
Effects of Dietary Omega-3 Enrichment on the Chemical Composition and the Pathogenic Microbiota of Ovine Milk
 , ,
, ,  ,
,  , , , , ,
, , , , ,
Abstract
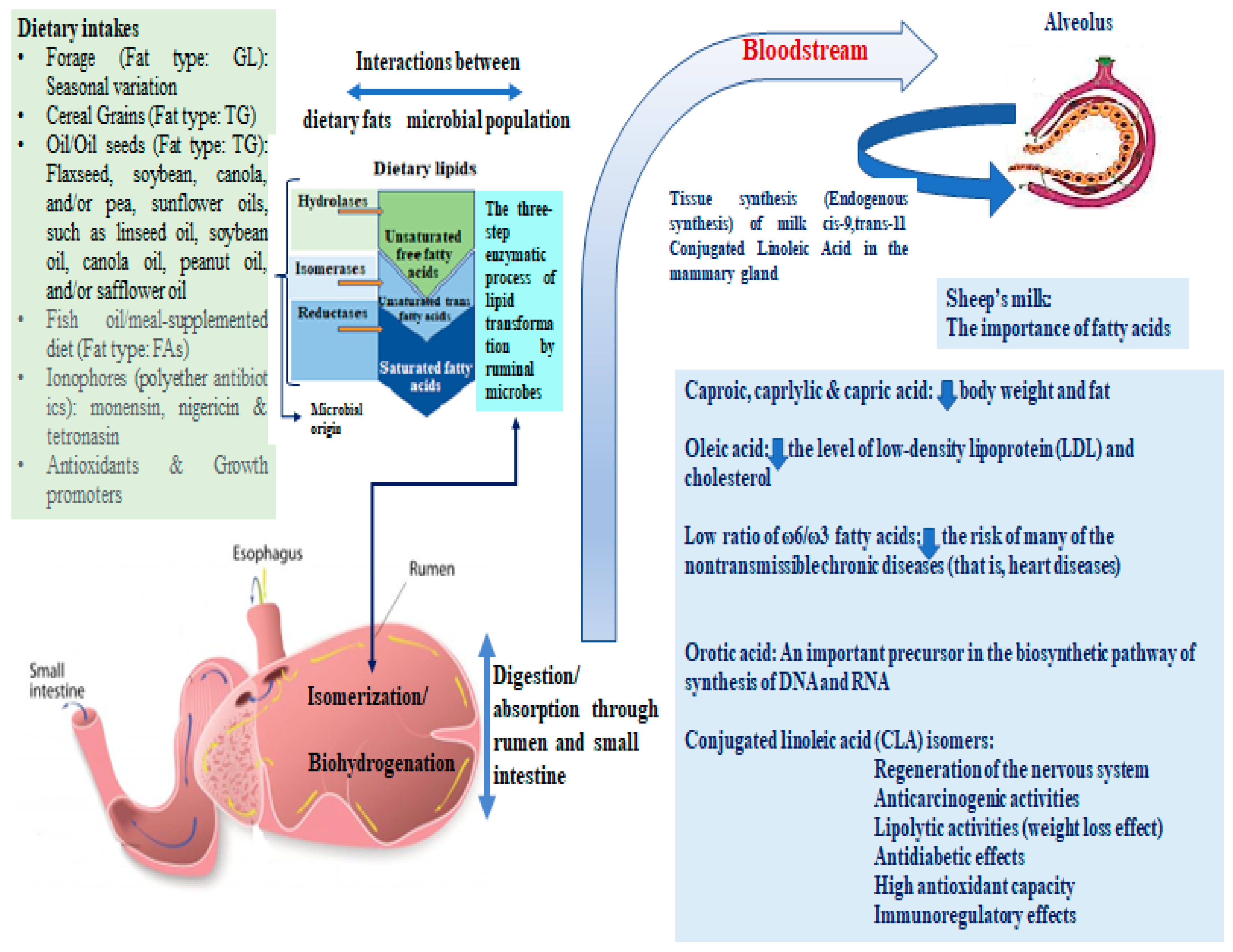
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.1.1. Animals, Experimental Diets, and Management
2.1.2. Milk Collection
2.1.3. Blood Sample Collection
2.2. Microbiological Analysis
Identification of Microbiota by the MALDI-TOF MS
2.3. Analysis of Samples by the California Mastitis Test and Categories of the Somatic Cell Count
2.4. Chemical Analysis/Biochemical Assay
2.4.1. Milk Samples
Determination of the Milk Samples FA Composition
2.4.2. Blood Samples
2.5. Calculations of Health Indexes
2.6. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
- −
- The supplementation of feeds rich in omega-3 FAs for a 30-day period increased the ovine milk fat and total solids content.
- −
- The milk’s FA profile was modified by the dietary omega-3 FA enrichment resulting in milk with increased amounts of UFA and improved atheromatic and thrombogenic indexes.
- −
- The pathogenic microbiota of the ovine milk was affected by the dietary omega-3 FA enrichment. Most pathogens were reduced or even eliminated in the second sampling, i.e., after 30 days of flaxseed and lupin administration. The biodiversity in terms of pathogenics for the udder species were altered towards a healthier direction. However, some pathogens remained while some other species emerged. Thus, the total effect can be possibly characterized as semi-protective for the udder.
