
Changes in Phenolic Compounds and Antioxidant Activity during Development of ‘Qiangcuili’ and ‘Cuihongli’ Fruit
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
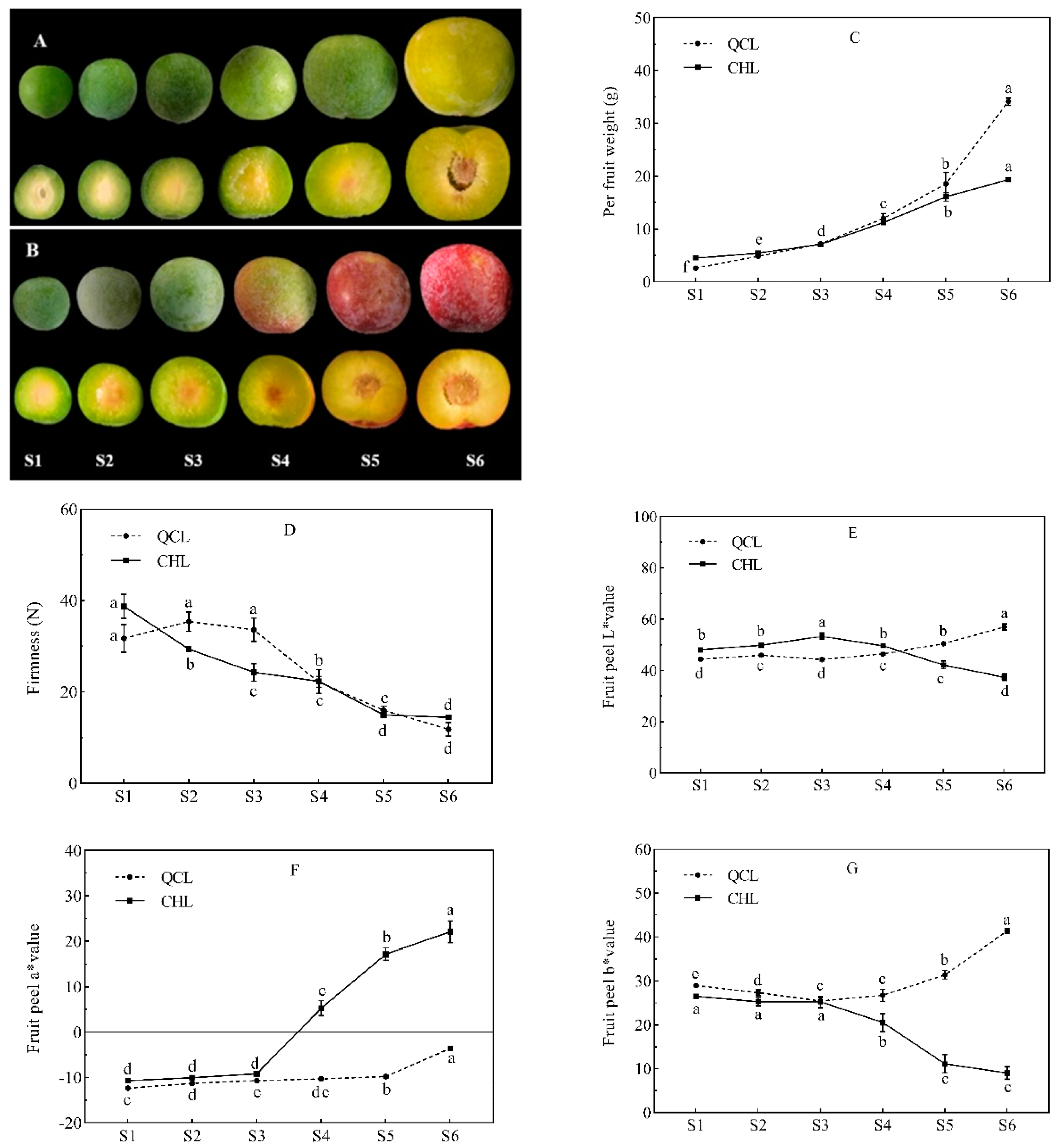
2.2. Determination of Appearance Quality
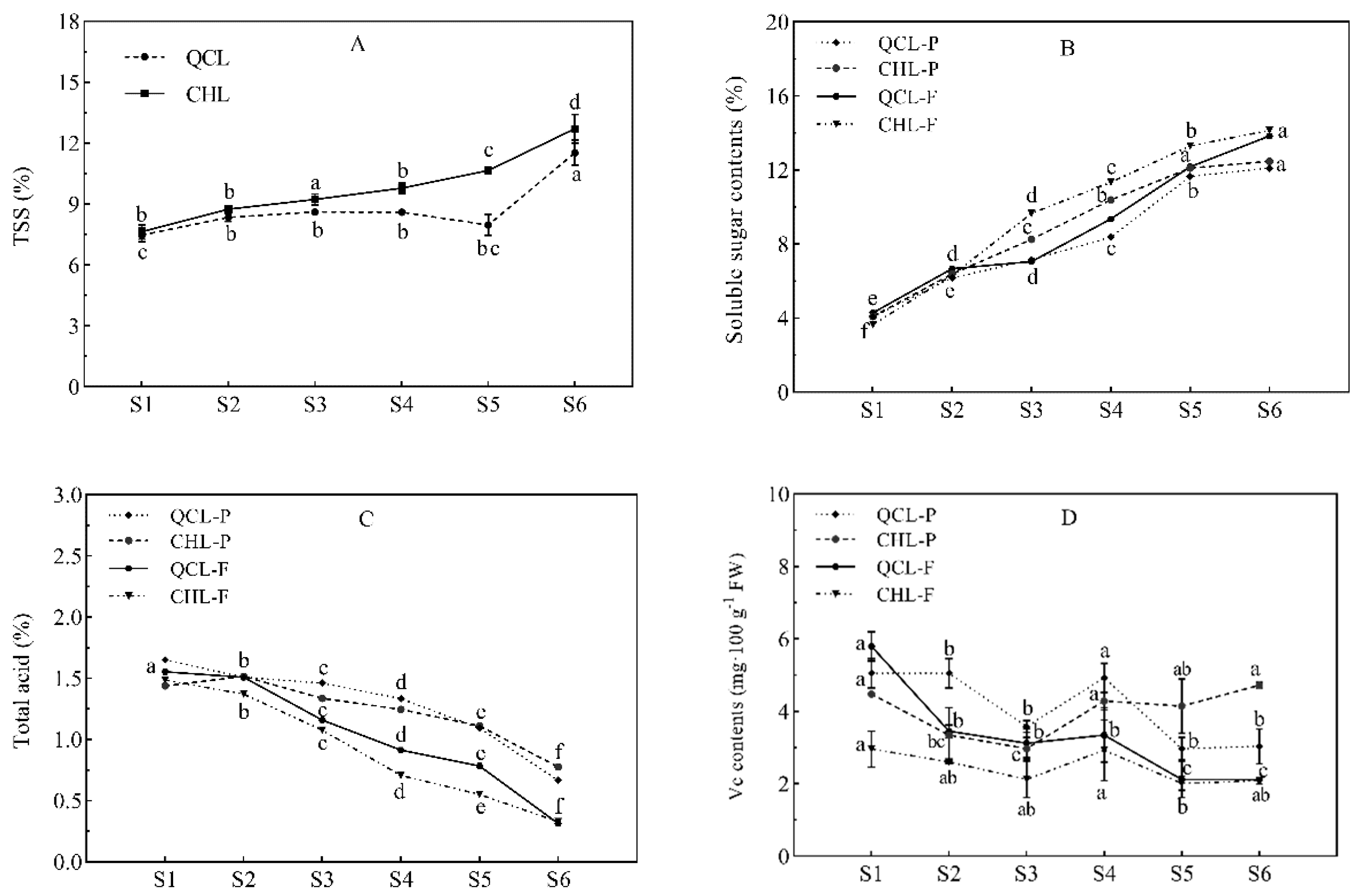
2.3. Determination of Total Soluble Solids (%), Soluble Sugars (%), Titratable Acidity (%), and Vitamin C Content
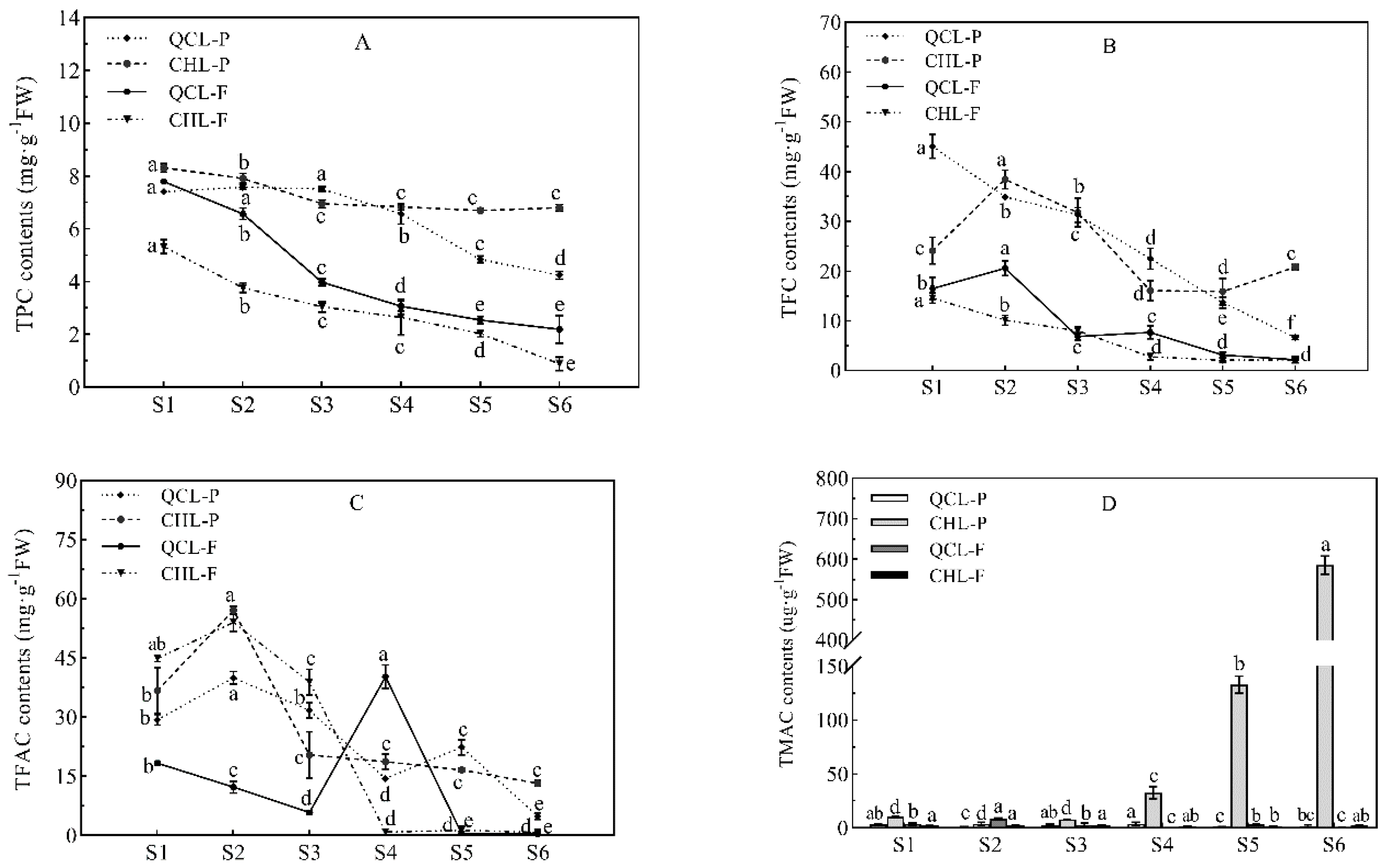
2.4. Determination of Total Phenolic Content
2.5. Determination of Flavonoid Content
2.6. Determination of Total Flavanol Content
2.7. Determination of Total Anthocyanin Content
2.8. Determination of Phenolic Components
2.9. Antioxidant Activity Analysis
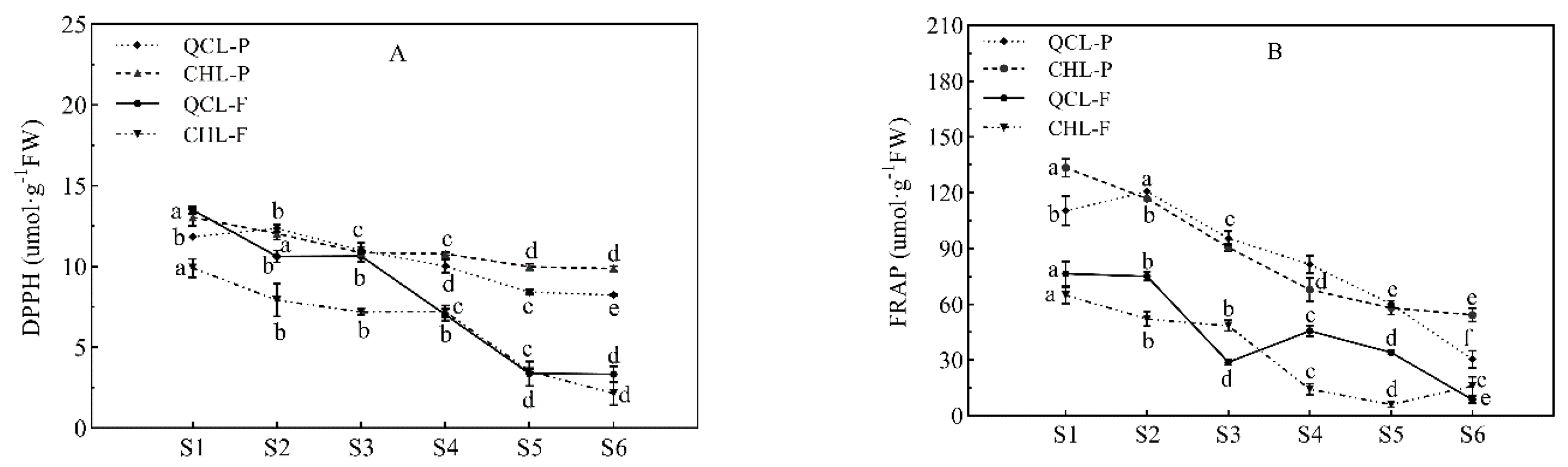
2.9.1. Determination of DPPH Scavenging Activity
2.9.2. Determination of Ferric Reducing Antioxidant Power (FRAP)
2.10. Analysis of Relative Expression of Structural Genes Related to the Synthesis of Phenolic Compounds
2.11. Statistical Analysis
3. Results
3.1. Changes in the Appearance Index during Fruit Development of the Two Plum Cultivars
3.2. Changes in TSS, SS, TA, and Vc Contents during Fruit Development of the Two Plum Cultivars
3.3. Changes in Phenolic Content during Fruit Development of the Two Plum Cultivars
3.4. Changes in the Phenolic Components during Fruit Development of the Two Plum Cultivars
3.5. Changes in the Antioxidant Capacity during ‘Qiangcuili’ and ‘Cuihongli’ Fruit Development
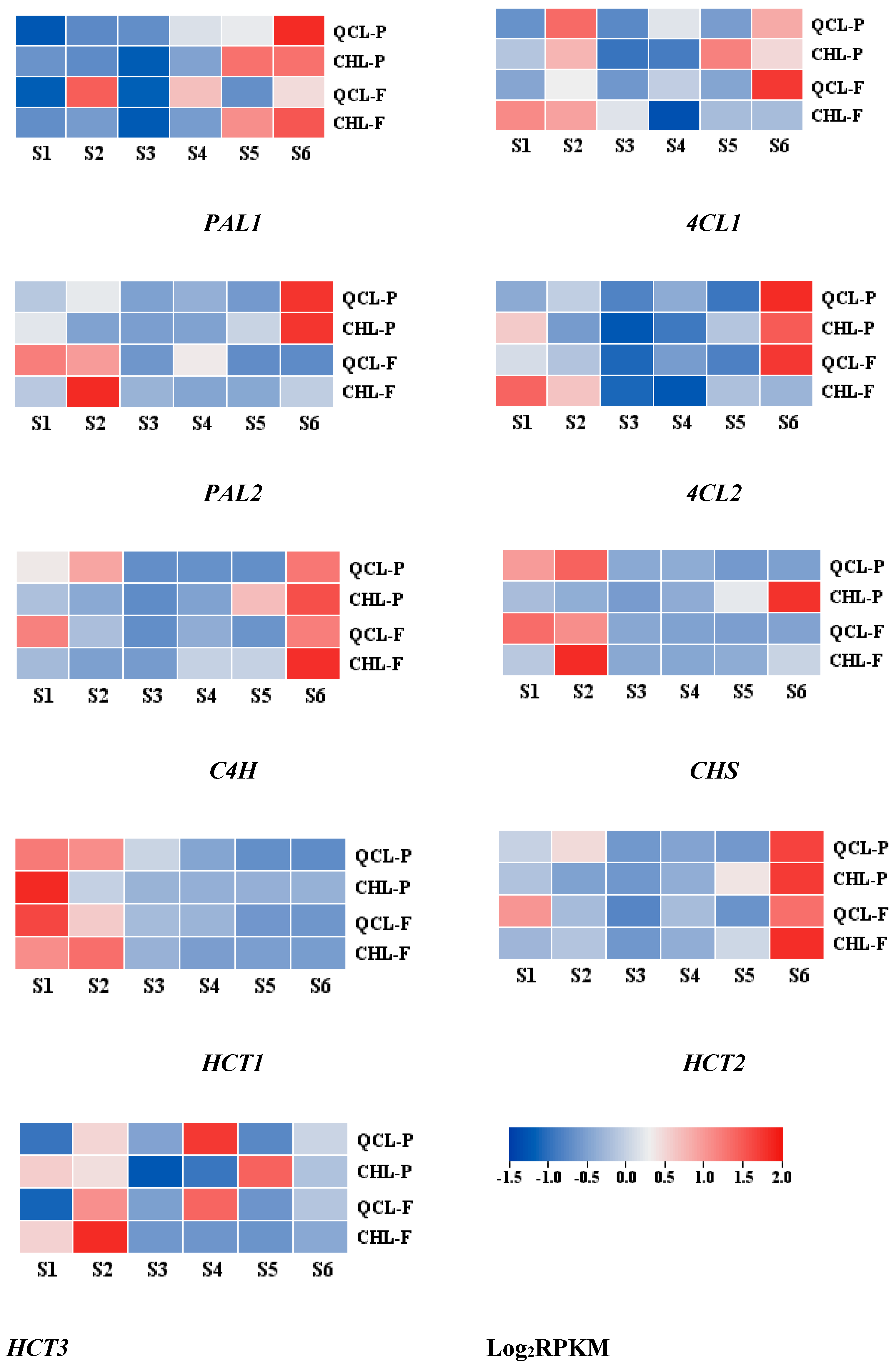
3.6. Analysis of the Development Process and Expression of Related Genes in the ‘Qiangcuili’ and ‘Cuihongli’ Peel and Flesh
3.7. Correlation Analysis of Phenolic Content and Related Gene Expression during Fruit Development of ‘Qiangcuili’ and ‘Cuihongli’
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pem, D.; Jeewon, R. Fruit and Vegetable Intake: Benefits and Progress of Nutrition Education Interventions−Narrative Review Article. Iran J. Public Health. 2015, 44, 1309–1321. [Google Scholar] [PubMed]
- Rodriguez−Mateos, A.; Vauzour, D.; Krueger, C.G.; Shanmuganayagam, D.; Reed, J.; Calani, L.; Mena, P.; Del Rio, D.; Crozier, A. Bioavailability, bioactivity and impact on health of dietary flavonoids and related compounds: An update. Arch. Toxicol. 2014, 88, 1803–1853. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Xu, B.T.; Xu, X.R.; Gan, R.Y.; Zhang, Y.; Xia, E.Q.; Li, H.B. Antioxidant capacities and total phenolic contents of 62 fruits. Food Chem. 2011, 129, 345–350. [Google Scholar] [CrossRef]
