chem. Toxicol. 2012, 50, 1722–1728. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, G.S.; Liu, C.; Su, M.; Venkatachalam, M.; Roux, K.H.; Sathe, S.K. Effects of the Maillard reaction on the immunoreactivity of Amandin in food matrices. J. Food Sci. 2017, 82, 2495–2503. [Google Scholar] [CrossRef] [PubMed]
- Willison, L.N.; Zhang, Q.; Su, M.; Teuber, S.S.; Sathe, S.K.; Roux, K.H. Conformational epitope mapping of Pru du 6, a major allergen from almond nut. Mol. Immunol. 2013, 55, 253–263. [Google Scholar] [CrossRef]
- Kennard, L.; Thomas, I.; Rutkowski, K.; Azzu, V.; Yong, P.F.K.; Kasternow, B.; Hunter, H.; Cabdi, N.M.O.; Nakonechna, A.; Wagner, A. A multicenter evaluation of diagnosis and management of Omega-5 Gliadin allergy (also known as wheat-dependent exercise-induced anaphylaxis) in 132 adults. J. Allergy Clin. Immunol. Pract. 2018, 6, 1892–1897. [Google Scholar] [CrossRef]
- Cianferoni, A. Wheat allergy: Diagnosis and management. J. Asthma Allergy 2016, 9, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Ashtari, S.; Pourhoseingholi, M.A.; Rostami, K.; Aghdaei, H.A.; Rostami-Nejad, M.; Busani, L.; Tavirani, M.R.; Zali, M.R. Prevalence of gluten-related disorders in Asia-Pacific region: A systematic review. J. Gastrointestin Liver Dis. 2019, 28, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Nwaru, B.I.; Hickstein, L.; Panesar, S.S.; Roberts, G.; Muraro, A.; Sheikh, A.; EAACI Food Allergy and Anaphylaxis Guidelines Group. Prevalence of common food allergies in Europe: A systematic review and meta-analysis. Allergy 2014, 69, 992–1007. [Google Scholar] [CrossRef]
- Nowak-Wegrzyn, A.; Fiocchi, A. Rare, medium, or well done? The effect of heating and food matrix on food protein allergenicity. Curr. Opin. Allergy Clin. Immunol. 2009, 9, 234–237. [Google Scholar] [CrossRef] [PubMed]
- Warren, C.M.; Chadha, A.S.; Sicherer, S.H.; Jiang, J.; Gupta, R.S. Prevalence and severity of sesame allergy in the United States. JAMA Netw. Open 2019, 2, e199144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, A.; Bahna, S.L. Hypersensitivities to sesame and other common edible seeds. Allergy 2016, 71, 1405–1413. [Google Scholar] [CrossRef]
- Sokol, K.; Rasooly, M.; Dempsey, C.; Lassiter, S.; Gu, W.; Lumbard, K.; Frischmeyer-Guerrerio, P.A. Prevalence and diagnosis of sesame allergy in children with IgE-mediated food allergy. Pediatr. Allergy Immunol. 2020, 31, 214–218. [Google Scholar] [CrossRef]
- Bedolla-Barajas, M.; Bedolla-Pulido, T.R.; Macriz-Romero, N.; Morales-Romero, J.; Robles-Figueroa, M. Prevalence of peanut, tree nut, sesame, and seafood allergy in Mexican adults. Rev. Investig. Clin. 2015, 67, 379–386. [Google Scholar]
- Garkaby, J.; Epov, L.; Musallam, N.; Almog, M.; Bamberger, E.; Mandelberg, A.; Dalal, I.; Kessel, A. The sesame-peanut conundrum in Israel: Reevaluation of food allergy prevalence in young children. J. Allergy Clin. Immunol. Pract. 2021, 9, 200–205. [Google Scholar] [CrossRef]
