
Enhancing the Nutritional and Functional Properties of Auricularia auricula through the Exploitation of Walnut Branch Waste
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Fungal Mycelium Growth Media
2.3. The Preparation of Liquid Strains and Mushroom Cultivation Process
2.4. Determination of Nutrient Elements in Walnut and Miscellaneous Sawdust
2.5. Determination of Extracellular Enzyme Activity
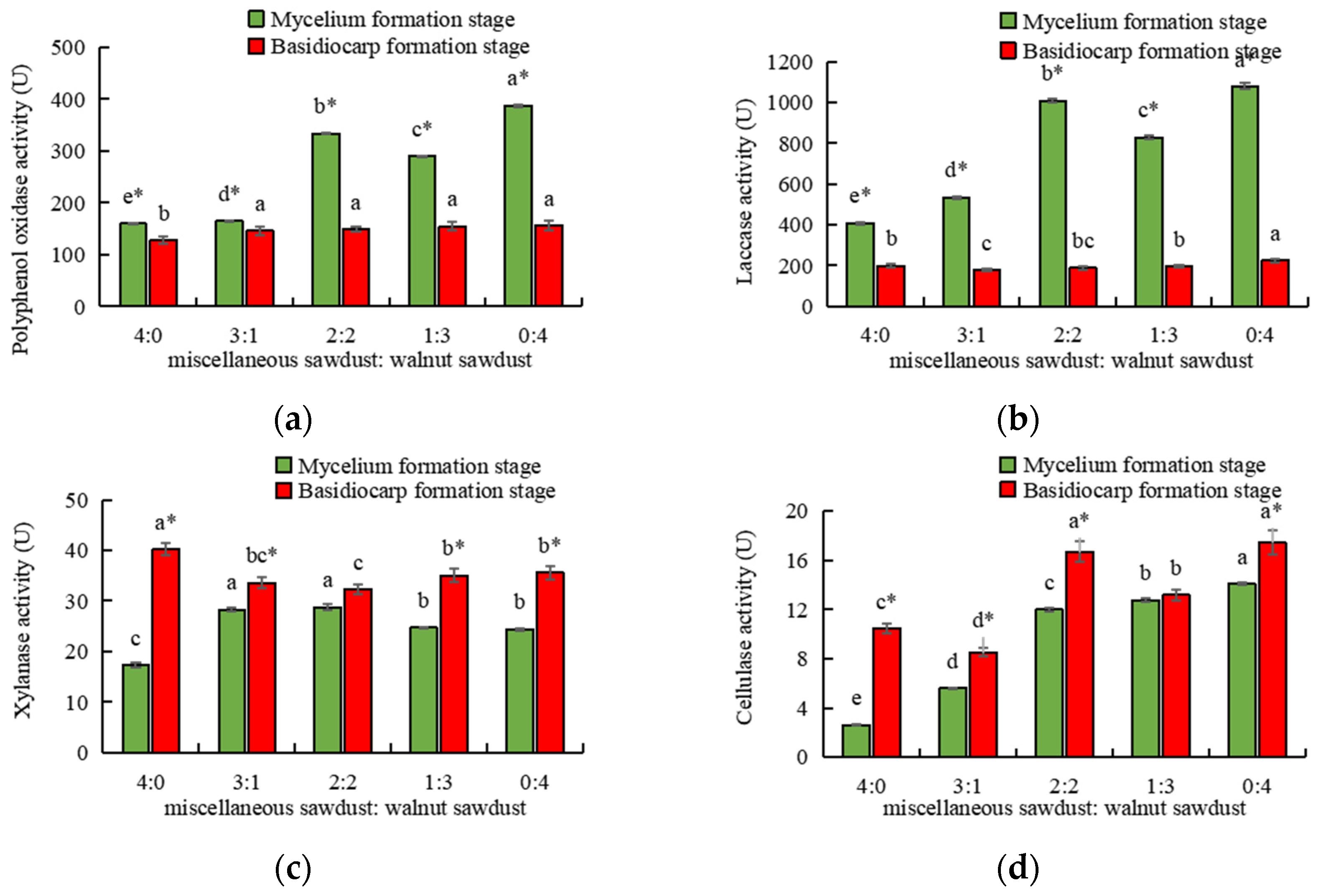
2.5.1. Determination of Laccase Activity
2.5.2. Determination of Polyphenol Oxidase Activity
2.5.3. Determination of Cellulase Activity
2.5.4. Determination of Xylanase Activity
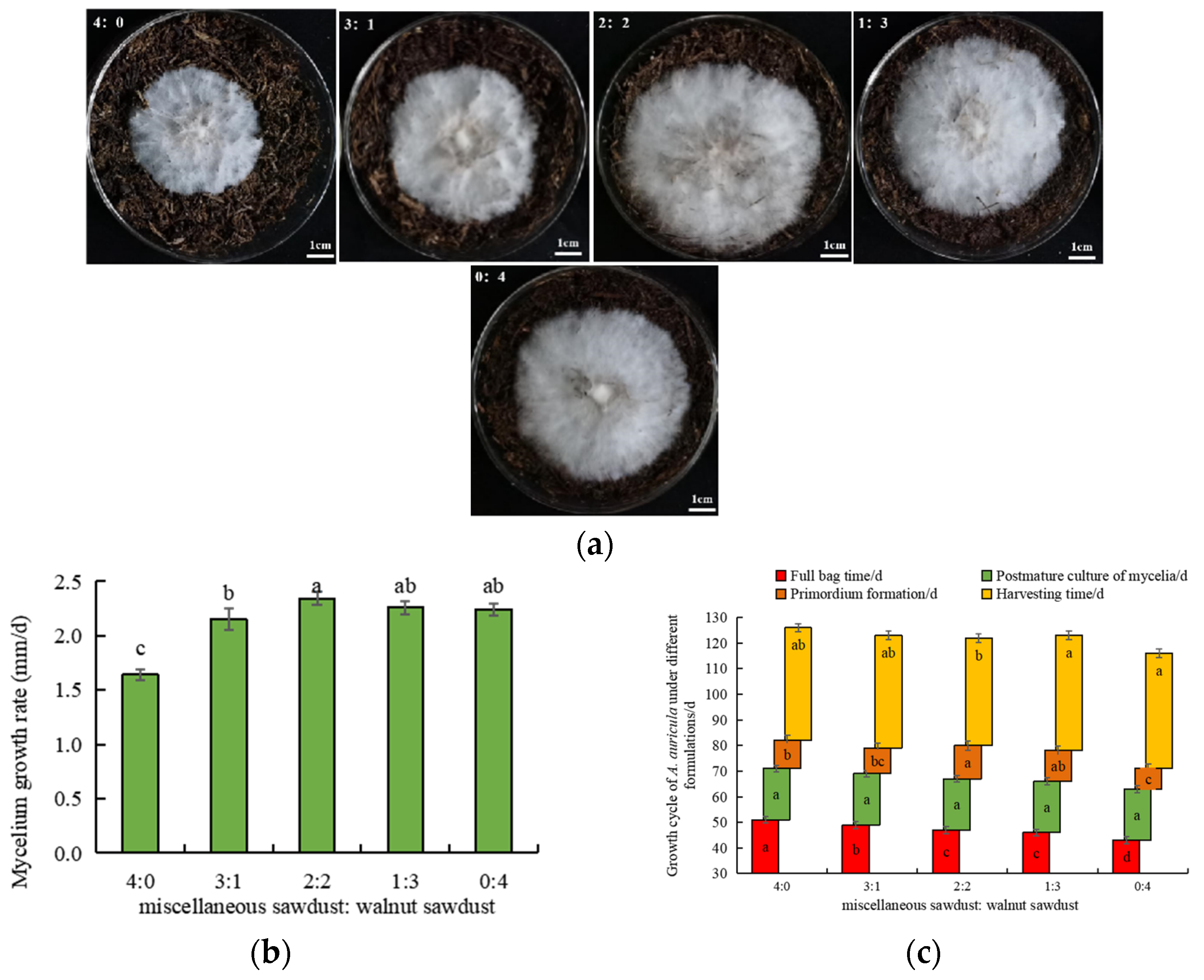
2.6. The Growth Evaluation of A. auricula
2.7. Determination of Production and Cultivation Benefit of A. auricula
2.8. Determination of Nutrient Elements in Basidiocarp of A. auricula
2.9. Comprehensive Evaluation of A. auricula Cultivated in Different Walnut Sawdust Ratio
2.10. Statistical Treatment of Experimental Data
3. Results and Discussion
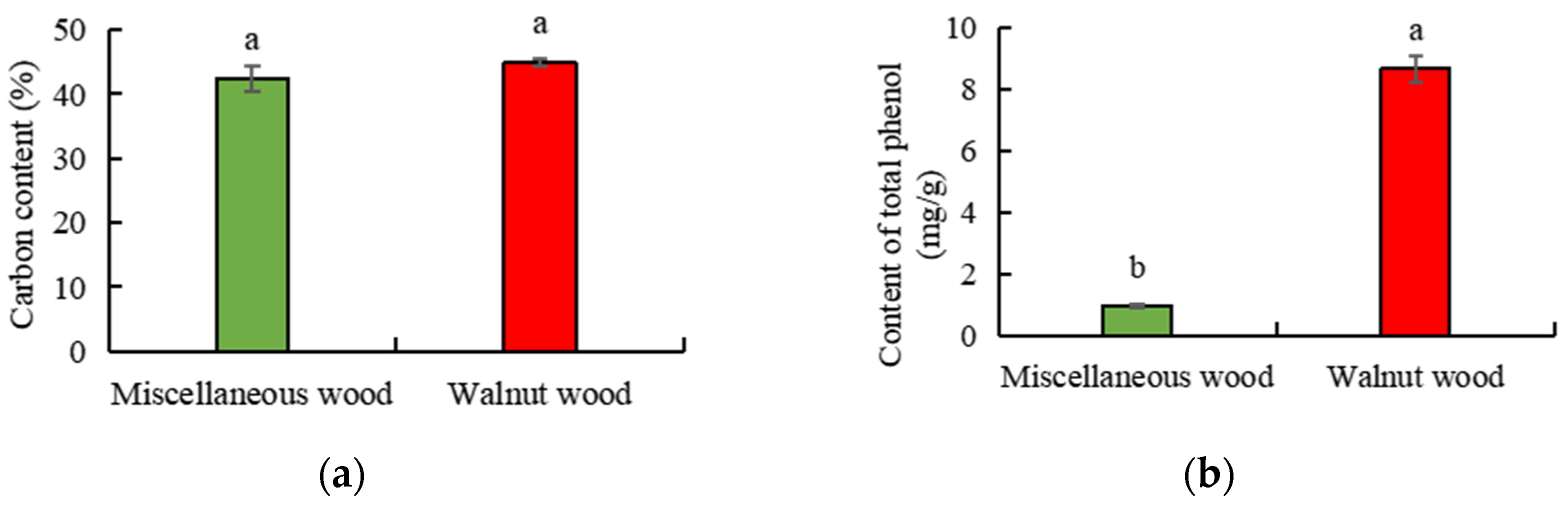
3.1. Comparison of Nutritional Components of Walnut and Miscellaneous Sawdust
3.2. The Effect of Walnut Sawdust Ratio on the Extracellular Enzyme Activities
3.3. Effect of Walnut Sawdust Addition Ratio on the Growth Cycle of A. auricula
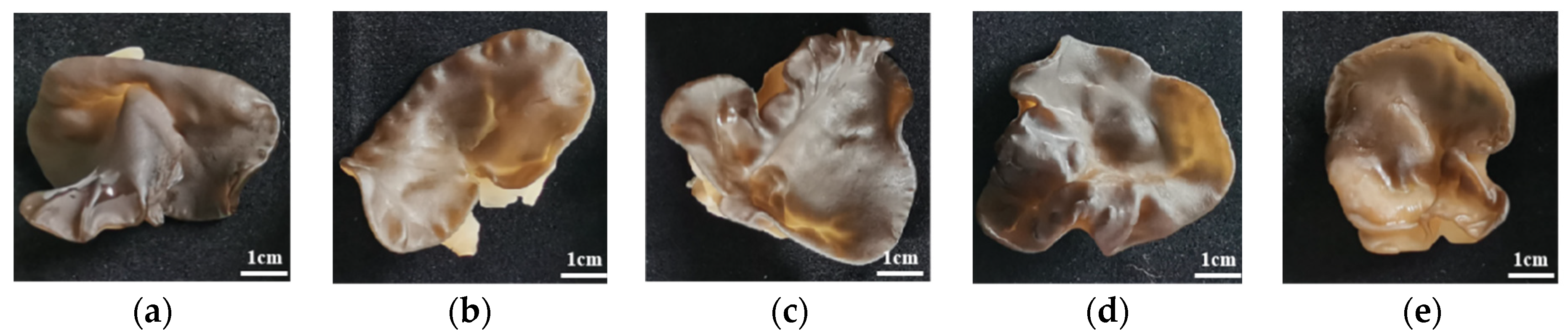
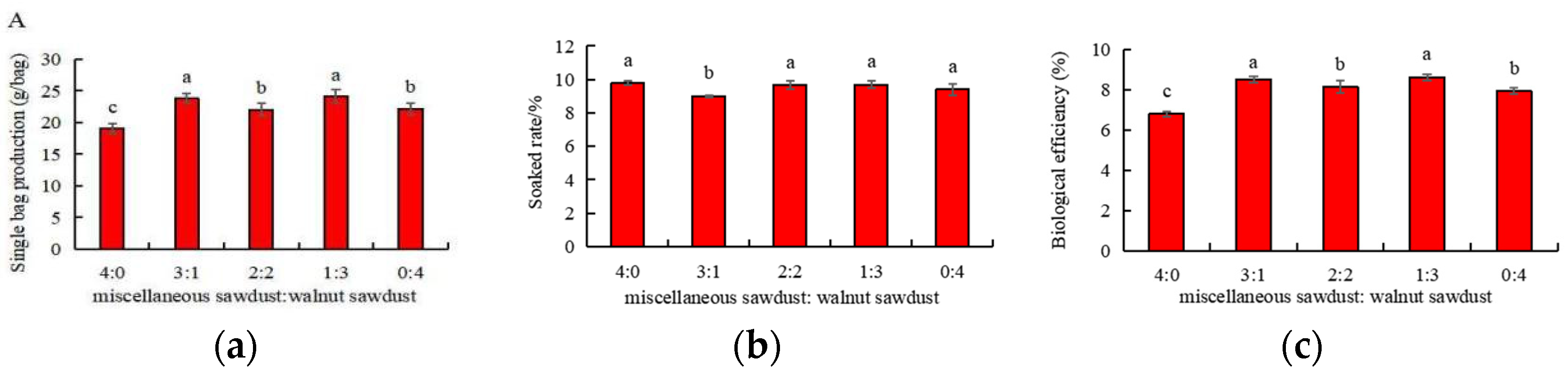
3.4. Effects of Walnut Sawdust Supplemental Ratio on Basidiocarp Agronomic Traits
3.5. Effects of Walnut Sawdust Added Ratio on Basidiocarp Nutrients and Mineral Nutrition
3.6. Comprehensive Evaluation of A. auricula Cultivated in Walnut Sawdust
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, Y.; Wang, L.; Zhang, D.; Li, R.; Cheng, T.; Zhang, Y.; Liu, X.; Wong, G.; Tang, Y.; Wang, H.; et al. Comparative transcription analysis reveals relationship of three major domesticated varieties of Auricularia auricula-judae. Sci. Rep. 2019, 9, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Y.C.; Huang, J.T.; Wang, X.N.; Wang, Y.Q.; Li, H.C.; Yue, T.L. Effect of sulfite treatment on the quality of black fungus. Food Chem. 2022, 385, 132685. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Sulayman, M.; Zhu, X.T.; Zhao, Y.C.; Yang, Z.L.; Hyde, K.D. Species clarification of the culinary Bachu mushroom in western China. Mycologia 2016, 108, 828–836. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.C. Quality management measures for the production of Auricularia auricula-judae edible fungus. Int. J. Med. Mushrooms 2017, 39, 38–39. [Google Scholar]
- Sun, S.; Zhang, X.; Chen, X.; Zhang, L.; Zhu, H. Production of natural edible melanin by Auricularia auricula and its physicochemical properties. Food Chem. 2016, 196, 486–492. [Google Scholar] [CrossRef]