- −
- These findings suggest that ovine milk could be beneficially modified by diets enriched in omega-3 FAs and could potentially be more appropriate for consumption by people belonging to high-risk groups for age-related and metabolic disorders.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Capper, J.L.; Wilkinson, R.G.; Mackenzie, A.M.; Sinclair, L.A. Polyunsaturated Fatty Acid Supplementation during Pregnancy Alters Neonatal Behavior in Sheep. J. Nutr. 2006, 136, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Bobiński, R.; Bobińska, J. Fatty acids of human milk—A review. Int. J. Vitam. Nutr. Res. 2020, 92, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Boquien, C.Y. Human Milk: An Ideal Food for Nutrition of Preterm Newborn. Front. Pediatr. 2018, 6, 295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietrzak-Fiećko, R.; Kamelska-Sadowska, A.M. The Comparison of Nutritional Value of Human Milk with Other Mammals’ Milk. Nutrients 2020, 12, 1404. [Google Scholar] [CrossRef] [PubMed]
- Flis, Z.; Molik, E. Importance of Bioactive Substances in Sheep’s Milk in Human Health. Int. J. Mol. Sci. 2021, 22, 4364. [Google Scholar] [CrossRef] [PubMed]
- Lund, J.; Rustan, A.C. Fatty Acids: Structures and Properties. In eLS.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2022. [Google Scholar] [CrossRef]
- Hanuš, O.; Samková, E.; Křížová, L.; Hasoňová, L.; Kala, R. Role of Fatty Acids in Milk Fat and the Influence of Selected Factors on Their Variability—A Review. Molecules 2018, 23, 1636. [Google Scholar] [CrossRef] [Green Version]
- Carta, G.; Murru, E.; Banni, S.; Manca, C. Palmitic Acid: Physiological Role, Metabolism and Nutritional Implications. Front. Physiol. 2017, 8, 902. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Luo, J. The Synthesis of Milk Medium-Chain Fatty Acids in Mammary Gland. In Fatty Acids [Internet]; Catala, A., Ed.; IntechOpen: London, UK, 2017; Available online: https://www.intechopen.com/chapters/54464 (accessed on 1 August 2022).
- Alshaikh, B.N.; Reyes Loredo, A.; Knauff, M.; Momin, S.; Moossavi, S. The Role of Dietary Fats in the Development and Prevention of Necrotizing Enterocolitis. Nutrients 2021, 14, 145. [Google Scholar] [CrossRef]
- Youdim, K.A.; Martin, A.; Joseph, J.A. Essential fatty acids and the brain: Possible health implications. Int. J. Dev. Neurosci. 2020, 18, 383–399. [Google Scholar] [CrossRef]
- Simopoulos, A.P. n−3 fatty acids and human health: Defining strategies for public policy. Lipids 2001, 36, S83–S89. [Google Scholar] [CrossRef]
- Pipoyan, D.; Stepanyan, S.; Stepanyan, S.; Beglaryan, M.; Costantini, L.; Molinari, R.; Merendino, N. The Effect of Trans Fatty Acids on Human Health: Regulation and Consumption Patterns. Foods 2021, 10, 2452. [Google Scholar] [CrossRef] [PubMed]
- Den Hartigh, L.J. Conjugated Linoleic Acid Effects on Cancer, Obesity, and Atherosclerosis: A Review of Pre-Clinical and Human Trials with Current Perspectives. Nutrients 2019, 11, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weylandt, K.H.; Serini, S.; Chen, Y.Q.; Su, H.-M.; Lim, K.; Cittadini, A.; Calviello, G. Omega-3 Polyunsaturated Fatty Acids: The Way Forward in Times of Mixed Evidence. BioMed Res. Int. 2015, 2015, 143109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reimers, A.; Ljung, H. The emerging role of omega-3 fatty acids as a therapeutic option in neuropsychiatric disorders. Ther. Adv. Psychopharmacol. 2019, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, T.S.; Giromini, C.; Rebucci, R.; Pistl, J.; Bhide, M.; Baldi, A. Role of omega-3 polyunsaturated fatty acids, citrus pectin, and milk-derived exosomes on intestinal barrier integrity and immunity in animals. J. Anim. Sci. Biotechnol. 2022, 13, 40. [Google Scholar] [CrossRef]
- Giosuè, A.; Calabrese, I.; Vitale, M.; Riccardi, G.; Vaccaro, O. Consumption of Dairy Foods and Cardiovascular Disease: A Systematic Review. Nutrients 2022, 14, 831. [Google Scholar] [CrossRef]
- Nguyen, Q.V.; Malau-Aduli, B.; Cavalieri, J.; Malau-Aduli, A.E.O.; Nichols, P. Enhancing Omega-3 Long-Chain Polyunsaturated Fatty Acid Content of Dairy-Derived Foods for Human Consumption. Nutrients 2019, 11, 743. [Google Scholar] [CrossRef] [Green Version]
- Bzikowska-Jura, A.; Czerwonogrodzka-Senczyna, A.; Jasińska-Melon, E.; Mojska, H.; Olędzka, G.; Wesołowska, A.; Szostak-Węgierek, D. The Concentration of Omega-3 Fatty Acids in Human Milk Is Related to Their Habitual but Not Current Intake. Nutrients 2019, 11, 1585. [Google Scholar] [CrossRef] [Green Version]
- Nudda, A.; Cannas, A.; Pulina, G. Nutritional approaches to improve the fatty acid profile of milk fat in sheep and goats. In Innovation for Sustainability in Sheep and Goats; Ruiz, R., López-Francos, A., López Marco, L., Eds.; CIHEAM: Zaragoza, Spain, 2019; pp. 73–82. Available online: http://om.ciheam.org/article.php?IDPDF=00007862 (accessed on 20 August 2022).
