
Lactic Acid Bacteria in Raw-Milk Cheeses: From Starter Cultures to Probiotic Functions


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
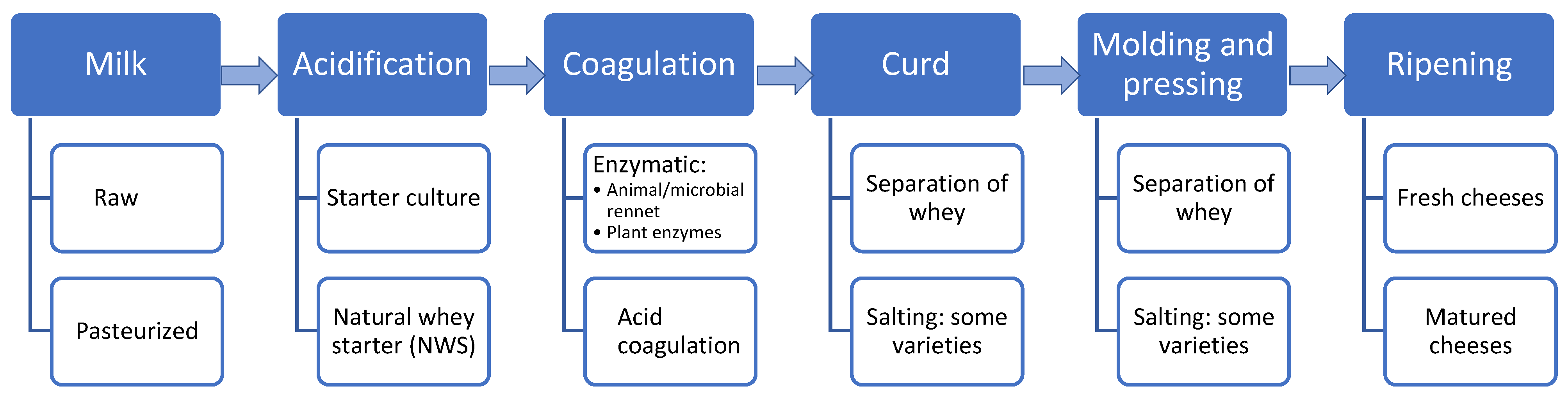
2. Raw-Milk Cheeses
3. LAB Characterization
LAB Taxonomy
4. Main LAB Identified in Raw-Milk Artisanal Cheeses
4.1. Lactococcus
4.2. Lactobacillus
4.3. Enterococcus
4.4. Streptococcus
4.5. Leuconostoc
5. LAB as Starter Cultures
5.1. Type of Starter Cultures
5.1.1. Mesophilic and Thermophilic Starter Cultures
5.1.2. Defined and Undefined Starter Cultures
5.1.3. Natural Whey Starter (NWS)
6. LAB as Adjunct Cultures
6.1. Selected Adjunct Cultures
6.2. Natural Adjunct Cultures
6.3. Charaterization of Adventitious NSLAB
7. Antimicrobial Activity of LAB
7.1. Antibacterial Activity
7.2. Antifungal Activity
7.3. Antimicrobial Metabolites Produced by LAB
7.3.1. Organic Acids
7.3.2. Hydrogen Peroxide
7.3.3. Diacetyl
7.3.4. Reuterin
7.3.5. Fatty Acids
7.3.6. Cyclic Dipeptides
7.3.7. Bacteriocins
8. Probiotic Potential of LAB
8.1. LAB Used as Probiotics
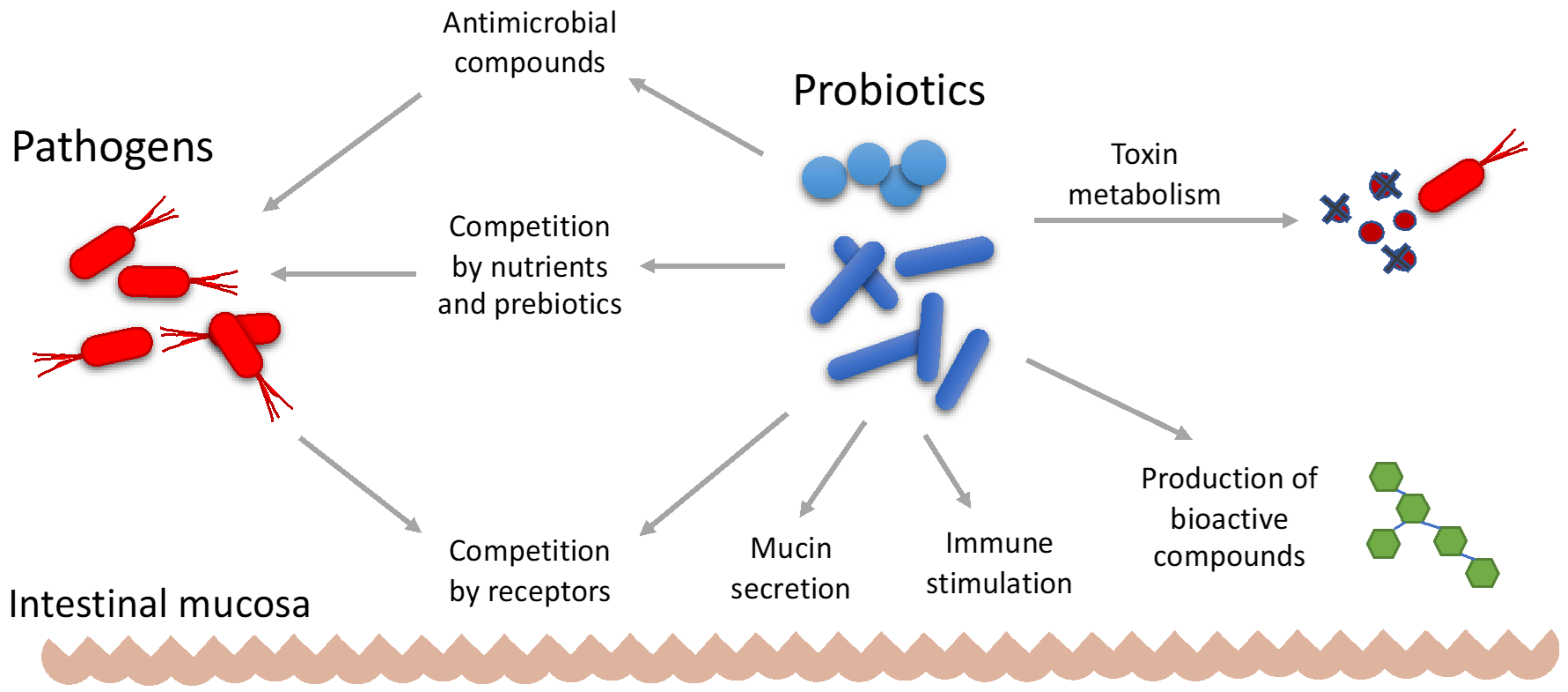
8.2. Mechanisms of Action of Probiotics
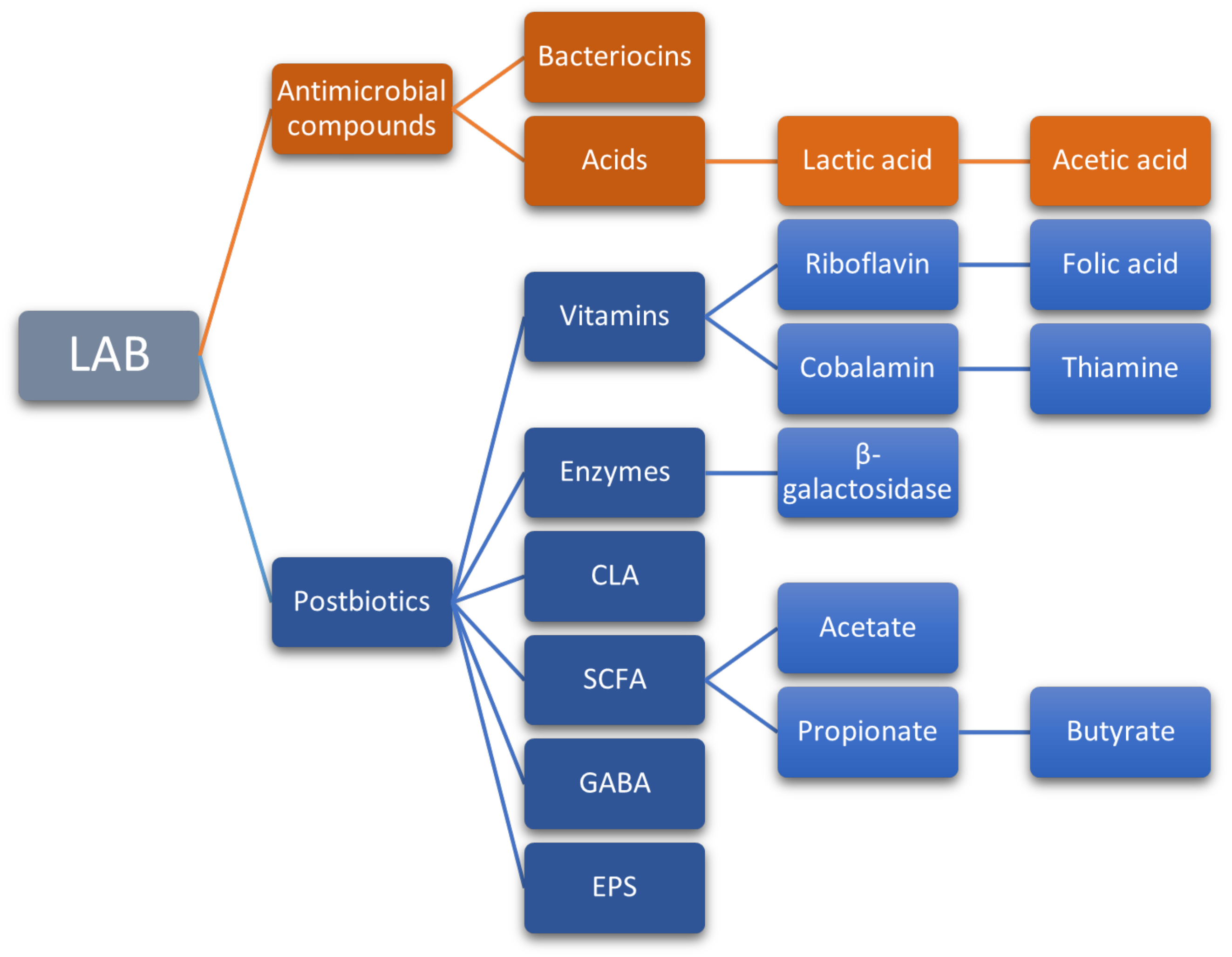
8.3. Bioactive Compounds Produced by Probiotic LAB
8.3.1. Bacteriocins
8.3.2. Short Chain Fatty Acids (SCFA)
8.3.3. Conjugated Linoleic Acid (CLA)
8.3.4. Gamma-Aminobutyric Acid (GABA)
8.3.5. Exopolysaccharides (EPS)
9. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Linares, D.M.; Gómez, C.; Renes, E.; Fresno, J.M.; Tornadijo, M.E.; Ross, R.P.; Stanton, C. Lactic acid bacteria and bifidobacteria with potential to design natural biofunctional health-promoting dairy foods. Front. Microbiol. 2017, 8, 846. [Google Scholar] [CrossRef] [PubMed]
- Shiby, V.; Mishra, H. Fermented milks and milk products as functional foods—A review. Crit. Rev. Food Sci. Nutr. 2013, 53, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Settanni, L.; Moschetti, G. Non-starter lactic acid bacteria used to improve cheese quality and provide health benefits. Food Microbiol. 2010, 27, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Biolcati, F.; Ferrocino, I.; Bottero, M.T.; Dalmasso, A. The Bacterial and Fungal Microbiota of “Robiola di Roccaverano” Protected Designation of Origin Raw Milk Cheese. Front. Microbiol. 2022, 12, 776862. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.H.; Cho, Y.S.; Rackerby, B.; Goddik, L.; Park, S.H. Shifts of microbiota during cheese production: Impact on production and quality. Appl. Microbiol. Biotechnol. 2021, 105, 2307–2318. [Google Scholar] [CrossRef]
- Endres, C.M.; Castro, Í.M.S.; Trevisol, L.D.; Severo, J.M.; Mann, M.B.; Varela, A.P.M.; Frazzon, A.P.G.; Mayer, F.Q.; Frazzon, J. Molecular characterization of the bacterial communities present in sheep’s milk and cheese produced in South Brazilian Region via 16S rRNA gene metabarcoding sequencing. LWT 2021, 147, 111579. [Google Scholar] [CrossRef]
- Biolcati, F.; Ferrocino, I.; Bottero, M.T.; Dalmasso, A. High-throughput sequencing approach to investigate Italian artisanal cheese production. J. Dairy Sci. 2020, 103, 10015–10021. [Google Scholar] [CrossRef]
- Falardeau, J.; Keeney, K.; Trmčić, A.; Kitts, D.; Wang, S. Farm-to-fork profiling of bacterial communities associated with an artisan cheese production facility. Food Microbiol. 2019, 83, 48–58. [Google Scholar] [CrossRef]
- Choi, J.; In Lee, S.; Rackerby, B.; Frojen, R.; Goddik, L.; Ha, S.-D.; Park, S.H. Assessment of overall microbial community shift during Cheddar cheese production from raw milk to aging. Appl. Microbiol. Biotechnol. 2020, 104, 6249–6260. [Google Scholar] [CrossRef]
- Cardinali, F.; Ferrocino, I.; Milanović, V.; Belleggia, L.; Corvaglia, M.R.; Garofalo, C.; Foligni, R.; Mannozzi, C.; Mozzon, M.; Cocolin, L. Microbial communities and volatile profile of Queijo de Azeitão PDO cheese, a traditional Mediterranean thistle-curdled cheese from Portugal. Food Res. Int. 2021, 147, 110537. [Google Scholar] [CrossRef]
- Riquelme, C.; Camara, S.; Dapkevicius, M.; Vinuesa, P.; da Silva, C.C.G.; Malcata, F.X.; Rego, O.A. Characterization of the bacterial biodiversity in Pico cheese (an artisanal Azorean food). Int. J. Food Microbiol. 2015, 192, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Alegría, Á.; Szczesny, P.; Mayo, B.; Bardowski, J.; Kowalczyk, M. Biodiversity in Oscypek, a traditional Polish cheese, determined by culture-dependent and-independent approaches. Appl. Environ. Microbiol. 2012, 78, 1890–1898. [Google Scholar] [CrossRef] [Green Version]