- Li, L.; Fan, X.Z.; Xiao, Y.; Zhou, Y.; Bian, Y.B. The physiological characteristics and genetic diversity analysis of Auricularia auricula-judae cultivated germplasm in China. Mycosystema 2010, 29, 644–652. [Google Scholar]
- Chen, S.Y.; Wu, Q.P.; Zhou, X.Y.; Que, S.H.; Qiu, W.Y. Research progress on cultivation of edible fungi with coniferous sawdust. J. Microbiol. 2002, 2, 49–52. [Google Scholar]
- Zhang, J.P.; Du, M.H. Application status and prospect of Pinus massoniana sawdust as the substrate for the cultivation of Edible fungi. Food Nutr. Sci. 2017, 8, 1105–1113. [Google Scholar]
- Liu, B.H.; Zhao, D.C.; Zhang, P.Y.; Liu, F.C.; Jia, M.; Liang, J. Seedling evaluation of six walnut rootstock species originated in China based on principal component analysis and cluster analysis. Sci. Hortic. 2020, 265, 109212. [Google Scholar] [CrossRef]
- Rugolo, M.; Lechner, B.; Mansilla, R.; Mata, G.; Rajchenberg, M. Evaluation of PleurotusostreatusBasidiomes production on Pinus sawdust and other agricultural and forestry wastes from Patagonia, Argrntina. Maderas-Ciens Technol. 2020, 22, 517–526. [Google Scholar]
- Galic, M.; Stajic, M.; Vukojevic, J.; Cilerdzic, J. Capacity of Auricularia auricula-judae to degrade agroforestry residues. Cellul. Chem. Technol. 2020, 54, 179–184. [Google Scholar] [CrossRef]
- Manavalan, T.; Manavalan, A.; Heese, K. Characterization of lignocellulolytic enzymes from white-rot fungi. Curr. Microbiol. 2015, 70, 485–498. [Google Scholar] [CrossRef]
- Liu, Z.B.; Xu, S.; Sun, X.Z.; Li, Y.; Song, B. Quality of Lentinus edodes cultivated with corn straw and safety evaluation of heavy metal elements. Chin. Edible Fungi 2021, 40, 38–46. [Google Scholar]
- Lakhtar, H.; Ismaili-Alaoui, M.; Philippoussis, A.; Perraud-Gaime, I.; Roussos, S. Screening of strains of Lentinula edodes grown on model olive mill wastewater in solid and liquid state culture for polyphenol biodegradation. Int. Biodeter. Biodegr. 2010, 64, 167–172. [Google Scholar] [CrossRef]
- Ntougias, S.; Baldrian, P.; Ehaliotis, C.; Nerud, F.; Antoniou, T.; Merhautova, V. Biodegradation and detoxification of olive mill wastewater by selected strains of the mushroom genera Ganoderma and Pleurotus. Chemosphere 2012, 88, 620–626. [Google Scholar] [CrossRef]
- Zervakis, G.; Yiatras, P.; Balis, C. Edible mushrooms from olive mill wastes. Int. Biodeter. Biodegr. 1996, 38, 237–243. [Google Scholar] [CrossRef]
- Philippoussis, A.; Diamantopoulou, P.; Papadopoulou, K.; Lakhtar, H.; Roussos, S.; Parissopoulos, G.; Papanikolaou, S. Biomass, laccase and endoglucanase production by Lentinula edodes during soild-state fermentation of reed grass, bean stalks and wheat straw residues. World J. Microbiol. Biotechnol. 2011, 27, 285–297. [Google Scholar] [CrossRef]
