
Assessment of the Bioaccessibility of Carotenoids in Goji Berry (Lycium barbarum L.) in Three Forms: In Vitro Digestion Model and Metabolomics Approach
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Sample Preparation
2.2. Chemicals and Reagents
2.3. Surface Color Measurement
2.4. Observation of the Epidermis Cells of LBL
2.5. Analysis of Carotenoid by High Resolution LC−Q−TOF−MS/MS
2.6. Differential Metabolite Analysis
2.7. In Vitro Digestion Model
2.8. Quantification of Bioaccessible Carotenoids
2.8.1. High−Performance Liquid Chromatography (HPLC)
2.8.2. Bioaccessibility Calculation
2.9. Determination of Total Carotenoids in Digestive Fluid
2.10. Determination of Antioxidant Activity
2.10.1. DPPH Free Radical Scavenging Capacity Assay
2.10.2. FRAP Iron Ion Reduction Capacity Measurement
2.10.3. ABTS Free Radical Scavenging Capacity Assay
2.11. Statistical Analysis
3. Results and Discussion
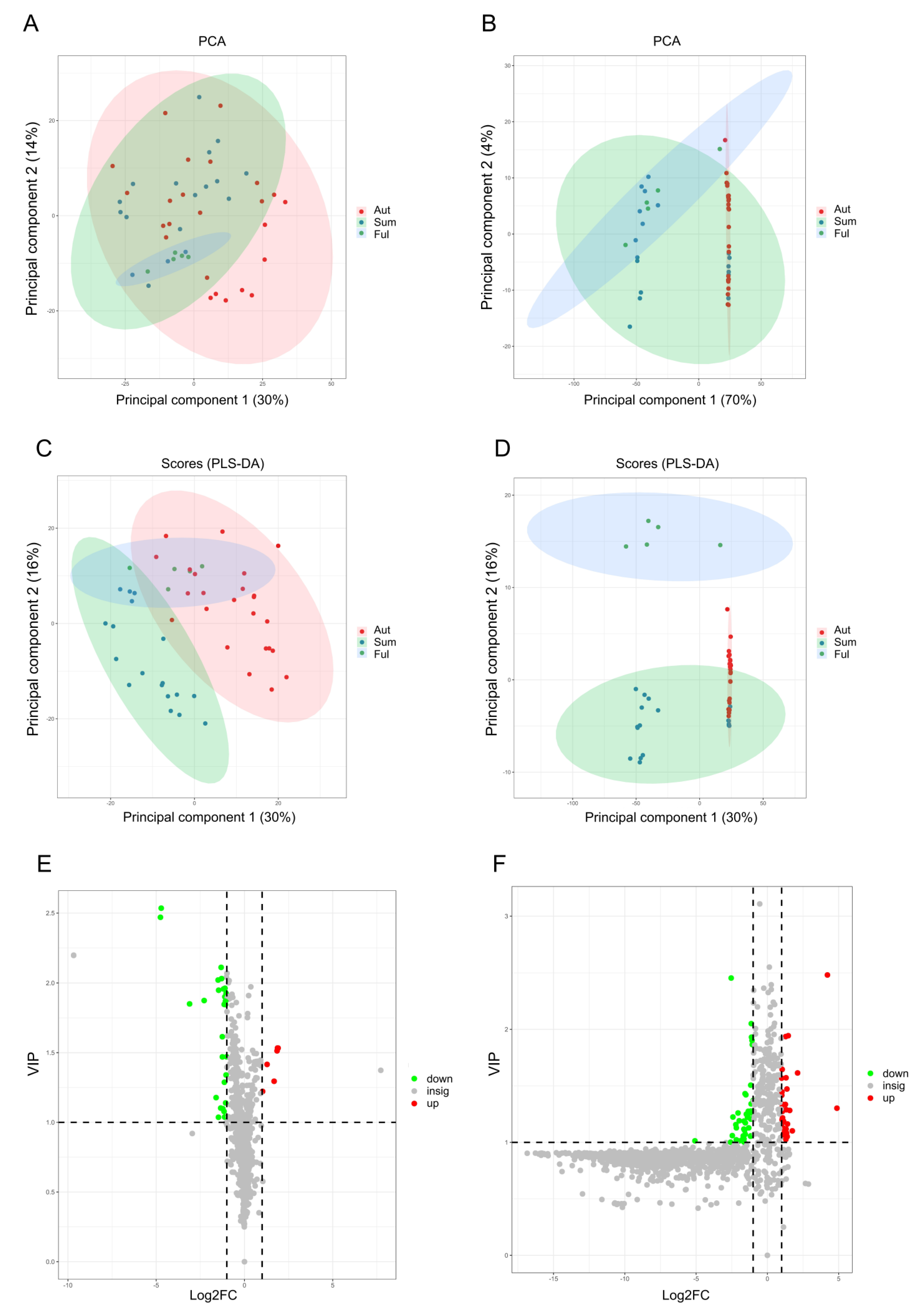
3.1. Screening and Identification of Carotenoids
3.2. Metabolism Profile of Carotenoids
3.3. Color and Cell Structure Changes of LBL
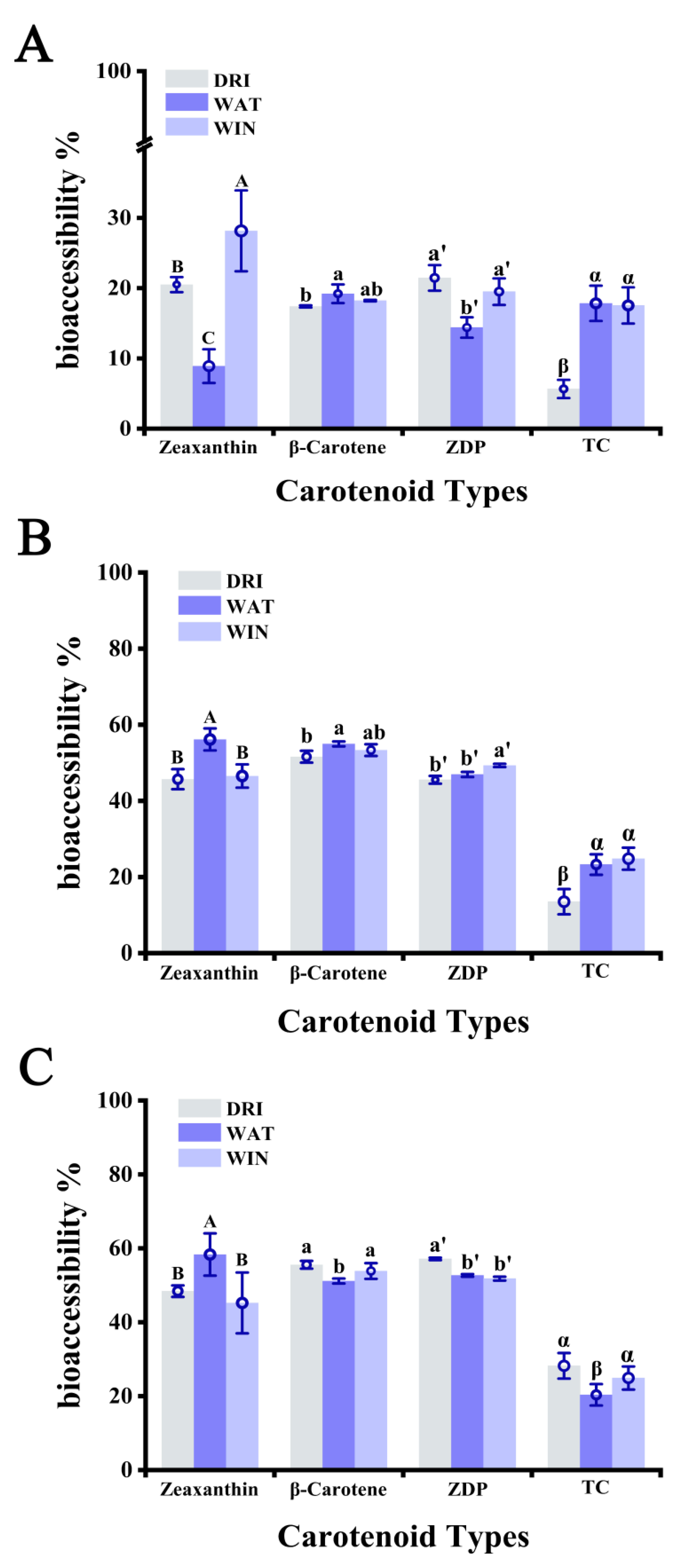
3.4. Bioaccessibility of Carotenoids
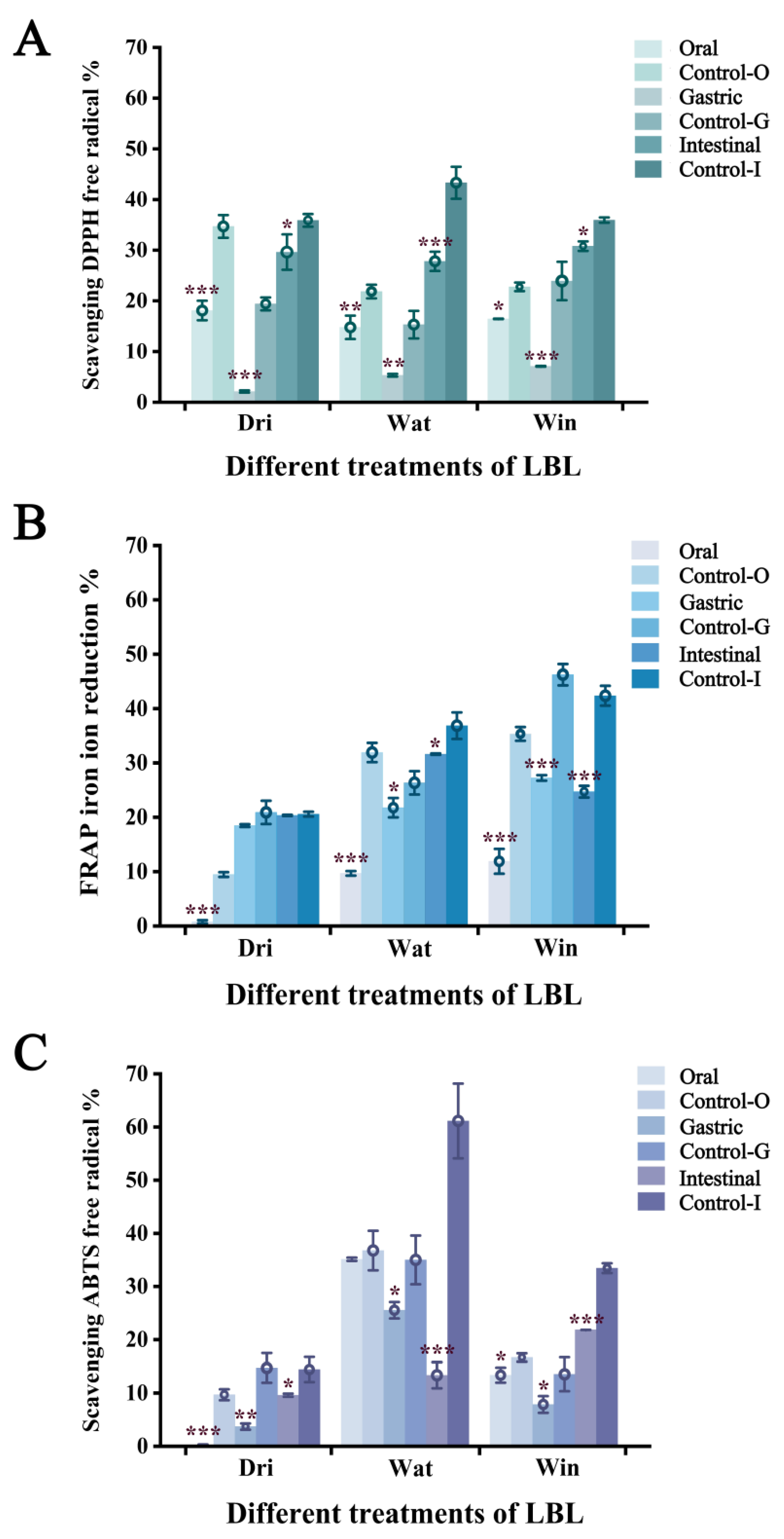
3.5. Antioxidant Activity of Carotenoids in Digestive Fluid
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mocan, C.; Moldovan, G.; Zengin, O.; Bender, M.; Locatelli, M.; Simirgiotis, A.; Atalay, D.C.; Vodnar, S.; Rohn, G. UHPLC-QTOF-MS analysis of bioactive constituents from two Romanian Goji (Lycium barbarum L.) berries cultivars and their antioxidant, enzyme inhibitory, and real-time cytotoxicological evaluation. Food Chem. Toxicol. 2018, 115, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Chen, G.J.; Yang, B.; Wu, Y.; Du, M.Y.; Kan, J.Q. Insights into the stability of carotenoids and capsaicinoids in water-based or oil-based chili systems at different processing treatments. Food Chem. 2021, 342, 128308. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R. Physicochemical, antioxidant properties of carotenoids and its optoelectronic and interaction studies with chlorophyll pigments. Sci. Rep. 2021, 11, 18365. [Google Scholar] [CrossRef]