- Noratto, G.; Porter, W.; Byrne, D.; Cisneros−Zevallos, L. Identifying peach and plum polyphenols with chemopreventive potential against estrogen—Independent breast cancer cells. J. Agric. Food Chem. 2009, 57, 5219–5226. [Google Scholar] [CrossRef]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1000–1001. [Google Scholar] [CrossRef]
- Vinson, J.A.; Su, X.; Zubik, L.; Bose, P. Phenol antioxidant quantity and quality in foods: Fruits. J. Agric. Food Chem. 2001, 49, 5315–5321. [Google Scholar] [CrossRef]
- Lee, S.H.; Lillehoj, H.S.; Cho, S.M.; Chun, H.K.; Park, H.J.; Lim, C.I.; Lillehoj, E.P. Immunostimulatory effects of oriental plum (Prunus salicina Lindl.). Comp. Immunol. Microbiol. Infect. Dis. 2009, 32, 407–417. [Google Scholar] [CrossRef]
- Utsunomiya, H.; Yamakawa, T.; Kamei, J.; Kadonosono, K.; Tanaka, S. Anti-hyperglycemic effects of plum in a rat model of obesity and type 2 diabetes, Wistar fatty rat. Biomed. Res. 2005, 26, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Shukitt-Hale, B.; Kalt, W.; Carey, A.N.; Vinqvist-Tymchuk, M.; Mcdonald, J.; Joseph, J.A. Plum juice, but not dried plum powder, is effective in mitigating cognitive deficits in aged rats. Nutrition 2009, 25, 567–573. [Google Scholar] [CrossRef]
- Kim, H.J.; Yu, M.H.; Lee, I.S. Inhibitory effects of methanol extract of plum (Prunus salicina L., cv.‘Soldam’) fruits against benzo (α) pyrene−induced toxicity in mice. Food Chem. Toxicol. 2008, 46, 3407–3413. [Google Scholar] [CrossRef]
- Wang, B.; Huang, Q.Y.; Venkitasamy, C.; Chai, H.; Gao, H.; Cheng, N.; Cao, W.; Lv, X.; Pan, Z. Changes in phenolic compounds and their antioxidant capacities in jujube (Ziziphus jujuba Miller.) during three edible maturity stages. Lwt Food Sci. Technol. 2016, 66, 56–62. [Google Scholar] [CrossRef]
- Metwally, A.; Finkemeier, I.; Georgi, M.; Dietz, K.J. Salicylic Acid Alleviates the Cadmium Toxicity in Barley Seedlings. Plant Physiol. 2003, 132, 272–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, H.; Yang, W.; Li, J.; Tang, W.; Gong, R. Transcriptomic and Metabolomic Analysis of Quality Changes during Sweet Cherry Fruit Development and Mining of Related Genes. Int. J. Mol. Sci. 2022, 23, 7402. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.; Sathasivam, R.; Nguyen, B.V.; Park, N.I.; Woo, S.H.; Park, S.U. Expression Analysis of Phenylpropanoid Pathway Genes and Metabolomic Analysis of Phenylpropanoid Compounds in Adventitious, Hairy, and Seedling Roots of Tartary Buckwheat. Plants 2021, 11, 90. [Google Scholar] [CrossRef] [PubMed]
- Ayhan, N.K. Assessment of Elemental Content, Antioxidant Activity and Total Phenolic Content of Vitis sylvestris Gmelin. J. Turk. Chem. Soc. Sect. A Chem. 2020, 7, 405–410. [Google Scholar] [CrossRef] [Green Version]