- Hu, X.Z. Research progress of lignin degrading enzyme. J. Anhui Agric. Sci. 2011, 39, 6326–6328, 6363. [Google Scholar]
- Sun, Y.Z.; Yang, L.X.; Wang, D.L. Relationship between walnut quinone and soil microbe population in Manchuria juglans forest. Chin. J. Appl. Ecol. 2013, 9, 1517–1522. [Google Scholar]
- Maria, V.O.; Raul, F.F.; Tomas, G.J.; Oscar, A.C.; Juan, A.; Anabela, M.A. Use of lignocellulosic wastes of pecan (Carya illinoinensis) in the cultivation of Ganoderma lucidum. Rev. Micol. 2018, 35, 103–109. [Google Scholar]
- Hao, Z.K.; Zhang, W.E.; Tian, F.H.; Wei, R.; Fu, L.R.; Pan, X.J. Comprehensive evaluation of the adaptability of 11 edible mushroom in Juglans sigillata Dode sawdust substrate. Seed 2022, 9, 60–68. [Google Scholar]
- Lu, M.; Yan, Q.L.; Chi, Y.; Zheng, H.Y.; Xiao, L.J. Production of liquid spawn of an edible mushroom, Sparassis latifolia by submerged fermentation and mycelial growth on pine wood sawdust. Sci. Hort. 2016, 209, 22–30. [Google Scholar]
- Singh, U.; Gautam, A.; Singha, T.K.; Tiwari, A.; Tiwari, P.; Sahai, V.; Sharma, S. Mass production of Pleurotuseryngii mycelia under submerged culture conditions with improved minerals and vitamin D2. LWT 2020, 131, 109665. [Google Scholar] [CrossRef]
- Cheng, Y.C.; Li, J.H.; Su, C.Z.; Zhang, Z.H.; Hu, K.H. Effects of temperature on the physiological activity of Mushroom at post-ripening stage. Edible Fungi China 2020, 39, 26–31, 36. [Google Scholar]
- Wang, M. Discussion on the method of determination of organic carbon content in soil by dimitric acid bell oxidation and external heating. Xinjiang YouseJinshu 2019, 42, 98–99. [Google Scholar]
- Yan, X.Z.; Liu, T.J.; Zhao, H.Y. The protein content in donkey-hide products is determined by the Keshi method. China Food Saf. Mag. 2020, 15, 120–122. [Google Scholar]
- Wang, J.L.; Han, G.; Yan, X.J.; Luo, J.Q.; Du, X.; Dong, H.Y. Determination of seven elements in artificial diamond by inductively coupled plasma atomic emission spectrometry. Metall. Anal. 2017, 03, 14. [Google Scholar]
- Wang, L.J.; Rao, F.J.; Sun, X.; Chen, H.T.; Qin, H.Y.; Zhan, L.X. Effect of transtubular succession on quality of Auricularia heimuer spawns. J. Jilin Agric. Univ. 2022, 44, 293–299. [Google Scholar]
- Liu, J.N.; Dang, A.L.; Zhang, P.Q.; Dai, X.D.; Zhang, J.C. Purification of Laccase from Auricularia auriculata and Characterization of part of Laccase. Fungus Res. 2012, 10, 234–239. [Google Scholar]
