
Synergetic Effect of Metschnikowia pulcherrima and Lachancea thermotolerans in Acidification and Aroma Compounds in Airén Wines


Abstract
:1. Introduction
2. Materials and Methods
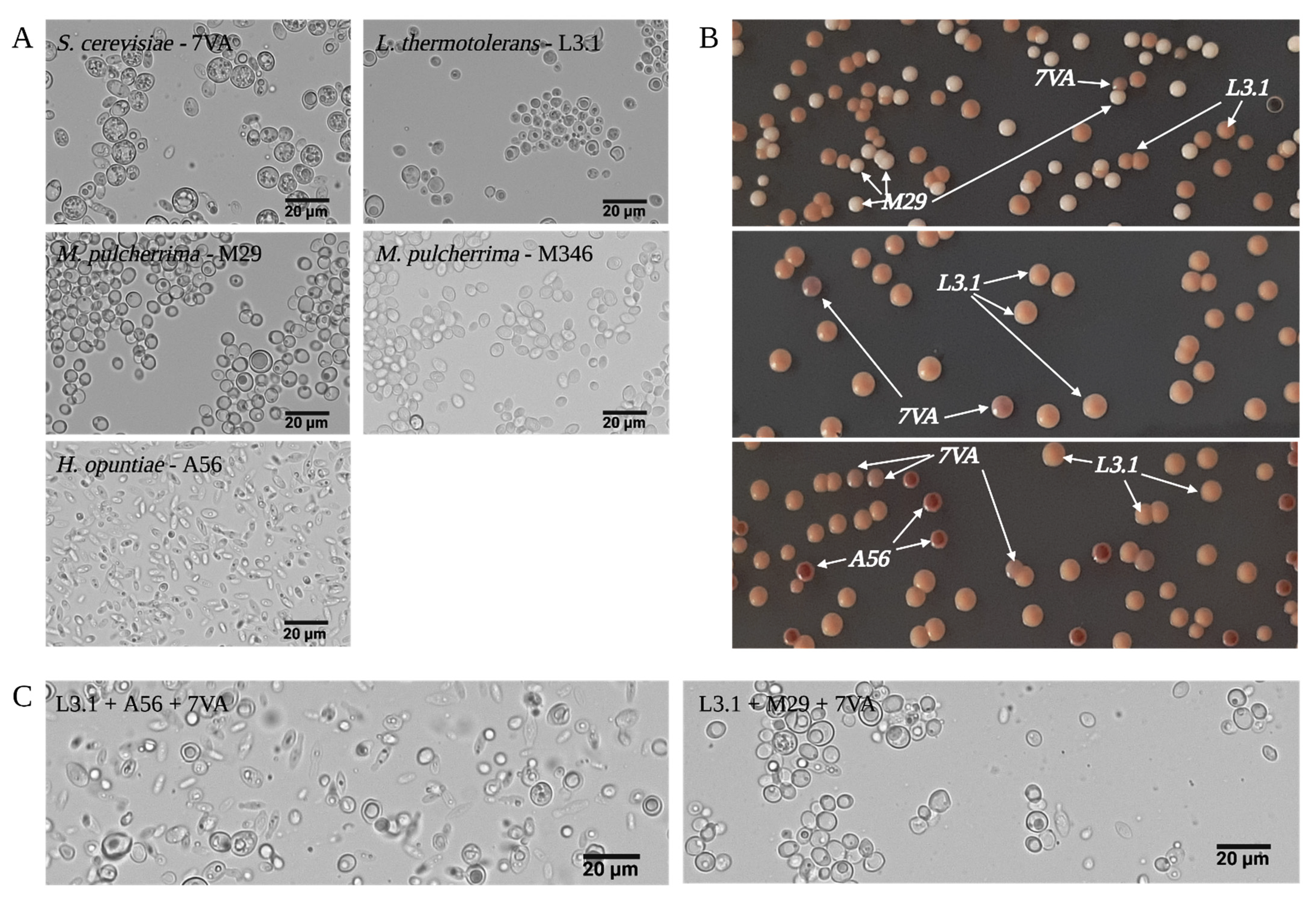
2.1. Yeast Strains and Growing Media
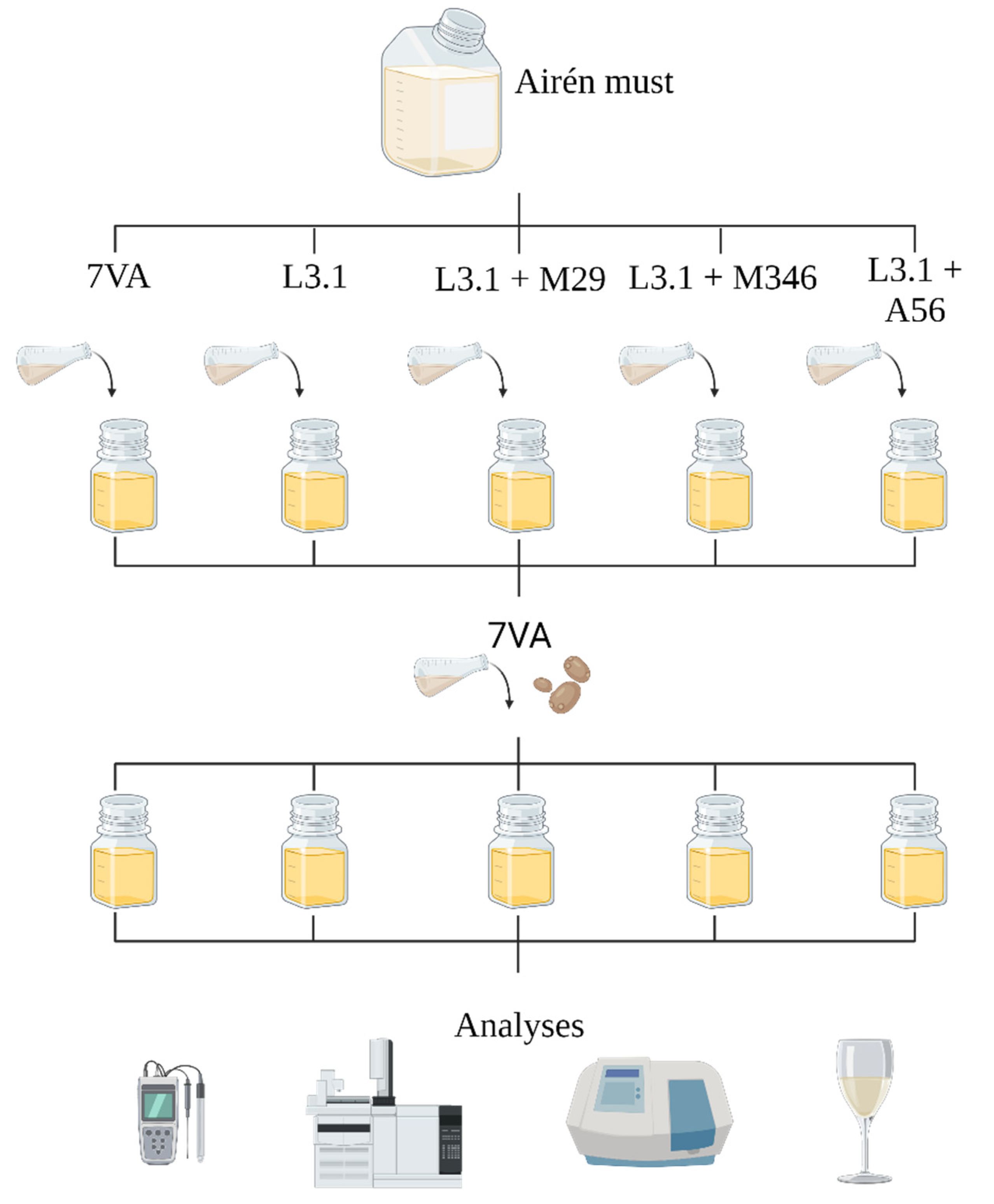
2.2. Must Fermentation
2.3. General Oenological Parameters
2.4. Aroma Volatile Compounds
2.5. Color Assessment
2.6. Sensory Evaluation
2.7. Statistical Analysis
3. Results and Discussion
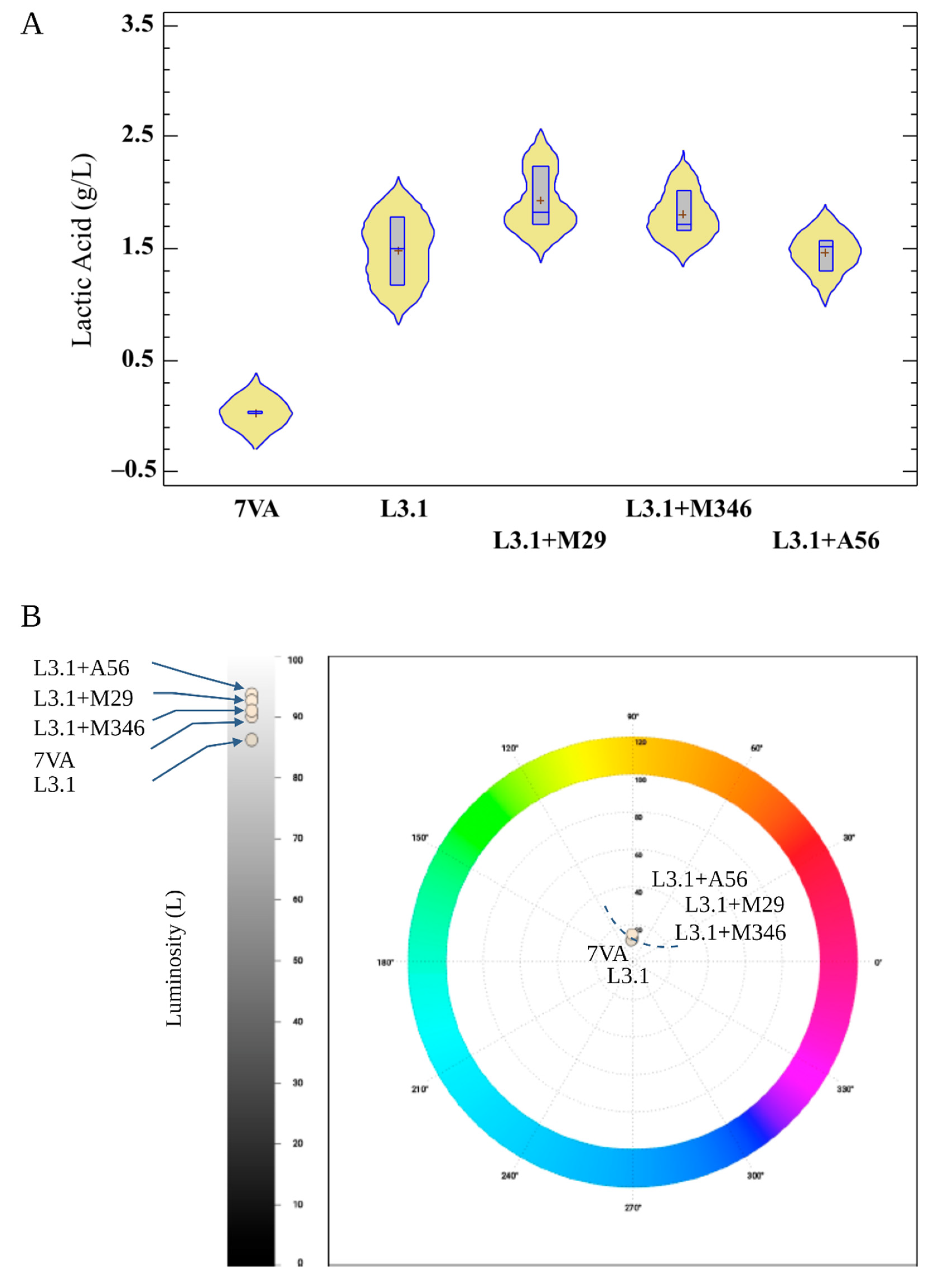
3.1. Wine Composition
3.2. Aromatic Profile
3.3. Color Assessment
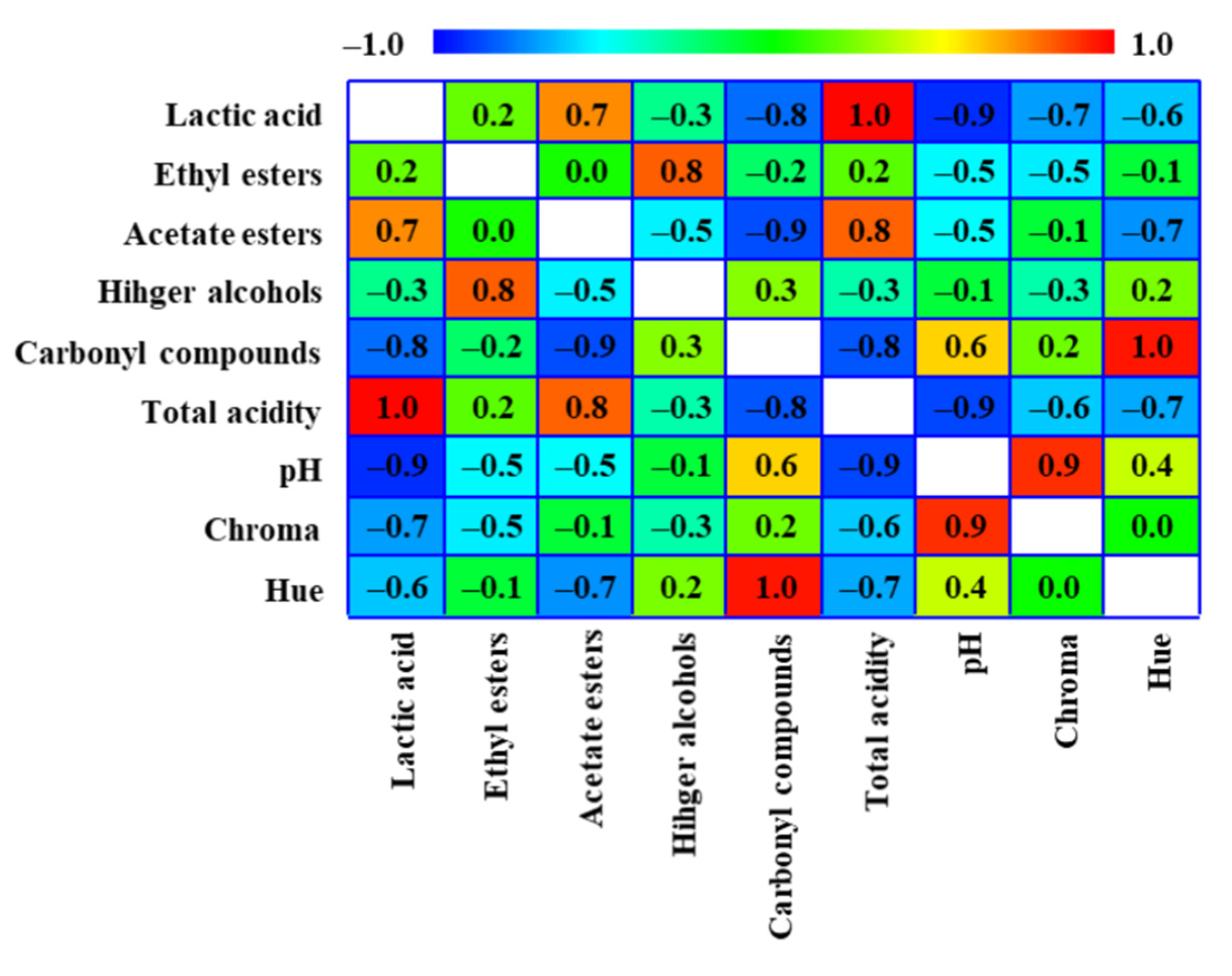
3.4. Variables Correlation
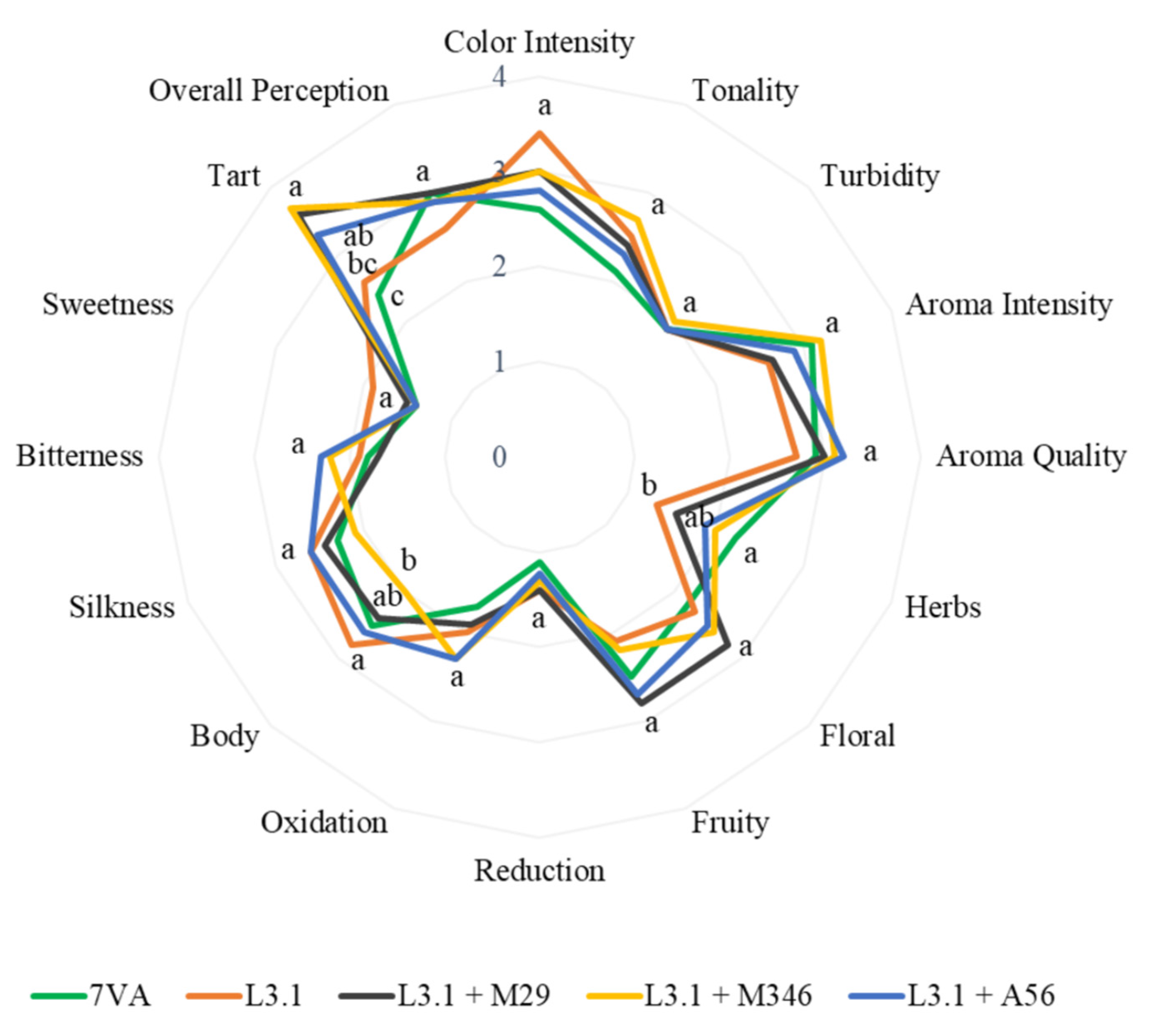
3.5. Wine Sensory Evaluation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Izquierdo Cañas, P.M.; Palacios García, A.T.; García Romero, E. Enhancement of flavour properties in wines using sequential inoculations of non-Saccharomyces (Hansenula and Torulaspora) and Saccharomyces yeast. VITIS J. Grapevine Res. 2011, 50, 177. [Google Scholar] [CrossRef]
- Escribano-Viana, R.; González-Arenzana, L.; Portu, J.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R. Wine aroma evolution throughout alcoholic fermentation sequentially inoculated with non-Saccharomyces/Saccharomyces yeasts. Food Res. Int. 2018, 112, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Bueno, J.E.; Peinado, R.A.; Medina, M.; Moreno, J. Effect of a short contact time with lees on volatile composition of Airen and Macabeo wines. Biotechnol. Lett. 2006, 28, 1007–1011. [Google Scholar] [CrossRef] [PubMed]