- Ercolini, D.; de Filippis, F.; La Storia, A.; Iacono, M. “Remake” by high-throughput sequencing of the microbiota involved in the production of water buffalo mozzarella cheese. Appl. Environ. Microbiol. 2012, 78, 8142–8145. [Google Scholar] [CrossRef] [Green Version]
- Jurášková, D.; Ribeiro, S.C.; Silva, C.C. Exopolysaccharides Produced by Lactic Acid Bacteria: From Biosynthesis to Health-Promoting Properties. Foods 2022, 11, 156. [Google Scholar] [CrossRef]
- Kanmani, P.; Satish Kumar, R.; Yuvaraj, N.; Paari, K.; Pattukumar, V.; Arul, V. Probiotics and its functionally valuable products—A review. Crit. Rev. Food Sci. Nutr. 2013, 53, 641–658. [Google Scholar] [CrossRef]
- FAO/WHO. Guidelines for the Evaluation of Probiotics in Food; Food and Health Agricultural Organisation of the United Nations: Rome, Italy, 2002. [Google Scholar]
- Coelho, M.C.; Silva, C.C.G.; Ribeiro, S.C.; Dapkevicius, M.; Rosa, H.J.D. Control of Listeria monocytogenes in fresh cheese using protective lactic acid bacteria. Int. J. Food Microbiol. 2014, 191, 53–59. [Google Scholar] [CrossRef]
- Ogawa, J.; Kishino, S.; Ando, A.; Sugimoto, S.; Mihara, K.; Shimizu, S. Production of conjugated fatty acids by lactic acid bacteria. J. Biosci. Bioeng. 2005, 100, 355–364. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, M.Y.; Ji, G.E.; Lee, Y.S.; Hwang, K.T. Production of γ-aminobutyric acid in black raspberry juice during fermentation by Lactobacillus brevis GABA100. Int. J. Food Microbiol. 2009, 130, 12–16. [Google Scholar] [CrossRef]
- Blom, U.; Weréen, P. Cheese and Cheese-Making—With Special Emphasis on Swedish Cheeses. Bioscience-Explained. 2002. Available online: https://bioscience-explained.org/content/cheeseEN.pdf (accessed on 3 March 2022).
- Fox, P.F.; McSweeney, P.L.; Cogan, T.M.; Guinee, T.P. Cheese: Chemistry, Physics and Microbiology, Volume 1: General Aspects; Elsevier: Amsterdam, The Netherlands, 2004. [Google Scholar]
- Reis, P.J.; Malcata, F.X. Current state of Portuguese dairy products from ovine and caprine milks. Small Rumin. Res. 2011, 101, 122–133. [Google Scholar] [CrossRef]
- Høier, E.; Janzen, T.; Rattray, F.; Sørensen, K.; Børsting, M.; Brockmann, E.; Johansen, E. The production, application and action of lactic cheese starter cultures. In Technology of Cheesemaking, 2nd ed.; Law, B.A., Tamime, A.Y., Eds.; Wiley-Blackwell: Chichester, UK, 2010; pp. 166–192. [Google Scholar]
- Quigley, L.; O’Sullivan, O.; Stanton, C.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. The complex microbiota of raw milk. FEMS Microbiol. Rev. 2013, 37, 664–698. [Google Scholar] [CrossRef] [Green Version]
- Fox, P.F.; Guinee, T.P.; Cogan, T.M.; McSweeney, P.L. Fundamentals of Cheese Science; Springer: New York, NY, USA, 2017. [Google Scholar]
- Blaya, J.; Barzideh, Z.; LaPointe, G. Symposium review: Interaction of starter cultures and nonstarter lactic acid bacteria in the cheese environment. J. Dairy Sci. 2018, 101, 3611–3629. [Google Scholar] [CrossRef] [PubMed]
- Gatti, M.; Bottari, B.; Lazzi, C.; Neviani, E.; Mucchetti, G. Invited review: Microbial evolution in raw-milk, long-ripened cheeses produced using undefined natural whey starters. J. Dairy Sci. 2014, 97, 573–591. [Google Scholar] [CrossRef] [Green Version]
- Guinee, T.; O’Brien, B. The quality of milk for cheese manufacture. In Technology of Cheesemaking, 2nd ed.; Law, B.A., Tamime, A.Y., Eds.; Wiley-Blackwell: Chichester, UK, 2010; pp. 1–67. [Google Scholar]
- Montel, M.-C.; Buchin, S.; Mallet, A.; Delbes-Paus, C.; Vuitton, D.A.; Desmasures, N.; Berthier, F. Traditional cheeses: Rich and diverse microbiota with associated benefits. Int. J. Food Microbiol. 2014, 177, 136–154. [Google Scholar] [CrossRef] [PubMed]
- Murtaza, M.A.; Ur-Rehman, S.; Anjum, F.M.; Huma, N.; Hafiz, I. Cheddar cheese ripening and flavor characterization: A review. Crit. Rev. Food Sci. Nutr. 2014, 54, 1309–1321. [Google Scholar] [CrossRef] [PubMed]
- Bottari, B.; Levante, A.; Neviani, E.; Gatti, M. How the fewest become the greatest. L. casei’s impact on long ripened cheeses. Front. Microbiol. 2018, 9, 2866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago-López, L.; Aguilar-Toalá, J.E.; Hernández-Mendoza, A.; Vallejo-Cordoba, B.; Liceaga, A.M.; González-Córdova, A.F. Invited review: Bioactive compounds produced during cheese ripening and health effects associated with aged cheese consumption. J. Dairy Sci. 2018, 101, 3742–3757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inácio, R.S.; Gomes, A.M.; Saraiva, J.A. Serra da Estrela cheese: A review. J. Food Process. Preserv. 2020, 44, e14412. [Google Scholar] [CrossRef]