- Yuan, B.; Zhou, S.Y.; Liu, C.W.; Zhang, S.; Yan, P.; Liu, A.L. Study on extraction technology and activity determination of polyphenol oxidase. Tea Commun. 2020, 47, 291–296. [Google Scholar]
- An, Q.; Wu, X.J.; Wu, B.; Dai, Y.C. Effects of different carbon and nitrogen sources on lignocellulosic activity of Flammulinavelutipes. Mycosystema 2015, 34, 761–771. [Google Scholar]
- Li, J.J.; Jiang, Y.C.; Meng, Q.X.; Dan, Q.K.; Wang, Y.; Zhu, S.W.; Hua, X.P. Optimum environment parameters of the growth of Auricularia auricula HW15. Edible Fungi China 2018, 37, 32–36. [Google Scholar] [CrossRef]
- Barros, L.; Venturini, B.A.; Baptista, P.; Estevinho, L.M.; Ferreira, I.C. Chemical composition and biological properties of Portuguese wild mushrooms, a comprehensive study. J. Agric Food Chem. 2008, 56, 3856–3862. [Google Scholar] [CrossRef]
- Song, J.L.; Yuan, W.D.; Zhou, Z.F.; Wang, W.K.; Lu, N.; Cheng, J.W.; Yan, J. Activities of extracellular enzymes and polysaccharides in liquid culture of Sanghuangporus. Mycosystema 2020, 39, 352–361. [Google Scholar]
- Wang, C.L.; Zhang, w.e.; Pan, X.J. Study on extraction process of Total Polyphenols from male inflorescences of Walnut. Food Ind. 2015, 36, 77–81. [Google Scholar]
- Wang, S.; Wang, Z.; Wang, P. Evaluation of wheat freezing resistance based on the responses of the physiological indices to low temperature stress. Acta Ecol. 2011, 31, 1064–1072. [Google Scholar]
- Zhou, G.S.; Mei, F.Z.; Zhou, Z.Q.; Zhu, X.T. Comprehensive evaluation of moisture tolerance of different wheat varieties (lines). J. Biomath. 2003, 18, 98–104. [Google Scholar]
- Gong, Z.G.; Wang, Y.F.; Wang, H.; Li, W.; Geng, M.J.; Zhang, W.M.; Liu, L. Research progress on mineral nutrition of walnut. Sci. Silvae Sin. 2021, 57, 178–190. [Google Scholar]
- Dean, H.; Colby. Plant Food Elements Withdrawn from the Soil by Fruits II California Walnut Growers Association. In The California Walnut; Hard Press Publishing: Miami, FL, USA, 2001; pp. 40–46. [Google Scholar]
- Srivastava, A.K.; Hu, C.X. Nutrient Management in Fruit Crops: Issues and Strategies. Indian J. Fertil. 2014, 10, 72–88. [Google Scholar]
- Stajić, M.; Persky, L.; Friesem, D.; Hadar, Y.; Wasser, S.P.; Nevo, E.; Vukojević, J. Effect of different carbon and nitrogen sources on laccase and peroxidases production by selected Pleurotus species. Enzym. Microb. Tech. 2006, 38, 65–73. [Google Scholar] [CrossRef]
- Osma, J.F.; Saravia, V.; Herrera, J.L.T.; Couto, S.R. Mandarin peelings: The best carbon source to produce laccase by static cultures of Trametespubescens. Chemosphere 2007, 67, 1677–1680. [Google Scholar] [CrossRef]