- Resende, M.; Paulo, L.; Antunes, P.; Miguel, P.C. High total phenol content, antioxidant activity and mineral content in ‘Sweetheart’ cherry peel. Acta Hortic. 2017, 1161, 673–676. [Google Scholar] [CrossRef]
- OrdóñezDíaz José, L.; Cardeñosa, V.; MuñozRedondo, J.M.; Ferreres, F.; Pereira, C.G.; Medrano, E.; MorenoRojas, J.M.; Moreno, D.A. Impact of Abiotic Stresses (Nitrogen Reduction and Salinity Conditions) on Phenolic Compounds and Antioxidant Activity of Strawberries. Processes 2021, 9, 1044. [Google Scholar] [CrossRef]
- Munekata, P.E.S.; Yilmaz, B.; Pateiro, M.; Kumar, M.; Domínguez, R.; Shariati, M.A.; Hano, C.; Lorenzo, J.M. Valorization of by−products from Prunus genus fruit processing: Opportunities and applications. Crit. Rev. Food Sci. Nutr. 2022, 14, 1–16. [Google Scholar] [CrossRef]
- Zhang, S.W.; Zong, Y.J.; Fang, C.Y.; Huang, S.H.; Li, J.; Xu, J.H.; Wang, Y.F.; Liu, C.H. Optimization of rapid determination of soluble sugar content in barley leaves by anthrone colorimetry. Food Res. Dev. 2020, 41, 196–200. [Google Scholar]
- Yang, X.Y.; Wang, Y.P.; Leng, P.; Wang, C.L.; Liu, S.T.; Hu, Y. Regulation of ABA on melon fruit maturation and softening. J. China Agric. Univ. 2010, 15, 25–32. [Google Scholar] [CrossRef]
- García−Ruiz, A.; González-Rompinelli, E.M.; Bartolomé, B.; Moreno-Arribas, M.V. Potential of wine−associated lactic acid bacteria to degrade biogenic amines. Int. J. Food Microbiol. 2011, 148, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Yong−Seo, P.; Soon−Teck, J.; Seong−Gook, K.; Buk, G.H.; Patricia, A.A.; Fernando, T.; Jerzy, D.; Jacek, N.; Shela, G. Antioxidants and proteins in ethylene−treated kiwifruits. Food Chem. 2007, 107, 640–648. [Google Scholar] [CrossRef]
- Li, Y.G.; Tanner, G.; Larkin, P. The DMACA–HCl Protocol and the Threshold Proanthocyanidin Content for Bloat Safety in Forage Legumes. J. Sci. Food Agric. 1996, 70, 89–101. [Google Scholar] [CrossRef]
- Johnson, J.; Collins, T.; Walsh, K.; Naiker, M. Solvent extractions and spectrophotometric protocols for measuring the total anthocyanin, phenols and antioxidant content in plums. Chem. Pap. 2020, 74, 4481–4492. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, W.; Yin, X.; Su, M.; Sun, C.; Li, X.; Chen, K. Phenolic composition and antioxidant properties of different peach [Prunus persica (L.) Batsch] cultivars in China. Int. J. Mol. Sci. 2015, 16, 5762–5778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brand−Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Lwt Food Sci. Technol. 1995, 28, 28–30. [Google Scholar] [CrossRef]
- Benzie, I.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Methods Enzymol. 1999, 299, 15–27. [Google Scholar] [CrossRef]
