
Genomic and Transcriptomic Analysis Reveal Multiple Strategies for the Cadmium Tolerance in Vibrio parahaemolyticus N10-18 Isolated from Aquatic Animal Ostrea gigas Thunberg
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. V. parahaemolyticus Strain and Cultural Conditions
2.2. Genomic DNA Preparation, Sequencing, Assembly, and Annotation
2.3. Comparative Genome Analysis
2.4. Phylogenetic Tree Analysis
2.5. Determination of Minimum Inhibitory Concentrations (MICs) of Antibiotics and Heavy Metals
2.6. Stress Conditions
2.7. Cell Membrane Permeability, Fluidity, and Surface Hydrophobicity Assays
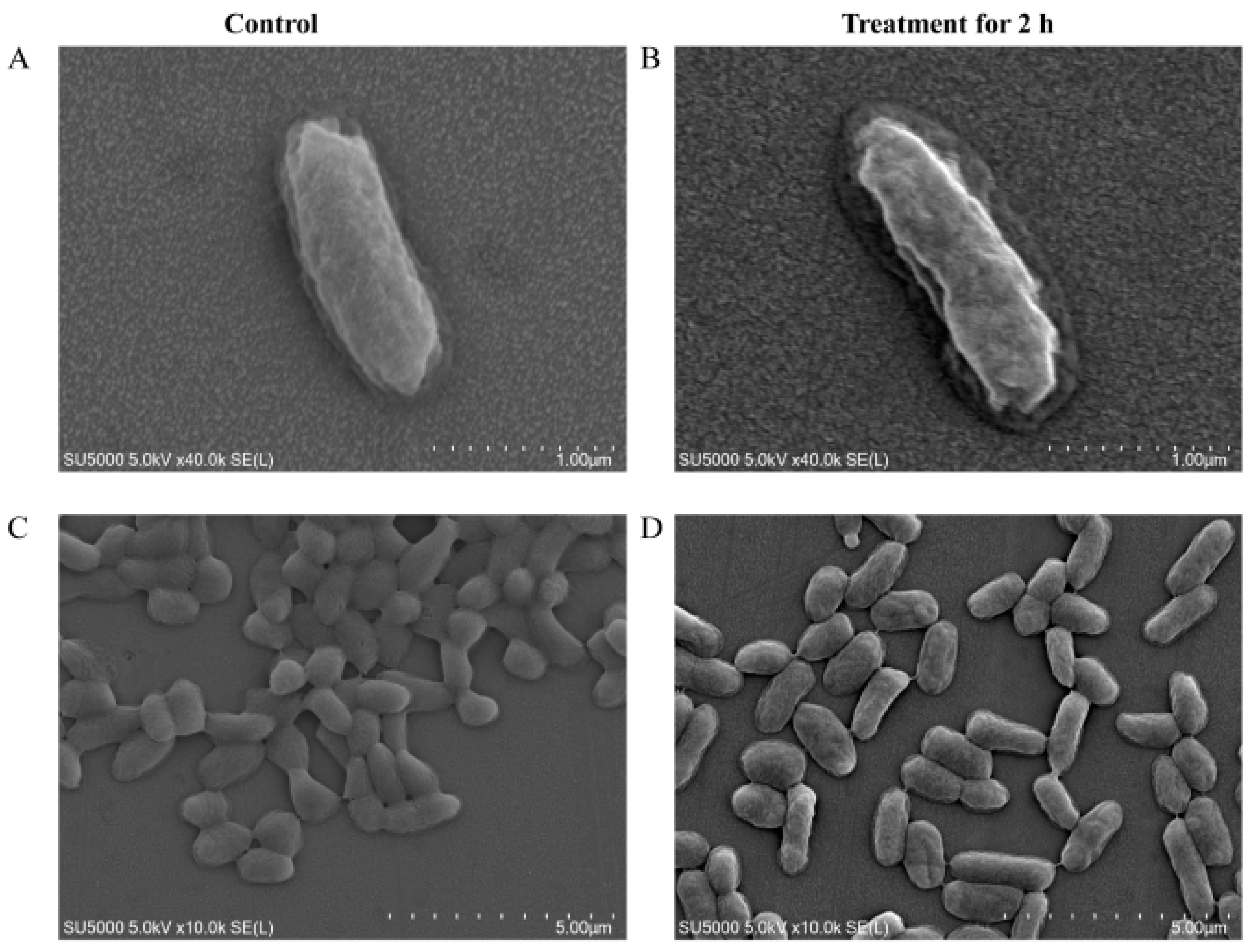
2.8. Scanning Electron Microscope (SEM) Assay
2.9. Illumina RNA Sequencing and Analysis
2.10. Statistical Analysis
3. Results
3.1. Genotype and Phenotype of V. parahaemolyticus N10-18
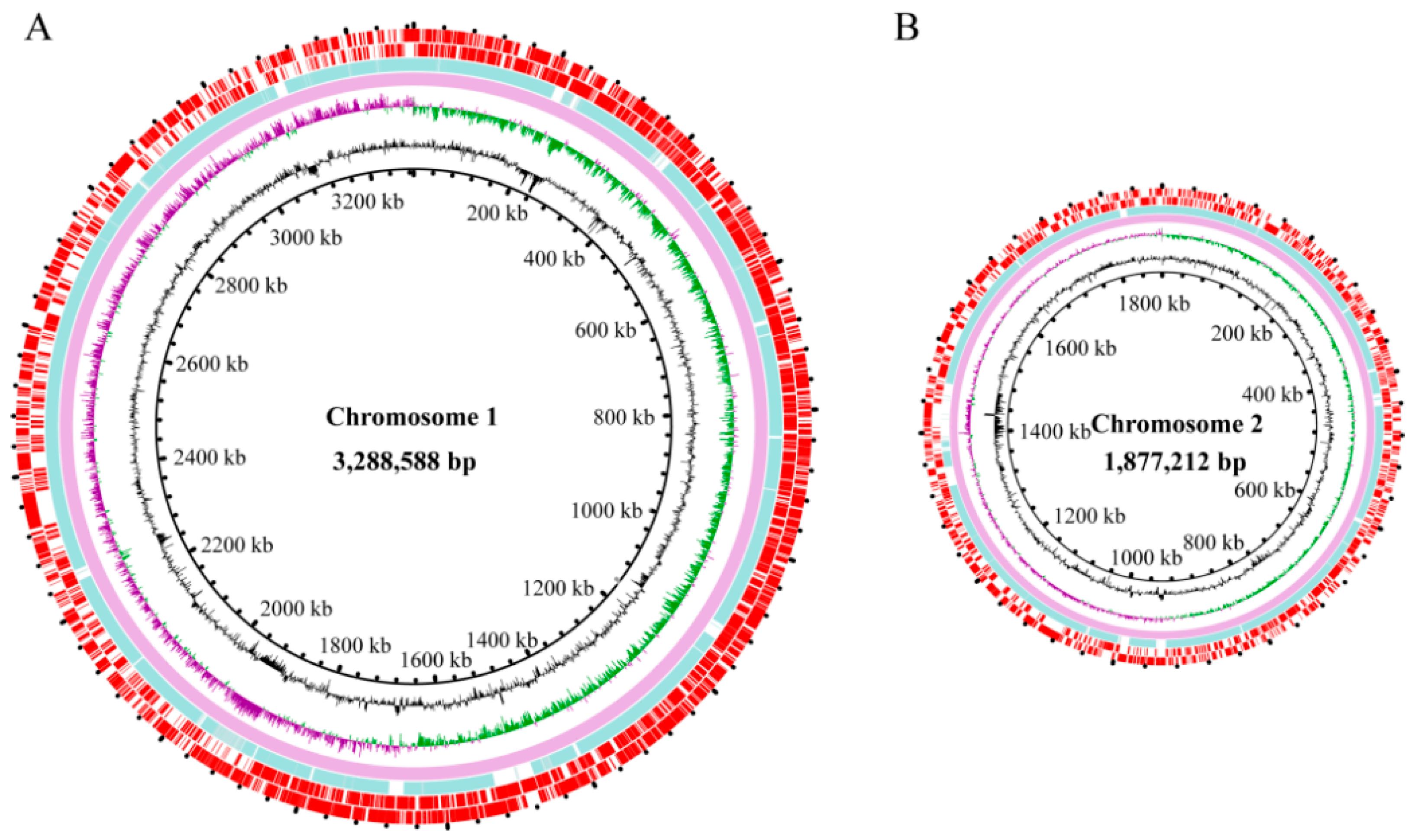
3.2. Genome Features of V. parahaemolyticus N10-18
3.3. Serotype and ST of V. parahaemolyticus N10-18
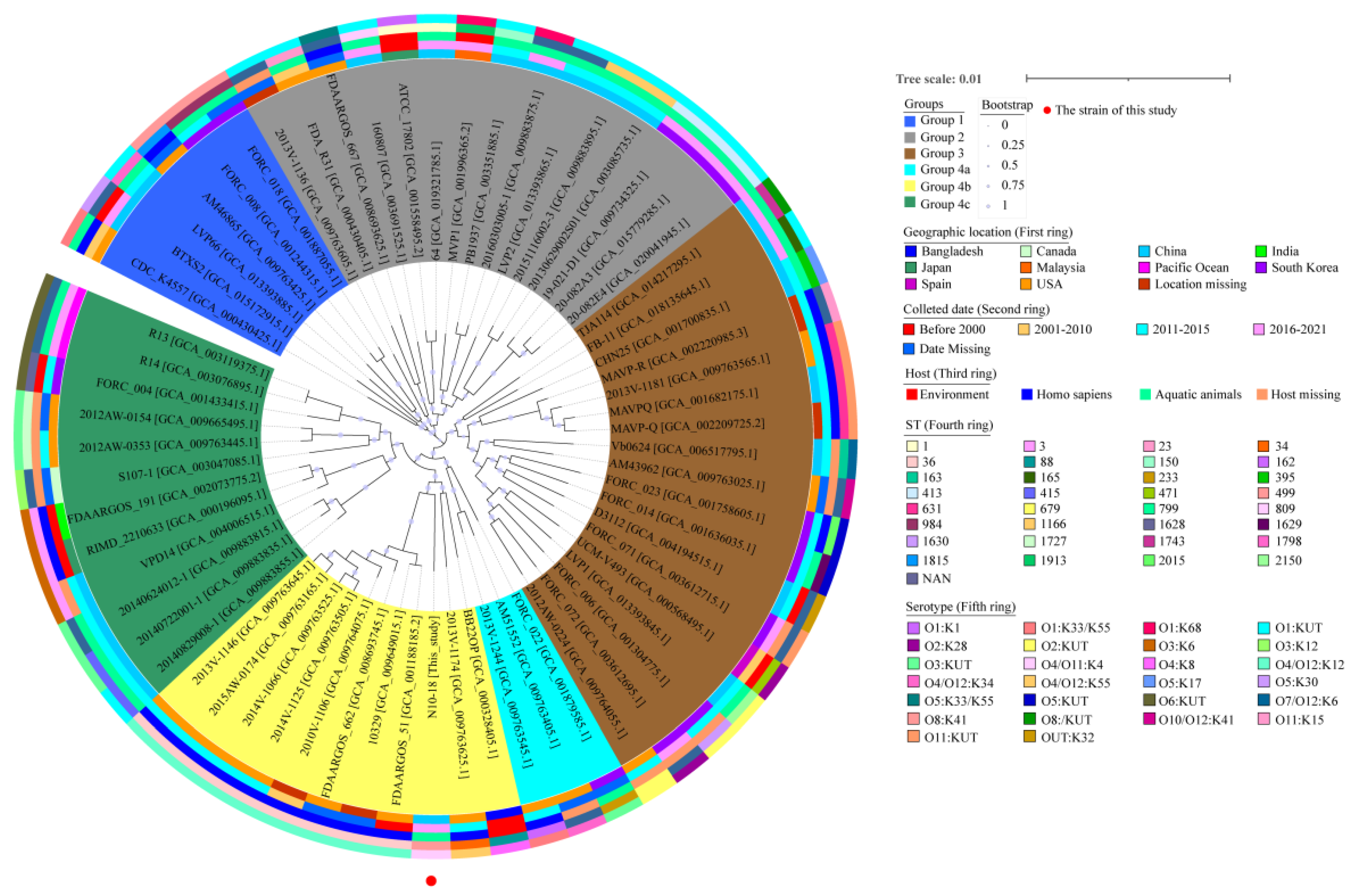
3.4. Phylogenetic Relatedness of V. parahaemolyticus N10-18
3.5. MGEs in the V. parahaemolyticus N10-18 Genome
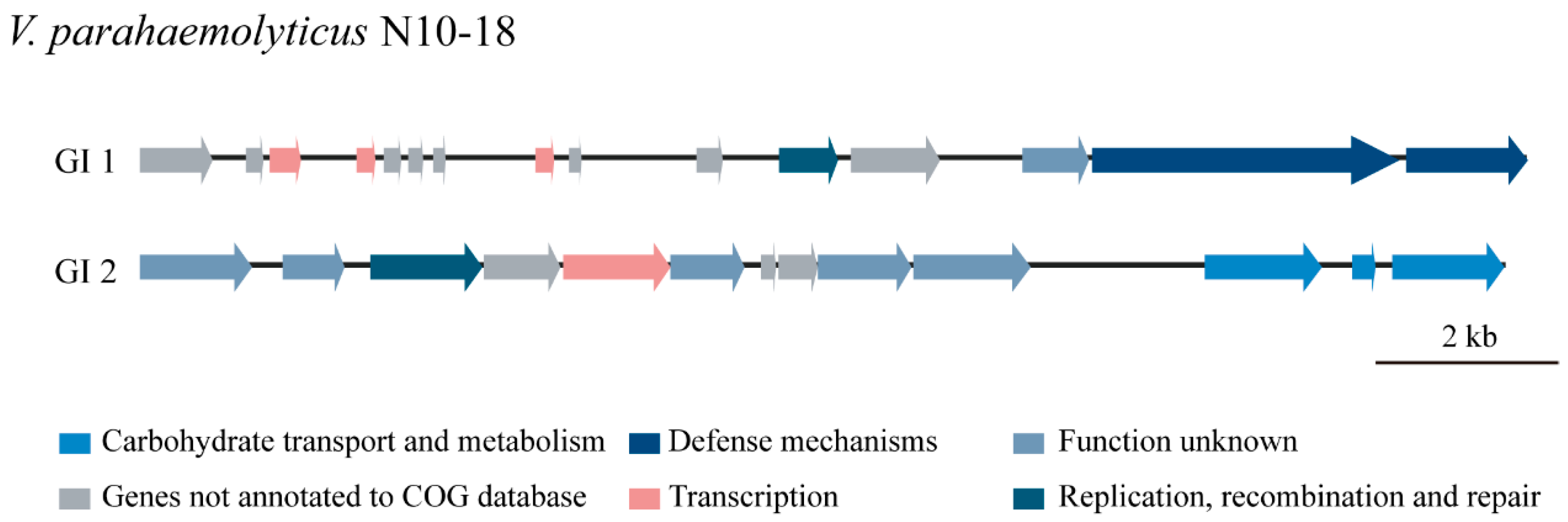
3.5.1. GIs
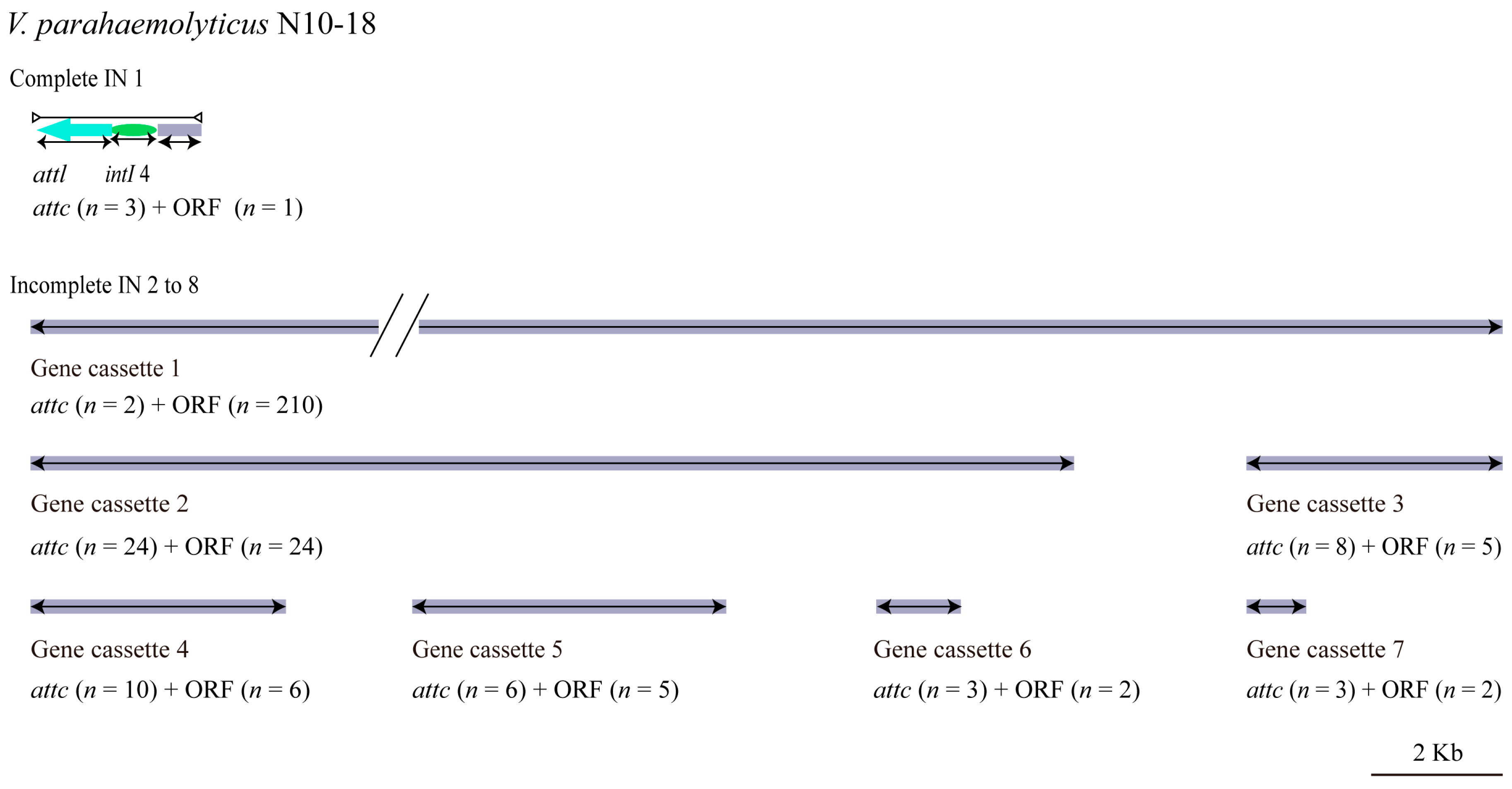
3.5.2. INs
3.5.3. ISs
3.6. Putative Virulence-Associated Genes
3.7. Heavy Metal and Antibiotic Resistance-Associated Genes
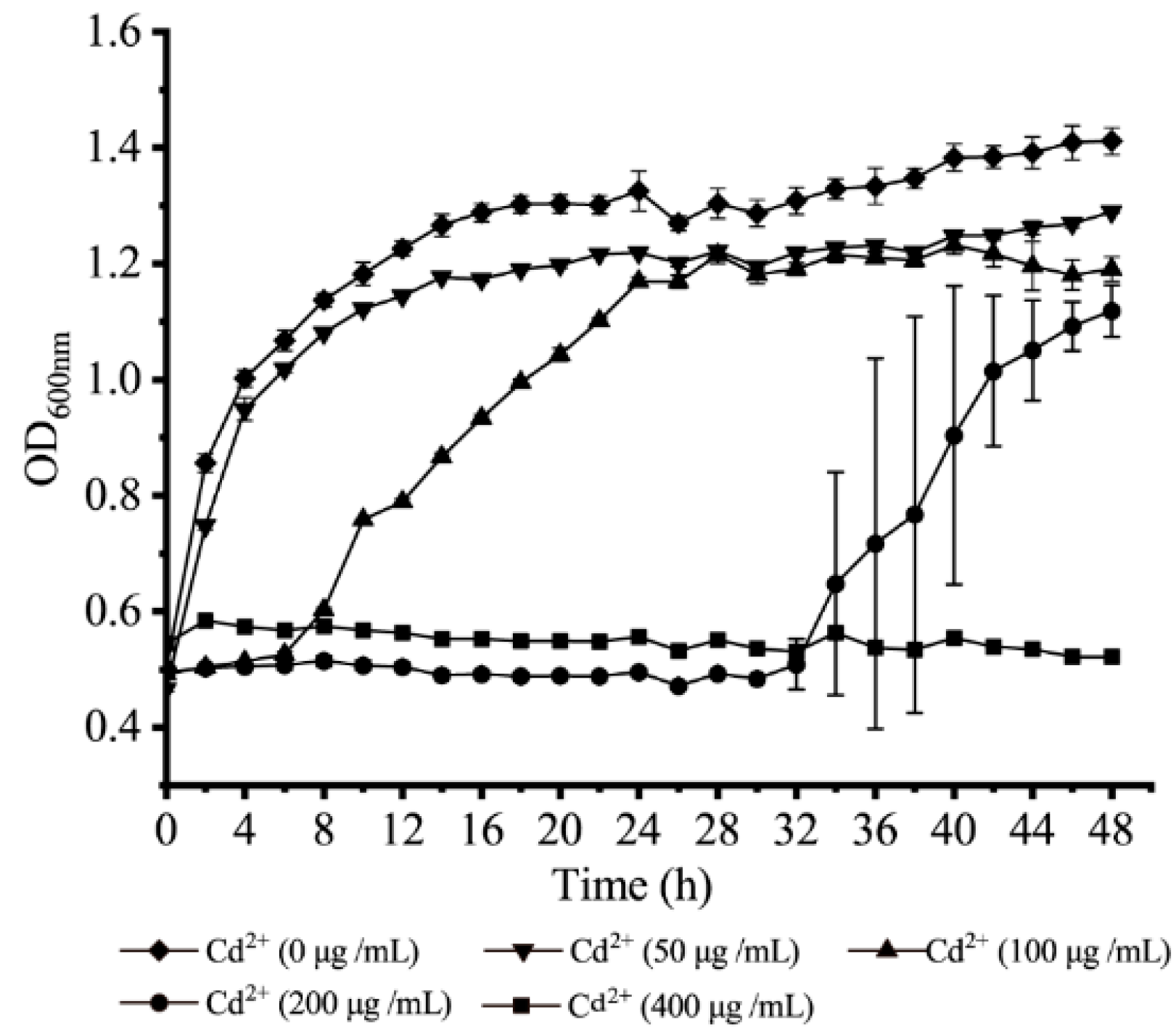
3.8. Survival of V. parahaemolyticus N10-18 under the Cd2+ (50 μg/mL) Stress
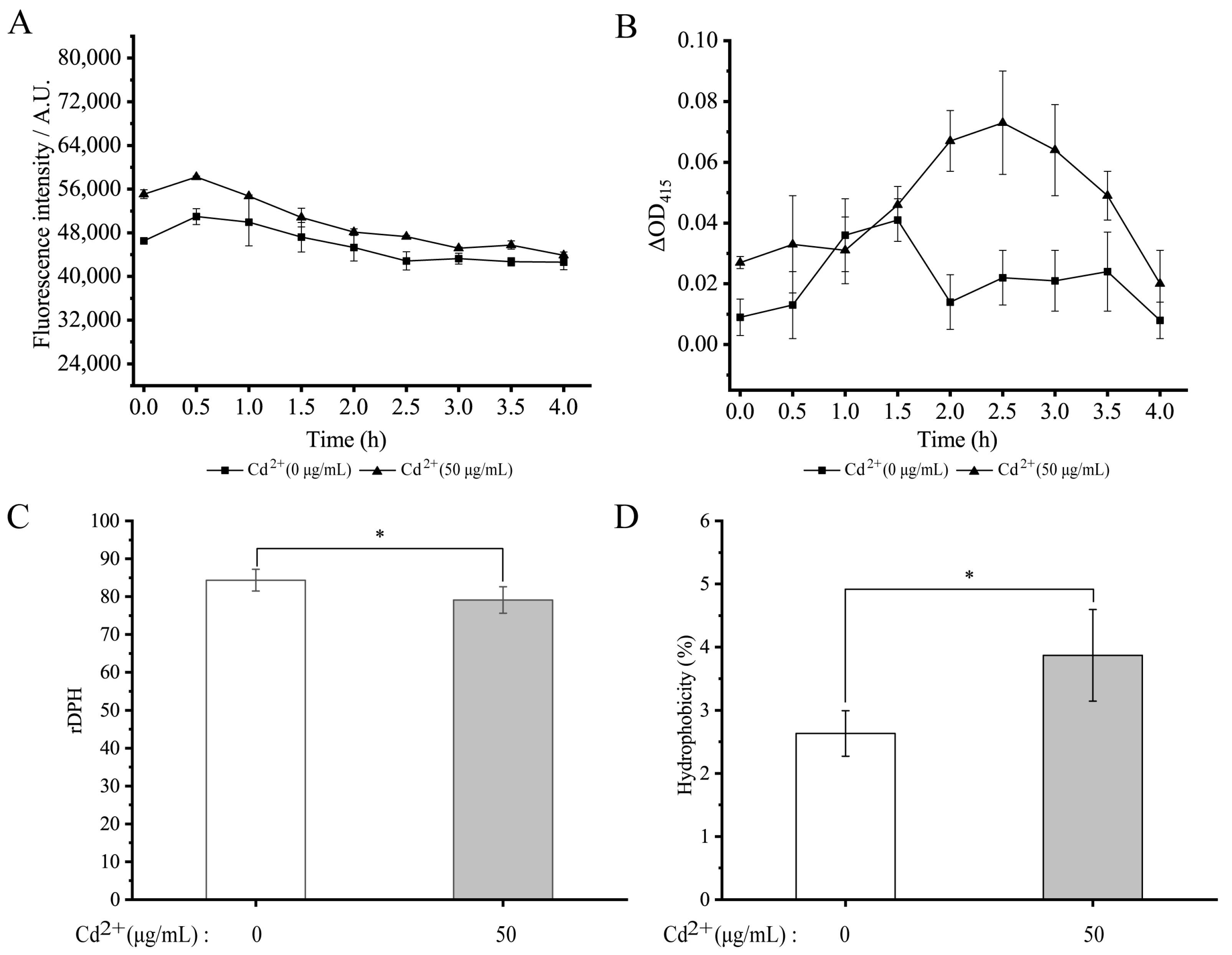
3.9. Changes in Cell Membrane Permeability and Fluidity and Cell Surface Hydrophobicity of V. parahaemolyticus N10-18 under the Cd2+ (50 μg/mL) Stress
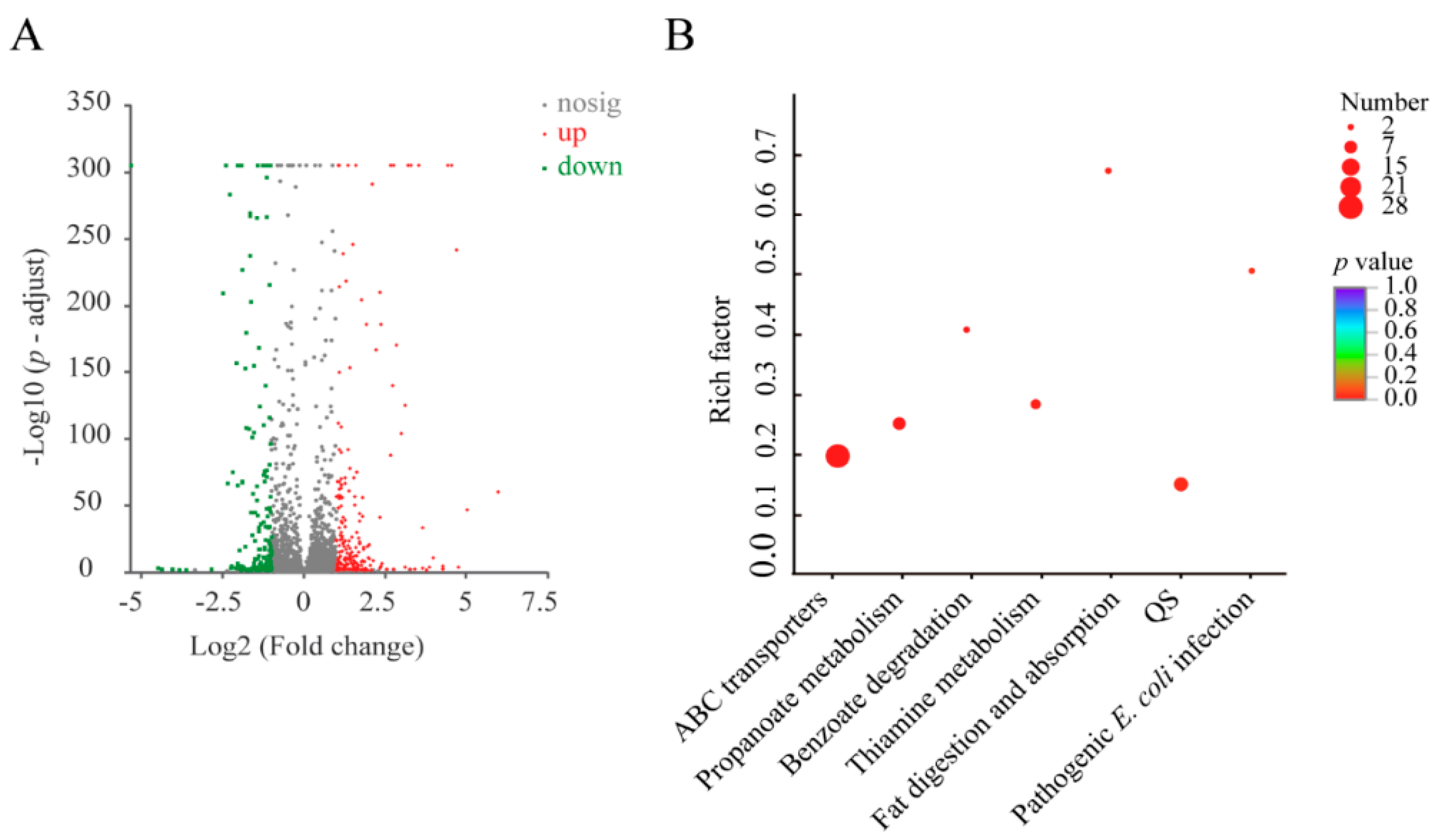
3.10. The Major Changed Metabolic Pathways Medicated by the Cd2+ (50 μg/mL) Stress in V. parahaemolyticus N10-18
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, J.; Bai, L.; Li, W.; Han, H.; Fu, P.; Ma, X.; Bi, Z.; Yang, X.; Zhang, X.; Zhen, S.; et al. Trends of foodborne diseases in China: Lessons from laboratory-based surveillance since 2011. Front. Med. 2017, 12, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Broberg, C.A.; Calder, T.J.; Orth, K. Vibrio parahaemolyticus cell biology and pathogenicity determinants. Microbes Infect. 2011, 13, 992–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujino, T.; Okuno, Y.; Nakada, D.; Aoyama, A.; Fukai, K.; Mukai, T.; Ueho, T. On the bacteriological examination of shirasu-food poisoning. Med. J. Osaka Univ. 1953, 4, 299–304. [Google Scholar]
- Ghenem, L.; Elhadi, N.; Alzahrani, F.; Nishibuchi, M. Vibrio parahaemolyticus: A Review on Distribution, Pathogenesis, Virulence Determinants and Epidemiology. Saudi J. Med. Med. Sci. 2017, 5, 93–103. [Google Scholar] [PubMed]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.-A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Zang, J.; Yu, W.; Shi, X.; Wu, Y. Occurrence and Identification of Pathogenic Vibrio Contaminants in Common Seafood Available in a Chinese Traditional Market in Qingdao, Shandong Province. Front. Microbiol. 2020, 11, 1488. [Google Scholar] [CrossRef] [PubMed]
- Eraghunath, P. Roles of thermostable direct hemolysin (TDH) and TDH-related hemolysin (TRH) in Vibrio parahaemolyticus. Front. Microbiol. 2015, 5, 805. [Google Scholar] [CrossRef]
- Li, Y.; Xie, T.; Pang, R.; Wu, Q.; Zhang, J.; Lei, T.; Xue, L.; Wu, H.; Wang, J.; Ding, Y.; et al. Food-Borne Vibrio parahaemolyticus in China: Prevalence, Antibiotic Susceptibility, and Genetic Characterization. Front. Microbiol. 2020, 11, 1670. [Google Scholar] [CrossRef]
- Dewi, R.R.; Hassan, L.; Daud, H.M.; Matori, M.F.; Nordin, F.; Ahmad, N.I.; Zakaria, Z. Prevalence and Antimicrobial Resistance of Escherichia coli, Salmonella and Vibrio Derived from Farm-Raised Red Hybrid Tilapia (Oreochromis spp.) and Asian Sea Bass (Lates calcarifer, Bloch 1970) on the West Coast of Peninsular Malaysia. Antibiotics 2022, 11, 136. [Google Scholar] [CrossRef]
- Prescott, J.; Barkovskii, A.L. In situ dynamics of Vibrio parahaemolyticus and Vibrio vulnificus in water, sediment and triploid Crassostrea virginica oysters cultivated in floating gear. J. Appl. Microbiol. 2021, 132, 3343–3354. [Google Scholar] [CrossRef]
- Pepi, M.; Focardi, S. Antibiotic-Resistant Bacteria in Aquaculture and Climate Change: A Challenge for Health in the Mediterranean Area. Int. J. Environ. Res. Public Health 2021, 18, 5723. [Google Scholar] [CrossRef]