- Hatti-Kaul, R.; Chen, L.; Dishisha, T.; Enshasy, H.E. Lactic acid bacteria: From starter cultures to producers of chemicals. FEMS Microbiol. Lett. 2018, 365, fny213. [Google Scholar] [CrossRef] [Green Version]
- Axelsson, L. Lactic acid bacteria: Classification and physiology. In Food Science and Technology; Marcel Dekker: New York, NY, USA, 2004; Volume 139, pp. 1–66. [Google Scholar]
- Saris, P.E. Biopreservation by lactic acid bacteria. In Dairy Microbiology and Biochemistry: Recent Developments; CRC Press: Boca Raton, FL, USA, 2014; p. 86. [Google Scholar]
- Bin Masalam, M.S.; Bahieldin, A.; Alharbi, M.G.; Al-Masaudi, S.; Al-Jaouni, S.K.; Harakeh, S.M.; Al-Hindi, R.R. Isolation, molecular characterization and probiotic potential of lactic acid bacteria in Saudi raw and fermented milk. Evid.-Based Complement. Altern. Med. 2018, 2018, 7970463. [Google Scholar] [CrossRef] [Green Version]
- Hippe, B.; Zwielehner, J.; Pirker, A.; Smith, W.M.; Haslberger, A.G. Detection and identification of probiotic microorganisms and other beneficial organisms from the human GI tract. In Probiotics; Springer: New York, NY, USA, 2011; pp. 57–86. [Google Scholar]
- Carr, F.J.; Chill, D.; Maida, N. The lactic acid bacteria: A literature survey. Crit. Rev. Microbiol. 2002, 28, 281–370. [Google Scholar] [CrossRef]
- Schleifer, K.; Ludwig, W. Phylogenetic relationships of lactic acid bacteria. In The Genera of Lactic Acid Bacteria; Springer: New York, NY, USA, 1995; pp. 7–18. [Google Scholar]
- Willey, J.M.; Sherwood, L.; Woolverton, C.J. Prescott’s Principles of Microbiology; McGraw-Hill: New York, NY, USA, 2009. [Google Scholar]
- Leroy, F.; de Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Bintsis, T. Lactic acid bacteria as starter cultures: An update in their metabolism and genetics. AIMS Microbiol. 2018, 4, 665. [Google Scholar] [CrossRef]
- Caplice, E.; Fitzgerald, G.F. Food fermentations: Role of microorganisms in food production and preservation. Int. J. Food Microbiol. 1999, 50, 131–149. [Google Scholar] [CrossRef]
- Topisirovic, L.; Kojic, M.; Fira, D.; Golic, N.; Strahinic, I.; Lozo, J. Potential of lactic acid bacteria isolated from specific natural niches in food production and preservation. Int. J. Food Microbiol. 2006, 112, 230–235. [Google Scholar] [CrossRef]
- Mayo, B.; Aleksandrzak-Piekarczyk, T.; Fernndez, M.; Kowalczyk, M.; Bardowski, J. Updates in the metabolism of lactic acid bacteria. In Biotechnology of Lactic Acid Bacteria: Novel Applications; Mozzi, F., Raya, R.R., Vignolo, G.M., Eds.; Blackwell Publishing: Hoboken, NJ, USA, 2010; pp. 3–33. [Google Scholar]
- Settanni, L.; Franciosi, E.; Cavazza, A.; Cocconcelli, P.S.; Poznanski, E. Extension of Tosèla cheese shelf-life using non-starter lactic acid bacteria. Food Microbiol. 2011, 28, 883–890. [Google Scholar] [CrossRef] [Green Version]
- Powthong, P.; Suntornthiticharoen, P. Isolation, identification and analysis of probiotic properties of lactic acid bacteria from selective various traditional thai fermented food and kefir. Pak. J. Nutr. 2015, 14, 67. [Google Scholar] [CrossRef] [Green Version]
- Collins, B.; Cotter, P.D.; Hill, C.; Ross, R.P. Applications of lactic acid bacteria-produced bacteriocins. In Biotechnology of Lactic Acid Bacteria: Novel Applications; Willey-Blackwell: New York, NY, USA, 2010; pp. 89–109. [Google Scholar]
- Nawaz, M.; Wang, J.; Zhou, A.; Ma, C.; Wu, X.; Moore, J.E.; Cherie Millar, B.; Xu, J. Characterization and transfer of antibiotic resistance in lactic acid bacteria from fermented food products. Curr. Microbiol. 2011, 62, 1081–1089. [Google Scholar] [CrossRef]
- Balciunas, E.M.; Martinez, F.A.C.; Todorov, S.D.; de Melo Franco, B.D.G.; Converti, A.; de Souza Oliveira, R.P. Novel biotechnological applications of bacteriocins: A review. Food Control 2013, 32, 134–142. [Google Scholar] [CrossRef]
- Authority, E.F.S. Introduction of a Qualified Presumption of Safety (QPS) approach for assessment of selected microorganisms referred to EFSA-Opinion of the Scientific Committee. EFSA J. 2007, 5, 587. [Google Scholar]