- Lai, G.; Pes, M.; Addis, M.; Pirisi, A. A Cluster Project Approach to Develop New Functional Dairy Products from Sheep and Goat Milk. Dairy 2020, 1, 10. [Google Scholar] [CrossRef]
- Moran, C.A.; Morlacchini, M.; Fusconi, G. Enhancing the DHA content in milk from dairy cows by feeding ALL-G-RICH™. J. Appl. Anim. Nutr. 2017, 5, e11. [Google Scholar] [CrossRef] [Green Version]
- Gagliostro, G.A.; Antonacci, L.E.; Pérez, C.D.; Rossetti, L.; Carabajal, A. Improving the Quality of Milk Fatty Acid in Dairy Cows Supplemented with Soybean Oil and DHA-Micro Algae in a Confined Production System. Agric. Sci. 2018, 09, 1115–1130. [Google Scholar] [CrossRef] [Green Version]
- Kiss, A.; Naár, Z.; Daróczi, L.; Kukovics, S.; Kukovics, F.; Mirmazloum, I.; Némedi, E. Changes of free fatty acid composition and number of lactic acid bacteria in three functional goat and sheep milk products fortified with inulin or fish oil. CyTA—J. Food 2019, 17, 51–59. [Google Scholar] [CrossRef] [Green Version]
- FAOSTAT. Statistical Database of the Food and Agriculture Organization of the United Nations. Available online: http://www.faostat.fao.org (accessed on 20 August 2022).
- Mollica, M.P.; Trinchese, G.; Cimmino, F.; Penna, E.; Cavaliere, G.; Tudisco, R.; Musco, N.; Manca, C.; Catapano, A.; Monda, M.; et al. Milk Fatty Acid Profiles in Different Animal Species: Focus on the Potential Effect of Selected PUFAs on Metabolism and Brain Functions. Nutrients 2021, 13, 1111. [Google Scholar] [CrossRef]
- Queiroz, M.P.; Lima, M.D.S.; Barbosa, M.Q.; de Melo, M.F.F.T.; Bertozzo, C.C.M.S.; de Oliveira, M.E.G.; Bessa, R.J.B.; Alves, S.P.A.; Souza, M.I.A.; Queiroga, R.C.R.D.E.; et al. Effect of Conjugated Linoleic Acid on Memory and Reflex Maturation in Rats Treated During Early Life. Front. Neurosci. 2019, 13, 370. [Google Scholar] [CrossRef] [PubMed]
- Destaillats, F.; Japiot, C.; Chouinard, P.Y.; Arul, J.; Angers, P. Short Communication: Rearrangement of Rumenic Acid in Ruminant Fats: A Marker of Thermal Treatment. J. Dairy Sci. 2005, 88, 1631–1635. [Google Scholar] [CrossRef] [Green Version]
- Balthazar, C.F.; Pimentel, T.C.; Ferrão, L.L.; Almada, C.N.; Santillo, A.; Albenzio, M.; Mollakhalili, N.; Mortazavian, A.M.; Nascimento, J.S.; Silva, M.C.; et al. Sheep Milk: Physicochemical Characteristics and Relevance for Functional Food Development. Compr. Rev. Food Sci. Food Saf. 2017, 16, 247–262. [Google Scholar] [CrossRef] [Green Version]
- Molkentin, J.; Precht, D. Validation of a gas-chromatographic method for the determination of milk fat contents in mixed fats by butyric acid analysis. Eur. J. Lipid Sci. Technol. 2020, 102, 194–201. [Google Scholar] [CrossRef]
- Lurueña-Martínez, M.A.; Revilla, I.; Severiano-Pérez, P.; Vivar-Quintana, A.M. The influence of breed on the organoleptic characteristics of Zamorano sheep’s raw milk cheese and its assessment by instrumental analysis. Int. J. Dairy Technol. 2010, 63, 216–223. [Google Scholar] [CrossRef]
- Malissiova, E.; Tzora, A.; Katsioulis, A.; Hatzinikou, M.; Tsakalof, A.; Arvanitoyannis, I.S.; Govaris, A.; Hadjichristodoulou, C. Relationship between production conditions and milk gross composition in ewe’s and goat’s organic and conventional farms in central Greece. Dairy Sci. Technol. 2015, 95, 437–450. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The Importance of the Omega-6/Omega-3 Fatty Acid Ratio in Cardiovascular Disease and Other Chronic Diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef]
- Premier Nutrition. Premier Atlas 2014. Ingredients Matrix; Premier Nutrition: Brereton, UK, 2014; Available online: https://www.premiernutrition.co.uk/news-insight/new-global-animal-feed-ingredients-matrix-launched (accessed on 30 August 2022).