- Ma, T.; Zhang, Y.; Hu, Q.; Han, M.; Li, X.; Zhang, Y.; Li, Z.; Shi, R. Accumulation Characteristics and Pollution Evaluation of Soil Heavy Metals in Different Land Use Types: Study on the Whole Region of Tianjin. Int. J. Environ. Res. Public Health 2022, 19, 10013. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Pan, B.; Shu, F.; Chen, X.; Xu, N.; Ni, J. Bioaccumulation of 35 metal(loid)s in organs of a freshwater mussel (Hyriopsis cumingii) and environmental implications in Poyang Lake, China. Chemosphere 2022, 307, 136150. [Google Scholar] [CrossRef] [PubMed]
- Balali-Mood, M.; Naseri, K.; Tahergorabi, Z.; Khazdair, M.R.; Sadeghi, M. Toxic Mechanisms of Five Heavy Metals: Mercury, Lead, Chromium, Cadmium, and Arsenic. Front. Pharmacol. 2021, 12, 643972. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Yu, T.; Yang, Y.; Yu, S.; Wu, J.; Lin, R.; Li, Y.; Fang, J.; Zhu, C. Co-occurrence of Antibiotic and Heavy Metal Resistance and Sequence Type Diversity of Vibrio parahaemolyticus Isolated From Penaeus vannamei at Freshwater Farms, Seawater Farms, and Markets in Zhejiang Province, China. Front. Microbiol. 2020, 11, 1294. [Google Scholar] [CrossRef]
- Fang, J.; Cheng, H.; Yu, T.; Jiang, H. Occurrence of Virulence Factors and Antibiotic and Heavy Metal Resistance in Vibrio parahaemolyticus Isolated from Pacific Mackerel at Markets in Zhejiang, China. J. Food Prot. 2020, 83, 1411–1419. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.-H.; Shin, Y.; Yu, H.; Kim, S.; So, J.-S. Antibiotic and heavy-metal resistance of Vibrio parahaemolyticus isolated from oysters in Korea. Mar. Pollut. Bull. 2018, 135, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zhang, X.; Wang, C.; Li, M.; Chen, J.; Xiong, J. Responses of sediment resistome, virulence factors and potential pathogens to decades of antibiotics pollution in a shrimp aquafarm. Sci. Total Environ. 2021, 794, 148760. [Google Scholar] [CrossRef] [PubMed]
- Prithvisagar, K.S.; Kumar, B.K.; Kodama, T.; Rai, P.; Iida, T.; Karunasagar, I.; Karunasagar, I. Whole genome analysis unveils genetic diversity and potential virulence determinants in Vibrio parahaemolyticus associated with disease outbreak among cultured Litopenaeus vannamei (Pacific white shrimp) in India. Virulence 2021, 12, 1936–1949. [Google Scholar] [CrossRef]
- Levy, S.E.; Boone, B.E. Next-Generation Sequencing Strategies. Cold Spring Harb. Perspect. Med. 2018, 9, a025791. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Peng, X.; Xie, L.; Chen, L. Survival and Genome Diversity of Vibrio parahaemolyticus Isolated from Edible Aquatic Animals. Diversity 2022, 14, 350. [Google Scholar] [CrossRef]
- Zhu, C.; Sun, B.; Liu, T.; Zheng, H.; Gu, W.; He, W.; Sun, F.; Wang, Y.; Yang, M.; Bei, W.; et al. Genomic and transcriptomic analyses reveal distinct biological functions for cold shock proteins (VpaCspA and VpaCspD) in Vibrio parahaemolyticus CHN25 during low-temperature survival. BMC Genom. 2017, 18, 436. [Google Scholar] [CrossRef] [Green Version]
- Su, C.; Chen, L. Virulence, resistance, and genetic diversity of Vibrio parahaemolyticus recovered from commonly consumed aquatic products in Shanghai, China. Mar. Pollut. Bull. 2020, 160, 111554. [Google Scholar] [CrossRef]
- Yao, W.; Yang, L.; Shao, Z.; Xie, L.; Chen, L. Identification of salt tolerance-related genes of Lactobacillus plantarum D31 and T9 strains by genomic analysis. Ann. Microbiol. 2020, 70, 10. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Wang, Y.; Yu, P.; Ren, S.; Zhu, Z.; Jin, Y.; Yan, J.; Peng, X.; Chen, L. Prophage-Related Gene VpaChn25_0724 Contributes to Cell Membrane Integrity and Growth of Vibrio parahaemolyticus CHN25. Front. Cell. Infect. Microbiol. 2020, 10, 595709. [Google Scholar] [CrossRef]
- Davis, E.M.; Sun, Y.; Liu, Y.; Kolekar, P.; Shao, Y.; Szlachta, K.; Mulder, H.L.; Ren, D.; Rice, S.V.; Wang, Z.; et al. SequencErr: Measuring and suppressing sequencer errors in next-generation sequencing data. Genome Biol. 2021, 22, 37. [Google Scholar] [CrossRef]
- Delcher, A.L.; Bratke, K.A.; Powers, E.C.; Salzberg, S.L. Identifying bacterial genes and endosymbiont DNA with Glimmer. Bioinformatics 2007, 23, 673–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, P.P.; Lowe, T.M. tRNAscan-SE: Searching for tRNA Genes in Genomic Sequences. Methods Mol. Biol. 2019, 1962, 1–14. [Google Scholar] [CrossRef]