- Capozzi, V.; Garofalo, C.; Chiriatti, M.A.; Grieco, F.; Spano, G. Microbial terroir and food innovation: The case of yeast biodiversity in wine. Microbiol. Res. 2015, 181, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Medina, K.; Boido, E.; Fariña, L.; Gioia, O.; Gomez, M.E.; Barquet, M.; Gaggero, C.; Dellacassa, E.; Carrau, F. Increased flavour diversity of Chardonnay wines by spontaneous fermentation and co-fermentation with Hanseniaspora vineae. Food Chem. 2013, 141, 2513–2521. [Google Scholar] [CrossRef]
- Martin, V.; Valera, M.; Medina, K.; Boido, E.; Carrau, F. Oenological Impact of the Hanseniaspora/Kloeckera Yeast Genus on Wines—A Review. Fermentation 2018, 4, 76. [Google Scholar] [CrossRef] [Green Version]
- Hranilovic, A.; Gambetta, J.M.; Schmidtke, L.; Boss, P.K.; Grbin, P.R.; Masneuf-Pomarede, I.; Bely, M.; Albertin, W.; Jiranek, V. Oenological traits of Lachancea thermotolerans show signs of domestication and allopatric differentiation. Sci. Rep. 2018, 8, 14812. [Google Scholar] [CrossRef] [Green Version]
- Morata, A.; Escott, C.; Bañuelos, M.A.; Loira, I.; Del Fresno, J.M.; González, C.; Suárez-Lepe, J.A. Contribution of Non-Saccharomyces Yeasts to Wine Freshness. A Review. Biomolecules 2020, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, M.; Velázquez, R. The Yeast Torulaspora delbrueckii: An Interesting But Difficult-To-Use Tool for Winemaking. Fermentation 2018, 4, 94. [Google Scholar] [CrossRef] [Green Version]