- Vyletělová, M.; Vlková, H.; Manga, I. Occurrence and characteristics of methicillin resistant Staphylococcus aureus and methicillin resistant coagulase-negative staphylococci in raw milk manufacturing. Czech J. Food Sci. 2011, 29, S11–S16. [Google Scholar] [CrossRef] [Green Version]
- Moura, G.S.; Gebreyes, W.A.; Marques, M.F.S.; Stipp, D.T.; Souza, F.N.; Da Costa, L.B.; Oliveira, C.J.B. Short communication: Occurrence of methicillin-resistant Staphylococcus aureus and coagulase-negative staphylococci in dairy goat herds in Ohio, United States. J. Dairy Sci. 2018, 101, 7804–7807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzora, A.; Nelli, A.; Voidarou, C.; Fthenakis, G.; Rozos, G.; Theodorides, G.; Bonos, E.; Skoufos, I. Microbiota “Fingerprint” of Greek Feta Cheese through Ripening. Appl. Sci. 2021, 11, 5631. [Google Scholar] [CrossRef]
- Tzora, A.; Nelli, A.; Voidarou, C.; Fotou, K.; Bonos, E.; Rozos, G.; Grigoriadou, K.; Papadopoulos, P.; Basdagianni, Z.; Giannenas, I.; et al. Impact of an Omega-3-Enriched Sheep Diet on the Microbiota and Chemical Composition of Kefalograviera Cheese. Foods 2022, 11, 843. [Google Scholar] [CrossRef] [PubMed]
- Fthenakis, G.C. Prevalence and aetiology of subclinical mastitis in ewes of southern Greece. Small Rumin. Res. 1994, 13, 293–300. [Google Scholar] [CrossRef]
- Zigo, F.; Vasiľ, M.; Takáč, L.; Zigová, M.; Elečko, J. Mastitis Pathogens Isolated from Raw Milk Samples on Sheep Farms Situated in Marginal Parts of Slovakia. Folia Vet. 2018, 62, 56–61. [Google Scholar] [CrossRef] [Green Version]
- Currò, S.; Manuelian, C.L.; Penasa, M.; Cassandro, M.; De Marchi, M. Technical note: Feasibility of near infrared trans-mittance spectroscopy to predict cheese ripeness. J. Dairy Sci. 2017, 100, 8759–8763. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- International Standard ISO 15884-IDF 182; Milk Fat-Preparation of Fatty Acid Methyl Esters. International Organisation for Standardisation (ISO): Geneva, Switzerland, 2002.
- Papaloukas, L.; Sinapis, E.; Arsenos, G.; Kyriakou, G.; Basdagianni, Z. Effect of season on fatty acid and terpene profiles of milk from Greek sheep raised under a semi-extensive production system. J. Dairy Res. 2016, 83, 375–382. [Google Scholar] [CrossRef]
- Ulbricht, T.L.V.; Southgate, D.A.T. Coronary heart disease: Seven dietary factors. Lancet 1991, 338, 982–992. [Google Scholar] [CrossRef]
- Paszczyk, B.; Łuczyńska, J. The Comparison of Fatty Acid Composition and Lipid Quality Indices in Hard Cow, Sheep, and Goat Cheeses. Foods 2020, 9, 1667. [Google Scholar] [CrossRef] [PubMed]
- Osmari, E.K.; Cecato, U.; Macedo, F.A.F.; Souza, N.E. Nutritional quality indices of milk fat from goats on diets supplemented with different roughages. Small Rumin. Res. 2011, 98, 128–132. [Google Scholar] [CrossRef]
- Markiewicz-Kęszycka, M.; Czyżak-Runowska, G.; Lipińska, P.; Wójtowski, J. Fatty Acid Profile of Milk—A Review. Bull. Vet. Inst. Puławy 2013, 57, 135–139. [Google Scholar] [CrossRef] [Green Version]
- Astrup, A.; Magkos, F.; Bier, D.M.; Brenna, J.T.; De Oliveira Otto, M.C.; Hill, J.O.; King, J.C.; Mente, A.; Ordovas, J.M.; Volek, J.S.; et al. Saturated Fats and Health: A Reassessment and Proposal for Food-Based Recommendations. J. Am. Coll. Cardiol. 2020, 76, 844–857. [Google Scholar] [CrossRef]
- Mccrorie, T.A.; Keaveney, E.M.; Wallace, J.M.W.; Binns, N.; Livingstone, M.B.E. Human health effects of conjugated linoleic acid from milk and supplements. Nutr. Res. Rev. 2011, 24, 206–227. [Google Scholar] [CrossRef] [Green Version]
- Basak, S.; Duttaroy, A.K. Conjugated Linoleic Acid and Its Beneficial Effects in Obesity, Cardiovascular Disease, and Cancer. Nutrients 2020, 12, 1913. [Google Scholar] [CrossRef]
- Santurino, C.; López-Plaza, B.; Fontecha, J.; Calvo, M.V.; Bermejo, L.M.; Gómez-Andrés, D.; Gómez-Candela, C. Consumption of Goat Cheese Naturally Rich in Omega-3 and Conjugated Linoleic Acid Improves the Cardiovascular and Inflammatory Biomarkers of Overweight and Obese Subjects: A Randomized Controlled Trial. Nutrients 2020, 12, 1315. [Google Scholar] [CrossRef]