- Edwards, C.H.; Ryden, P.; Mandalari, G.; Butterworth, P.J.; Ellis, P.R. Structure-function studies of chickpea and durum wheat uncover mechanisms by which cell wall properties influence starch bioaccessibility. Nat. Food 2021, 2, 118. [Google Scholar] [CrossRef] [PubMed]
- Widomska, J.; Paul Sangiovanni, J.; Subczynski, W.K. Why is zeaxanthin the most concentrated xanthophyll in the central fovea? Nutrients 2020, 12, 1333. [Google Scholar] [CrossRef]
- Hempel, J.; Schdle, C.N.; Sprenger, J.; Heller, A.; Schweiggert, R.M. Ultrastructural deposition forms and bioaccessibility of carotenoids and carotenoid esters from goji berries (Lycium barbarum L.). Food Chem. 2017, 218, 525–533. [Google Scholar] [CrossRef]
- Kan, X.H.; Yan, Y.M.; Ran, L.; Lu, W.L.; Mi, J.; Zhang, Z.J.; Li, X.Y.; Zeng, X.X.; Cao, Y.L. Ultrasonic-assisted extraction and high-speed counter-current chromatography purification of zeaxanthin dipalmitate from the fruits of Lycium barbarum L. Food Chem. 2020, 310, 125854. [Google Scholar] [CrossRef] [PubMed]
- Fratianni, A.; Niro, S.; Alam, M.D.R.; Cinquanta, L.; Matteo, M.D.; Adiletta, G.; Panfili, G. Effect of a physical pre-treatment and drying on carotenoids of goji berries (Lycium barbarum L.). LWT Food Sci. Technol. 2018, 92, 318–323. [Google Scholar] [CrossRef]
- Chen, X.; Gao, H.; Chen, Z.; Li, T.; Zhang, Z.; Yun, Z.; Jiang, Y. Metabolic variations in the pulp of four litchi cultivars during pulp breakdown. Food Res. Int. 2021, 140, 110080. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Kunze, D.; Shixin, H.; Rui, Y.; Lina, Q.; Jin, L.; Yanxu, C. A comprehensive strategy integrating metabolomics with multiple chemometric for discovery of function related active markers for assessment of foodstuffs: A case of hawthorn (Crataegus cuneata) fruits. Food Chem. 2022, 383, 132464. [Google Scholar]
- Ze, Y.; Huijun, G.; Yueming, J. Insights into metabolomics in quality attributes of postharvest fruit. Biomolecules 2022, 45, 100836. [Google Scholar]