- Loira, I.; Vejarano, R.; Bañuelos, M.A.; Morata, A.; Tesfaye, W.; Uthurry, C.; Villa, A.; Cintora, I.; Suárez-Lepe, J.A. Influence of sequential fermentation with Torulaspora delbrueckii and Saccharomyces cerevisiae on wine quality. LWT Food Sci. Technol. 2014, 59, 915–922. [Google Scholar] [CrossRef]
- Velázquez, R.; Zamora, E.; álvarez, M.L.; Hernández, L.M.; Ramírez, M. Effects of new Torulaspora delbrueckii killer yeasts on the must fermentation kinetics and aroma compounds of white table wine. Front. Microbiol. 2015, 6, 1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sipiczki, M. Metschnikowia strains isolated from botrytized grapes antagonize fungal and bacterial growth by iron depletion. Appl. Environ. Microbiol. 2006, 72, 6716–6724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oro, L.; Ciani, M.; Comitini, F. Antimicrobial activity of Metschnikowia pulcherrima on wine yeasts. J. Appl. Microbiol. 2014, 116, 1209–1217. [Google Scholar] [CrossRef] [PubMed]
- Zott, K.; Thibon, C.; Bely, M.; Lonvaud-Funel, A.; Dubourdieu, D.; Masneuf-Pomarede, I. The grape must non-Saccharomyces microbial community: Impact on volatile thiol release. Int. J. Food Microbiol. 2011, 151, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Loira, I.; Escott, C.; del Fresno, J.M.; Bañuelos, M.A.; Suárez-Lepe, J.A. Applications of Metschnikowia pulcherrima in Wine Biotechnology. Fermentation 2019, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- Rowan, N.J. Pulsed light as an emerging technology to cause disruption for food and adjacent industries—Quo vadis? Trends Food Sci. Technol. 