- Morato-Martínez, M.; López-Plaza, B.; Santurino, C.; Palma-Milla, S.; Gómez-Candela, C. A Dairy Product to Reconstitute Enriched with Bioactive Nutrients Stops Bone Loss in High-Risk Menopausal Women without Pharmacological Treatment. Nutrients 2020, 12, 2203. [Google Scholar] [CrossRef]
- Moatsou, G.; Samolada, M.; Katsabeki, A.; Anifantakis, E. Casein fraction of ovine milk from indigenous Greek breeds. Le Lait 2004, 84, 285–296. [Google Scholar] [CrossRef]
- Barłowska, J.; Szwajkowska, M.; Litwińczuk, Z.; Król, J. Nutritional Value and Technological Suitability of Milk from Various Animal Species Used for Dairy Production. Compr. Rev. Food Sci. Food Saf. 2011, 10, 291–302. [Google Scholar] [CrossRef]
- Haenlein, G.F.W.; Wendorff, W.L. Sheep Milk. In Handbook of Milk of Non-Bovine Mammals; Park, Y.W., Haenlein, G.F.W., Eds.; Blackwell Publishing: Ames, IA, USA, 2006; pp. 137–194. [Google Scholar]
- Park, Y.W. Rheological characteristics of goat and sheep milk. Small Rumin. Res. 2007, 68, 73–87. [Google Scholar] [CrossRef]
- Park, Y.W. Overview of Bioactive Components in Milk and Dairy Products. In Bioactive Components in Milk and Dairy Products; Park, Y.W., Ed.; Wiley-Blackwell: Ames, IA, USA, 2009; pp. 3–4. Print ISBN 9780813819822|Online ISBN 9780813821504. [Google Scholar] [CrossRef] [Green Version]
- Caroprese, M.; Albenzio, M.; Bruno, A.; Fedele, V.; Santillo, A.; Sevi, A. Effect of solar radiation and flaxseed supplementation on milk production and fatty acid profile of lactating ewes under high ambient temperature. J. Dairy Sci. 2011, 94, 3856–3867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nudda, A.; Cannas, A.; Correddu, F.; Atzori, A.S.; Lunesu, M.F.; Battacone, G.; Pulina, G. Sheep and Goats Respond Differently to Feeding Strategies Directed to Improve the Fatty Acid Profile of Milk Fat. Animals 2020, 10, 1290. [Google Scholar] [CrossRef] [PubMed]
- Pulina, G.; Nudda, A.; Battacone, G.; Cannas, A. Effects of nutrition on the contents of fat, protein, somatic cells, aromatic compounds, and undesirable substances in sheep milk. Anim. Feed. Sci. Technol. 2006, 131, 255–291. [Google Scholar] [CrossRef]
- Vlaeminck, B.; Fievez, V.; Tamminga, S.; Dewhurst, R.J.; van Vuuren, A.; De Brabander, D.; Demeyer, D. Milk odd- and branched-chain fatty acids in relation to the rumen fermentation pattern. J. Dairy Sci. 2006, 89, 3954–3964. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, K.; Hao, X.; Xin, H. The relationships between odd- and branched-chain fatty acids to ruminal fermentation parameters and bacterial populations with different dietary ratios of forage and concentrate. J. Anim. Physiol. Anim. Nutr. 2017, 101, 1103–1114. [Google Scholar] [CrossRef]
- Taormina, V.M.; Unger, A.L.; Schiksnis, M.R.; Torres-Gonzalez, M.; Kraft, J. Branched-Chain Fatty Acids—An Underexplored Class of Dairy-Derived Fatty Acids. Nutrients 2020, 12, 2875. [Google Scholar] [CrossRef]
- Czauderna, M.; Kowalczyk, J.; Niedźwiedzka, K.M.; Wąsowska, I.; Pastuszewska, B. Conjugated linoleic acid (CLA) content and fatty acids composition of muscle in rats fed isomers of CLA and selenium. J. Anim. Feed. Sci. 2004, 13, 183–196. [Google Scholar] [CrossRef]
- Martysiak-Żurowska, D.; Kiełbratowska, B.; Szlagatys-Sidorkiewicz, A. The content of conjugated linoleic acid and vaccenic acid in the breast milk of women from Gdansk and the surrounding district, as well as in, infant formulas and follow-up formulas. nutritional recommendation for nursing women. Dev. Period Med. 2018, 22, 128–134. [Google Scholar] [CrossRef]
- Kharitonov, E. The Processes of Nutrition and Metabolism Affecting the Biosynthesis of Milk Components and Vitality of Cows with High- and Low-Fat Milk. Animals 2022, 12, 604. [Google Scholar] [CrossRef]
- Hervás, G.; Toral, P.G.; Fernández-Díez, C.; Badia, A.D.; Frutos, P. Effect of Dietary Supplementation with Lipids of Different Unsaturation Degree on Feed Efficiency and Milk Fatty Acid Profile in Dairy Sheep. Animals 2021, 11, 2476. [Google Scholar] [CrossRef] [PubMed]
- Kompan, D.; Komprej, A. The Effect of Fatty Acids in Goat Milk on Health. In Milk Production—An Up-to-Date Overview of Animal Nutrition, Management and Health [Internet]; Chaiyabutr, N., Ed.; IntechOpen: London, UK, 2012; Available online: https://www.intechopen.com/chapters/39464 (accessed on 30 August 2022).