- Jensen, L.J.; Julien, P.; Kuhn, M.; von Mering, C.; Muller, J.; Doerks, T.; Bork, P. eggNOG: Automated construction and annotation of orthologous groups of genes. Nucleic Acids Res. 2007, 36, D250–D254. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Yu, P.; Zheng, H.; Gu, W.; He, W.; Tang, Y.; Wang, Y.; Dong, Y.; Peng, X.; She, Q.; et al. Comparative genomics for non-O1/O139 Vibrio cholerae isolates recovered from the Yangtze River Estuary versus V. cholerae representative isolates from serogroup O1. Mol. Genet. Genom. 2018, 294, 417–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertelli, C.; Laird, M.R.; Williams, K.P.; Simon Fraser University Research Computing Group; Lau, B.Y.; Hoad, G.; Winsor, G.L.; Brinkman, F.S.L. IslandViewer 4: Expanded prediction of genomic islands for larger-scale datasets. Nucleic Acids Res. 2017, 45, W30–W35. [Google Scholar] [CrossRef] [PubMed]
- Fouts, D.E. Phage_Finder: Automated identification and classification of prophage regions in complete bacterial genome sequences. Nucleic Acids Res. 2006, 34, 5839–5851. [Google Scholar] [CrossRef]
- Cury, J.; Jové, T.; Touchon, M.; Néron, B.; Rocha, E.P. Identification and analysis of integrons and cassette arrays in bacterial genomes. Nucleic Acids Res. 2016, 44, 4539–4550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siguier, P.; Gourbeyre, E.; Chandler, M. Bacterial insertion sequences: Their genomic impact and diversity. FEMS Microbiol. Rev. 2014, 38, 865–891. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Jiang, Y.; Shi, X.; Li, Y.; Jiang, M.; Lin, Y.; Qiu, Y.; Zuo, L.; Deng, Y.; Lin, Z.; et al. Simultaneous Identification of Clinically Common Vibrio parahaemolyticus Serotypes Using Probe Melting Curve Analysis. Front. Cell. Infect. Microbiol. 2019, 9, 385. [Google Scholar] [CrossRef]
- Bian, S.; Zeng, W.; Li, Q.; Li, Y.; Wong, N.-K.; Jiang, M.; Zuo, L.; Hu, Q.; Li, L. Genetic Structure, Function, and Evolution of Capsule Biosynthesis Loci in Vibrio parahaemolyticus. Front. Microbiol. 2021, 11, 546150. [Google Scholar] [CrossRef]
- Chen, Y.; Dai, J.; Jr, J.G.M.; Johnson, J.A. Genetic analysis of the capsule polysaccharide (K antigen) and exopolysaccharide genes in pandemic Vibrio parahaemolyticus O3:K6. BMC Microbiol. 2010, 10, 374. [Google Scholar] [CrossRef] [Green Version]
- Lun, J.; Zhang, S.; Dong, Y.; Xie, S.; Mai, H.; Mei, Y.; Hu, Z. Immunological cross-reactivity and cross-protection of outer membrane protein OmpU among Vibrio species. Acta Microbiol. Sin. 2016, 56, 867–879. [Google Scholar]
- Guo, X.; Liu, B.; Chen, M.; Wang, Y.; Wang, L.; Chen, H.; Wang, Y.; Tu, L.; Zhang, X.; Feng, L. Genetic and serological identification of three Vibrio parahaemolyticus strains as candidates for novel provisional O serotypes. Int. J. Food Microbiol. 2017, 245, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, Y.; Yao, W.; Wu, T.; Zhu, Q.; Zhang, Y.; Ye, H.; Wang, R.; Zheng, S.; Yu, F.; et al. A new emerging serotype of Vibrio parahaemolyticus in China is rapidly becoming the main epidemic strain. Clin. Microbiol. Infect. 2019, 26, 644.e1–644.e7. [Google Scholar] [CrossRef]
- González-Escalona, N.; Martinez-Urtaza, J.; Romero, J.; Espejo, R.T.; Jaykus, L.-A.; DePaola, A. Determination of Molecular Phylogenetics of Vibrio parahaemolyticus Strains by Multilocus Sequence Typing. J. Bacteriol. 2008, 190, 2831–2840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic orthology inference for comparative genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef] [PubMed]
- Buck, J. The Plate Count in Aquatic Microbiology. Nativ. Aquat. Bact. Enumer. Act. Ecol. 1979, 1, 19–28. [Google Scholar] [CrossRef]
- Harman, R.M.; Yang, S.; He, M.K.; Van de Walle, G.R. Antimicrobial peptides secreted by equine mesenchymal stromal cells inhibit the growth of bacteria commonly found in skin wounds. Stem Cell Res. Ther. 2017, 8, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, H.R.; Sugimoto, Y.; Aoki, T. Ovotransferrin antimicrobial peptide (OTAP-92) kills bacteria through a membrane damage mechanism. Biochim. Biophys. Acta (BBA) Gen. Subj. 2000, 1523, 196–205. [Google Scholar] [CrossRef]
- Voss, D.; Montville, T.J. 1,6-Diphenyl-1,3,5-hexatrine as a reporter of inner spore membrane fluidity in Bacillus subtilis and Alicyclobacillus acidoterrestris. J. Microbiol. Methods 2014, 96, 101–103. [Google Scholar] [CrossRef]
- Yan, F.; Dang, Q.; Liu, C.; Yan, J.; Wang, T.; Fan, B.; Cha, D.; Li, X.; Liang, S.; Zhang, Z. 3,6-O-[N-(2-Aminoethyl)-acetamide-yl]-chitosan exerts antibacterial activity by a membrane damage mechanism. Carbohydr. Polym. 2016, 149, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Wang, Y.; Sun, M.; Xu, Y.; Chen, L. Antibacterial Activity and Components of the Methanol-Phase Extract from Rhizomes of Pharmacophagous Plant Alpinia officinarum Hance. Molecules 2022, 27, 4308. [Google Scholar] [CrossRef] [PubMed]
