
Dietary Supplement of Amomum villosum Lour. Polysaccharide Attenuates Ulcerative Colitis in BALB/c Mice
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Extraction of AVLP
2.3. Monosaccharide Composition Analysis
2.4. Molecular Weight Analysis
2.5. FT-IR Spectrometry
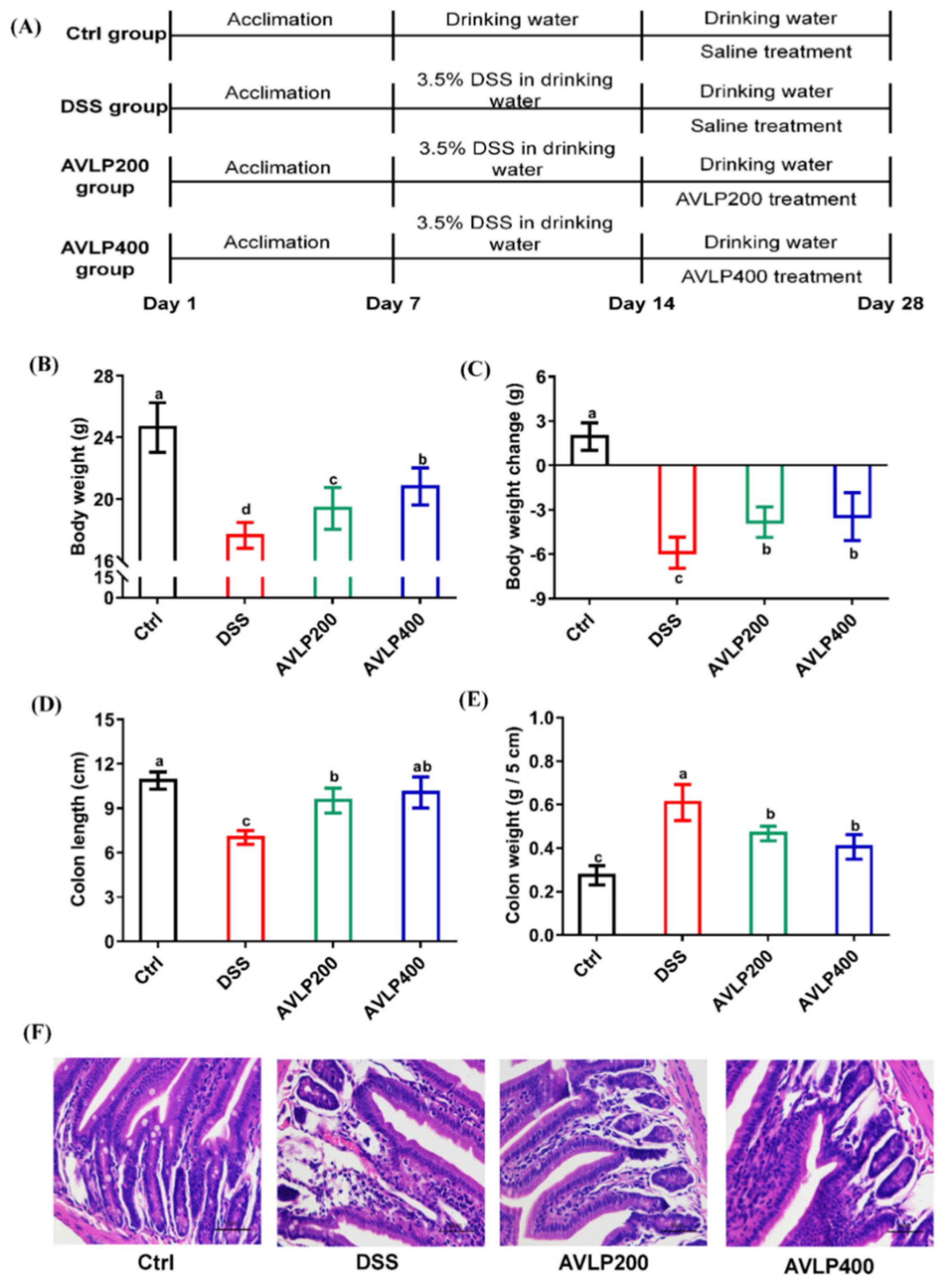
2.6. Animal Experiment
2.7. Serum Parameter Analysis
2.8. Histopathology Analysis
2.9. Immunohistochemistry Analysis
(percentage of strong intensity × 3)
2.10. Quantitative Reverse Transcription PCR Analysis
2.11. Intestinal Flora Analysis
2.12. Statistical Analysis
3. Results and Discussion
3.1. Characterization of AVLP
3.2. Effects of AVLP on Pathological Changes in Colitis Mice
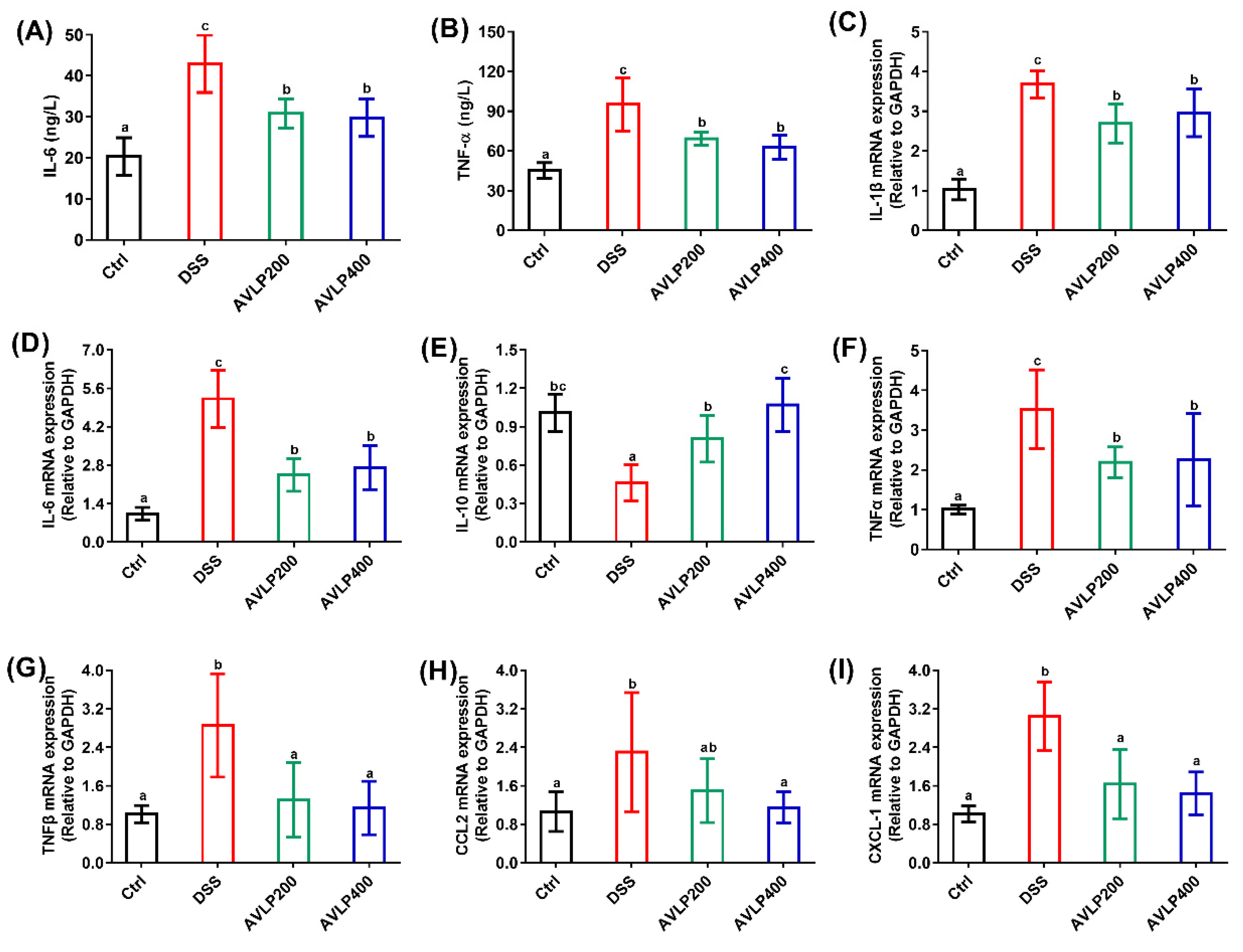
3.3. Effects of AVLP on Inflammatory Cytokines and Chemokines in Colitis Mice
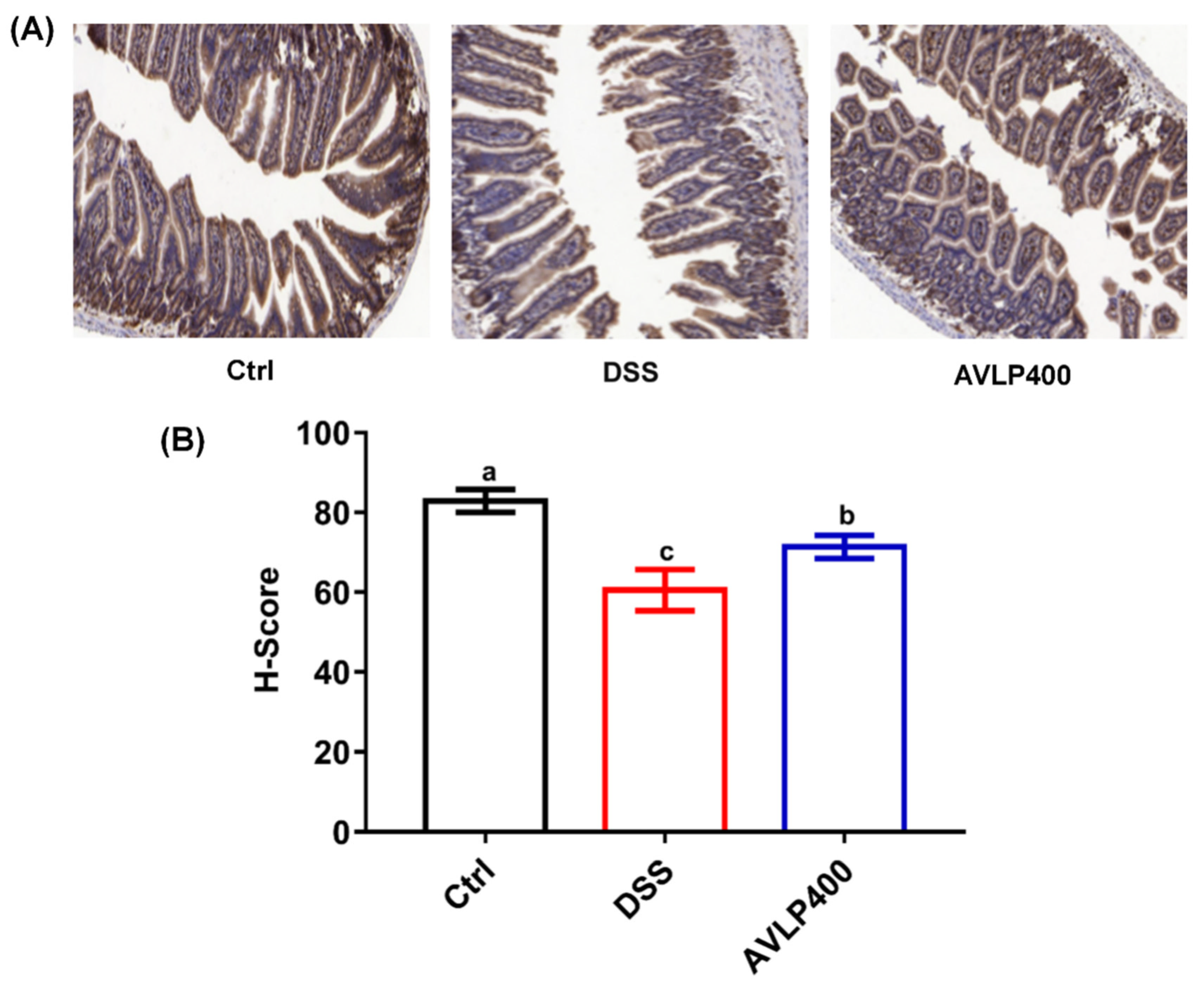
3.4. Effect of AVLP on ZO-1 Protein Expression in Colitis Mice
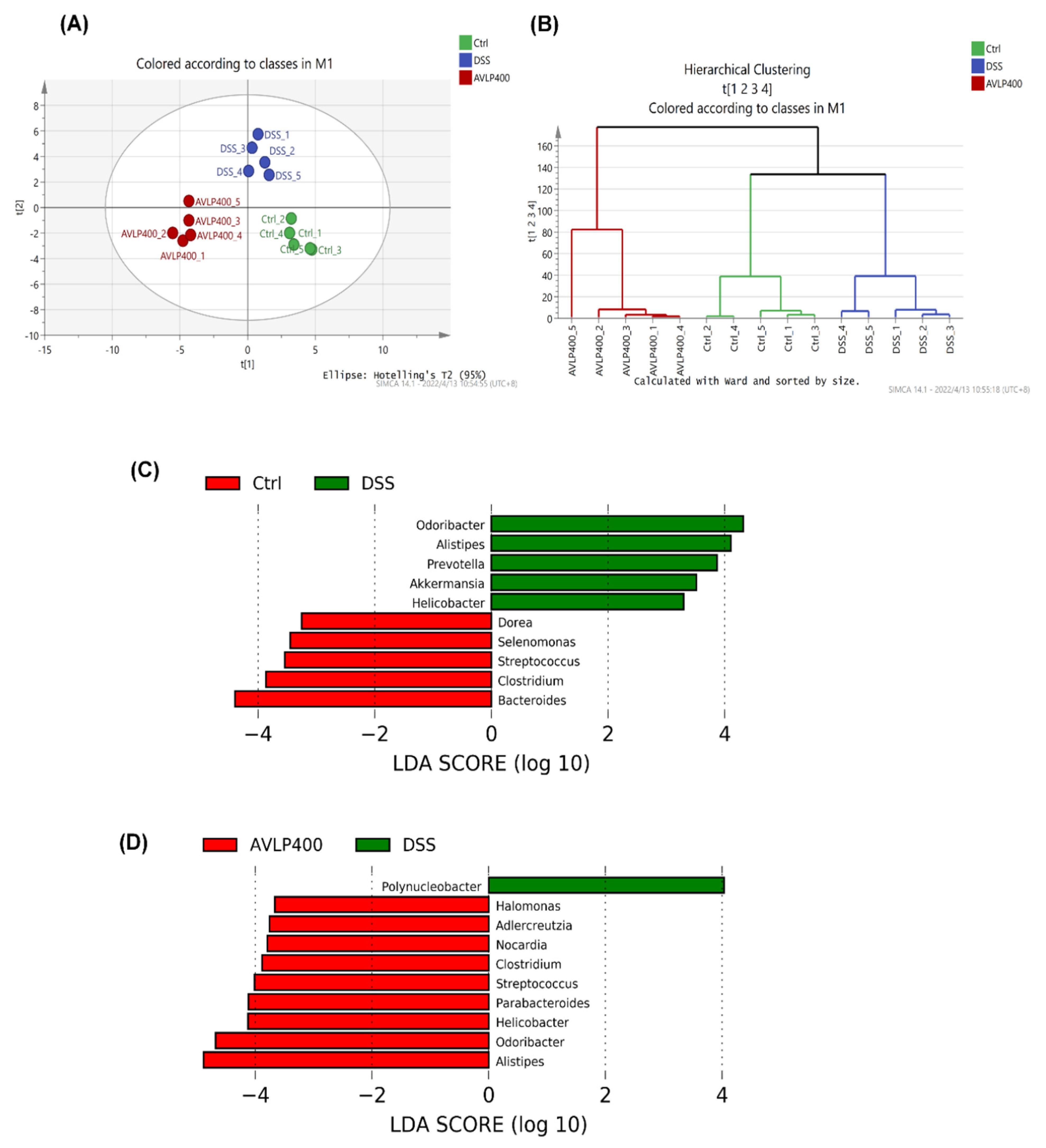
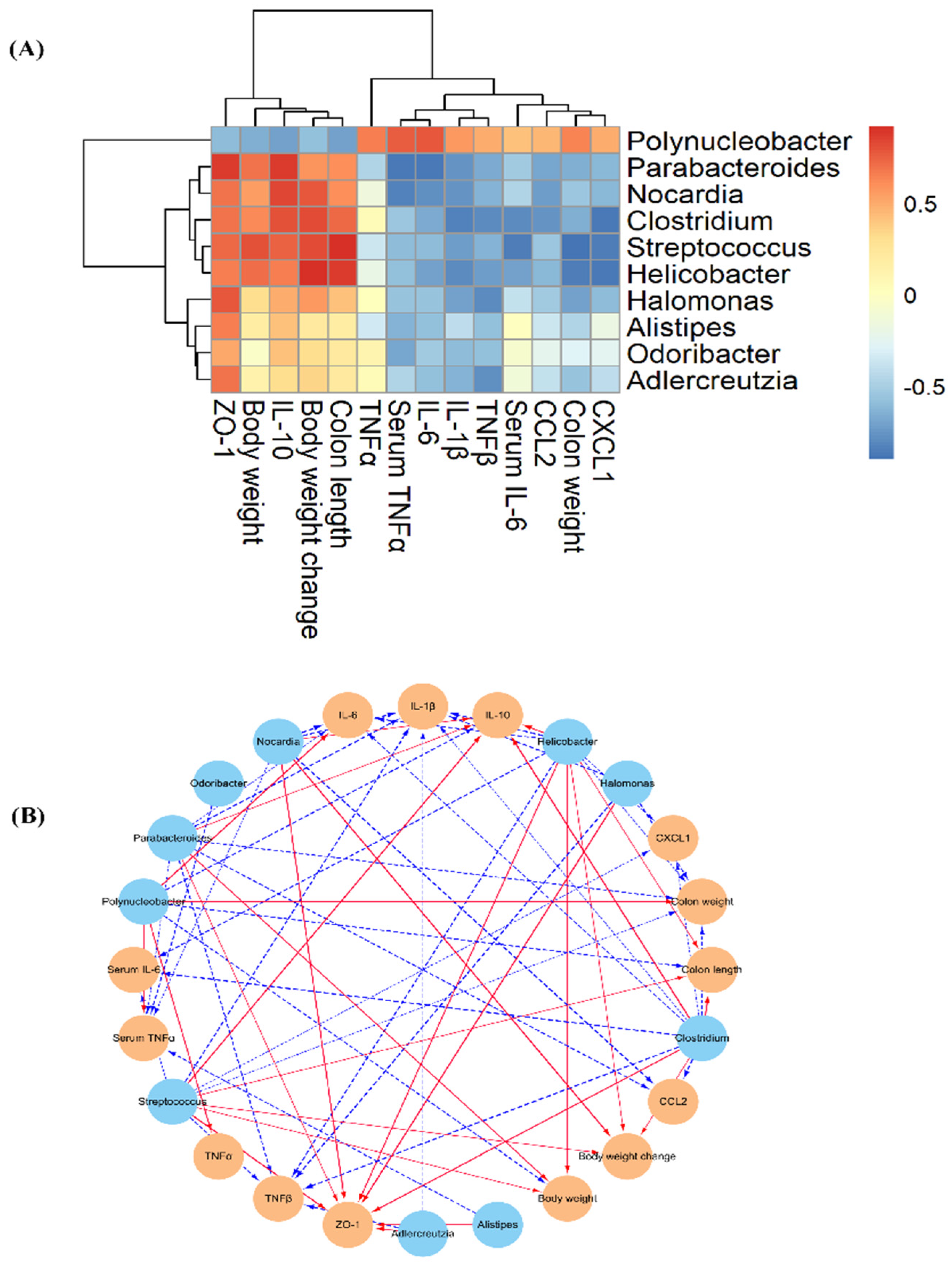
3.5. Effect of AVLP on Gut Flora in Colitis Mice
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Marina, L.; Verna, G.; Cavalcanti, E.; Santis, S.D.; Armentano, R.; Tafaro, A.; Lippolis, A.; Campiglia, P.; Gasbarrini, A.; Mastronardi, M.; et al. Interleukin 1beta Blockade Reduces Intestinal Inflammation in a Murine Model of Tumor Necrosis Factor-Independent Ulcerative Colitis. Cell Mol. Gastroenterol. Hepatol. 2022, 14, 151–171. [Google Scholar] [CrossRef]
- Cui, G.; Yuan, A. A Systematic Review of Epidemiology and Risk Factors Associated with Chinese Inflammatory Bowel Disease. Front. Med. 2018, 5, 183. [Google Scholar] [CrossRef] [PubMed]
- Feuerstein, J.-D.; Isaacs, K.-L.; Schneider, Y.; Siddique, S.-M.; Falck, Y.-Y.; Singh, S. AGA Clinical Practice Guidelines on the Management of Moderate to Severe Ulcerative Colitis. Gastroenterology 2020, 158, 1450–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyams, J.S.; Turner, D.; Cohen, S.A.; Szakos, E.; Kowalskaduplaga, K.; Ruemmele, F.; Croft, N.M.; Korczowski, B.; Lawrence, P.; Bhatia, S.; et al. Pharmacokinetics, Safety, and Efficacy of Intravenous Vedolizumab in Paediatric Patients with Ulcerative Colitis or Crohn’s Disease: Results from the Phase 2 HUBBLE Study. J. Crohn’s Colitis 2022, 16, 1243–1254. [Google Scholar] [CrossRef] [PubMed]