- Valvo, M.A.; Bella, M.; Scerra, M.; Biondi, L. Effects of ewe feeding system (grass vs concentrate) on milk fatty acid composition. In Advanced Nutrition and Feeding Strategies to Improve Sheep and Goat; Priolo, A., Biondi, L., Ben Salem, H., Morand-Fehr, P., Eds.; CIHEAM: Zaragoza, Spain, 2007; pp. 227–231. [Google Scholar]
- Toral, P.G.; Hervás, G.; Frutos, P. Effect of lipid supplementation on the endogenous synthesis of milk cis-9, trans-11 conjugated linoleic acid in dairy sheep and goats: A tracer assay with 13C-vaccenic acid. J. Dairy Sci. 2022, 105, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Sampelayo, M.S.; Chilliard, Y.; Schmidely, P.; Boza, J. Influence of type of diet on the fat constituents of goat and sheep milk. Small Rumin. Res. 2007, 68, 42–63. [Google Scholar] [CrossRef]
- Zervas, G.; Tsiplakou, E. The effect of feeding systems on the characteristics of products from small ruminants. Small Rumin. Res. 2011, 101, 140–149. [Google Scholar] [CrossRef]
- Thum, C.; Roy, N.C.; Everett, D.W.; Mcnabb, W.C. Variation in milk fat globule size and composition: A source of bioactives for human health. Crit. Rev. Food Sci. Nutr. 2021, 1–27. [Google Scholar] [CrossRef]
- Fragkou, I.; Skoufos, J.; Cripps, P.J.; Kyriazakis, I.; Papaioannou, N.; Boscos, C.M.; Tzora, A.; Fthenakis, G.C. Differences in susceptibility to Manhaemia haemolytica—Associated mastitis, between two breeds of dairy sheep. J. Dairy Res. 2007, 74, 349–355. [Google Scholar] [CrossRef]
- Fragkou, I.A.; Gougoulis, D.A.; Billinis, C.; Mavrogianni, V.S.; Bushnell, M.J.; Cripps, P.J.; Tzora, A.; Fthenakis, G.C. Transmission of Mannheimia haemolytica from the tonsils of lambs to the teat of ewes during sucking. Vet. Microbiol. 2011, 148, 66–74. [Google Scholar] [CrossRef]
- Scott, P. Antibiotic Treatment Response of Chronic Lung Diseases of Adult Sheep in the United Kingdom Based upon Ultrasonographic Findings. Vet. Med. Int. 2014, 2014, 537501. [Google Scholar] [CrossRef]
- Ghazvini, K.; Karbalaei, M.; Kianifar, H.; Keikha, M. The first report of Streptococcus pluranimalium infection from Iran: A case report and literature review. Clin. Case Rep. 2019, 7, 1858–1862. [Google Scholar] [CrossRef] [Green Version]
- Chinnam, B.K.; Nelapati, S.; Tumati, S.R.; Bobbadi, S.; Peddada, V.C.; Bodempudi, B. Detection of β-Lactamase-Producing Proteus mirabilis Strains of Animal Origin in Andhra Pradesh, India and Their Genetic Diversity. J. Food Prot. 2021, 84, 1374–1379. [Google Scholar] [CrossRef]
- Wicaksono, D.P.; Washio, J.; Abiko, Y.; Domon, H.; Takahashi, N. Nitrite Production from Nitrate and Its Link with Lactate Metabolism in Oral Veillonella spp. Appl. Environ. Microbiol. 2020, 86, e01255-20. [Google Scholar] [CrossRef] [PubMed]
- Fehr, K.; Moossavi, S.; Sbihi, H.; Boutin, R.C.; Bode, L.; Robertson, B.; Yonemitsu, C.; Field, C.J.; Becker, A.B.; Mandhane, P.J.; et al. Breastmilk Feeding Practices Are Associated with the Co-Occurrence of Bacteria in Mothers’ Milk and the Infant Gut: The CHILD Cohort Study. Cell Host Microbe 2020, 28, 285–297.e4. [Google Scholar] [CrossRef] [PubMed]
- Libera, K.; Konieczny, K.; Grabska, J.; Smulski, S.; Szczerbal, I.; Szumacher-Strabel, M.; Pomorska-Mól, M. Potential Novel Biomarkers for Mastitis Diagnosis in Sheep. Animals 2021, 11, 2783. [Google Scholar] [CrossRef] [PubMed]