- Almuhaideb, E.; Chintapenta, L.K.; Abbott, A.; Parveen, S.; Ozbay, G. Assessment of Vibrio parahaemolyticus levels in oysters (Crassostrea virginica) and seawater in Delaware Bay in relation to environmental conditions and the prevalence of molecular markers to identify pathogenic Vibrio parahaemolyticus strains. PLoS ONE 2020, 15, e0242229. [Google Scholar] [CrossRef] [PubMed]
- Ciufo, S.; Kannan, S.; Sharma, S.; Badretdin, A.; Clark, K.; Turner, S.; Brover, S.; Schoch, C.L.; Kimchi, A.; DiCuccio, M. Using average nucleotide identity to improve taxonomic assignments in prokaryotic genomes at the NCBI. Int. J. Syst. Evol. Microbiol. 2018, 68, 2386–2392. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.; Guo, X.; Tian, X.; Liu, F.; Wang, L.; Wu, J.; Zhang, S.; Li, S.; Liu, B. Developing a novel molecular serotyping system based on capsular polysaccharide synthesis gene clusters of Vibrio parahaemolyticus. Int. J. Food Microbiol. 2019, 309, 108332. [Google Scholar] [CrossRef]
- Chi, P.-S.; Wong, H.-C. Variation of genomic islands and flanking fragments in Vibrio parahaemolyticus isolates from environmental and clinical sources in Taiwan. Int. J. Food Microbiol. 2017, 259, 68–76. [Google Scholar] [CrossRef] [PubMed]
- An, X.-L.; Chen, Q.; Zhu, D.; Zhu, Y.-G.; Gillings, M.R.; Su, J.-Q. Impact of Wastewater Treatment on the Prevalence of Integrons and the Genetic Diversity of Integron Gene Cassettes. Appl. Environ. Microbiol. 2018, 84, e02766-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabbagh, P.; Rajabnia, M.; Maali, A.; Ferdosi-Shahandashti, E. Integron and its role in antimicrobial resistance: A literature review on some bacterial pathogens. Iran. J. Basic Med. Sci. 2021, 24, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.M.; Collis, C.M. Mobile gene cassettes and integrons: Capture and spread of genes by site-specific recombination. Mol. Microbiol. 1995, 15, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Mazel, D.; Dychinco, B.; Webb, V.A.; Davies, J. A Distinctive Class of Integron in the Vibrio cholerae Genome. Science 1998, 280, 605–608. [Google Scholar] [CrossRef]
- De Nisco, N.J.; Casey, A.K.; Kanchwala, M.; Lafrance, A.E.; Coskun, F.S.; Kinch, L.N.; Grishin, N.V.; Xing, C.; Orth, K. Manipulation of IRE1-Dependent MAPK Signaling by a Vibrio Agonist-Antagonist Effector Pair. mSystems 2021, 6, e00872-20. [Google Scholar] [CrossRef] [PubMed]
- Lian, L.; Xue, J.; Li, W.; Ren, J.; Tang, F.; Liu, Y.; Xue, F.; Dai, J. VscF in T3SS1 Helps to Translocate VPA0226 in Vibrio parahaemolyticus. Front. Cell. Infect. Microbiol. 2021, 11, 652432. [Google Scholar] [CrossRef] [PubMed]
- Gu, D.; Zhang, Y.; Wang, Q.; Zhou, X. S-nitrosylation-mediated activation of a histidine kinase represses the type 3 secretion system and promotes virulence of an enteric pathogen. Nat. Commun. 2020, 11, 5777. [Google Scholar] [CrossRef]
- Rahman, M.M.; Machuca, M.A.; Roujeinikova, A. Bioinformatics analysis and biochemical characterisation of ABC transporter-associated periplasmic substrate-binding proteins ModA and MetQ from Helicobacter pylori strain SS1. Biophys. Chem. 2021, 272, 106577. [Google Scholar] [CrossRef] [PubMed]
- Krachler, A.M.; Ham, H.; Orth, K. Outer membrane adhesion factor multivalent adhesion molecule 7 initiates host cell binding during infection by Gram-negative pathogens. Proc. Natl. Acad. Sci. USA 2011, 108, 11614–11619. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.-K.; Wang, C.-J.; Chew, Y.; Wang, P.-C.; Yin, H.-S.; Kao, M.-C. Functional characterization of Helicobacter pylori 26695 sedoheptulose 7-phosphate isomerase encoded by hp0857 and its association with lipopolysaccharide biosynthesis and adhesion. Biochem. Biophys. Res. Commun. 2016, 477, 794–800. [Google Scholar] [CrossRef] [PubMed]
- Eisemann, T.; Langelier, M.-F.; Pascal, J.M. Structural and functional analysis of parameters governing tankyrase-1 interaction with telomeric repeat-binding factor 1 and GDP-mannose 4,6-dehydratase. J. Biol. Chem. 2019, 294, 14574–14590. [Google Scholar] [CrossRef]
- Cesur, M.F.; Siraj, B.; Uddin, R.; Durmuş, S.; Çakır, T. Network-Based Metabolism-Centered Screening of Potential Drug Targets in Klebsiella pneumoniae at Genome Scale. Front. Cell. Infect. Microbiol. 2020, 9, 447. [Google Scholar] [CrossRef] [Green Version]