- Peery, A.-F.; Crockett, S.-D.; Murphy, C.-C.; Jensen, E.-T.; Kim, H.-P.; Egberg, M.-D.; Lund, J.-L.; Moon, A.-M.; Pate, V.; Barnes, E.-L.; et al. Burden and Cost of Gastrointestinal, Liver, and Pancreatic Diseases in the United States: Update 2021. Gastroenterology 2021, 162, 621–644. [Google Scholar] [CrossRef]
- Niu, W.; Chen, W.; Xu, R.; Dong, H.; Yang, F.; Wang, Y.; Zhang, Z.; Ju, J. Polysaccharides from natural resources exhibit great potential in the treatment of ulcerative colitis: A review. Carbohydr. Polym. 2021, 254, 117189. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, J.; Li, J.; Zhu, J.; Wang, R.; Xi, Q.; Wu, H.; Shi, T.; Chen, W. Astragalus polysaccharide prevents ferroptosis in a murine model of experimental colitis and human Caco-2 cells via inhibiting NRF2/HO-1 pathway. Eur. J. Pharmacol. 2021, 911, 174518. [Google Scholar] [CrossRef]
- Han, H.-S.; Kim, S.-Y.; Shin, J.-S.; Lee, H.-H.; Chung, K.-S.; Rhee, Y.K.; Cho, C.-W.; Hong, H.-D.; Lee, K.-T. Polysaccharide fraction isolated from the leaves of Hordeum vulgare L. protects against colonic inflammation of systemic immune responses. J. Funct. Foods 2021, 87, 104765. [Google Scholar] [CrossRef]
- Yu, J.; Dong, X.-D.; Jiao, J.-S.; Ji, H.-Y.; Liu, A.-J. Antitumor and immunoregulatory activities of a novel polysaccharide from Astragalus membranaceus on S180 tumor-bearing mice. Int. J. Biol. Macromol. 2021, 189, 930–938. [Google Scholar] [CrossRef]
- Miao, X.-P.; Sun, X.-N.; Cui, L.-J.; Cao, Q.-F.; Zhuang, G.-F.; Deng, T.-Z.; Zhang, D.-Y. Suppressive effect of pectic polysaccharides extracted from Rauwolfia verticillata (Lour.) Baill. var. hainanensis Tsiang on inflammation by regulation of NF– κ B pathway and interleukin–17 in mice with dextran sulphatesodium–induced ulcerative colitis. Asian Pac. J. Trop. Med. 2015, 8, 147–152. [Google Scholar] [CrossRef]
