
Pea-Tea Intercropping Improves Tea Quality through Regulating Amino Acid Metabolism and Flavonoid Biosynthesis
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. RNA Extraction and cDNA Library Construction
2.3. Transcriptome Sequencing and Data Assessment
2.4. Differentially Expressed Genes Identification and Function Annotation
2.5. Quantitative Real-Time PCR Verification
2.6. Metabolite Extraction and Metabolome Analysis
2.7. Detection of Tea Quality Components
2.8. Statistical Analysis
3. Results
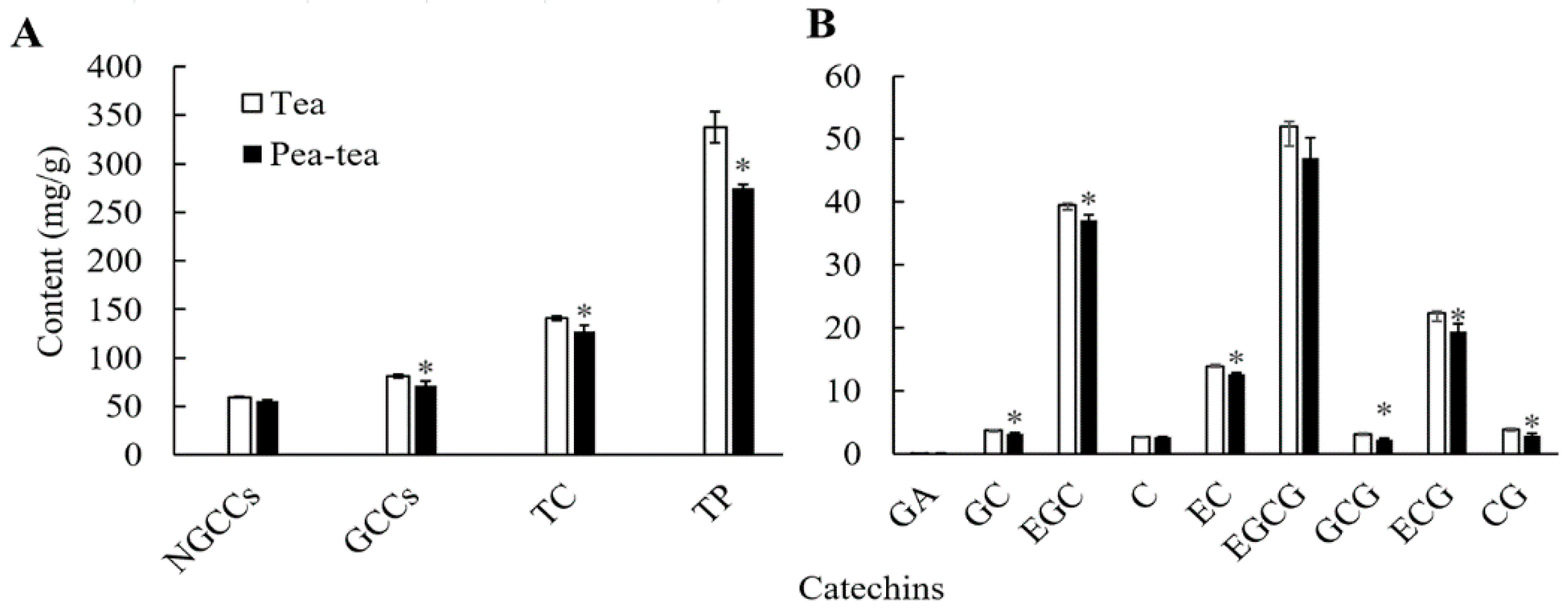
3.1. Tea Quality Components in Pea-Tea Intercropping Plants
3.2. Quality of Transcriptomic Data
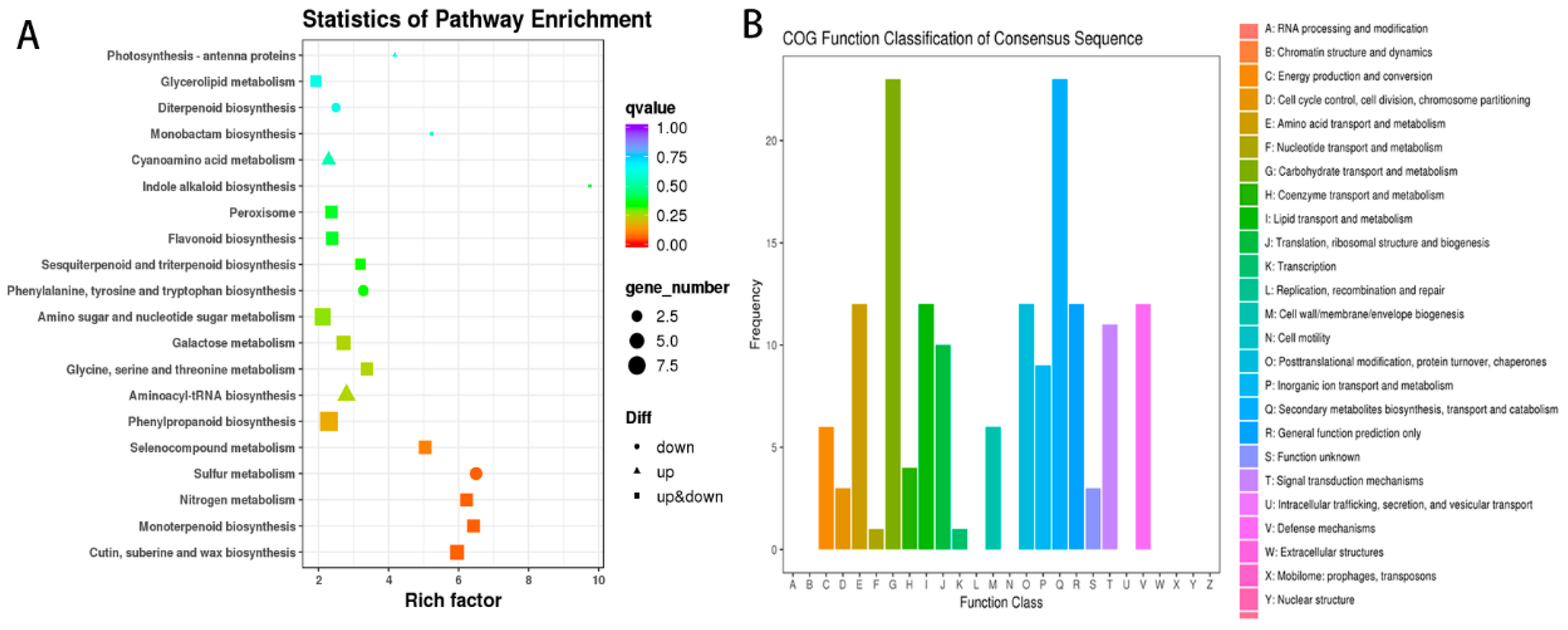
3.3. Identification of Differentially Expressed Genes
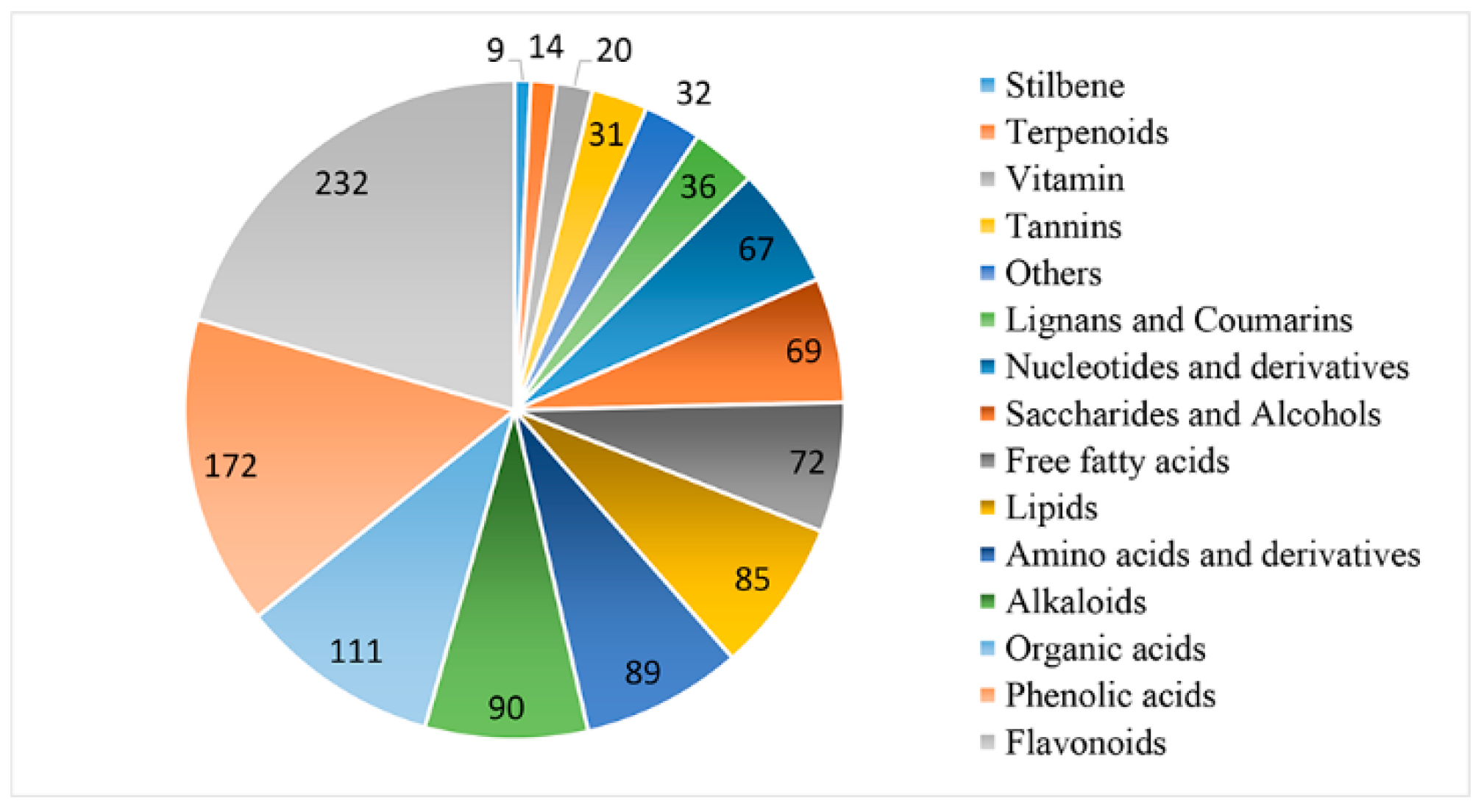
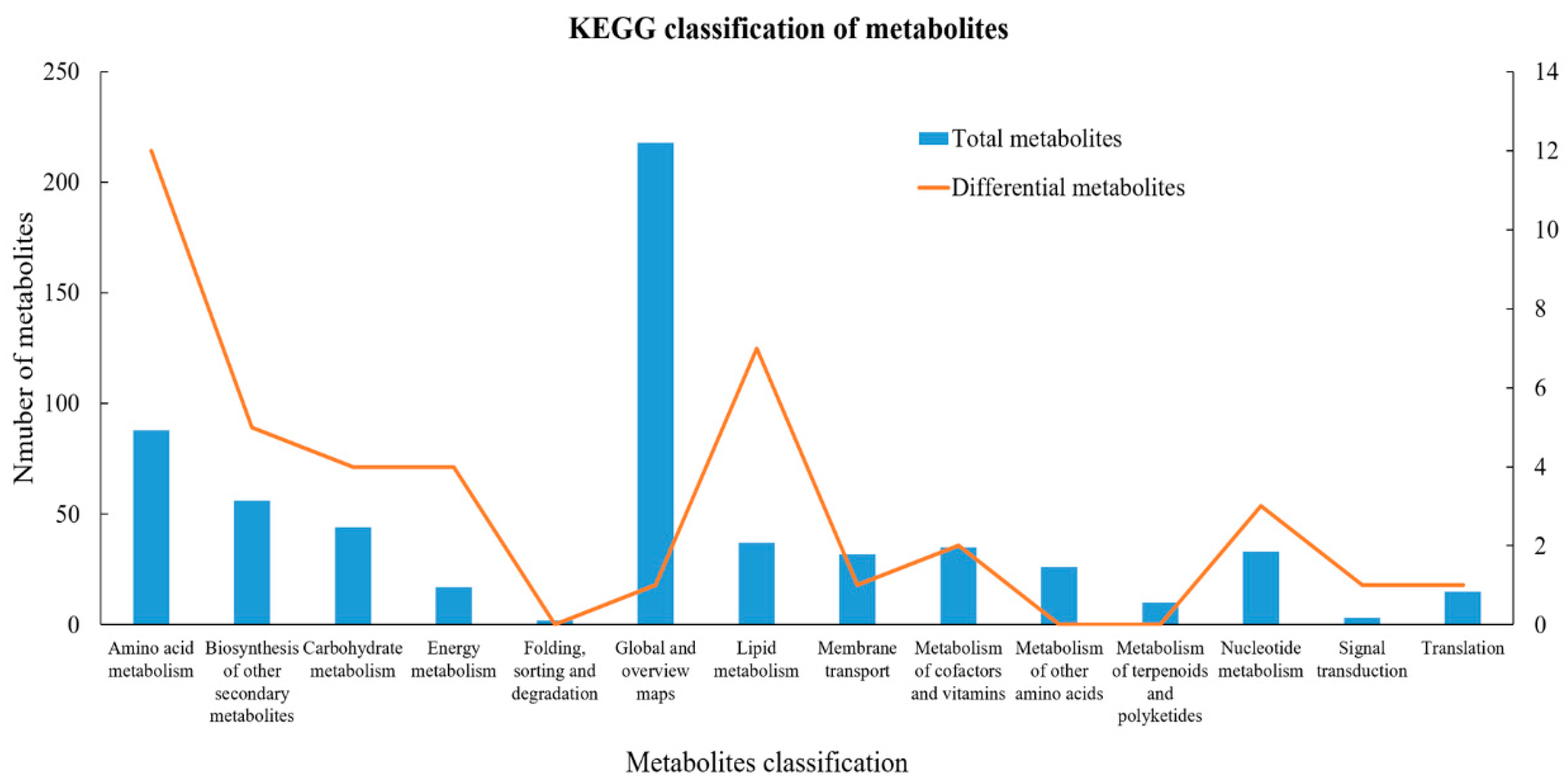
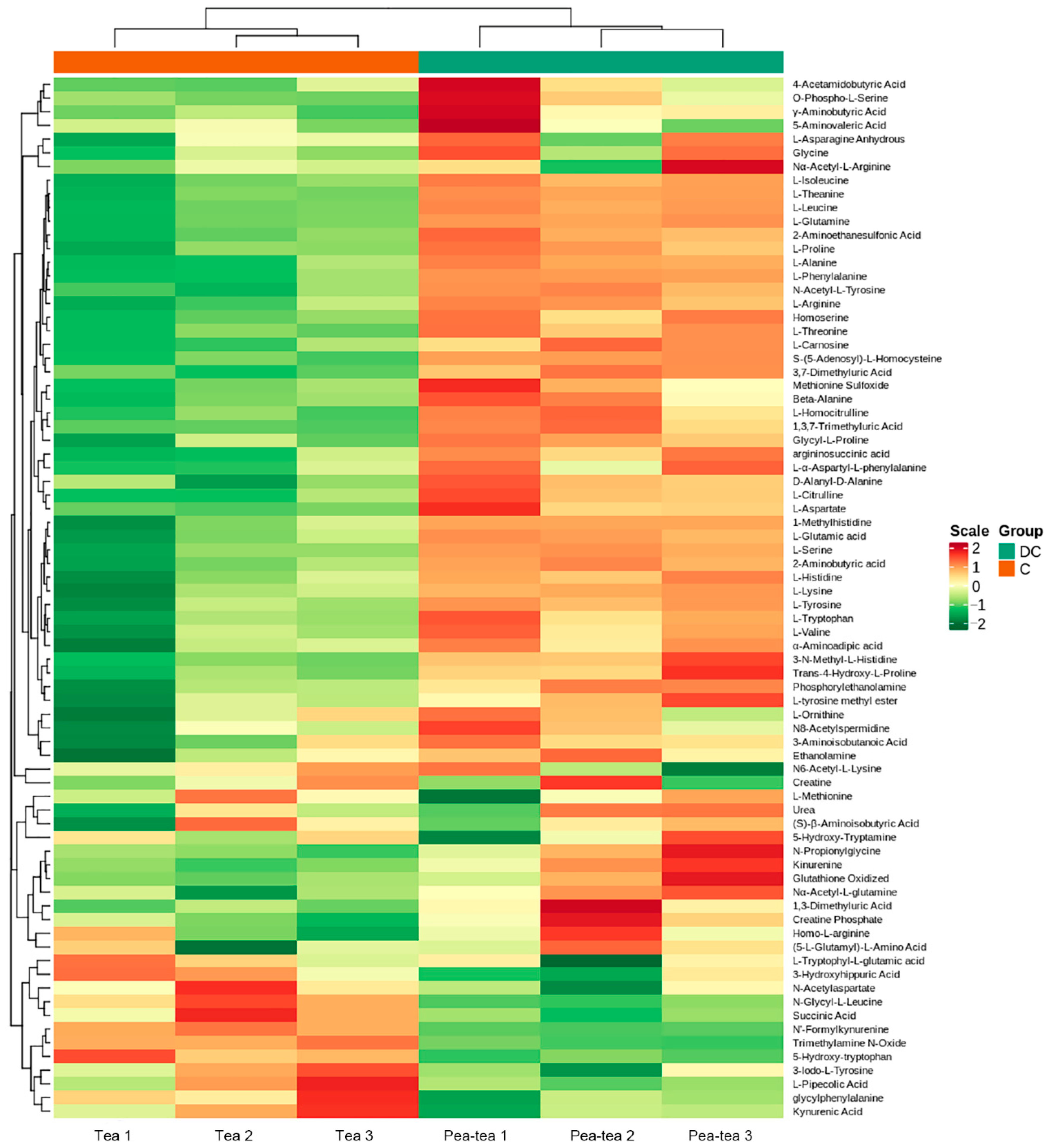
3.4. Differential Metabolite Identification
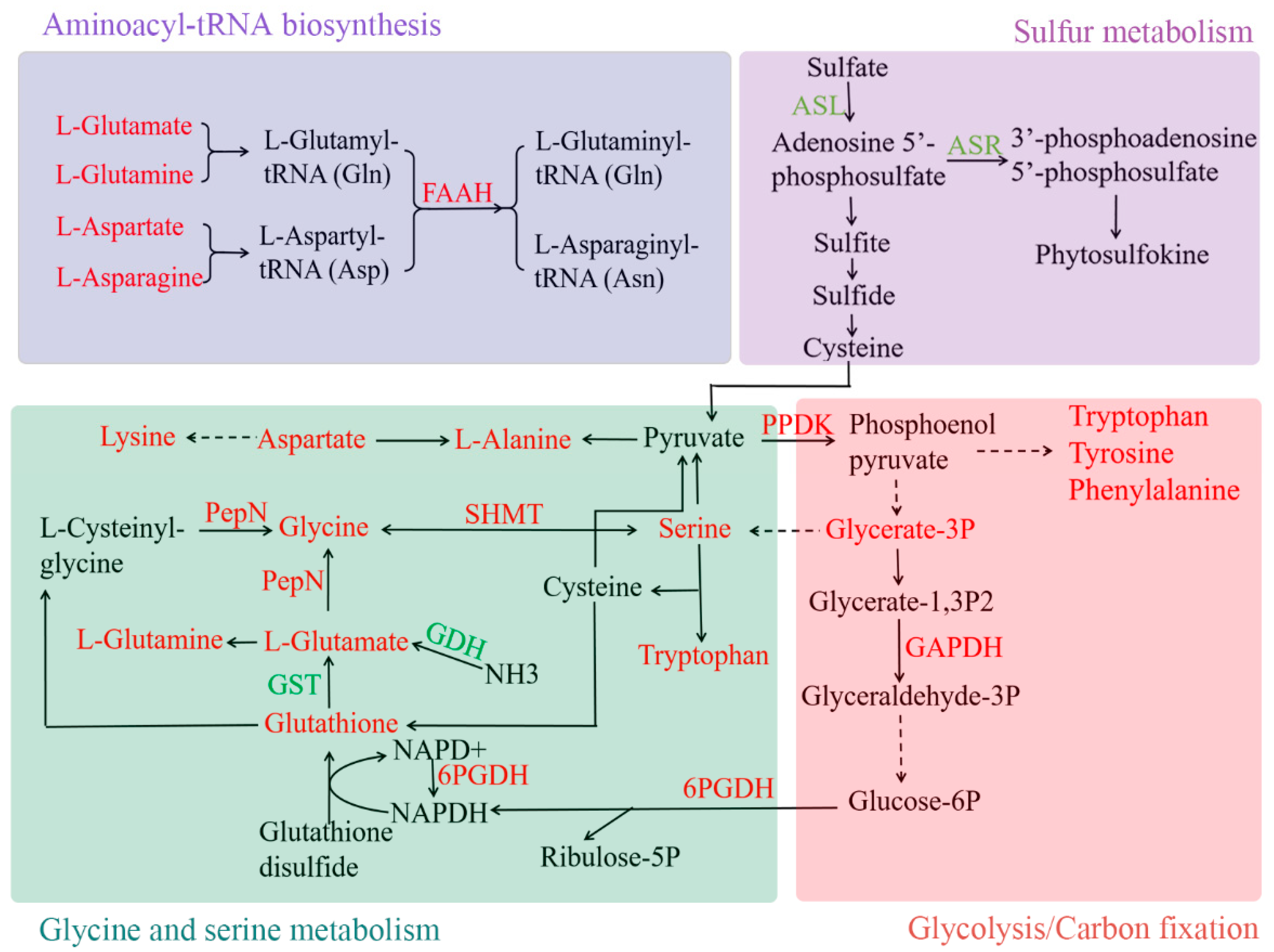
3.5. Amino Acid Metabolism in Pea Intercropped Tea Plants
3.6. Carbohydrate Transport and Metabolism in Pea Intercropped Tea Plants
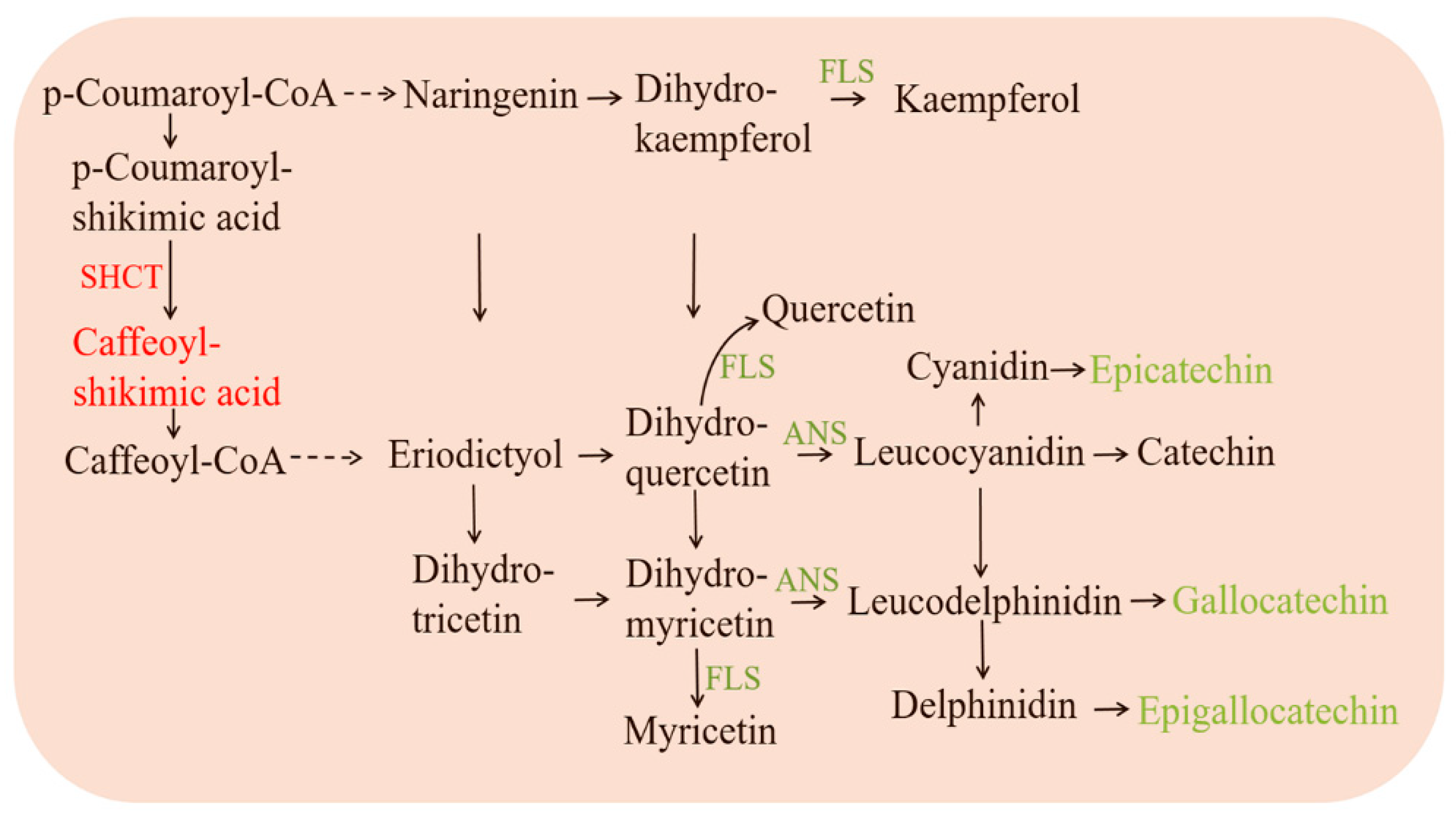
3.7. Flavonoid Metabolism in Pea Intercropped Tea Plants
4. Discussion
4.1. Pea Intercropping Promoted the Accumulation of Amino Acids in Tea Plants
4.2. Pea Intercropping Slowed down the Catechin Biosynthesis in Tea Plants