- Hu, S.; Liu, C.; Liu, X. Innovative Application of Metabolomics on Bioactive Ingredients of Foods. Foods 2022, 11, 2974. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ming, R.; Yao, S.; Li, L.; Huang, R.; Tan, Y. Identification of anthocyanins in the fruits of Kadsura coccinea using UPLC-MS/MS-based metabolomics. Biochem. Syst. Ecol. 2021, 98, 104324. [Google Scholar] [CrossRef]
- Feng, L.; Tang, N.; Liu, R.; Nie, R.; Guo, Y.; Liu, R.; Chang, M. Effects of different processing methods on bioactive substances and antioxidation properties of Lycium barbarum (goji berry) from China. Food Biosci. 2021, 42, 101048. [Google Scholar] [CrossRef]
- Ropelewska, E.; Sabanci, K.; Aslan, M.F. The Changes in Bell Pepper Flesh as a Result of Lacto-Fermentation Evaluated Using Image Features and Machine Learning. Foods 2022, 11, 2956. [Google Scholar] [CrossRef]
- Zou, S.C.; Wu, J.C.; Shahid, M.Q.; He, Y.H.; Lin, S.Q.; Liu, Z.H.; Yang, X.H. Identification of key taste components in loquat using widely targeted metabolomics. Food Chem. 2020, 323, 126822. [Google Scholar] [CrossRef]
- Becerra-Martínez, E.; Florentino-Ramos, E.; Pérez-Hernández, N.; Zepeda-Vallejo, G.; Villa-Ruano, L.N.; Velázquez-Ponce, M. 1H NMR-based metabolomic fingerprinting to determine metabolite levels in serrano peppers (Capsicum annum L.) grown in two different regions. Food Res. Int. 2017, 102, 163–170. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Lebensm. Wiss. Technol. Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free. Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- De Rosso, V.V.; Mercadante, A.Z. Identifification and quantifification of carotenoids, by HPLC-PDA-MS/MS from Amazonian fruits. J. Agric. Food Chem. 2007, 55, 5062–5072. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, J.U.; Wahl, S.; Wurschum, T.; Longin, C.F.H.; Carle, R.; Schweiggert, R.M. Lutein and lutein esters in whole grain flours made from 75 genotypes of 5 Triticum species grown at multiple sites. J. Agric. Food Chem. 2015, 63, 5061–5071. [Google Scholar] [CrossRef]