2019, 88, 316–332. [Google Scholar] [CrossRef]
- Escott, C.; Vaquero, C.; del Fresno, J.M.; Bañuelos, M.A.; Loira, I.; Han, S.; Bi, Y.; Morata, A.; Suárez-Lepe, J.A. Pulsed light effect in red grape quality and fermentation. Food Bioprocess Technol. 2017, 10, 1540–1547. [Google Scholar] [CrossRef]
- Santamera, A.; Escott, C.; Loira, I.; Del Fresno, J.M.; González, C.; Morata, A. Pulsed light: Challenges of a non-thermal sanitation technology in the winemaking industry. Beverages 2020, 6, 45. [Google Scholar] [CrossRef]
- Abalos, D.; Vejarano, R.; Morata, A.; González, C.; Suárez-Lepe, J.A. The use of furfural as a metabolic inhibitor for reducing the alcohol content of model wines. Eur. Food Res. Technol. 2011, 232, 663–669. [Google Scholar] [CrossRef]
- Escribano-Viana, R.; González-Arenzana, L.; Garijo, P.; López, R.; Santamaría, P.; Gutiérrez, A.R. Selection Process of a Mixed Inoculum of Non-Saccharomyces Yeasts Isolated in the D.O.Ca. Rioja. Fermentation 2021, 7, 148. [Google Scholar] [CrossRef]
- Vaquero, C.; Loira, I.; Heras, J.M.; Carrau, F.; González, C.; Morata, A. Biocompatibility in Ternary Fermentations With Lachancea thermotolerans, Other Non-Saccharomyces and Saccharomyces cerevisiae to Control pH and Improve the Sensory Profile of Wines From Warm Areas. Front. Microbiol. 2021, 12, 656262. [Google Scholar] [CrossRef] [PubMed]