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wu, T.; Zou, R.; Pu, D.; Lan, Z.; Zhao, B. Non-targeted and targeted metabolomics profiling of tea plants (Camellia sinensis) in response to its intercropping with Chinese chestnut. BMC Plant Biol. 2021, 21, 55. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Qin, Y.; Li, M. Intercropping of tea (Camellia sinensis L.) and Chinese chestnut: Variation in the structure of rhizosphere bacterial communities. J. Soil Sci. Plant Nutr. 2021, 21, 2178–2190. [Google Scholar] [CrossRef]
- Guo, Z.; Zhang, Y.; Deegen, P.; Uibrig, H. Economic analyses of rubber and tea plantations and rubber-tea intercropping in Hainan, China. Agrofor. Sys. 2006, 66, 117–127. [Google Scholar] [CrossRef]
- Bai, Y.C.; Li, B.X.; Xu, C.Y.; Raza, M.; Wang, Q.; Wang, Q.Z.; Fu, Y.N.; Hu, J.Y.; Imoulan, A.; Hussain, M.; et al. Intercropping walnut and tea: Effects on soil nutrients, enzyme activity, and microbial communities. Front. Microbiol. 2022, 13, 852342. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Liu, Z.; Wang, C.; Zhong, Z. Effects of intercropping with persimmon on the rhizosphere environment of tea. Front. Biol. China 2006, 1, 407–410. [Google Scholar] [CrossRef]