- Franz, C.; Cho, G.-S.; Holzapfel, W.H.; Gálvez, A. Safety of lactic acid bacteria. In Biotechnology of Lactic Acid Bacteria: Novel Applications; Mozzi, F., Raya, R.R., Vignolo, G.M., Eds.; Blackwell Publishing: Hoboken, NJ, USA, 2010; Volume 2, pp. 341–359. [Google Scholar]
- Von Wright, A.; Axelsson, L. Lactic acid bacteria: An introduction. In Lactic Acid Bacteria: Microbiological and Functional Aspects; Lahtinen, S., Ouwehand, A.C., Salminen, S., von Wright, A., Eds.; CRC Press: Boca Raton, FL, USA, 2011; Volume 4, pp. 1–16. [Google Scholar]
- Mattarelli, P.; Holzapfel, W.; Franz, C.M.; Endo, A.; Felis, G.E.; Hammes, W.; Pot, B.; Dicks, L.; Dellaglio, F. Recommended minimal standards for description of new taxa of the genera Bifidobacterium, Lactobacillus and related genera. Int. J. Syst. Evol. Microbiol. 2014, 64, 1434–1451. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.; Harris, H.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Huang, H.-Y.; Huang, S.-Y.; Chen, P.-Y.; King, V.; Lin, Y.-P.; Tsen, J.-H. Basic characteristics of Sporolactobacillus inulinus BCRC 14647 for potential probiotic properties. Curr. Microbiol. 2007, 54, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Irlinger, F.; Mounier, J. Microbial interactions in cheese: Implications for cheese quality and safety. Curr. Opin. Biotechnol. 2009, 20, 142–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gori, K.; Ryssel, M.; Arneborg, N.; Jespersen, L. Isolation and identification of the microbiota of Danish farmhouse and industrially produced surface-ripened cheeses. Microb. Ecol. 2013, 65, 602–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeluri Jonnala, B.; McSweeney, P.L.; Sheehan, J.J.; Cotter, P.D. Sequencing of the cheese microbiome and its relevance to industry. Front. Microbiol. 2018, 9, 1020. [Google Scholar] [CrossRef] [Green Version]
- Masoud, W.; Takamiya, M.; Vogensen, F.K.; Lillevang, S.; Al-Soud, W.A.; Sørensen, S.J.; Jakobsen, M. Characterization of bacterial populations in Danish raw milk cheeses made with different starter cultures by denaturating gradient gel electrophoresis and pyrosequencing. Int. Dairy J. 2011, 21, 142–148. [Google Scholar] [CrossRef]
- Afshari, R.; Pillidge, C.J.; Read, E.; Rochfort, S.; Dias, D.A.; Osborn, A.M.; Gill, H. New insights into cheddar cheese microbiota-metabolome relationships revealed by integrative analysis of multi-omics data. Sci. Rep. 2020, 10, 3164. [Google Scholar] [CrossRef] [Green Version]
- Nero, L.A.; Andretta, M.; Almeida, T.T.; Ferreira, L.R.; Camargo, A.C.; Yamatogi, R.S.; Carvalho, A.F.; Call, D.R. Lactic microbiota of the minas artisanal cheese produced in the serro region, Minas Gerais, Brazil. LWT 2021, 148, 111698. [Google Scholar] [CrossRef]
- Camargo, A.C.; Costa, E.A.; Fusieger, A.; de Freitas, R.; Nero, L.A.; de Carvalho, A.F. Microbial shifts through the ripening of the “Entre Serras” Minas artisanal cheese monitored by high-throughput sequencing. Food Res. Int. 2021, 139, 109803. [Google Scholar] [CrossRef]
- Teuber, M. The genus Lactococcus. In The Genera of Lactic Acid Bacteria; Wood, B.J., Holzapfel, W., Eds.; Springer Science & Business Media: New York, NY, USA, 1995; Volume 2. [Google Scholar]
- Fox, P.; Guinee, T.; Cogan, T.; McSweeney, P. Fundamentals of Cheese Science; Aspen Publishers Inc.: Gaithersburg, MD, USA, 2000; pp. 98–137. [Google Scholar]
- Noda, S.; Sakamoto, M.; Aihara, C.; Yuki, M.; Katsuhara, M.; Ohkuma, M. Lactococcus termiticola sp. nov., isolated from the gut of the wood-feeding higher termite Nasutitermes takasagoensis. Int. J. Syst. Evol. Microbiol. 2018, 68, 3832–3836. [Google Scholar] [CrossRef]
- Heo, J.; Cho, H.; Tamura, T.; Saitou, S.; Park, K.; Kim, J.-S.; Hong, S.-B.; Kwon, S.-W.; Kim, S.-J. Lactococcus allomyrinae sp. nov., isolated from gut of larvae of Allomyrina dichotoma. Int. J. Syst. Evol. Microbiol. 2019, 69, 3682–3688. [Google Scholar] [CrossRef]
- Vinderola, G.; Ouwehand, A.; Salminen, S.; von Wright, A. Lactic Acid Bacteria: Microbiological and Functional Aspects; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Kongo, J.M.; Ho, A.J.; Malcata, F.X.; Wiedmann, M. Characterization of dominant lactic acid bacteria isolated from Sao Jorge cheese, using biochemical and ribotyping methods. J. Appl. Microbiol. 2007, 103, 1838–1844. [Google Scholar] [CrossRef]