- Li, P.; Wang, H.; Liu, G.; Li, X.; Yao, J. The effect of carbon source succession on laccase activity in the co-culture process of Ganoderma lucidum and a yeast. Enzym. Microb. Technol. 2011, 48, 1–6. [Google Scholar] [CrossRef]
- Adil, B.; Xiang, Q.J.; He, M.L.; Wu, Y.T.; Asghar, M.A.; Arshad, M.; Qin, P.; Gu, Y.F.; Yu, X.M.; Zhao, K.; et al. Effect of sodium and calcium on polysaccharide production and the activities of enzymes involved in the polysaccharide synthesis of Lentinus edodes. AMB Expr. 2020, 10, 47. [Google Scholar] [CrossRef] [Green Version]
- Han, Z.H.; Zhang, P.Q.; Kong, X.H.; Ma, Q.F.; Dai, X.D.; Zhang, J.C. Study on changes of extracellular enzyme activity in Auricularia auriculata and comparison of cultivated characters. Chin. J. Edible Fungi 2007, 14, 41–46. [Google Scholar]
- Wang, Y.J.; Fu, L.Z.; Han, Z.H.; Dai, X.D.; Kong, X.H.; Zhang, J.C. Research progress of the extracellular enzyme of Auricularia auricular. Heilongjiang Sci. 2011, 2, 37–39. [Google Scholar]
- Fang, H.Y.; Ren, Z.M.; Meng, X.X.; Dai, J.J.; Li, S.J.; Li, Z.W.; Li, X. The relationship between extracellular enzymes changes and agronomic traits in Auricularia fuscosuccinea. Mol. Plant Breed. 2018, 16, 4431–4435. [Google Scholar]
- Alcalde, M. Laccases: Biological Functions, Molecular Structure and Industrial Applications. In Industrial Enzyms; Polaina, J., MacCabe, A.P., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 461–476. [Google Scholar]
- Si, J.; Cui, B.K.; He, S.; Dai, Y.C. Optimization of conditions for laccase production by Perenniporiasubacida and its application in dye decolorization. Chin. J. Appl. Environ. Biol. 2011, 17, 736–741. [Google Scholar]
- Wu, Y.; Ma, H.F.; Cao, Y.J.; Si, J.; Cui, B.K. Advances on properties, production, purification and immobilization of fungal laccase. Biotechnol. Bull. 2019, 35, 1–10. [Google Scholar]
- Wu, Z.W. Study on the Effects of Cultivation Formula, Cu(2+) and Mn(2+) on The Growth and Development of Pleurotuscylindracea. Master’s Thesis, Hunan Agricultural University, Changsha, Hunan, 2012. [Google Scholar]
- Zhu, C.W.; Bao, G.W.; Huang, S. Pleurotusostreatus laccase for different heavy metals stress response. J. Environ. Sci. Res. 2015, 28, 1631–1637. [Google Scholar]
- Pandey, R.K.; Tewari, S.; Tewari, L. Lignolytic mushroom Lenzites elegans WDP2: Laccase production, characterization, and bioremediation of synthetic dyes. Ecotoxicol. Environ. Saf. 2018, 158, 50–58. [Google Scholar] [CrossRef]
- Xu, X.H.; Chen, A.; Pei, Y.; Wu, L.J.; Wen, Y.; Zhang, W.P. Analysis of Extracellular Enzymes Activity of Liquid and Solid Strains of Lentinus edodes in Different Media. Mol. Plant Breed. 2021, 12, 6933–6941. [Google Scholar]