- Van Breemen, R.B.; Dong, L.; Pajkovic, N.D. Atmospheric pressure chemicalionization tandem mass spectrometry of carotenoids. Int. J. Mass Spectrom. 2012, 312, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Xiaogang, H.; Xiang, H.; Jiaping, Y.; Yulong, F.; Ganghui, C. Rapid prediction method of α-Glycosidase inhibitory activity of Coreopsis tinctoria extract from different habitats by near infrared spectroscopy. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2022, 268, 120601. [Google Scholar]
- Magwaza, S.L.; Naidoo, S.I.M.; Laurie, S.M.; Laing, M.D.; Shimelis, H. Development of NIRS models for rapid quantification of protein content in sweetpotato [Ipomoea batatas (L.) LAM.]. LWT Food Sci. Technol. 2016, 72, 63–70. [Google Scholar] [CrossRef]
- Ma, W.-P.; Ni, Z.-J.; Li, H.; Chen, M. Changes of the main carotenoid pigment contents during the drying processes of the different harvest stage fruits of Lycium barbarum L. Agric. Sci. China 2008, 7, 363–369. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, Y.H.; Xu, H.; Zhao, X.L.; Tan, Y.T.; Li, P.P.; Li, D.D.; Tao, Y.M.; Liu, D.H. Fruit softening correlates with enzymatic activities and compositional changes in fruit cell wall during growing in Lycium barbarum L. Int. J. Food Sci. Technol. 2021, 56, 3044–3054. [Google Scholar] [CrossRef]
- Cristina, J.; Domenico, M.; Jordi, M.; Ana, J.-G. Carotenoids present in goji berries Lycium barbarum L. are suitable to protect against mycotoxins effects: An in vitro study of bioavailability. J. Funct. Foods 2022, 92, 105049. [Google Scholar]
- Zaripheh, S.; Erdman, J.W. Factors that influence the bioavailablity of xanthophylls. J. Nutr. 2002, 132, 531S–534S. [Google Scholar] [CrossRef] [Green Version]
- Bouyahya, A.; El Omari, N.; Hakkur, M.; El Hachlafi, N.; Charfi, S.; Balahbib, A.; Guaouguaou, F.-E.; Bakrim, S. Sources, health benefits, and biological properties of zeaxanthin. Trends Food Sci. Technol. 2021, 118, 519–538. [Google Scholar] [CrossRef]