- Lahtinen, S.; Ouwehand, A.C.; Salminen, S.; von Wright, A. Lactic Acid Bacteria: Microbiological And Functional Aspects; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Samedi, L.; Charles, A.L. Isolation and characterization of potential probiotic Lactobacilli from leaves of food plants for possible additives in pellet feeding. Ann. Agric. Sci. 2019, 64, 55–62. [Google Scholar] [CrossRef]
- Bamunuarachchige, T.; Wickramasinghe, H.; Dissanayaka, D.; Wickramarathna, N. Genetic Engineering of Probiotic Microorganisms. In Probiotics; Springer: New York, NY, USA, 2011; pp. 109–138. [Google Scholar]
- Otieno, D.O. Biology of prokaryotic probiotics. In Probiotics; Springer: New York, NY, USA, 2011; pp. 1–28. [Google Scholar]
- Hammes, W.P.; Vogel, R.F. The genus lactobacillus. In The Genera of Lactic Acid Bacteria; Springer: New York, NY, USA, 1995; pp. 19–54. [Google Scholar]
- Barrangou, R.; Lahtinen, S.J.; Ibrahim, F.; Ouwehand, A.C. Genus lactobacillus. In Lactic Acid Bacteria; CRC Press: Boca Raton, FL, USA, 2012; p. 7. [Google Scholar]
- Salvetti, E.; Torriani, S.; Felis, G.E. The genus Lactobacillus: A taxonomic update. Probiot. Antimicrob. Proteins 2012, 4, 217–226. [Google Scholar] [CrossRef]
- Claesson, M.J.; van Sinderen, D.; O’Toole, P.W. The genus Lactobacillus—A genomic basis for understanding its diversity. FEMS Microbiol. Lett. 2007, 269, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Lavanya, K.; Dayakar, Y. Isolation and characterization of probiotic bacteria from the soil samples of the coastal areas of (Gudur division, Nellore Dt.) for utilization in Shrimp farming. Int. J. Fish. Aquafic Stud. 2017, 5, 371–376. [Google Scholar]
- Devriese, L.; Pot, B. The genus enterococcus. In The Genera of Lactic Acid Bacteria; Springer: New York, NY, USA, 1995; pp. 327–367. [Google Scholar]
- Franz, C.; Holzapfel, W.H. The genus Enterococcus: Biotechnological and safety issues. In Food Science and Technology; Salminen, S., von Wright, A., Ouwehand, A., Eds.; CRC Press: Boca Raton, FL, USA, 2004; Volume 139, pp. 199–248. [Google Scholar]
- Giraffa, G. Functionality of enterococci in dairy products. Int. J. Food Microbiol. 2003, 88, 215–222. [Google Scholar] [CrossRef]
- Sarantinopoulos, P.; Kalantzopoulos, G.; Tsakalidou, E. Citrate metabolism by Enterococcus faecalis FAIR-E 229. Appl. Environ. Microbiol. 2001, 67, 5482–5487. [Google Scholar] [CrossRef] [Green Version]
- Wouters, J.T.; Ayad, E.H.; Hugenholtz, J.; Smit, G. Microbes from raw milk for fermented dairy products. Int. Dairy J. 2002, 12, 91–109. [Google Scholar] [CrossRef]
- Cocolin, L.; Foschino, R.; Comi, G.; Fortina, M.G. Description of the bacteriocins produced by two strains of Enterococcus faecium isolated from Italian goat milk. Food Microbiol. 2007, 24, 752–758. [Google Scholar] [CrossRef]
- Hardie, J.; Whiley, R. The genus streptococcus. In The Genera of Lactic Acid Bacteria; Springer: New York, NY, USA, 1995; pp. 55–124. [Google Scholar]
- Tagg, J.R.; Wescombe, P.A.; Burton, J.P.; Wescombe, P. Streptococcus: A brief update on the current taxonomic status of the genus. In Lactic Acid Bacteria: Microbiological and Functional Aspects; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2011; pp. 123–146. [Google Scholar]
- Dellaglio, F.; Dicks, L.; Torriani, S. The genus leuconostoc. In The Genera of Lactic Acid Bacteria; Springer: New York, NY, USA, 1995; pp. 235–278. [Google Scholar]
- Huys, G.; Leisner, J.; Björkroth, J. The lesser LAB gods: Pediococcus, Leuconostoc, Weissella, Carnobacterium, and affiliated genera. In Lactic Acid Bacteria: Microbiological and Functional Aspectspp; Lahtinen, S., Ouwehand, A.C., Salminen, S., von Wright, A., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 93–121. [Google Scholar]
- Hemme, D.; Foucaud-Scheunemann, C. Leuconostoc, characteristics, use in dairy technology and prospects in functional foods. Int. Dairy J. 2004, 14, 467–494. [Google Scholar] [CrossRef]
- Liu, S.-Q.; Asmundson, R.; Holland, R.; Crow, V. Acetaldehyde metabolism by Leuconostoc mesenteroides subsp. cremoris under stress conditions. Int. Dairy J. 1997, 7, 175–183. [Google Scholar] [CrossRef]
- Villani, F.; Aponte, M.; Blaiotta, G.; Mauriello, G.; Pepe, O.; Moschetti, G. Detection and characterization of a bacteriocin, garviecin L1-5, produced by Lactococcus garvieae isolated from raw cow’s milk. J. Appl. Microbiol. 2001, 90, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Villani, F.; Moschetti, G.; Blaiotta, G.; Coppola, S. Characterization of strains of Leuconostoc mesenteroides by analysis of soluble whole-cell protein pattern, DNA fingerprinting and restriction of ribosomal DNA. J. Appl. Microbiol. 1997, 82, 578–588. [Google Scholar] [CrossRef]