- Ridler, A.L.; Rout-Brown, G.; Flay, K.J.; Velathanthiri, N.; Grinberg, A. Defects and bacterial pathogens in udders of non-dairy breed ewes from New Zealand. N. Z. J. Agric. Res. 2022, 65, 163–171. [Google Scholar] [CrossRef]


{kind=link}
| Ingredients (On Fresh Weight Basis) | Control Diet (g/Day/ewe) | Experimental Diet (g/Day/ewe) |
|---|---|---|
| Lucerne hay | 1.200 | 1.200 |
| Barley straw | 300 | 300 |
| Corn grain | 540 | 590 |
| Barley grain | 330 | 150 |
| Wheat bran | 180 | 110 |
| Sunflower seed meal (36% crude protein) | 30 | 180 |
| Soybean meal (47% crude protein) | 300 | 150 |
| Cotton seed | 30 | 80 |
| Flaxseed | - | 75 |
| Lupin seed | - | 75 |
| Molasses | 20 | 20 |
| Premix 1 containing vitamins and inorganic minerals | 60 | 60 |
| Total dry matter intake/day | 2630 | 2640 |
| Chemical Analysis (%) | ||
| Dry matter | 87.4 | 87.5 |
| Crude protein (N × 6.25) | 16.1 | 16.1 |
| Ether extract | 2.82 | 4.01 |
| Neutral detergent fiber | 33.8 | 35.1 |
| Acid detergent fiber | 19.5 | 19.9 |
| Acid detergent lignin | 4.51 | 4.98 |
| Ash | 6.55 | 6.52 |
| Starch | 18.0 | 16.1 |
| Calculated analysis | ||
| Calcium (g/kg) | 11.5 | 11.6 |
| Phosphorus (total) (g/kg) | 3.82 | 3.98 |
| PDI (g/kg DM) | 89.4 | 87.5 |
| PDIA (g/kg DM) | 44.3 | 42.7 |
| UFL | 0.71 | 0.71 |
| Total saturated fatty acids (%) | 0.48 | 0.59 |
| Total unsaturated fatty acids (%) | 1.69 | 2.72 |
| Omega-6 fatty acids (%) | 0.99 | 1.15 |
| Omega-3 fatty acids (%) | 0.28 | 0.82 |
| Omega-6/Omega-3 | 3.53 | 1.41 |
| Milk Analysis | Control Diet Day 30 of Trial | Experimental Diet Day 60 of Trial | SEM | p |
|---|---|---|---|---|
| Milk amount at morning milking (mL) | 1375.0 | 1371.4 | 47.03 | 0.971 |
| Milk total mesophilic flora (Log CFU/g) | 4.119 | 3.808 | 0.1740 | 0.381 |
| Milk fat (%) | 3.85 a | 5.50 b | 0.203 | <0.001 |
| Milk protein (%) | 4.59 | 4.66 | 0.071 | 0.648 |
| Milk lactose (%) | 5.36 | 5.43 | 0.035 | 0.277 |
| Milk total solids (%) | 14.57 a | 16.22 b | 0.223 | <0.001 |
| Milk solids nonfat (%) | 10.62 | 10.76 | 0.072 | 0.359 |
| Milk Fatty Acid Profile (% of Total Fatty Acids) | Control Diet Day 30 of Trial | Experimental Diet Day 60 of Trial | SEM | p |
|---|---|---|---|---|
| C4:0; Butyric | 0.73 | 0.90 | 0.122 | 0.479 |
| C6:0; Caproic | 4.25 | 4.43 | 0.320 | 0.790 |
| C8:0; Caprylic | 4.43 | 4.51 | 0.242 | 0.867 |
| C10:0; Capric | 8.84 a | 11.47 b | 0.672 | 0.048 |
| C12:0; Lauric | 5.24 | 4.96 | 0.843 | 0.870 |
| C14:0; Myristic | 11.56 | 11.16 | 0.500 | 0.695 |
| C14:1; Myristoleic | 0.43 b | 0.29 a | 0.036 | 0.047 |
| C15:0; Pentadecylic | 1.15 | 1.15 | 0.084 | 0.985 |
| C15:1; Pentadecenoic acid | 0.12 | 0.10 | 0.021 | 0.650 |
| C16:0; Palmitic | 26.22 b | 22.92 a | 0.689 | 0.013 |
| C16:1; Palmitoleic | 0.99 | 0.82 | 0.098 | 0.405 |
| C17:0; Margaric | 0.72 | 0.58 | 0.071 | 0.346 |
| C17:1 ω10 cis; Heptadecenoic acid | 0.15 | 0.11 | 0.030 | 0.522 |
| C18:0; Stearic | 12.87 b | 9.51 a | 0.766 | 0.025 |
| C18:1 ω9 trans; Elaidic | 1.17 | 0.98 | 0.138 | 0.488 |
| C18:1; ω11 trans; Vaccenic acid | 2.09 | 2.29 | 0.273 | 0.725 |
| C18:1 ω9 cis; Oleic | 16.23 a | 18.56 b | 0.559 | 0.034 |
| C18:2 trans; Conjugated linoleic acid | 0.11 | 0.26 | 0.050 | 0.124 |
| C18:2 ω6 trans; Linoelaidic | 0.29 a | 1.83 b | 0.247 | 0.001 |
| C18:2 ω6 cis; Linoleic | 2.21 | 2.20 | 0.223 | 0.978 |
| C18:3 ω3; Linolenic acid | 0.17 a | 0.73 b | 0.065 | <0.001 |
| Short-chain fatty acids—SCFA | 4.98 | 5.33 | 0.349 | 0.624 |
| Medium-chain saturated fatty acids—MCFA | 18.51 | 20.94 | 1.155 | 0.303 |