- Marchetti, N.; Bonetti, G.; Brandolini, V.; Cavazzini, A.; Maietti, A.; Meca, G.; Manes, J. Stinging nettle (Urtica dioica L.) as a functional food additive in egg pasta: Enrichment and bioaccessibility of Lutein and beta-carotene. J. Funct. Foods 2018, 47, 547–553. [Google Scholar] [CrossRef]
- Mapelli-Brahm, P.; Barba, F.J.; Remize, F.; Garcia, C.; Fessard, A.; Mousavi Khaneghah, A.; Sant’Ana, A.S.; Melendez-Martinez, A.J. The impact of fermentation processes on the production, retention and bioavailability of carotenoids: An overview. Trends Food Sci. Technol. 2020, 99, 389–401. [Google Scholar] [CrossRef]
- Gouado, I.; Schweigert, F.J.; Ejoh, R.A.; Tchouanguep, M.F.; Camp, J.V. Systemic levels of carotenoids from mangoes and papaya consumed in three forms (juice, fresh and dry slice). Eur. J. Clin. Nutr. 2007, 61, 1180–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, L.; Roberta, R.H.; Carl, L.; Keen, L.; Morse, S.; Glenn, Y.; Robert, M.H. Goji berry intake increases macular pigment optical density in healthy adults: A randomized pilot trial. Nutrients 2021, 13, 4409. [Google Scholar]
- Yonekura, L.; Nagao, A. Intestinal absorption of dietary carotenoids. Mol. Nutr. Food Res. 2007, 51, 107–115. [Google Scholar] [CrossRef]
- Huo, T.; Ferruzzi, M.G.; Schwartz, S.J.; Failla, M.L. Impact of fatty acyl composition and quantity of triglycerides on bioaccessibility of dietary carotenoids. J. Agric. Food Chem. 2007, 55, 8950–8957. [Google Scholar] [CrossRef] [Green Version]
- Colle, I.J.P.; Lemmens, L.; Van Buggenhout, S.; Met, K.; Van Loey, A.M.; Hendrickx, M.E. Processing tomato pulp in the presence of lipids: The impact on lycopene bioaccessibility. Food Res. Int. 2013, 51, 32–38. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carotenoids | m/z | Rt (Min) | Ionic Adducts | Secondary Fragments | Ionization Mode |
|---|---|---|---|---|---|
| New yellow mass | 601.4 | 3.43 | [M+H]+ | 583/565/491/393 | APCI+ |
| 13− or −13−(cis)−Zeaxanthin | 569.4 | 5.74 | [M+H]+ | 469/533/551 | APCI+/ESI+ |