- Ma, Y.-H.; Fu, S.-L.; Zhang, X.-P.; Zhao, K.; Chen, H.Y.H. Intercropping improves soil nutrient availability, soil enzyme activity and tea quantity and quality. App. Soil Ecol. 2017, 119, 171–178. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, C.; Xu, Y.; Huang, X.; Zhang, L.; Mu, W. Effects of intercropping tea with aromatic plants on population dynamics of arthropods in Chinese tea plantations. J. Pest Sci. 2017, 90, 227–237. [Google Scholar] [CrossRef]
- Zou, Y.; Shen, F.; Zhong, Y.; Lv, C.; Pokharel, S.S.; Fang, W.; Chen, F. Impacts of Intercropped Maize Ecological Shading on Tea Foliar and Functional Components, Insect Pest Diversity and Soil Microbes. Plants 2022, 11, 1883. [Google Scholar] [CrossRef]
- Duan, Y.; Shang, X.; Liu, G.; Zou, Z.; Zhu, X.; Ma, Y.; Li, F.; Fang, W. The effects of tea plants-soybean intercropping on the secondary metabolites of tea plants by metabolomics analysis. BMC Plant Biol. 2021, 21, 482. [Google Scholar] [CrossRef]
- Duan, Y.; Shen, J.; Zhang, X.; Wen, B.; Ma, Y.; Wang, Y.; Fang, W.; Zhu, X. Effects of soybean–tea intercropping on soil-available nutrients and tea quality. Acta Physiol. Plant. 2019, 41, 140. [Google Scholar] [CrossRef]
- Huang, Z.; Cui, C.; Cao, Y.; Dai, J.; Cheng, X.; Hua, S.; Wang, W.; Duan, Y.; Petropoulos, E.; Wang, H.; et al. Tea plant–legume intercropping simultaneously improves soil fertility and tea quality by changing Bacillus species composition. Hort. Res. 2022, 9, uhac046. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Dong, X.; Wang, Y.; Maker, G.; Agarwal, M.; Ding, Z. Tea-soybean intercropping improves tea quality and nutrition uptake by inducing changes of rhizosphere bacterial communities. Microorganisms 2022, 10, 2149. [Google Scholar] [CrossRef] [PubMed]
- Farooq, T.H.; Kumar, U.; Mo, J.; Shakoor, A.; Wang, J.; Rashid, M.H.U.; Tufail, M.A.; Chen, X.; Yan, W. Intercropping of peanut–tea enhances soil enzymatic activity and soil nutrient status at different soil profiles in subtropical southern China. Plants 2021, 10, 881. [Google Scholar] [CrossRef] [PubMed]
- Xia, E.; Tong, W.; Hou, Y.; An, Y.; Chen, L.; Wu, Q.; Liu, Y.; Yu, J.; Li, F.; Li, R.; et al. The Reference Genome of Tea Plant and Resequencing of 81 Diverse Accessions Provide Insights into Its Genome Evolution and Adaptation. Mol. Plant 2020, 13, 1013–1026. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotech. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Florea, L.; Song, L.; Salzberg, S.L. Thousands of exon skipping events differentiate among splicing patterns in sixteen human tissues. F1000 Res. 2013, 2, 188. [Google Scholar] [CrossRef] [Green Version]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D.J.M. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ma, Q.; Li, H.; Zou, Z.; Arkorful, E.; Lv, Q.; Zhou, Q.; Chen, X.; Sun, K.; Li, X. Transcriptomic analyses identify albino-associated genes of a novel albino tea germplasm ‘Huabai 1’. Hort. Res. 2018, 5, 54. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef]