| Long-chain saturated fatty acids—LCFA | 76.47 | 73.47 | 1.318 | 0.265 |
| Saturated fatty acids—SFA | 76.00 b | 71.59 a | 0.889 | 0.010 |
| Unsaturated fatty acids—UFA | 23.95 b | 28.15 a | 0.862 | 0.011 |
| UFA/SFA ratio | 0.32 a | 0.40 b | 0.017 | 0.014 |
| Monounsaturated fatty acids—MUFA | 21.18 | 23.14 | 0.649 | 0.133 |
| Polyunsaturated fatty acids—PUFA | 2.78 a | 5.01 b | 0.349 | <0.001 |
| PUFA ω3 | 0.17 a | 0.73 b | 0.065 | <0.001 |
| PUFA ω6 | 2.50 a | 4.02 b | 0.287 | 0.005 |
| ω6/ω3 | 17.32 b | 5.85 a | 2.047 | 0.003 |
| Δ-9 dehydrogenase C14:1/C14:0 | 0.04 | 0.03 | 0.004 | 0.136 |
| Δ-9 dehydrogenase C16:1/C16:0 | 0.04 | 0.04 | 0.004 | 0.923 |
| Atherogenicity index (C12:0 + 4 × C14:0 + C16:0)/UFA | 3.30 b | 2.67 a | 0.151 | 0.034 |
| Thrombogenic index (C14:0 + C16:0 + C18:0)/(0.5 × MUFA + 0.5 × PUFAω6 + 3 × PUFAω3 + ω3/ω6) | 4.11 b | 2.79 a | 0.176 | <0.001 |
| Blood Parameters | Control Diet Day 30 of Trial | Experimental Diet Day 60 of Trial | SEM | p |
|---|---|---|---|---|
| Albumin—ALB (g/DL) | 3.0 | 2.9 | 0.06 | 0.762 |
| Alanine aminotransferase—ALT (U/L) | 59.6 | 66.9 | 2.65 | 0.174 |
| Aspartate aminotransferase—AST (U/L) | 130.7 | 125.2 | 7.37 | 0.720 |
| Glucose—GLU (mg/DL) | 62.7 | 63.4 | 2.86 | 0.907 |
| Cholesterol—CHOL (mg/DL) | 44.1 a | 57.3 b | 2.20 | 0.001 |
| Creatine kinase—CK (U/L) | 74.9 | 81.9 | 3.55 | 0.337 |
| Species | Control Diet Day 30 of Trial | Experimental Diet Day 60 of Trial | Isolated Only on Day 30 | Isolated Both on Day 30 and Day 60 | Isolated Only on Day 60 |
|---|---|---|---|---|---|
| Corynebacterium stationis | 9 | 7 | 4 | 5 | 2 |
| Staphylococcus xylosus | 8 | 4 | 6 | 2 | 2 |
| Staphylococcus epidermidis | 8 | 4 | 7 | 1 | 3 |
| Staphylococcus chromogenes | 2 | - | - | - | - |
| Staphylococcus auricularis | 1 | - | - | -- | - |
| Staphylococcus petrasii | 1 | - | - | - | - |
| Staphylococcus haemolyticus | - | 3 | - | - | - |
| Staphylococcus aureus | - | 1 | - | - | - |
| Staphylococcus caprae | - | 1 | - | - | - |
| Manchemia haemolytica | 1 | - | - | - | - |
| Streptococcus pluranimalis | - | 1 | - | - | - |
| Streptococcus parasanguinus | - | 1 | - | - | - |
| Proteus mirabilis | - | 1 | - | - | - |
| Veillonella parvula | - | 1 | - | - | - |
| Enterobacter kobei | - | 1 | - | - | - |
| Enterobacter cloacae | - | 1 | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tzora, A.; Voidarou, C.; Giannenas, I.; Bonos, E.; Fotou, K.; Nelli, A.; Grigoriadou, K.; Karamoutsios, A.; Basdagianni, Z.; Dokou, S.; et al. Effects of Dietary Omega-3 Enrichment on the Chemical Composition and the Pathogenic Microbiota of Ovine Milk. Foods 2022, 11, 3736. https://doi.org/10.3390/foods11223736
Tzora A, Voidarou C, Giannenas I, Bonos E, Fotou K, Nelli A, Grigoriadou K, Karamoutsios A, Basdagianni Z, Dokou S, et al. Effects of Dietary Omega-3 Enrichment on the Chemical Composition and the Pathogenic Microbiota of Ovine Milk. Foods. 2022; 11(22):3736. https://doi.org/10.3390/foods11223736
Chicago/Turabian StyleTzora, Athina, Chrysoula (Chrysa) Voidarou, Ilias Giannenas, Eleftherios Bonos, Konstantina Fotou, Aikaterini Nelli, Katerina Grigoriadou, Achilleas Karamoutsios, Zoitsa Basdagianni, Stella Dokou, and et al. 2022. "Effects of Dietary Omega-3 Enrichment on the Chemical Composition and the Pathogenic Microbiota of Ovine Milk" Foods 11, no. 22: 3736. https://doi.org/10.3390/foods11223736