- Carlson, J.; Gould, T.; Slavin, J. In vitro analysis of partially hydrolyzed guar gum fermentation on identified gut microbiota. Anaerobe 2016, 42, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Chen, S.; Chen, J.; Lin, J.; Xiong, Q.; Yang, Y.; Yuan, J.; Zhou, L.; He, L.; Hou, S.; et al. Therapeutic roles of polysaccharides from Dendrobium Officinaleon colitis and its underlying mechanisms. Carbohydr. Polym. 2018, 185, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Bie, Y.; Wang, D.; Fan, W.-T. Effects of Dandelion polysaccharide on IL-6 / STAT3 signaling pathway in ulcerative colitis rats. China Acad. J. Electron. Publ. House 2017, 33, 422–425. (In Chinese) [Google Scholar] [CrossRef]
- Commission, Chinese Pharmacopoeia. Pharmacopoeia of the People’s Republic of China; China Medical Science Press: Beijing, China, 2015. (In Chinese) [Google Scholar]
- Yue, J.; Zhang, S.; Zheng, B.; Faisal, R.; Luo, Z.; Li, X.; Zhang, Y.; Nie, Q.; Qiu, M. Efficacy and Mechanism of Active Fractions in Fruit of Amomum villosum Lour. for Gastric Cancer. J. Cancer 2021, 12, 5991–5998. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Ni, W.; Yang, C.; Zhang, T.; Lu, S.; Zhao, R.; Mao, X.; Yu, J. Therapeutic Effect of Amomum villosum on Inflammatory Bowel Disease in Rats. Front. Pharmacol. 2018, 9, 639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.-J.; Weng, W.-F.; Zhao, H.-N.; Wen, J.-F.; Li, R.; Li, J.-N.; Zeng, C.-B.; Ji, S.-G. Application of Fourier transform near-infrared spectroscopy combined with GC in rapid and simultaneous determination of essential components in Amomum villosum. Spectrochim. Acta Mol. Biomol. Spectrosc. 2021, 251, 119426. [Google Scholar] [CrossRef] [PubMed]
- Jia, R.-B.; Wu, J.; Li, Z.-R.; Ou, Z.-R.; Lin, L.; Sun, B.; Zhao, M. Structural characterization of polysaccharides from three seaweed species and their hypoglycemic and hypolipidemic activities in type 2 diabetic rats. Int. J. Biol. Macromol. 2020, 155, 1040–1049. [Google Scholar] [CrossRef]
- Jia, R.-B.; Li, Z.-R.; Wu, J.; Ou, Z.-R.; Liao, B.; Sun, B.; Lin, L.; Zhao, M. Mitigation mechanisms of Hizikia fusifarme polysaccharide consumption on type 2 diabetes in rats. Int. J. Biol. Macromol. 2020, 164, 2659–2670. [Google Scholar] [CrossRef]