- Maina, N.H.; Tenkanen, M.; Maaheimo, H.; Juvonen, R.; Virkki, L. NMR spectroscopic analysis of exopolysaccharides produced by Leuconostoc citreum and Weissella confusa. Carbohydr. Res. 2008, 343, 1446–1455. [Google Scholar] [CrossRef]
- Smid, E.J.; Erkus, O.; Spus, M.; Wolkers-Rooijackers, J.; Alexeeva, S.; Kleerebezem, M. Functional implications of the microbial community structure of undefined mesophilic starter cultures. Microb. Cell Factories 2014, 13, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Rzepkowska, A.; Zielińska, D.; Ołdak, A.; Kołożyn-Krajewska, D. Organic whey as a source of Lactobacillus strains with selected technological and antimicrobial properties. Int. J. Food Sci. Technol. 2017, 52, 1983–1994. [Google Scholar] [CrossRef]
- Beresford, T.P.; Fitzsimons, N.A.; Brennan, N.L.; Cogan, T.M. Recent advances in cheese microbiology. Int. Dairy J. 2001, 11, 259–274. [Google Scholar] [CrossRef]
- Carminati, D.; Giraffa, G.; Quiberoni, A.; Binetti, A.; Suárez, V.; Reinheimer, J. Advances and trends in starter cultures for dairy fermentations. In Biotechnology of Lactic Acid Bacteria: Novel Applications; Mozzi, F., Raya, R.R., Vignolo, G.M., Eds.; Blackwell Publishing: Hoboken, NJ, USA, 2010; Volume 177, pp. 177–192. [Google Scholar]
- Law, B. Cheese-ripening and cheese flavour technology. In Technology of Cheesemaking. Blackwell, MA, USA; Law, B.A., Tamime, A.Y., Eds.; Blackwell Publishing: Hoboken, NJ, USA, 2010; pp. 231–359. [Google Scholar]
- Beresford, T.; Williams, A. The microbiology of cheese ripening. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands, 2004; Volume 1, pp. 287–318. [Google Scholar]
- Fox, P.F.; Cogan, T.M.; Guinee, T.P. Factors that affect the quality of cheese. In Cheese; Elsevier: Amsterdam, The Netherlands, 2017; pp. 617–641. [Google Scholar]
- Pintado, M.; Da Cruz, A.G.; de Sá, P. Cheese microbiology. In Dairy Microbiology and Biochemistry: Recent Developments; Özer, B.H., Akdemir-Evrendilek, G., Eds.; CRC Press: Boca Raton, FL, USA, 2014; Volume 1, pp. 113–133. [Google Scholar]
- Câmara, S.; Dapkevicius, A.; Riquelme, C.; Elias, R.B.; Silva, C.; Malcata, F.X.; Dapkevicius, M. Potential of lactic acid bacteria from Pico cheese for starter culture development. Food Sci. Technol. Int. 2019, 25, 303–317. [Google Scholar] [CrossRef]
- Callanan, M.; Ross, R. Starter cultures: Genetics. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands, 2004; Volume 4, pp. 149–161. [Google Scholar]
- Parente, E.; Cogan, T. Starter cultures: General aspects. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands, 2004; Volume 1, pp. 123–148. [Google Scholar]
- Vázquez-Velázquez, R.; Salvador-Figueroa, M.; Adriano-Anaya, L.; DeGyves–Córdova, G.; Vázquez-Ovando, A. Use of starter culture of native lactic acid bacteria for producing an artisanal Mexican cheese safe and sensory acceptable. CyTA-J. Food 2018, 16, 460–468. [Google Scholar] [CrossRef] [Green Version]
- Ben Braiek, O.; Smaoui, S. Enterococci: Between emerging pathogens and potential probiotics. BioMed Res. Int. 2019, 2019, 5938210. [Google Scholar] [CrossRef] [PubMed]
- Marshall, V.M. Inoculated ecosystems in a milk environment. J. Appl. Bacteriol. 1992, 73, 127s–135s. [Google Scholar] [CrossRef]
- Mayra-Makinen, A.; Bigret, M. Industrial use and production of lactic acid bacteria. In Lactic Acid Bacteria: Microbiological and Functional Aspects; Salminen, S., von Wright, A., Ouwehand, A., Eds.; Marcel Dekker: New York, NY, USA, 2004; Volume 139, pp. 175–198. [Google Scholar]
- Ustunol, Z.; Özer, B.; Akdemir-Evrendilek, G. Dairy starter cultures. In Dairy Microbiology and Biochemistry: Recent Developments; Özer, B.H., Akdemir-Evrendilek, G., Eds.; CRC Press: Boca Raton, FL, USA, 2014; Volume 39. [Google Scholar]
- Kelleher, P.; Murphy, J.; Mahony, J.; van Sinderen, D. Next-generation sequencing as an approach to dairy starter selection. Dairy Sci. Technol. 2015, 95, 545–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.; Law, B. The origins, development and basic operations of cheesemaking technology. In Technology of Cheesemaking, 2nd ed.; Law, B.A., Tamime, A.Y., Eds.; Wiley-Blackwell: Chichester, UK, 2010; pp. 68–97. [Google Scholar]