- Shu, Y.; Wan, T.J.; Ya, N.Y.; Yu, H.F. An Optimized CTAB Method for Genomic DNA Extraction from Freshly−picked Pinnae of Fern, Adiantum capillus-veneris L. Bio-Protocol 2018, 8, e2906. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.; Lee, T.; Cheng, C.H.; Buble, K.; Zheng, P.; Yu, J.; Humann, J.; Ficklin, S.P.; Gasic, K.; Scott, K.; et al. 15 years of GDR: New data and functionality in the Genome Database for Rosaceae. Nucleic Acids Res. 2019, 47, D1137–D1145. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real−time quantitative PCR and the 2−11CT Method. Methods. 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Duan, C.; Qiu, X.; Xiong, B.; Zhang, H.; Wang, Z. Fruit Textural Characteristics of 23 Plum (Prunus salicina Lindl) Cultivars: Evaluation and Cluster Analysis. HortScience 2021, 56, 7. [Google Scholar] [CrossRef]
- Huang, X.; Wu, Y.; Zhang, S.; Yang, H.; Wu, W.; Lyu, L.; Li, W. Variation in Bioactive Compounds and Antioxidant Activity of Rubus Fruits at Different Developmental Stages. Foods 2022, 11, 1169. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, X.; Li, L.; Aghdam, M.S.; Wei, X.; Liu, J.; Xu, Y.; Luo, Z. Elevated CO2 delayed the chlorophyll degradation and anthocyanin accumulation in postharvest strawberry fruit. Food Chem. 2019, 285, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Rupasinghe, H.; Jayasankar, S.; Lay, W. Variation in total phenolics and antioxidant capacity among European plum genotypes. Sci. Hortic. 2006, 108, 240–246. [Google Scholar] [CrossRef]
- Slimestad, R.; Vangdal, E.; Brede, C. Analysis of Phenolic Compounds in Six Norwegian Plum Cultivars (Prunus domestica L.). J. Agric. Food Chem. 2009, 57, 11370–11375. [Google Scholar] [CrossRef]
- Aliyazicioglu, R.; Yildiz, O.; Sahin, H.; Eyupoglu, O.E.; Ozkan, M.T.; Karaoglu, S.A. Phenolic Components and Antioxidant Activity of Prunus spinosafrom Gumushane, Turkey. Chem. Nat. Compd. 2015, 51, 346–349. [Google Scholar] [CrossRef]
- Cosmulescu, S.; Trandafir, I.; Nour, V.; Botu, M. Total Phenolic, Flavonoid Distribution and Antioxidant Capacity in Skin, Pulp and Fruit Extracts of Plum Cultivars. J. Food Biochem. 2015, 39, 64–69. [Google Scholar] [CrossRef]
- Wang, Y.; Qi, D.; Wang, S.; Cao, X.; Ye, Y.; Suo, Y. Comparison of Phenols Content and Antioxidant Activity of Fruits from Different Maturity Stages of Ribes stenocarpum Maxim. Molecules 2018, 23, 3148. [Google Scholar] [CrossRef] [Green Version]
- Wojdyło, A.; Oszmiański, J. Antioxidant Activity Modulated by Polyphenol Contents in Apple and Leaves during Fruit Development and Ripening. Antioxidants 2020, 9, 567. [Google Scholar] [CrossRef]