- Fatemeh, N.B.; Tahmasebi, E.S. FT-IR study of the polysaccharides isolated from the skin juice, gel juice, and flower of Aloe vera tissues affected by fertilizer treatment. Org. Med. Chem. Lett. 2012, 2, 33. [Google Scholar] [CrossRef] [Green Version]
- Kalkan, I.H.; Dagli, U. What is the most accurate method for the diagnosis of cytomegalovirus (CMV) enteritis or colitis? Turk. J. Gastroenterol. 2010, 21, 83–86. [Google Scholar] [CrossRef]
- Obermeier, F.; Kojouharoff, G.; Hans, W.; SchÖlmerich, J.; Gross, V.; Falk, W. Interferon-gamma (IFN-γ)- and tumour necrosis factor (TNF)-induced nitric oxide as toxic effector molecule in chronic dextran sulphate sodium (DSS)-induced colitis in mice. Clin. Exp. Immunol. 2001, 116, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Todenhöfer, T.; Gratzke, C. Re: Prostate-specific Membrane Antigen Heterogeneity and DNA Repair Defects in Prostate Cancer. Eur. Urol. 2020, 77, 559–560. [Google Scholar] [CrossRef] [PubMed]
- Robin, G.; Berry, L.-D.; Aisner, D.L.; Sheren, J.; Boyle, T.; Bunn, P.A.; Johnson, B.E.; David, J.K.; Drilon, A.; Sholl, L.M.; et al. MET IHC Is a Poor Screen for MET Amplification or MET Exon 14 Mutations in Lung Adenocarcinomas: Data from a Tri-Institutional Cohort of the Lung Cancer Mutation Consortium. J. Thorac. Oncol. 2019, 14, 1666–1671. [Google Scholar] [CrossRef]
- Sharma, A.; Tirpude, N.V.; Kulurkar, P.M.; Sharma, R.; Padwad, Y. Berberis lycium fruit extract attenuates oxi-inflammatory stress and promotes mucosal healing by mitigating NF-kappaB/c-Jun/MAPKs signalling and augmenting splenic Treg proliferation in a murine model of dextran sulphate sodium-induced ulcerative colitis. Eur. J. Nutr. 2020, 59, 2663–2681. [Google Scholar] [CrossRef]
- Suo, S.; Lai, Y.; Li, M.; Song, Q.; Cai, J.; Zhao, J.; Yang, Q.; Ung, C.O.L.; Hu, H. Phytochemicals, pharmacology, clinical application, patents, and products of Amomi fructus. Food Chem. Toxicol. 2018, 119, 31–36. [Google Scholar] [CrossRef]
- Song, S.; Nishikawa, T.; Isobe, T.; Ito, H.; Tanikawa, M.; Yoshizaki, K. Analysis of mechanism of different responses of acute-phase proteins to IL-6 and TNFα in chronic inflammatory diseases. Nihon Rinsho Men’eki Gakkai Sokai Shorokushu 2011, 39, 123. [Google Scholar] [CrossRef]