- Li, P.H.; Gunawardana, N.; Thomas, I.; Ue, K.L.; Siew, L.; Watts, T.J.; Bintcliffe, K.; Haque, R.; Rutkowski, K.; Skypala, I.; et al. Sesame allergy in adults: Investigation and outcomes of oral food challenges. Ann. Allergy Asthma Immunol. 2017, 119, 285–287. [Google Scholar] [CrossRef] [Green Version]
- Berk, E.; Hamzalıoğlu, A.; Gökmen, V. Investigations on the Maillard Reaction in sesame (Sesamumindicum L.) seeds induced by roasting. J. Agric. Food Chem. 2019, 67, 4923–4930. [Google Scholar] [CrossRef]
- Dreskin, S.C.; Koppelman, S.J.; Andorf, S.; Nadeau, K.C.; Kalra, A.; Braun, W.; Negi, S.S.; Chen, X.; Schein, C.H. The importance of the 2S albumins for allergenicity and cross-reactivity of peanuts, tree nuts, and sesame seeds. J. Allergy Clin. Immunol. 2021, 147, 1154–1163. [Google Scholar] [CrossRef] [PubMed]
- Messina, M.; Rogero, M.M.; Fisberg, M.; Waitzberg, D. Health impact of childhood and adolescent soy consumption. Nutr. Rev. 2017, 75, 500–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, D.; Adányi, N.; Takács, K.; Maczó, A.; Nagy, A.; Gelencsér, É.; Pachner, M.; Lauter, K.; Baumgartner, S.; Vollmann, J. Development of soybeans with low P34 allergen protein concentration for reduced allergenicity of soy foods. J. Sci. Food Agric. 2017, 97, 1010–1017. [Google Scholar] [CrossRef] [PubMed]
- Saeed, H.; Gagnon, C.; Cober, E.; Gleddie, S. Using patient serum to epitope map soybean glycinins reveals common epitopes shared with many legumes and tree nuts. Mol. Immunol. 2016, 70, 125–133. [Google Scholar] [CrossRef] [PubMed]
- He, M.X.; Xi, J. Identification of an IgE epitope of soybean allergen Gly m Bd 60K. LWT 2020, 133, 110131. [Google Scholar] [CrossRef]
- Bu, G.; Zhang, N.; Chen, F. The influence of glycosylation on the antigenicity, allergenicity, and structural properties of 11S-lactose conjugates. Food Res. Int. 2015, 76 Pt 3, 511–517. [Google Scholar] [CrossRef]
- Bu, G.; Zhu, T.; Chen, F. The structural properties and antigenicity of soybean glycinin by glycation with xylose. J. Sci. Food Agric. 2017, 97, 2256–2262. [Google Scholar] [CrossRef]
- Babiker, E.E.; Hiroyuki, A.; Matsudomi, N.; Iwata, H.; Ogawa, T.; Bando, N.; Kato, A. Effect of polysaccharide conjugation or transglutaminase treatment on the allergenicity and functional properties of soy protein. J. Agric. Food Chem. 1998, 46, 866–871. [Google Scholar] [CrossRef]
- Usui, M.; Tamura, H.; Nakamura, K.; Ogawa, T.; Muroshita, M.; Azakami, H.; Kanuma, S.; Kato, A. Enhanced bactericidal action and masking of allergen structure of soy protein by attachment of chitosan through maillard-type protein-polysaccharide conjugation. Nahrung 2004, 48, 69–72. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gou, J.; Liang, R.; Huang, H.; Ma, X. Maillard Reaction Induced Changes in Allergenicity of Food. Foods 2022, 11, 530. https://doi.org/10.3390/foods11040530
Gou J, Liang R, Huang H, Ma X. Maillard Reaction Induced Changes in Allergenicity of Food. Foods. 2022; 11(4):530. https://doi.org/10.3390/foods11040530
Chicago/Turabian StyleGou, Jingkun, Rui Liang, Houjin Huang, and Xiaojuan Ma. 2022. "Maillard Reaction Induced Changes in Allergenicity of Food" Foods 11, no. 4: 530. https://doi.org/10.3390/foods11040530