- Guiducci, G.; Paone, A.; Tramonti, A.; Giardina, G.; Rinaldo, S.; Bouzidi, A.; Magnifico, M.C.; Marani, M.; Menendez, J.A.; Fatica, A. The moonlighting RNA-binding activity of cytosolic serine hydroxymethyltransferase contributes to control compartmentalization of serine metabolism. Nucleic Acids Res. 2019, 47, 4240–4254. [Google Scholar] [CrossRef] [PubMed]
- Costenoble, R.; Picotti, P.; Reiter, L.; Stallmach, R.; Heinemann, M.; Sauer, U.; Aebersold, R. Comprehensive quantitative analysis of central carbon and amino-acid metabolism in Saccharomyces cerevisiae under multiple conditions by targeted proteomics. Mol. Syst. Biol. 2011, 7, 464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueroa, C.M.; Feil, R.; Ishihara, H.; Watanabe, M.; Kölling, K.; Krause, U.; Höhne, M.; Encke, B.; Plaxton, W.C.; Zeeman, S.C. Trehalose 6–phosphate coordinates organic and amino acid metabolism with carbon availability. Plant J. 2016, 85, 410–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz-Bertomeu, J.S.; Cascales-Miñana, B.; Mulet, J.M.; Baroja-Fernández, E.; Pozueta-Romero, J.; Kuhn, J.M.; Segura, J.; Ros, R. Plastidial glyceraldehyde-3-phosphate dehydrogenase deficiency leads to altered root development and affects the sugar and amino acid balance in Arabidopsis. Plant Physiol. 2009, 151, 541–558. [Google Scholar] [CrossRef] [Green Version]