- Vaquero, C.; Loira, I.; Bañuelos, M.A.; Heras, J.M.; Cuerda, R.; Morata, A. Industrial Performance of Several Lachancea thermotolerans Strains for pH Control in White Wines from Warm Areas. Microorganisms 2020, 8, 830. [Google Scholar] [CrossRef] [PubMed]
- Hranilovic, A.; Albertin, W.; Capone, D.L.; Gallo, A.; Grbin, P.R.; Danner, L.; Bastian, S.E.P.; Masneuf-Pomarede, I.; Coulon, J.; Bely, M.; et al. Impact of Lachancea thermotolerans on chemical composition and sensory profiles of Merlot wines. Food Chem. 2021, 349, 129015. [Google Scholar] [CrossRef] [PubMed]
- Canonico, L.; Comitini, F.; Ciani, M. Metschnikowia pulcherrima selected strain for ethanol reduction in wine: Influence of cell immobilization and aeration condition. Foods 2019, 8, 378. [Google Scholar] [CrossRef] [Green Version]
- Castrillo, D.; Rabuñal, E.; Neira, N.; Blanco, P. Oenological potential of non-Saccharomyces yeasts to mitigate effects of climate change in winemaking: Impact on aroma and sensory profiles of Treixadura wines. FEMS Yeast Res. 2019, 19, 65. [Google Scholar] [CrossRef]
- Porter, T.J.; Divol, B.; Setati, M.E. Lachancea yeast species: Origin, biochemical characteristics and oenological significance. Food Res. Int. 2019, 119, 378–389. [Google Scholar] [CrossRef]