- Kav, T.; Akyol, A.; Aksoy, E.; Ozer, C.; Torgutalp, M.; Sivri, B. P029 Azelnidipine, a novel calcium channel blocker, ameliorates severity of colitis in DSS induced colitis in mice possibly by modulating tissue levels of TNF-alpha and IL-6. J. Crohn’s Colitis 2017, 11, S93–S94. [Google Scholar] [CrossRef] [Green Version]
- Neurath, M.-F. Cytokines in inflammatory bowel disease, Nature reviews. Immunology 2014, 14, 328–342. [Google Scholar] [CrossRef]
- Li, S.; Huo, X.; Qi, Y.; Ren, D.; Li, Z.; Qu, D.; Sun, Y. The Protective Effects of Ginseng Polysaccharides and Their Effective Subfraction against Dextran Sodium Sulfate-Induced Colitis. Foods 2022, 11, 890. [Google Scholar] [CrossRef]
- Han, G.; Cho, H.; Kim, H.; Jang, Y.; Jang, H.; Kim, D.-E.; Kim, E.-S.; Kim, E.-H.; Hwang, K.-Y.; Kim, K.; et al. Bovine colostrum derived-exosomes prevent dextran sulfate sodium-induced intestinal colitis via suppression of inflammation and oxidative stress. Biomater. Sci. 2022, 10, 2076–2087. [Google Scholar] [CrossRef]
- Cristina, S.R.; Rodrigo, M.; Carla, M.F.; Freire, B.A.; Cypriano, D.R.; Geraldo, P.M.; Calixto, J.B. Oral administration of the flavonoid myricitrin prevents dextran sulfate sodium-induced experimental colitis in mice through modulation of PI3K/Akt signaling pathway. Mol. Nutr. Food Res. 2013, 57, 1938–1949. [Google Scholar] [CrossRef]
- Torres, H.A.; Katelyn, R.-H.; Antonia, B.-M.; David, B. Tnfaip3 drives colitis through death cell and TNFα production. Inflamm. Bowel Dis. 2022, 28, S61. [Google Scholar] [CrossRef]
- Mahida, Y.-R.; Wu, K.; Jewell, D.-P. Enhanced production of interleukin 1-beta by mononuclear cells isolated from mucosa with active ulcerative colitis of Crohn’s disease. Gut 1989, 30, 835–838. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.-H.; Myunghoo, K.; Linchun, C.; Adrien, A.; Fatima, B.-S.; Daniel, F.-Z.; Kendra, N.; Samuel, B.-S.; Gretchen, E.-D. Interleukin-1β secretion induced by mucosa-associated gut commensal bacteria promotes intestinal barrier repair. Gut Microbes 2022, 14, 2014772. [Google Scholar] [CrossRef] [PubMed]