- Pratelli, R.; Pilot, G. Regulation of amino acid metabolic enzymes and transporters in plants. J. Exp. Bot. 2014, 65, 5535–5556. [Google Scholar] [CrossRef]
- Sharkey, T.D. Pentose phosphate pathway reactions in photosynthesizing cells. Cells 2021, 10, 1547. [Google Scholar] [CrossRef]
- Kopriva, S.; Rennenberg, H. Control of sulphate assimilation and glutathione synthesis: Interaction with N and C metabolism. J. Exp. Bot. 2004, 55, 1831–1842. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Chaouch, S.; Han, Y.; Neukermans, J.; Marquez-Garcia, B.; Queval, G.; Foyer, C.H. Glutathione in plants: An integrated overview. Plant Cell Environ. 2012, 35, 454–484. [Google Scholar] [CrossRef]
- Sylvestre-Gonon, E.; Law, S.R.; Schwartz, M.; Robe, K.; Keech, O.; Didierjean, C.; Dubos, C.; Rouhier, N.; Hecker, A. Functional, structural and biochemical features of plant serinyl-glutathione transferases. Front. Plant Sci. 2019, 10, 608. [Google Scholar] [CrossRef] [Green Version]
- Kopriva, S.; Malagoli, M.; Takahashi, H. Sulfur nutrition: Impacts on plant development, metabolism, and stress responses. J. Exp. Bot. 2019, 70, 4069–4073. [Google Scholar] [CrossRef]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M.J.P.S. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Tu, S.; Xiao, F.; Min, X.; Chen, H.; Fan, X.; Cao, K. Catechin attenuates coronary heart disease in a rat model by inhibiting inflammation. Cardiovasc. Toxicol. 2018, 18, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Shim, W.; Kim, C.E.; Choi, S.Y.; Lee, H.; Yang, J. Therapeutic efficacy of nanocomplex of poly(ethylene glycol) and catechin for dry eye disease in a mouse model. Investig. Ophthalmol. Vis. Sci. 2017, 58, 1682–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pervin, M.; Unno, K.; Ohishi, T.; Tanabe, H.; Miyoshi, N.; Nakamura, Y. Beneficial effects of green tea catechins on neurodegenerative diseases. Molecules 2018, 23, 1297. [Google Scholar] [CrossRef] [Green Version]
- Potapovich, A.I.; Kostyuk, V.A. Comparative study of antioxidant properties and cytoprotective activity of flavonoids. Biochemistry 2003, 68, 514–519. [Google Scholar] [CrossRef]
- Zhang, L.; Cao, Q.-Q.; Granato, D.; Xu, Y.-Q.; Ho, C.-T. Association between chemistry and taste of tea: A review. Trends. Food Sci. Technol. 2020, 101, 139–149. [Google Scholar] [CrossRef]
- Ho, C.-T.; Zheng, X.; Li, S. Tea aroma formation. Food Sci. Hum. Wellness 2015, 4, 9–27. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Reads | Mapped Reads | Unique Mapped Reads | GC Content | % ≥ Q30 |
|---|---|---|---|---|---|
| Tea1 | 47,836,620 | 42,016,678 (87.83%) | 35,436,061 (74.08%) | 45.38% | 95.49% |