- Escribano-Viana, R.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R.; González-Arenzana, L. Do Non-Saccharomyces Yeasts Work Equally with Three Different Red Grape Varieties? Fermentation 2019, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Gore-Lloyd, D.; Sumann, I.; Brachmann, A.O.; Schneeberger, K.; Ortiz-Merino, R.A.; Moreno-Beltrán, M.; Schläfli, M.; Kirner, P.; Santos Kron, A.; Rueda-Mejia, M.P.; et al. Snf2 controls pulcherriminic acid biosynthesis and antifungal activity of the biocontrol yeast Metschnikowia pulcherrima. Mol. Microbiol. 2019, 112, 317–332. [Google Scholar] [CrossRef] [Green Version]
- Peinado, R.; Moreno, J.; Medina, M.; Mauricio, J. Changes in volatile compounds and aromatic series in sherry wine with high gluconic acid levels subjected to aging by submerged flor yeast cultures. Biotechnol. Lett. 2004, 26, 757–762. [Google Scholar] [CrossRef]
- Li, H.; Guo, A.; Wang, H. Mechanisms of oxidative browning of wine. Food Chem. 2008, 108, 1–13. [Google Scholar] [CrossRef]
- Medina-Plaza, C.; García-Cabezón, C.; García-Hernández, C.; Bramorski, C.; Blanco-Val, Y.; Martín-Pedrosa, F.; Kawai, T.; de Saja, J.A.; Rodríguez-Méndez, M.L. Analysis of organic acids and phenols of interest in the wine industry using Langmuir–Blodgett films based on functionalized nanoparticles. Anal. Chim. Acta 2015, 853, 572–578. [Google Scholar] [CrossRef]
- Mierczynska-Vasilev, A.; Bindon, K.; Gawel, R.; Smith, P.; Vasilev, K.; Butt, H.J.; Hans, J.; Koynov, K. Fluorescence correlation spectroscopy to unravel the interactions between macromolecules in wine. Food Chem. 2021, 352, 129343. [Google Scholar] [CrossRef] [PubMed]