- Pal, C.; Bengtsson-Palme, J.; Rensing, C.; Kristiansson, E.; Larsson, D.G.J. BacMet: Antibacterial biocide and metal resistance genes database. Nucleic Acids Res. 2013, 42, D737–D743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.W.; Glickmann, E.; Cooksey, D.A. Chromosomal locus for cadmium resistance in Pseudomonas putida consisting of a cadmium-transporting ATPase and a MerR family response regulator. Appl. Environ. Microbiol. 2001, 67, 1437–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Wang, L.; Zheng, X.; Cohen, M.; Li, X. Cross-Kingdom Comparative Transcriptomics Reveals Conserved Genetic Modules in Response to Cadmium Stress. mSystems 2021, 6, e0118921. [Google Scholar] [CrossRef]
- Che, R.-X.; Xing, X.-X.; Liu, X.; Qu, Q.-W.; Chen, M.; Yu, F.; Ma, J.-X.; Chen, X.-R.; Zhou, Y.-H.; God’Spower, B.-O.; et al. Analysis of multidrug resistance in Streptococcus suis ATCC 700794 under tylosin stress. Virulence 2019, 10, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Kary, S.C.; Yoneda, J.R.K.; Olshefsky, S.C.; Stewart, L.A.; West, S.B.; Cameron, A.D.S. The Global Regulatory Cyclic AMP Receptor Protein (CRP) Controls Multifactorial Fluoroquinolone Susceptibility in Salmonella enterica Serovar Typhimurium. Antimicrob. Agents Chemother. 2017, 61, e01666-17. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.-W.; Zhu, J.-H.; Javid, B. Clinically relevant mutations in mycobacterial LepA cause rifampicin-specific phenotypic resistance. Sci. Rep. 2020, 10, 8402. [Google Scholar] [CrossRef]
- Ortiz-Padilla, M.; Portillo-Calderón, I.; Maldonado, N.; Rodríguez-Martínez, J.; de Gregorio-Iaria, B.; Merino-Bohórquez, V.; Rodríguez-Baño, J.; Pascual, Á.; Docobo-Pérez, F. Role of inorganic phosphate concentrations in in vitro activity of fosfomycin. Clin. Microbiol. Infect. 2021, 28, 302.e1–302.e4. [Google Scholar] [CrossRef]
- El-Razik, K.A.; Arafa, A.; Hedia, R.H.; Ibrahim, E. Tetracycline resistance phenotypes and genotypes of coagulase-negative staphylococcal isolates from bubaline mastitis in Egypt. Veter World 2017, 10, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Moseng, M.A.; Lyu, M.; Pipatpolkai, T.; Glaza, P.; Emerson, C.C.; Stewart, P.L.; Stansfeld, P.J.; Yu, E.W. Cryo-EM Structures of CusA Reveal a Mechanism of Metal-Ion Export. mBio 2021, 12, e00452-21. [Google Scholar] [CrossRef] [PubMed]
- Ackerley, D.F.; Gonzalez, C.F.; Keyhan, M.; Blake, R., 2nd; Matin, A. Mechanism of chromate reduction by the Escherichia coli protein, NfsA, and the role of different chromate reductases in minimizing oxidative stress during chromate reduction. Environ. Microbiol. 2004, 6, 851–860. [Google Scholar] [CrossRef]
- Alboghobeish, H.; Tahmourespour, A.; Doudi, M. The study of Nickel Resistant Bacteria (NiRB) isolated from wastewaters polluted with different industrial sources. J. Environ. Health Sci. Eng. 2014, 12, 44. [Google Scholar] [CrossRef] [Green Version]
- Gu, R.; Gao, J.; Dong, L.; Liu, Y.; Li, X.; Bai, Q.; Jia, Y.; Xiao, H. Chromium metabolism characteristics of coexpression of ChrA and ChrT gene. Ecotoxicol. Environ. Saf. 2020, 204, 111060. [Google Scholar] [CrossRef] [PubMed]
- Pombinho, R.; Camejo, A.; Vieira, A.; Reis, O.; Carvalho, F.; Almeida, M.T.; Pinheiro, J.C.; Sousa, S.; Cabanes, D. Listeria monocytogenes CadC Regulates Cadmium Efflux and Fine-tunes Lipoprotein Localization to Escape the Host Immune Response and Promote Infection. J. Infect. Dis. 2017, 215, 1468–1479. [Google Scholar] [CrossRef] [Green Version]
- Decré, D.; Arlet, G.; Bergogne-Bérézin, E.; Philippon, A. Identification of a carbenicillin-hydrolyzing beta-lactamase in Alcaligenes denitrificans subsp. xylosoxydans. Antimicrob. Agents Chemother. 1995, 39, 771–774. [Google Scholar] [CrossRef] [Green Version]
- Håkonsholm, F.; Lunestad, B.T.; Aguirre Sánchez, J.R.; Martinez-Urtaza, J.; Marathe, N.P.; Svanevik, C.S. Vibrios from the Norwegian marine environment: Characterization of associated antibiotic resistance and virulence genes. Microbiologyopen 2020, 9, e1093. [Google Scholar] [CrossRef] [PubMed]
- Bessa, L.J.; Ferreira, M.; Gameiro, P. Evaluation of membrane fluidity of multidrug-resistant isolates of Escherichia coli and Staphylococcus aureus in presence and absence of antibiotics. J. Photochem. Photobiol. B Biol. 2018, 181, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.R.; Garcia, G.E.; Geissler, P.L. Membrane hydrophobicity determines the activation free energy of passive lipid transport. Biophys. J. 2021, 120, 3718–3731. [Google Scholar] [CrossRef]