- Kasama, T.; Umemura, M.; Isojima, S.; Furuya, H.; Yanai, R.; Isozaki, T. Clinical Significance of Cytokines and Chemokines in Neuropsychiatric Systemic Lupus Erythematosus. Curr. Rheumatol. Rev. 2013, 8, 296–306. [Google Scholar] [CrossRef]
- Reynaud, J.-M.; François, J.-J.; Welsch, J.-C.; Branka, H. Human herpesvirus 6A infection in CD46 transgenic mice: Viral persistence in the brain and increased production of proinflammatory chemokines via Toll-like receptor 9. J. Virol. 2014, 88, 5421–5436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Meng, S.; Jiang, H.; Chen, T.; Wu, W. Exosomes derived from interleukin-10-treated dendritic cells can inhibit trinitrobenzene sulfonic acid-induced rat colitis. Scand. J. Gastroenterol. 2010, 45, 1168–1177. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Chen, Y.; Huang, C.; Wei, M.; Li, T.; Lv, Y.; Song, Q.; Mo, S. Adipose-derived mesenchymal stem cells may reduce intestinal epithelial damage in ulcerative colitis by communicating with macrophages and blocking inflammatory pathways: An analysis in silico. Aging 2022, 14, 2665–2677. [Google Scholar] [CrossRef]
- Nasu, Y.; Ido, A.; Tanoue, S.; Hashimoto, S.; Sasaki, F.; Takami, Y.; Kanmura, S.; Setoyama, H.; Funakawa, K.; Moriuchi, A.; et al. T1777 Hepatocyte Growth Factor Stimulates the Migration of Gastric Epithelial Cells by Altering the Intracellular Localization of the Tight Junction Protein ZO-1. Gastroenterology 2010, 138, S576. [Google Scholar] [CrossRef]
- Li, Y.; Li, Z.; Xie, C.-Y.; Fan, J.; Lv, J.; Xu, X.-J.; Lv, J.; Kuai, W.-T.; Jia, Y.-T. Gegen Qinlian decoction enhances immunity and protects intestinal barrier function in colorectal cancer patients via gut microbiota. World J. Gastroenterol. 2020, 26, 7633–7651. [Google Scholar] [CrossRef]
- Zhong, G.; Zhang, J.; Guo, Y.; Wang, Y.; Wu, M.; Ren, J.; Li, Y.; Zhang, X.; Zhou, B.; Zhao, W.; et al. IF-1 inactivation attenuates experimental colitis through downregulation of neutrophil infiltration in colon mucosa. Int. Immunopharmacol. 2021, 99, 107980. [Google Scholar] [CrossRef] [PubMed]