| All−trans−zeaxanthin | 569.4 | 5.77 | [M+H]+ | 551/533/543/469 | APCI+/ESI+ |
| 9− or 9−(cis)−Zeaxanthin | 569.4 | 5.79 | [M+H]+ | 551/533 | APCI+ |
| β−Cryptophanin | 535.4 | 10.49 | [M+H−H2O]+ | 461/497/535 | APCI+ |
| α−Carotene | 537.3 | 13.56 | [M+H]+ | 135/331/399 | APCI+ |
| 9− or 9−(cis)−β−carotene | 537.4 | 14.78 | [M+H]+ | 445/444/413 | APCI+ |
| All−trans−β−carotene | 537.4 | 15.62 | [M+H]+ | 444/413 | APCI+/ESI+ |
| β−Cryptoxanthin monopalmitate | 791.6 | 45.32 | [M+H]+ | 790 | APCI+ |
| Zeaxanthin monopalmitate | 1062 | 46.93 | [M+FA−H]− | 551 | APCI+/ESI+ |
| Zeaxanthin dipalmitate | 1300.1 | 48.76 | [M+FA−H]− | 533/789 | APCI+/ESI+ |
| Zeaxanthin glucomannan | 965.72 | 8.48 | [M+H−H2O]+ | ESI+ | |
| 1′−Hydroxy−γ−carotenoid glucomannan | 697.48 | 4.82 | [M−H2O−H]+ | 279/78/97 | ESI+ |
| Treatments | Parameters | |||
|---|---|---|---|---|
| L* | a* | b* | C* | |
| DRI | 42.63 ± 1.89 a | 46.62 ± 1.01 a | 31.72 ± 2.00 a | 56.38 ± 1.54 a |
| WAT | 42.80 ± 1.31 a | 39.97 ± 3.01 b | 30.32 ± 1.74 a | 50.16 ± 3.37 b |
| WIN | 39.96 ± 1.14 a | 30.28 ± 1.54 c | 19.01 ± 2.08 b | 35.75 ± 2.19 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Z.; Ma, Y.; Liu, J.; Fan, Y.; Zheng, A.; Gao, P.; Wang, L.; Liu, D. Assessment of the Bioaccessibility of Carotenoids in Goji Berry (Lycium barbarum L.) in Three Forms: In Vitro Digestion Model and Metabolomics Approach. Foods 2022, 11, 3731. https://doi.org/10.3390/foods11223731
Hu Z, Ma Y, Liu J, Fan Y, Zheng A, Gao P, Wang L, Liu D. Assessment of the Bioaccessibility of Carotenoids in Goji Berry (Lycium barbarum L.) in Three Forms: In Vitro Digestion Model and Metabolomics Approach. Foods. 2022; 11(22):3731. https://doi.org/10.3390/foods11223731
Chicago/Turabian StyleHu, Ziying, Yanan Ma, Jun Liu, Yijun Fan, Anran Zheng, Pengyan Gao, Liang Wang, and Dunhua Liu. 2022. "Assessment of the Bioaccessibility of Carotenoids in Goji Berry (Lycium barbarum L.) in Three Forms: In Vitro Digestion Model and Metabolomics Approach" Foods 11, no. 22: 3731. https://doi.org/10.3390/foods11223731