- Blanco, P.; Rabuñal, E.; Neira, N.; Castrillo, D. Dynamic of Lachancea thermotolerans Population in Monoculture and Mixed Fermentations: Impact on Wine Characteristics. Beverages 2020, 6, 36. [Google Scholar] [CrossRef]
- Vilela, A. Lachancea thermotolerans, the Non-Saccharomyces yeast that reduces the volatile acidity of wines. Fermentation 2018, 4, 56. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 7VA | L3.1 | L3.1 + M29 | L3.1 + M346 | L3.1 + A56 | |
|---|---|---|---|---|---|
| Ethanol (% v/v) | 12.6 ± 0.1a | 9.6 ± 0.1d | 11.9 ± 0.1c | 12.1 ± 0.1b | 12.1 ± 0.0bc |
| Glucose and Fructose (g/L) | 1.9 ± 0.02c | 2.0 ± 0.01b | 2.3 ± 0.02a | 1.5 ± 0.02d | 2.0 ± 0.01b |
| Lactic acid (g/L) | 0.0 + 0.01b | 1.5 + 0.31a | 1.9 + 0.28a | 1.8 + 0.28a | 1.5 + 0.15a |
| Acetic acid (g/L) | 0.4 ± 0.01a | 0.3 ± 0.01b | 0.2 ± 0.00d | 0.2 ± 0.00e | 0.2 ± 0.01c |
| Glycerol (g/L) | 5.1 ± 0.1a | 5.1 ± 0.2a | 5.4 ± 0.1a | 5.1 ± 0.1a | 5.4 ± 0.3a |
| Total Acidity 1 (g/L) | 3.7 ± 0.0d | 4.5 ± 0.0b | 4.8 ± 0.1a | 4.9 ± 0.1a | 4.3 ± 0.0c |
| pH | 3.8 ± 0.01a | 3.6 ± 0.02b | 3.6 ± 0.02b | 3.6 ± 0.01b | 3.6 ± 0.02b |
| Methanol (mg/L) | 125.8 ± 3.7ab | 114.9 ± 14.5bc | 100.4 ± 2.3d | 104.8 ± 3.4cd | 129.7 ± 4.1a |
| Ethyl esters (mg/L) | 1.5 ± 0.3c | 23.0 ± 9.7b | 42.4 ± 10.3a | 38.7 ± 10.7a | 9.5 ± 2.0bc |
| Acetate esters (mg/L) | 18.4 ± 4.8d | 40.7 ± 2.6bc | 41.7 ± 5.9ab | 33.7 ± 3.2c | 48.2 ± 2.5a |
| Higher alcohols (mg/L) | 148.2 ± 2.4c | 230.8 ± 9.0a | 214.1 ± 6.2b | 218.9 ± 9.8ab | 220.8 ± 6.1ab |