- Coppola, R.; Nanni, M.; Iorizzo, M.; Sorrentino, A.; Sorrentino, E.; Grazia, L. Survey of lactic acid bacteria isolated during the advanced stages of the ripening of Parmigiano Reggiano cheese. J. Dairy Res. 1997, 64, 305–310. [Google Scholar] [CrossRef]
- Casey, M.G.; Häni, J.P.; Gruskovnjak, J.; Schaeren, W.; Wechsler, D. Characterisation of the non-starter lactic acid bacteria (NSLAB) of Gruyère PDO cheese. Le Lait 2006, 86, 407–414. [Google Scholar] [CrossRef] [Green Version]
- Cogan, T.; Beresford, T.; Steele, J.; Broadbent, J.; Shah, N.; Ustunol, Z. Invited review: Advances in starter cultures and cultured foods. J. Dairy Sci. 2007, 90, 4005–4021. [Google Scholar] [CrossRef] [Green Version]
- De Angelis, M.; Corsetti, A.; Tosti, N.; Rossi, J.; Corbo, M.R.; Gobbetti, M. Characterization of non-starter lactic acid bacteria from Italian ewe cheeses based on phenotypic, genotypic, and cell wall protein analyses. Appl. Environ. Microbiol. 2001, 67, 2011–2020. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.G.; Banks, J.M. Proteolytic and other hydrolytic enzyme activities in non-starter lactic acid bacteria (NSLAB) isolated from Cheddar cheese manufactured in the United Kingdom. Int. Dairy J. 1997, 7, 763–774. [Google Scholar] [CrossRef]
- Fitzsimons, N.A.; Cogan, T.M.; Condon, S.; Beresford, T. Spatial and temporal distribution of non-starter lactic acid bacteria in Cheddar cheese. J. Appl. Microbiol. 2001, 90, 600–608. [Google Scholar] [CrossRef]
- Broadbent, J.R.; Houck, K.; Johnson, M.; Oberg, C. Influence of adjunct use and cheese microenvironment on nonstarter bacteria in reduced-fat Cheddar-type cheese. J. Dairy Sci. 2003, 86, 2773–2782. [Google Scholar] [CrossRef] [Green Version]
- Fitzsimons, N.; Cogan, T.; Condon, S.; Beresford, T. Phenotypic and genotypic characterization of non-starter lactic acid bacteria in mature cheddar cheese. Appl. Environ. Microbiol. 1999, 65, 3418–3426. [Google Scholar] [CrossRef] [Green Version]
- Gobbetti, M.; de Angelis, M.; Di Cagno, R.; Mancini, L.; Fox, P.F. Pros and cons for using non-starter lactic acid bacteria (NSLAB) as secondary/adjunct starters for cheese ripening. Trends Food Sci. Technol. 2015, 45, 167–178. [Google Scholar] [CrossRef]
- Domingos-Lopes, M.; Stanton, C.; Ross, P.; Dapkevicius, M.; Silva, C. Genetic diversity, safety and technological characterization of lactic acid bacteria isolated from artisanal Pico cheese. Food Microbiol. 2017, 63, 178–190. [Google Scholar] [CrossRef]
- Terzić-Vidojević, A.; Mihajlović, S.; Uzelac, G.; Golić, N.; Fira, Đ.; Kojić, M.; Topisirović, L. Identification and characterization of lactic acid bacteria isolated from artisanal white brined Golija cows’ milk cheeses. Arch. Biol. Sci. 2014, 66, 179–192. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Juanes, F.; Teixeira-Martín, V.; González-Buitrago, J.M.; Velázquez, E.; Flores-Félix, J.D. Identification of species and subspecies of lactic acid bacteria present in spanish cheeses type “Torta” by MALDI-TOF MS and pheS gene analyses. Microorganisms 2020, 8, 301. [Google Scholar] [CrossRef] [Green Version]
- Biolcati, F.; Andrighetto, C.; Bottero, M.T.; Dalmasso, A. Microbial characterization of an artisanal production of Robiola di Roccaverano cheese. J. Dairy Sci. 2020, 103, 4056–4067. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Mills, D.A. Facility-specific “house” microbiome drives microbial landscapes of artisan cheesemaking plants. Appl. Environ. Microbiol. 2013, 79, 5214–5223. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Hoover, D. Bacteriocins and their food applications. Compr. Rev. Food Sci. Food Saf. 2003, 2, 82–100. [Google Scholar]
- Galvez, A.; Lopez, R.L.; Abriouel, H.; Valdivia, E.; Omar, N.B. Application of bacteriocins in the control of foodborne pathogenic and spoilage bacteria. Crit. Rev. Biotechnol. 2008, 28, 125–152. [Google Scholar] [CrossRef]
- Pinto, M.S.; de Carvalho, A.F.; dos Santos Pires, A.C.; de Paula, J.C.J.; Sobral, D.; Magalhães, F.A.R. Survival of Listeria innocua in Minas Traditional Serro cheese during ripening. Food Control 2009, 20, 1167–1170. [Google Scholar] [CrossRef]
- Al-Gamal, M.S.; Ibrahim, G.A.; Sharaf, O.M.; Radwan, A.A.; Dabiza, N.M.; Youssef, A.M.; El-Ssayad, M.F. The protective potential of selected lactic acid bacteria against the most common contaminants in various types of cheese in Egypt. Heliyon 2019, 5, e01362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gobbetti, M.; Di Cagno, R.; Calasso, M.; Neviani, E.; Fox, P.F.; de Angelis, M. Drivers that establish and assembly the lactic acid bacteria biota in cheeses. Trends Food Sci. Technol. 2018, 78, 244–254. [Google Scholar] [CrossRef]