- Rapozo, D.C.; Bernardazzi, C.; De Souza, H.S. Diet and microbiota in inflammatory bowel disease: The gut in disharmony. World J. Gastroenterol. 2017, 23, 2124–2140. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Zu, X.-P.; Xie, H.-S.; Jin, H.-Z.; Yang, N.; Liu, X.-R.; Zhang, W.-D. Research progress in mechanism of intestinal microorganisms in human diseases. Yao Xue Xue Bao = Acta Pharm. Sin. 2016, 51, 843–852. (In Chinese) [Google Scholar] [CrossRef]
- Dou, X.; Ma, Z.; Yan, D.; Gao, N.; Li, Z.; Li, Y.; Feng, X.; Meng, L.; Shan, A. Sodium butyrate alleviates intestinal injury and microbial flora disturbance induced by lipopolysaccharides in rats. Food Funct. 2022, 13, 1360–1369. [Google Scholar] [CrossRef]
- Li, S.; Heng, X.; Guo, L.; James, L.-D.; Chu, W. SCFAs improve disease resistance via modulate gut microbiota, enhance immune response and increase antioxidative capacity in the host. Fish Shellfish. Immunol. 2022, 120, 560–568. [Google Scholar] [CrossRef]
- Gu, J.; Zhao, L.; Chen, Y.Z.; Guo, Y.X.; Sun, Y.; Guo, Q.; Duan, G.X.; Li, C.; Tang, Z.B.; Zhang, Z.X.; et al. Preventive effect of sanguinarine on intestinal injury in mice exposed to whole abdominal irradiation. Biomed. Pharmacother. 2022, 146, 112496. [Google Scholar] [CrossRef]
- Guo, C.; Wang, Y.; Zhang, S.; Zhang, X.; Du, Z.; Xi, M.; Ding, L. Crataegus pinnatifida polysaccharide alleviates colitis via modulation of gut microbiota and SCFAs metabolism. Int. J. Biol. Macromol. 2021, 181, 357–368. [Google Scholar] [CrossRef]
- Marteau, P. Bacterial flora in inflammatory bowel disease. Dig. Dis. 2009, 27 (Suppl. 1), 99–103. [Google Scholar] [CrossRef]
- Zubin, G.; Alicia, K.; Morrison, M.; Graham, L.; Radford, S.; Naoki, F.; Lisa, S.; Peter, L. The Relative Abundances of Dorea and Faecalibacterium spp. in the Mucosa Associated Microbiome of Newly Diagnosed Children with Crohn’s Disease Are Differentially Affected by Exclusive Enteral Nutrition. Gastroenterology 2016, 150, S132–S133. [Google Scholar] [CrossRef]