- He, J.G.; Cheng, Y.D.; Guan, J.F.; Ge, W.Y.; Zhao, Z. Changes of chlorogenic acid content and its synthesis−associated genes expression in Xuehua pear fruit during development. J. Integr. Agr. 2017, 16, 471–477. [Google Scholar] [CrossRef]
- González, M.; Salazar, E.; Cabrera, S.; Olea, P.; Carrasco, B. Analysis of anthocyanin biosynthesis genes expression profiles in contrasting cultivars of Japanese plum (Prunus salicina L.) during fruit development. Gene Expr. Patterns 2016, 21, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Remberg, S.F.; Måge, F.; Haffner, K.; Blomhoff, R. Highbush blueberries vaccinium corymbosum l. raspberries Rubus idaeus l. and black currants Ribes nigrum l. influence of cultivar on antioxidant activity and other quality parameters. Acta Hortic. 2007, 74, 259–266. [Google Scholar] [CrossRef]
- Sun, Y.; Li, M.; Mitra, S.; Hafiz, M.R.; Debnath, B.; Lu, X.; Jian, H.; Qiu, D. Comparative Phytochemical Profiles and Antioxidant Enzyme Activity Analyses of the Southern Highbush Blueberry (Vaccinium corymbosum) at Different Developmental Stages. Molecules 2018, 23, 2209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Wang, D.; Tan, C.; Hu, Y.; Sundararajan, B.; Zhou, Z. Profiling of Flavonoid and Antioxidant Activity of Fruit Tissues from 27 Chinese Local Citrus Cultivars. Plants 2020, 9, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bate, N.J.; Orr, J.; Ni, W.; Meromi, A.; Nadler-Hassar, T.; Doerner, P.W.; Dixon, R.A.; Lamb, C.J.; Elkind, Y. Quantitative relationship between phenylalanine ammonia−lyase levels and phenylpropanoid accumulation in transgenic tobacco identifies a rate−determining step in natural product synthesis. Proc. Natl. Acad. Sci. USA 1994, 9, 1069–1619. [Google Scholar] [CrossRef] [Green Version]
- Lepelley, M.; Cheminade, G.; Tremillon, N.; Simkin, A.; Caillet, V.; James, M.C. Chlorogenic acid synthesis in coffee: An analysis of CGA content and real−time RT−PCR expression of HCT, HQT, C3H1, and CCoAOMT1 genes during grain development in C. canephora. Plant Sci. 2007, 172, 978–996. [Google Scholar] [CrossRef]
- Pu, G.; Wang, P.; Zhou, B.; Liu, Z.; Xiang, F. Cloning and characterization of Lonicera japonica p−coumaroyl ester 3−hydroxylase which is involved in the biosynthesis of chlorogenic acid. Biosci. Biotechnol. Biochem. 2013, 77, 1403–1409. [Google Scholar] [CrossRef] [Green Version]
- Soltani, B.M.; Ehlting, J.; Hamberger, B.; Douglas, C.J. Multiple cis regulatory elements regulated is distinct and complex patterns of developmental and wound induced expression of Arabidopsis thaliana 4CL gene family members. Planta 2006, 224, 1226–1238. [Google Scholar] [CrossRef]