- He, C.L.; Li, M.; Tian, J.H.; Dong, Y.N.; Li, S.M. Study on Effective Cultivation Formula Selection and Extracellular Enzymes Activity of Lentinus edodes with Grape Sawdust. Edible Fungi China 2019, 38, 43–49. [Google Scholar]
- Zhao, C.M.; Du, F.; Zou, Y.J.; Hu, Q.X.; Zheng, S.Y. Study on the relationship between mycelial growth and lignin degrading enzymes of Pleurotuseryngii based on different substrates. Edible Fungi China 2020, 39, 109–114. [Google Scholar]
- Rani, P.; Kalyani, N.; Prathiba, K. Evaluation of Lignocellulosci wastes for production of Edible Mushrooms. Appl. Biochem. 2008, 151, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Tanesaka, E.; Takeda, H.; Yoshida, M. Phenol-Oxidizing enzyme expression in Lentinula edodes by the addition of sawdust extract, aromatic compounds, or copper in liquid culture media. Biocontrol Sci. 2013, 18, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Isikhuemhen, O.S.; Mikiashvilli, N.A. Lignocellulolytic enzyme activity, substrate utilization, and mushroom yield by Pleurotusostreatus cultivated on substrate containing anaerobic digester solids. J. Ind. Microbiol. Biotechnol. 2009, 36, 1353–1362. [Google Scholar] [CrossRef]
- Koutrotsios, G.; Tagkouli, D.; Bekiaris, G.; Kaliora, A.; Tsiaka, T.; Tsiantas, K.; Chatzipavlidis, I.; Zoumpoulakis, P.; Kalogeropoulos, N.; Zervakis, G.I. Enhancing the nutritional and functional properties of Pleurotuscitrinopileatus mushrooms through the exploitation of winery and olive mill wastes. Food Chem. 2022, 370, 131022. [Google Scholar] [CrossRef]
- Basso, V.; Schiavenin, C.; Mendonca, S.; Goncalves, F.; Salvador, M.; Camassola, M. Chemical features and antioxidant profile by Schizophyllun commune produced on different agroindustrial wastes and byproducts of biodiesel production. Food Chem. 2020, 329, 127089. [Google Scholar] [CrossRef]
- Dubost, N.J.; Ou, B.; Beelman, R.B. Quantification of polyphenols and ergothioneine in cultivated mushrooms and correlation to total antioxidant capacity. Food Chem. 2007, 105, 727–735. [Google Scholar] [CrossRef]
- Gambato, G.; Todescato, K.; Paväo, E.M.; Scortegagna, A.; Fontana, R.C.; Salvador, M.; Camassola, M. Evaluation of productivity and antioxidant profile of solid-state cultivated macrofungi Pleurotusalbidus and Pycnoporussanguineus. Bioresour. Technol. 2016, 207, 46–51. [Google Scholar] [CrossRef]
- Tang, Q.Y.; Feng, M.G. Practical Statistical Analysis and Its DPS Data Processing System; Science Press: Beijing, China, 2002. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Macro mineral element | Species | N (mg/g) | P (mg/g) | K (mg/g) | Ca (mg/g) | Mg (mg/g) |
| WS | 5.31 ± 0.14 a | 0.95 ± 0.04 a | 2.50 ± 0.07 a | 10.48 ± 0.22 a | 1.23 ± 0.04 a | |