- Zhengwei, Z.; Fang, W.; Lee, H.Y.; Yang, Z. Responses of Azorhizobium caulinodans to cadmium stress. FEMS Microbiol. Ecol. 2005, 54, 455–461. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, L.; Liu, P.; Jin, Y.; Qin, S.; Chen, L. Identification of Antibacterial Components in the Methanol-Phase Extract from Edible Herbaceous Plant Rumex madaio Makino and Their Antibacterial Action Modes. Molecules 2022, 27, 660. [Google Scholar] [CrossRef]
- Garvey, G.S.; Rocco, C.J.; Escalante-Semerena, J.C.; Rayment, I. The three-dimensional crystal structure of the PrpF protein of Shewanella oneidensis complexed withtrans-aconitate: Insights into its biological function. Protein Sci. 2007, 16, 1274–1284. [Google Scholar] [CrossRef] [Green Version]
- Adekoya, O.A.; Helland, R.; Willassen, N.-P.; Sylte, I. Comparative sequence and structure analysis reveal features of cold adaptation of an enzyme in the thermolysin family. Proteins Struct. Funct. Bioinform. 2005, 62, 435–449. [Google Scholar] [CrossRef] [PubMed]
- Rodionov, D.A.; Leyn, S.A.; Li, X.; Rodionova, I.A. A novel transcriptional regulator related to thiamine phosphate synthase controls thiamine metabolism genes in Archaea. J. Bacteriol. 2017, 199, e00743. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; de Groot, A.; Boeren, S.; Abee, T.; Smid, E.J. Lactococcus lactis Mutants Obtained from Laboratory Evolution Showed Elevated Vitamin K2 Content and Enhanced Resistance to Oxidative Stress. Front. Microbiol. 2021, 12, 746770. [Google Scholar] [CrossRef]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metal toxicity and the environment. Mol. Clin. Environ. Toxicol. 2012, 101, 133–164. [Google Scholar] [CrossRef] [Green Version]
- Boyd, R.S. Heavy Metal Pollutants and Chemical Ecology: Exploring New Frontiers. J. Chem. Ecol. 2010, 36, 46–58. [Google Scholar] [CrossRef]
- Chibani, C.M.; Roth, O.; Liesegang, H.; Wendling, C.C. Genomic variation among closely related Vibrio alginolyticus strains is located on mobile genetic elements. BMC Genom. 2020, 21, 354. [Google Scholar] [CrossRef]
- Baltazar, M.; Bourgeois-Nicolaos, N.; Larroudé, M.; Couet, W.; Uwajeneza, S.; Doucet-Populaire, F.; Ploy, M.-C.; Da Re, S. Activation of class 1 integron integrase is promoted in the intestinal environment. PLoS Genet. 2022, 18, e1010177. [Google Scholar] [CrossRef]
- He, Z.; Li, T.; Wang, J.; Luo, D.; Ning, N.; Li, Z.; Chen, F.; Wang, H. AtaT Improves the Stability of Pore-Forming Protein EspB by Acetylating Lysine 206 to Enhance Strain Virulence. Front. Microbiol. 2021, 12, 627141. [Google Scholar] [CrossRef]
- Zhang, Y.; Deng, Y.; Feng, J.; Guo, Z.; Chen, H.; Wang, B.; Hu, J.; Lin, Z.; Su, Y. Functional characterization of VscCD, an important component of the type III secretion system of Vibrio harveyi. Microb. Pathog. 2021, 157, 104965. [Google Scholar] [CrossRef]
- Nadella, R.K.; Panda, S.K.; Badireddy, M.R.; Kurcheti, P.P.; Raman, R.P.; Mothadaka, M.P. Multi-drug resistance, integron and transposon-mediated gene transfer in heterotrophic bacteria from Penaeus vannamei and its culture environment. Environ. Sci. Pollut. Res. 2022, 29, 37527–37542. [Google Scholar] [CrossRef]
- Liu, B.-T.; Liao, X.-P.; Yang, S.-S.; Wang, X.-M.; Li, L.-L.; Sun, J.; Yang, Y.-R.; Fang, L.-X.; Li, L.; Zhao, D.-H.; et al. Detection of mutations in the gyrA and parC genes in Escherichia coli isolates carrying plasmid-mediated quinolone resistance genes from diseased food-producing animals. J. Med. Microbiol. 2012, 61, 1591–1599. [Google Scholar] [CrossRef]
- Liu, B.-T.; Liao, X.-P.; Liao, S.-D.; Sun, N.; Zhang, M.-J.; Yang, S.-S.; Li, L.-L.; Sun, J.; Yang, Y.-R.; Liu, Y.-H. Plasmid-mediated quinolone resistance determinant qepA1 and extended-spectrum β-lactamase gene bla CTX-M-14 co-located on the same plasmid in two Escherichia coli strains from China. J. Med. Microbiol. 2012, 61, 603–605. [Google Scholar] [CrossRef]
- Liu, Q.; Han, L.; Li, B.; Sun, J.; Ni, Y. Virulence Characteristic and MLST-agr Genetic Background of High-Level Mupirocin-Resistant, MRSA Isolates from Shanghai and Wenzhou, China. PLoS ONE 2012, 7, e37005. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.-W.; Chang, H.-J.; Chia, J.-H.; Kuo, A.-J.; Wu, T.-L.; Lee, M.-H. Outcomes and characteristics of ertapenem-nonsusceptible Klebsiella pneumoniae bacteremia at a university hospital in Northern Taiwan: A matched case-control study. J. Microbiol. Immunol. Infect. 2012, 45, 113–119. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Bao, Z.; Feng, H.; Chen, L.; Li, Q. Nitric oxide enhances resistance of Pleurotus eryngii to cadmium stress by alleviating oxidative damage and regulating of short-chain dehydrogenase/reductase family. Environ. Sci. Pollut. Res. 2022, 29, 53036–53049. [Google Scholar] [CrossRef]