- Léchaudel, M.; Darnaudery, M.; Joët, T.; Fournier, P.; Joas, J. Genotypic and environmental effects on the level of ascorbic acid, phenolic compounds and related gene expression during pineapple fruit development and ripening. Plant Physiol. Biochem. 2018, 130, 127–138. [Google Scholar] [CrossRef]
- Chen, Z.F.; Deng, H.H.; Xiong, B.; Li, S.C.; Yang, L.C.; Yang, Y.T.; Huang, S.J.; Tan, L.P.; Sun, G.C.; Wang, Z.H. Rootstock Effects on Anthocyanin Accumulation and Associated Biosynthetic Gene Expression and Enzyme Activity during Fruit Development and Ripening of Blood Oranges. Agriculture 2022, 12, 342. [Google Scholar] [CrossRef]
- Rahim, M.A.; Busatto, N.; Trainotti, L. Regulation of anthocyanin biosynthesis in peach fruits. Planta 2014, 240, 913–929. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.H.; Li, W.F.; Mao, J.; Li, W.; Zuo, C.W.; Zhao, X.; Dawuda, M.M.; Shi, X.Y.; Chen, B.H. Synthesis of light−inducible and light−independent anthocyanins regulated by specific genes in grape ‘Marselan’ (V. vinifera L.). PeerJ 2019, 7, e6521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Wang, D.; Liu, J.; Yu, X.; Wang, R.; Wei, Y.; Wen, C.; Ouyang, Z. Transcriptomic analysis of key genes involved in chlorogenic acid biosynthetic pathway and characterization of MaHCT from Morus alba L. Protein Expr. Purif. 2019, 156, 25–35. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Developmental Site | Species | Phenolic Acids (mg·100 g−1 FW) | Flavonol Component (mg·100 g−1 FW) | Flavanol Component (mg·100 g−1 FW) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gallic Acid | Neochlorogenic Acid | Chlorogenic Acid | Vanillic Acid | Ferulic Acid | Benzoic Acid | Rutin | Quercetin | Procyanidin B1 | ||
| QCL-P | S1 | 2.10 ± 0.06 a | 37.4 ± 0.79 a | 51.99 ± 4.59 a | 3.84 ± 0.12 b | 2.19 ± 0.06 a | 5.70 ± 0.18 b | 3.43 ± 0.14 b | ND | 174.68 ± 10.23 a |
| S2 | 1.66 ± 0.02 b | 24.87 ± 0.05 b | 35.10 ± 1.41 b | 35.15 ± 1.83 a | 1.51 ± 0.17 b | 20.22 ± 0.11 a | 17.64 ± 0.55 a | 15.83 ± 0.85 a | 66.55 ± 4.33 d | |
| S3 | 1.07 ± 0.03 c | 18.88 ± 0.55 c | 24.68 ± 0.42 c | 2.74 ± 0.11 bc | 1.25 ± 0.02 c | 4.39 ± 0.35 c | 1.93 ± 0.07 cd | 2.51 ± 0.08 b | 145.67 ± 5.08 b | |
| S4 | 0.98 ± 0.02 c | 14.19 ± 0.50 d | 18.41 ± 1.62 c | 0.90 ± 0.02 c | 1.1 ± 0.07 cd | 5.40 ± 0.19 b | 1.18 ± 0.06 d | 0.55 ± 0.01 c | 151.13 ± 6.92 b | |
| S5 | 0.64 ± 0.03 d | 8.70 ± 0.42 e | 10.60 ± 0.33 d | 1.5 ± 0.02 bc | 0.88 ± 0.01 de | 4.12 ± 0.20 c | 1.61 ± 0.04 cd | 1.31 ± 0.01 c | 97.64 ± 3.48 c | |
| S6 | 0.50 ± 0.00 e | 5.18 ± 0.32 f | 7.82 ± 0.37 d | 0.82 ± 0.03 c | 0.70 ± 0.03 e | 3.28 ± 0.03 d | 2.06 ± 0.20 c | 0.70 ± 0.04 c | 65.30 ± 4.59 d | |
| CHL-P | S1 | 4.44 ± 0.08 a | 3.44 ± 0.20 b | 5.63 ± 0.11 c | ND | 1.05 ± 0.04 c | 11.28 ± 0.21 b | 3.13 ± 0.27 c | ND | 5.63 ± 0.01 c |