| MS | 4.31 ± 0.10 b | 0.26 ± 0.01 b | 1.38 ± 0.02 b | 6.74 ± 0.23 b | 0.82 ± 0.02 b | |
| Micro mineral element | Species | Fe (mg/kg) | Mn (mg/kg) | Cu (mg/kg) | Zn (mg/kg) | B (mg/kg) |
| WS | 678.43 ± 23.83 b | 11.16 ± 0.68 a | 252.62 ± 3.87 a | 28.90 ± 1.07 b | 195.23 ± 2.66 b | |
| MS | 800.43 ± 60.96 a | 6.50 ± 0.94 b | 237.76 ± 6.44 b | 59.72 ± 1.93 a | 223.32 ± 3.22 a |
| MS: WS | Protein/% | Fat/% | Polysaccharide (mg/g) | Total Phenols (mg/100 g) | Total Flavonoids (mg/100 g) | Total Sugar (g/100 g) | Moisture Content/% |
|---|---|---|---|---|---|---|---|
| 4:0 | 9.82 ±0.05 a | 1.08 ± 0.03 c | 87.19 ± 1.70 c | 131.77 ± 8.73 d | 95.55 ± 8.13 b | 65.55 ± 2.32 a | 89.76 ± 0.12 a |
| 3:1 | 9.74 ± 0.10 a | 1.20 ± 0.03 b | 89.13 ± 1.08 c | 201.14 ± 8.93 a | 78.18 ± 7.29 c | 63.30 ± 1.91 a | 89.37 ± 0.39 a |
| 2:2 | 9.65 ± 0.19 a | 1.33 ± 0.03 a | 95.99 ± 2.60 b | 188.93 ± 9.78 ab | 117.01 ± 7.53 a | 56.18 ± 2.09 b | 89.66 ± 0.25 a |
| 1:3 | 9.76 ± 0.07 a | 1.33 ± 0.02 a | 99.70 ± 1.73 a | 172.44 ± 9.36 bc | 90.80 ± 7.48 bc | 55.55 ± 1.88 b | 88.89 ± 0.07 b |
| 0:4 | 9.79 ± 0.03 a | 1.24 ± 0.05 b | 88.73 ± 2.39 c | 158.28 ± 8.59 c | 103.61 ± 7.85 ab | 54.66 ± 2.05 b | 89.67 ± 0.22 a |
| MS: WS | Ca (mg/g) | Mg (mg/g) | Fe (mg/kg) | Mn (mg/kg) | Zn (mg/kg) | Cu (mg/kg) |
|---|---|---|---|---|---|---|
| 4:0 | 2.85 ± 0.07 b | 1.37 ± 0.03 c | 431.77 ± 15.81 bc | 9.73 ± 0.47 c | 41.56 ± 0.78 c | 216.67 ± 1.80 bc |
| 3:1 | 2.23 ± 0.05 c | 1.26 ± 0.03 d | 425.52 ± 14.77 c | 9.27 ± 0.49 c | 48.96 ± 0.81 a | 212.50 ± 1.56 c |
| 2:2 | 2.74 ± 0.10 b | 1.41 ± 0.05 bc | 465.10 ± 25.07 b | 11.23 ± 0.98 b | 39.69 ± 0.58 c | 221.35 ± 4.77 b |
| 1:3 | 3.36 ± 0.15 a | 1.49 ± 0.06 b | 545.94 ± 28.31 a | 12.94 ± 0.77 a | 37.01 ± 1.15 d | 227.22 ± 2.36 a |
| 0:4 | 3.27 ± 0.10 a | 1.65 ± 0.05 a | 427.91 ± 12.93 c | 11.43 ± 0.77 b | 46.21 ± 1.08 b | 218.36 ± 1.80 b |
| Correlation Coefficient | Single Bag Production | Soaked Rate | Protein | Fat | Total Sugar | Polysaccharide | Total Phenols | Total Flavonoids | Ca | Fe | Mg | Mn | Cu | Zn |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Single bag production | 1 | |||||||||||||
| Soaked rate | −0.471 | 1 | ||||||||||||
| Protein | −0.523 | 0.063 | 1 | |||||||||||
| Fat | 0.793 | 0.072 | −0.702 | 1 | ||||||||||
| Total sugar | −0.552 | −0.173 | 0.382 | −0.859 | 1 | |||||||||
| Polysaccharide | 0.628 | 0.377 | −0.545 | 0.861 | −0.624 | 1 | ||||||||
| Total phenols | 0.843 | −0.622 | −0.762 | 0.604 | −0.242 | 0.387 | 1 | |||||||
| Total flavonoids | −0.226 | 0.584 | −0.458 | 0.369 | −0.529 | 0.228 | −0.156 | 1 | ||||||
| Ca | −0.025 | 0.611 | 0.330 | 0.326 | −0.650 | 0.393 | −0.512 | 0.345 | 1 | |||||