- Khan, Z.; Rehman, A.; Nisar, M.A.; Zafar, S.; Zerr, I. Biosorption behavior and proteomic analysis of Escherichia coli P4 under cadmium stress. Chemosphere 2017, 174, 136–147. [Google Scholar] [CrossRef]
- Bhuiyan, N.A.; Ansaruzzaman, M.; Kamruzzaman, M.; Alam, K.; Chowdhury, N.R.; Nishibuchi, M.; Faruque, S.M.; Sack, D.A.; Takeda, Y.; Nair, G.B. Prevalence of the Pandemic Genotype of Vibrio parahaemolyticus in Dhaka, Bangladesh, and Significance of Its Distribution across Different Serotypes. J. Clin. Microbiol. 2002, 40, 284–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delmar, J.A.; Su, C.-C.; Yu, E.W. Structural mechanisms of heavy-metal extrusion by the Cus efflux system. BioMetals 2013, 26, 593–607. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, G.; Xiong, A.; Uebel, M.; Singh, V.K.; Jayaswal, R.K. Molecular Characterization of the Iron-Hydroxamate Uptake System in Staphylococcus aureus. Appl. Environ. Microbiol. 2001, 67, 1001–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abushattal, S.; Vences, A.; Osorio, C.R. A virulence gene typing scheme for Photobacterium damselae subsp. piscicida, the causative agent of fish photobacteriosis, reveals a high prevalence of plasmid-encoded virulence factors and of type III secretion system genes. Aquaculture 2020, 521, 735057. [Google Scholar] [CrossRef]
- Akeda, Y.; Okayama, K.; Kimura, T.; Dryselius, R.; Kodama, T.; Oishi, K.; Iida, T.; Honda, T. Identification and characterization of a type III secretion-associated chaperone in the type III secretion system 1 of Vibrio parahaemolyticus. FEMS Microbiol. Lett. 2009, 296, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Tam, V.; Suzuki, M.; Coughlin, M.; Saslowsky, D.; Biswas, K.; Lencer, W.; Faruque, S.M.; Mekalanos, J.J. Functional Analysis of VopF Activity Required for Colonization in Vibrio cholerae. mBio 2010, 1, e00289-10. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, R.; Qi, X.; Zhou, B.; Wang, J.; Chen, Y.; Zhang, H. Epidemiology of foodborne disease outbreaks caused by Vibrio parahaemolyticus during 2010–2014 in Zhejiang Province, China. Food Control. 2017, 77, 110–115. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genome Feature | V. parahaemolyticus N10-18 |
|---|---|
| Genome size (bp) | 4,910,080 |
| G + C (%) | 45.46 |
| DNA Scaffold | 70 |
| Total predicted gene | 4653 |
| Protein-coding gene | 4565 |
| RNA gene | 143 |
| Genes assigned to COG | 3843 |
| Genes with unknown function | 722 |
| Transposase gene | 10 |
| GI | 2 |
| Prophage | 0 |
| IN | 8 |
| IS | 1 |
| Heavy Metal and Antibiotic Agent | Resistance Gene | Reference |
|---|---|---|
| Heavy metal | ||
| As | arsCR, pstABCS | [65] |
| Cu | actP, copAB, corC, cueR, cusABRS, cutAC | [65,73] |
| Cr | nfsA | [65,74] |
| Ni | nirBD | [65,75] |
| Zn | zntAR, znuABC, zur | [66,67] |
| Zn, Hg | smtA | [65] |
| Cr, Fe | chrAR | [65,76] |
| W, Mo | modABC | [65] |
| Cr, Te, Se | recG, ruvB | [65] |
| Cd, Zn, Pb | cadC | [65,77] |
| Cd, Zn, Hg, Cu | dsbABC | [65] |
| Antimicrobial agent | ||
| Beta-lactamases | blaCARB-21 | [78] |
| Elfamycin | tuf | [68] |
| Fluoroquinolone | crp | [69] |
| Fosfomycin | UhpT | [71] |
| Peptide, rifamycin | rpoB | [70] |
| Tetracycline | Tet (34), Tet (35) | [72,79] |
| Metabolic Pathway | Gene | Gene ID | Fold Change | Description |
|---|---|---|---|---|
| ABC transporters | livH | Vp_N10_18_2959 | 0.061 | Branched-chain amino acid ABC transporter permease |
| znuB | Vp_N10_18_4101 | 0.081 | Metal ABC transporter permease | |
| malE | Vp_N10_18_1557 | 0.252 | Maltose ABC transporter substrate-binding protein MalE | |
| malK | Vp_N10_18_1556 | 0.263 | Maltose/maltodextrin import ATP-binding protein MalK | |
| rbsB | Vp_N10_18_3026 | 0.325 | Ribose ABC transporter substrate-binding protein RbsB | |
| aapP | Vp_N10_18_2527 | 0.355 | Arginine ABC transporter ATP-binding protein | |
| rbsD | Vp_N10_18_3023 | 0.378 | D-ribose pyranase | |
| rbsC | Vp_N10_18_3025 | 0.428 | Ribose ABC transporter permease | |
| aapJ | Vp_N10_18_2530 | 0.441 | Amino acid ABC transporter substrate-binding protein | |
| oppF | Vp_N10_18_2154 | 0.478 | Hypothetical protein VIBHAR_00643 | |
| yejA | Vp_N10_18_2156 | 0.494 | Extracellular solute-binding protein | |