| S2 | 4.19 ± 0.11 b | 3.47 ± 0.15 b | 5.29 ± 0.16 c | 0.90 ± 0.02 b | 1.40 ± 0.13 ab | 10.89 ± 0.46 b | 2.55 ± 0.15 c | ND | 5.29 ± 0.01 d | |
| S3 | 2.89 ± 0.10 c | 2.77 ± 0.10 c | 10.60 ± 0.33 a | ND | 1.34 ± 0.04 b | 4.98 ± 0.13 d | 11.33 ± 1.53 b | ND | 10.60 ± 0.02 a | |
| S4 | 2.06 ± 0.02 d | 1.47 ± 0.14 d | 3.16 ± 0.31 d | ND | 0.47 ± 0.01 d | 6.70 ± 0.64 c | 2.05 ± 0.04 c | ND | 3.15 ± 0.02 f | |
| S5 | ND | 3.36 ± 0.19 b | 3.65 ± 0.10 d | ND | 1.57 ± 0.02 a | 16.97 ± 0.85 a | 10.66 ± 0.62 b | 10.05±0.47 a | 3.67 ± 0.05 e | |
| S6 | ND | 7.95 ± 0.05 a | 8.07 ± 0.28 b | 18.67 ± 1.36 a | 1.34 ± 0.04 b | 18.37 ± 0.53 a | 17.07 ± 0.78 a | 4.86 ± 0.29 b | 8.02 ± 0.03 b | |
| QCL-F | S1 | ND | 24.79 ± 2.66 a | 14.96 ± 0.79 a | 5.51 ± 0.31 a | 1.94 ± 0.06 a | 8.68 ± 0.23 c | 3.34 ± 0.07 b | ND | 52.02 ± 7.18 a |
| S2 | ND | 20.76 ± 0.39 b | 5.12 ± 0.29 b | 3.69 ± 0.10 b | 1.46 ± 0.07 b | 19.59 ± 0.57 a | 7.15 ± 0.29 a | ND | 33.02 ± 0.79 b | |
| S3 | ND | 9.89 ± 1.25 c | 4.10 ± 0.15 b | 1.03 ± 0.03 c | 1.15 ± 0.04 c | 12.43 ± 0.41 b | 1.68 ± 0.03 c | ND | 26.33 ± 1.93 bc | |
| S4 | ND | 7.08 ± 0.76 c | 2.92 ± 0.31 c | 0.94 ± 0.01 c | 0.83 ± 0.04 d | 7.46 ± 0.08 d | ND | ND | 20.35 ± 2.70 c | |
| S5 | ND | 2.99 ± 0.55 d | 1.64 ± 0.19 d | ND | ND | 3.71 ± 0.33 e | ND | ND | 10.36 ± 1.56 d | |
| S6 | ND | 2.68 ± 0.13 d | 0.91 ± 0.04 de | ND | ND | ND | ND | ND | 6.70 ± 0.35 de | |
| CHL-F | S1 | ND | 1.74 ± 0.31 a | 1.41 ± 0.06 a | 0.83 ± 0.13 a | 0.73 ± 0.02 b | 5.10 ± 0.17 a | 4.46 ± 0.17 a | ND | 4.40 ± 0.22 d |
| S2 | ND | 1.48 ± 0.13 b | 1.05 ± 0.05 b | 1.16 ± 0.01 b | 0.44 ± 0.01 c | 3.50 ± 0.26 b | 2.02 ± 0.12 b | ND | 4.32 ± 0.04 d | |
| S3 | ND | ND | ND | ND | 1.30 ± 0.03 a | ND | 1.49 ± 0.05 c | ND | 35.38 ± 0.01 a | |
| S4 | ND | ND | ND | ND | ND | ND | ND | ND | 6.70 ± 0.12 c | |
| S5 | ND | ND | ND | ND | ND | ND | ND | ND | 11.64 ± 0.01 b | |
| S6 | ND | 2.10 ± 0.20 b | ND | ND | ND | ND | ND | ND | ND | |
| TPC | TFC | TFAC | TMAC | ||
|---|---|---|---|---|---|
| QCL-P | DPPH | 0.950 ** | 0.936 ** | 0.824 ** | 0.094 |
| FRAP | 0.908 ** | 0.495 * | 0.770 ** | −0.582 * | |
| CHL-P | DPPH | 0.891 ** | 0.796 ** | 0.307 | 0.489 * |
| FRAP | 0.913 ** | 0.839 ** | 0.746 ** | 0.395 | |
| QCL-F | DPPH | 0.952 ** | 0.935 ** | 0.888 ** | 0.107 |
| FRAP | 0.938 ** | 0.632 ** | 0.798 ** | −0.610 ** | |
| CHL-F | DPPH | 0.894 ** | 0.929 ** | 0.428 | 0.660 ** |
| FRAP | 0.844 ** | 0.972 ** | 0.944 ** | 0.588 * | |
| PAL1 | PAL3 | C4H | 4CL1 | 4CL2 | CHS | HCT1 | HCT2 | HCT3 | ||
|---|---|---|---|---|---|---|---|---|---|---|
| QCL−P | TPC | −0.819 ** | −0.498 * | −0.161 | −0.124 | −0.525 * | 0.617 ** | 0.697 ** | −0.435 | 0.054 |