- Dziarski, R.; Park, S.-Y.; Kashyap, D.-R.; Dowd, S.-E.; Gupta, D. Pglyrp-Regulated Gut Microflora Prevotella falsenii, Parabacteroides distasonis and Bacteroides eggerthii Enhance and Alistipes finegoldii Attenuates Colitis in Mice. PLoS ONE 2016, 11, e0146162. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.; Zang, S.; Wei, J.; Yu, H.; Yang, Z.; Wu, H.; Kang, Y.; Tao, H.; Yang, M.; Jin, L.; et al. High-throughput sequencing provides insights into oral microbiota dysbiosis in association with inflammatory bowel disease. Genomics 2021, 113, 664–676. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Hand, T.-W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanoue, T.; Atarashi, K.; Honda, K. Development and maintenance of intestinal regulatory T cells. Nat. Rev. Immunol. 2016, 16, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Jia, C.; Zhang, X.; Wang, Z.; Li, Y.-P.; Fang, X. Identification of Key Genes Related with Aspartic Acid Metabolism and Corresponding Protein Expression in Human Colon Cancer with Postoperative Prognosis and the Underlying Molecular Pathways Prediction. Front. Cell Dev. Biol. 2022, 10, 812271. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| Il-1β | ATGCCACCTTTTGACAGTGATG | GATGTGCTGCTGCGAGATTT |
| IL-6 | GACTTCCATCCAGTTGCCTT | ACAACTCTTTTCTCATTTCCACGA |
| IL-10 | CTTACTGACTGGCATGAGGATCA | GCAGCTCTAGGAGCATGTGG |
| TNFα | AGCCGATGGGTTGTACCTTG | ATAGCAAATCGGCTGACGGT |
| TNFβ | GGAGGCATGTTCGGTAGTGG | CCCTGCGTTGGATTTCGTG |
| CCL2 | CCAGCAAGATGATCCCAATGAGT | CCATTCCTTCTTGGGGTCAGC |
| CXCL-1 | ACCCAAACCGAAGTCATAGCC | ACTTGGGGACACCTTTTAGCATC |
| GAPDH | TGTTTCCTCGTCCCGTAGACA | AACAATCTCCACTTTGCCACT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, D.; Zeng, J.; Guan, J.; Xu, Y.; Jia, R.-B.; Chen, J.; Jiang, G.; Zhou, C. Dietary Supplement of Amomum villosum Lour. Polysaccharide Attenuates Ulcerative Colitis in BALB/c Mice. Foods 2022, 11, 3737. https://doi.org/10.3390/foods11223737
Luo D, Zeng J, Guan J, Xu Y, Jia R-B, Chen J, Jiang G, Zhou C. Dietary Supplement of Amomum villosum Lour. Polysaccharide Attenuates Ulcerative Colitis in BALB/c Mice. Foods. 2022; 11(22):3737. https://doi.org/10.3390/foods11223737
Chicago/Turabian StyleLuo, Donghui, Jiao Zeng, Jingjing Guan, Yuanyuan Xu, Rui-Bo Jia, Jin Chen, Guili Jiang, and Chunxia Zhou. 2022. "Dietary Supplement of Amomum villosum Lour. Polysaccharide Attenuates Ulcerative Colitis in BALB/c Mice" Foods 11, no. 22: 3737. https://doi.org/10.3390/foods11223737