| Carbonyl compounds 2 (mg/L) | 64.8 ± 2.5a | 33.6 ± 6.0c | 41.5 ± 1.5c | 33.5 ± 2.5b | 38.5 ± 5.9bc |
| Total Polyphenols Index | 9.3 ± 0.1d | 17.4 ± 0.1a | 11.6 ± 0.1c | 11.5 ± 0.1c | 13.5 ± 0.1b |
| Colour Intensity 3 | 0.41 ± 0.00b | 0.52 ± 0.06a | 0.34 ± 0.00c | 0.42 ± 0.00b | 0.30 ± 0.00c |
| Chroma 4 | 12.7 ± 0.1a | 9.4 ± 0.1e | 10.8 ± 0.0b | 10.1 ± 0.0d | 10.4 ± 0.0c |
| Hue 4 (º) | 86.5 ± 0.01a | 85.4 ± 0.02b | 83.1 ± 0.02d | 83.1 ± 0.01d | 83.6 ± 0.02c |
| Luminosity 4 (L) | 90.6 ± 0.1d | 86.8 ± 0.1e | 93.1 ± 0.1b | 91.5 ± 0.1c | 94.1 ± 0.1a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escott, C.; Vaquero, C.; Loira, I.; López, C.; González, C.; Morata, A. Synergetic Effect of Metschnikowia pulcherrima and Lachancea thermotolerans in Acidification and Aroma Compounds in Airén Wines. Foods 2022, 11, 3734. https://doi.org/10.3390/foods11223734
Escott C, Vaquero C, Loira I, López C, González C, Morata A. Synergetic Effect of Metschnikowia pulcherrima and Lachancea thermotolerans in Acidification and Aroma Compounds in Airén Wines. Foods. 2022; 11(22):3734. https://doi.org/10.3390/foods11223734
Chicago/Turabian StyleEscott, Carlos, Cristian Vaquero, Iris Loira, Carmen López, Carmen González, and Antonio Morata. 2022. "Synergetic Effect of Metschnikowia pulcherrima and Lachancea thermotolerans in Acidification and Aroma Compounds in Airén Wines" Foods 11, no. 22: 3734. https://doi.org/10.3390/foods11223734