| TFC | −0.868 ** | −0.438 | −0.049 | −0.233 | −0.488 * | 0.720 ** | 0.793 ** | −0.358 | −0.193 | |
| TFAC | −0.793 ** | −0.517 * | −0.091 | −0.07 | −0.591 ** | 0.680 ** | 0.703 ** | −0.409 | −0.25 | |
| TMAC | −0.068 | −0.224 | −0.278 | −0.353 | −0.136 | −0.132 | 0.061 | −0.243 | −0.315 | |
| CHL−P | TPC | −0.494 | −0.171 | −0.374 | 0.036 | 0.080 | −0.355 | 0.827 ** | −0.381 | 0.220 |
| TFC | −0.603 | −0.316 | −0.454 | −0.019 | −0.266 | −0.352 | 0.096 | −0.433 | −0.173 | |
| TFAC | −0.488 | −0.365 | −0.440 | 0.218 | −0.131 | −0.442 | 0.415 | −0.484 | 0.253 | |
| TMAC | 0.745 ** | 0.910 ** | 0.891 ** | 0.336 | 0.744 ** | 0.988 ** | −0.299 | 0.959 ** | 0.038 | |
| QCL−F | TPC | −0.069 | 0.857 ** | 0.285 | −0.336 | −0.069 | 0.958 ** | 0.936 ** | 0.182 | −0.136 |
| TFC | 0.237 | 0.888 ** | 0.131 | −0.228 | −0.11 | 0.916 ** | 0.813 ** | 0.074 | 0.199 | |
| TFAC | 0.245 | 0.557 ** | −0.095 | −0.233 | −0.227 | 0.15 | 0.256 | −0.041 | 0.573 * | |
| TMAC | 0.288 | 0.504 * | −0.182 | −0.175 | −0.205 | 0.691 ** | 0.451 | −0.25 | 0.169 | |
| CHL−F | TPC | −0.646 | 0.269 | −0.655 | 0.453 | 0.586 * | 0.249 | 0.774 ** | −0.573 | 0.565 * |
| TFC | −0.575 | 0.398 | −0.508 | 0.633** | 0.683 ** | 0.369 | 0.858 ** | −0.391 | 0.643 ** | |
| TFAC | −0.615 | 0.621 ** | −0.590 | 0.634** | 0.533 * | 0.590 * | 0.849 ** | −0.427 | 0.750 ** | |
| TMAC | −0.163 | 0.300 | −0.006 | 0.507 * | 0.331 | 0.339 | 0.427 | 0.151 | 0.451 |
| HCT1 | HCT2 | HCT3 | ||
|---|---|---|---|---|
| QCL−P | NCA | 0.853 ** | −0.598 ** | 0.066 |
| CA | 0.857 ** | −0.549 ** | 0.095 | |
| CHL−P | NCA | −0.071 | 0.892 ** | 0.167 |
| CA | −0.084 | 0.099 | −0.444 | |
| QCL−F | NCA | 0.763 ** | −0.534 ** | 0.121 |
| CA | 0.964 ** | −0.371 | 0.165 | |
| CHL−F | NCA | 0.753 ** | −0.339 | 0.290 |
| CA | 0.841 ** | −0.325 | 0.331 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Pu, J.; Tang, Y.; Wang, M.; Tian, K.; Wang, Y.; Luo, X.; Deng, Q. Changes in Phenolic Compounds and Antioxidant Activity during Development of ‘Qiangcuili’ and ‘Cuihongli’ Fruit. Foods 2022, 11, 3198. https://doi.org/10.3390/foods11203198
Zhang H, Pu J, Tang Y, Wang M, Tian K, Wang Y, Luo X, Deng Q. Changes in Phenolic Compounds and Antioxidant Activity during Development of ‘Qiangcuili’ and ‘Cuihongli’ Fruit. Foods. 2022; 11(20):3198. https://doi.org/10.3390/foods11203198
Chicago/Turabian StyleZhang, Huifen, Jing Pu, Yan Tang, Miao Wang, Kun Tian, Yongqing Wang, Xian Luo, and Qunxian Deng. 2022. "Changes in Phenolic Compounds and Antioxidant Activity during Development of ‘Qiangcuili’ and ‘Cuihongli’ Fruit" Foods 11, no. 20: 3198. https://doi.org/10.3390/foods11203198