| aapQ | Vp_N10_18_2529 | 0.496 | Amino acid ABC transporter permease | |
| mlaF | Vp_N10_18_2720 | 0.500 | ATP-binding cassette domain-containing protein | |
| proV | Vp_N10_18_0094 | 2.147 | Glycine betaine/L-proline transport ATP binding subunit | |
| afuA | Vp_N10_18_1887 | 2.243 | Iron ABC transporter substrate-binding protein | |
| fhuB | Vp_N10_18_1520 | 2.270 | Fe3+-hydroxamate ABC transporter permease FhuB | |
| thiY | Vp_N10_18_1092 | 2.400 | Hypothetical protein | |
| oppB | Vp_N10_18_3430 | 2.402 | Oligopeptide ABC transporter permease OppB | |
| znuA | Vp_N10_18_4099 | 2.594 | Metal ABC transporter substrate-binding protein | |
| artI | Vp_N10_18_0733 | 3.101 | Arginine ABC transporter substrate-binding protein | |
| artM | Vp_N10_18_0735 | 3.204 | Arginine transporter permease subunit ArtM | |
| thiZ | Vp_N10_18_1090 | 3.271 | Hydrogenase expression protein | |
| thiX | Vp_N10_18_1091 | 3.567 | ABC transporter permease | |
| - | Vp_N10_18_1522 | 3.891 | Iron (III) ABC transporter ATP-binding protein | |
| artP | Vp_N10_18_0732 | 4.015 | Arginine ABC transporter ATP-binding protein ArtP | |
| znuB | Vp_N10_18_1681 | 6.403 | Zinc ABC transporter permease subunit ZnuB | |
| znuC | Vp_N10_18_1680 | 9.190 | Zinc ABC transporter ATP-binding protein ZnuC | |
| znuA | Vp_N10_18_1679 | 11.609 | Zinc ABC transporter substrate-binding protein ZnuA | |
| Propanoate metabolism | puuE | Vp_N10_18_2902 | 0.069 | Aspartate aminotransferase family protein |
| prpE | Vp_N10_18_0011 | 0.330 | AMP-binding protein | |
| acnD | Vp_N10_18_0013 | 0.370 | Fe/S-dependent 2-methylisocitrate dehydratase AcnD | |
| pdhB | Vp_N10_18_0742 | 0.382 | Alpha-ketoacid dehydrogenase subunit beta | |
| prpF | Vp_N10_18_0012 | 0.431 | 2-Methylaconitate cis-trans isomerase PrpF | |
| gabT | Vp_N10_18_0139 | 0.432 | 4-Aminobutyrate--2-oxoglutarate transaminase | |
| prpC | Vp_N10_18_0015 | 0.438 | 2-Methylcitrate synthase | |
| Benzoate degradation | pcaH | Vp_N10_18_0737 | 0.49 | Dioxygenase family protein |
| pcaC | Vp_N10_18_2971 | 2.003 | Carboxymuconolactone decarboxylase family protein | |
| Thiamine metabolism | thiC | Vp_N10_18_4412 | 2.116 | Phosphomethylpyrimidine synthase ThiC |
| thiE | Vp_N10_18_4413 | 2.247 | Thiamine phosphate synthase | |
| thiD | Vp_N10_18_1089 | 2.555 | Bifunctional hydroxymethylpyrimidine Kinase/phosphomethylpyrimidine kinase | |
| tenA | Vp_N10_18_1094 | 2.615 | Thiaminase II | |
| thiE | Vp_N10_18_1096 | 2.740 | Thiamine phosphate synthase | |
| Fat digestion and absorption | atoB | Vp_N10_18_3849 | 0.371 | 3-Ketoacyl-CoA thiolase @ Acetyl-CoA Acetyltransferase |
| atoB | Vp_N10_18_2988 | 4.215 | Thiolase family protein | |
| Quorum sensing | - | Vp_N10_18_2155 | 0.424 | ABC transporter ATP-binding protein |
| ribA | Vp_N10_18_1217 | 0.453 | GTP cyclohydrolase II | |
| - | Vp_N10_18_1879 | 0.472 | ABC transporter ATP-binding protein | |
| - | Vp_N10_18_2632 | 0.486 | Sigma 54-interacting transcriptional regulator | |
| - | Vp_N10_18_1219 | 2.140 | Sugar ABC transporter ATP-binding protein | |
| - | Vp_N10_18_0181 | 2.378 | Polyamine ABC transporter substrate-binding protein | |
| ribA | Vp_N10_18_2468 | 2.675 | GTP cyclohydrolase II | |
| - | Vp_N10_18_1876 | 2.918 | ABC transporter permease | |
| - | Vp_N10_18_2783 | 9.727 | ABC transporter permease | |
| Pathogenic Escherichia coli infection | gapA | Vp_N10_18_3876 | 2.086 | Glyceraldehyde-3-phosphate dehydrogenase |
| yscF | Vp_N10_18_0060 | 5.836 | Type III secretion system needle filament protein VscF |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, P.; Yang, L.; Wang, J.; Su, C.; Qin, S.; Zeng, C.; Chen, L. Genomic and Transcriptomic Analysis Reveal Multiple Strategies for the Cadmium Tolerance in Vibrio parahaemolyticus N10-18 Isolated from Aquatic Animal Ostrea gigas Thunberg. Foods 2022, 11, 3777. https://doi.org/10.3390/foods11233777
Yu P, Yang L, Wang J, Su C, Qin S, Zeng C, Chen L. Genomic and Transcriptomic Analysis Reveal Multiple Strategies for the Cadmium Tolerance in Vibrio parahaemolyticus N10-18 Isolated from Aquatic Animal Ostrea gigas Thunberg. Foods. 2022; 11(23):3777. https://doi.org/10.3390/foods11233777
Chicago/Turabian StyleYu, Pan, Lianzhi Yang, Juanjuan Wang, Chenli Su, Si Qin, Chaoxi Zeng, and Lanming Chen. 2022. "Genomic and Transcriptomic Analysis Reveal Multiple Strategies for the Cadmium Tolerance in Vibrio parahaemolyticus N10-18 Isolated from Aquatic Animal Ostrea gigas Thunberg" Foods 11, no. 23: 3777. https://doi.org/10.3390/foods11233777