| Mg | 0.0179 | 0.31 | 0.235 | 0.332 | −0.763 | 0.138 | −0.370 | 0.471 | 0.868 | 1 | ||||
| Fe | 0.4924 | 0.4629 | −0.194 | 0.662 | −0.494 | 0.925 * | 0.125 | 0.016 | 0.553 | 0.188 | 1 | |||
| Mn | 0.4396 | 0.4855 | −0.151 | 0.765 | −0.850 | 0.807 | −0.007 | 0.352 | 0.840 | 0.679 | 0.834 | 1 | ||
| Cu | 0.3137 | 0.6695 | −0.196 | 0.694 | −0.684 | 0.878 | −0.0703 | 0.369 | 0.766 | 0.482 | 0.917 * | 0.947 * | 1 | |
| Zn | 0.0254 | −0.8754 | 0.193 | −0.421 | 0.329 | −0.759 | 0.2408 | −0.420 | −0.564 | −0.179 | −0.808 | −0.670 | −0.882 * | 1 |
| Index | Z1 | Z2 | Z3 | |
|---|---|---|---|---|
| Eigenvalues | 7.086 | 3.734 | 1.700 | |
| Contribution ratio/% | 50.617 | 26.671 | 12.143 | |
| Cumulative contribution ratio/% | 50.617 | 77.287 | 89.430 | |
| Eigenvector | Single bag production | 0.167 | 0.426 | 0.154 |
| Soaked rate | 0.206 | −0.354 | −0.326 | |
| Protein | −0.133 | −0.363 | 0.172 | |
| Fat | 0.313 | 0.271 | 0.096 | |
| Total sugar | −0.313 | −0.080 | −0.387 | |
| Polysaccharide | 0.335 | 0.165 | −0.232 | |
| Total phenols | 0.335 | 0.165 | −0.232 | |
| Total flavonoids | 0.177 | −0.130 | 0.061 | |
| Ca | 0.274 | −0.296 | 0.246 | |
| Mg | 0.208 | −0.223 | 0.533 | |
| Fe | 0.320 | 0.039 | −0.249 | |
| Mn | 0.364 | −0.044 | 0.123 | |
| Cu | 0.363 | −0.077 | −0.120 | |
| Zn | −0.292 | 0.160 | 0.419 |
| MS:WS | Z1 | Z2 | Z3 | U(X1) | U(X2) | U(X3) | D | Rank |
|---|---|---|---|---|---|---|---|---|
| 4:0 | 243.489 | 71.249 | −151.019 | 0.083 | 0.000 | 0.777 | 0.172 | 5 |
| 3:1 | 238.782 | 113.547 | −145.267 | 0.000 | 1.000 | 0.931 | 0.441 | 3 |
| 2:2 | 269.266 | 102.425 | −156.569 | 0.539 | 0.737 | 0.630 | 0.611 | 2 |
| 1:3 | 295.305 | 100.569 | −180.233 | 1.000 | 0.693 | 0.000 | 0.749 | 1 |
| 0:4 | 249.094 | 86.704 | −142.658 | 0.182 | 0.365 | 1.000 | 0.368 | 4 |
| Index weight | 50.617 | 26.671 | 12.143 | 0.566 | 0.298 | 0.136 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, Z.; Zhang, W.; Tian, F.; Wei, R.; Pan, X. Enhancing the Nutritional and Functional Properties of Auricularia auricula through the Exploitation of Walnut Branch Waste. Foods 2022, 11, 3242. https://doi.org/10.3390/foods11203242
Hao Z, Zhang W, Tian F, Wei R, Pan X. Enhancing the Nutritional and Functional Properties of Auricularia auricula through the Exploitation of Walnut Branch Waste. Foods. 2022; 11(20):3242. https://doi.org/10.3390/foods11203242
Chicago/Turabian StyleHao, Zhenkun, Wen’e Zhang, Fenghua Tian, Rong Wei, and Xuejun Pan. 2022. "Enhancing the Nutritional and Functional Properties of Auricularia auricula through the Exploitation of Walnut Branch Waste" Foods 11, no. 20: 3242. https://doi.org/10.3390/foods11203242