| Tea2 | 53,223,798 | 46,833,314 (87.99%) | 39,326,279 (73.89%) | 46.04% | 95.21% |
| Tea3 | 55,349,774 | 48,631,380 (87.86%) | 40,979,079 (74.04%) | 45.80% | 95.18% |
| Pea-tea1 | 51,540,604 | 45,241,568 (87.78%) | 38,288,792 (74.29%) | 45.69% | 94.72% |
| Pea-tea2 | 58,111,972 | 50,998,587 (87.76%) | 42,914,459 (73.85%) | 45.77% | 94.89% |
| Pea-tea3 | 55,017,918 | 48,264,631 (87.73%) | 40,734,571 (74.04%) | 45.35% | 95.23% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Q.; Song, L.; Niu, Z.; Qiu, Z.; Sun, H.; Ren, Z.; Wu, H.; Wang, Y.; Mei, H.; Li, X.; et al. Pea-Tea Intercropping Improves Tea Quality through Regulating Amino Acid Metabolism and Flavonoid Biosynthesis. Foods 2022, 11, 3746. https://doi.org/10.3390/foods11223746
Ma Q, Song L, Niu Z, Qiu Z, Sun H, Ren Z, Wu H, Wang Y, Mei H, Li X, et al. Pea-Tea Intercropping Improves Tea Quality through Regulating Amino Acid Metabolism and Flavonoid Biosynthesis. Foods. 2022; 11(22):3746. https://doi.org/10.3390/foods11223746
Chicago/Turabian StyleMa, Qingping, Laichao Song, Zhanhai Niu, Ziyuan Qiu, Haiwei Sun, Zhihong Ren, Huanhuan Wu, Yu Wang, Huiling Mei, Xinghui Li, and et al. 2022. "Pea-Tea Intercropping Improves Tea Quality through Regulating Amino Acid Metabolism and Flavonoid Biosynthesis" Foods 11, no. 22: 3746. https://doi.org/10.3390/foods11223746