- O’Connor, P.M.; Ross, R.P.; Hill, C.; Cotter, P.D. Antimicrobial antagonists against food pathogens: A bacteriocin perspective. Curr. Opin. Food Sci. 2015, 2, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Dalié, D.; Deschamps, A.; Richard-Forget, F. Lactic acid bacteria–Potential for control of mould growth and mycotoxins: A review. Food Control 2010, 21, 370–380. [Google Scholar] [CrossRef]
- Ryan, L.A.; Zannini, E.; Dal Bello, F.; Pawlowska, A.; Koehler, P.; Arendt, E.K. Lactobacillus amylovorus DSM 19280 as a novel food-grade antifungal agent for bakery products. Int. J. Food Microbiol. 2011, 146, 276–283. [Google Scholar] [CrossRef]
- Blagojev, N.; Škrinjar, M.; Vesković-Moračanin, S.; Šošo, V. Control of mould growth and mycotoxin production by lactic acid bacteria metabolites. Rom. Biotechnol. Lett. 2012, 17, 7219–7226. [Google Scholar]
- Sevgi, E.; Tsveteslava, I.-I. Antifungal activity of lactic acid bacteria, isolated from Bulgarian wheat and rye flour. J. Life Sci. 2015, 9, 1–6. [Google Scholar]
- Sedaghat, H.; Eskandari, M.H.; Moosavi-Nasab, M.; Shekarforoush, S.S. Application of non-starter lactic acid bacteria as biopreservative agents to control fungal spoilage of fresh cheese. Int. Dairy J. 2016, 56, 87–91. [Google Scholar] [CrossRef]
- Perczak, A.; Goliński, P.; Bryła, M.; Waśkiewicz, A. The efficiency of lactic acid bacteria against pathogenic fungi and mycotoxins. Arh. Za Hig. Rada I Toksikol. 2018, 69, 32–44. [Google Scholar] [CrossRef] [Green Version]
- Ouiddir, M.; Bettache, G.; Salas, M.L.; Pawtowski, A.; Donot, C.; Brahimi, S.; Mabrouk, K.; Coton, E.; Mounier, J. Selection of Algerian lactic acid bacteria for use as antifungal bioprotective cultures and application in dairy and bakery products. Food Microbiol. 2019, 82, 160–170. [Google Scholar] [CrossRef]
- Hammami, R.; Fliss, I.; Corsetti, A. Application of protective cultures and bacteriocins for food biopreservation. Front. Microbiol. 2019, 10, 1561. [Google Scholar] [CrossRef] [Green Version]
- Benkerroum, N.; Ghouati, Y.; Ghalfi, H.; Elmejdoub, T.; Roblain, D.; Jacques, P.; Thonart, P. Biocontrol of Listeria monocytogenes in a model cultured milk (lben) by in situ bacteriocin production from Lactococcus lactis ssp. lactis. Int. J. Dairy Technol. 2002, 55, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, S.C.; O’Connor, P.M.; Ross, R.P.; Stanton, C.; Silva, C.C.G. An anti-listerial Lactococcus lactis strain isolated from Azorean Pico cheese produces lacticin 481. Int. Dairy J. 2016, 63, 18–28. [Google Scholar] [CrossRef]
- Mills, S.; Serrano, L.M.; Griffin, C.; O’Connor, P.M.; Schaad, G.; Bruining, C.; Hill, C.; Ross, R.P.; Meijer, W.C. Inhibitory activity of Lactobacillus plantarum LMG P-26358 against Listeria innocua when used as an adjunct starter in the manufacture of cheese. Microb. Cell Factories 2011, 10, S7. [Google Scholar] [CrossRef] [Green Version]
- Pingitore, E.V.; Todorov, S.D.; Sesma, F.; de Melo Franco, B.D.G. Application of bacteriocinogenic Enterococcus mundtii CRL35 and Enterococcus faecium ST88Ch in the control of Listeria monocytogenes in fresh Minas cheese. Food Microbiol. 2012, 32, 38–47. [Google Scholar] [CrossRef]
- Vandera, E.; Lianou, A.; Kakouri, A.; Feng, J.; Koukkou, A.-I.; Samelis, J. Enhanced control of Listeria monocytogenes by Enterococcus faecium KE82, a multiple enterocin–producing strain, in different milk environments. J. Food Prot. 2017, 80, 74–85. [Google Scholar] [CrossRef]
- Ribeiro, S.C.; Ross, R.P.; Stanton, C.; Silva, C.C. Characterization and application of antilisterial enterocins on model fresh cheese. J. Food Prot. 2017, 80, 1303–1316. [Google Scholar] [CrossRef]
- Barrios, M.J.; Medina, L.; Lopez, M.C.; Jordano, R. Fungal biota isolated from Spanish cheeses. J. Food Saf. 1998, 18, 151–157. [Google Scholar] [CrossRef]
- Kure, C.F.; Skaar, I. The fungal problem in cheese industry. Curr. Opin. Food Sci. 2019, 29, 14–19. [Google Scholar] [CrossRef]
- Garnier, L.; Valence, F.; Pawtowski, A.; Auhustsinava-Galerne, L.; Frotté, N.; Baroncelli, R.; Deniel, F.; Coton, E.; Mounier, J. Diversity of spoilage fungi associated with various French dairy products. Int. J. Food Microbiol. 2017, 241, 191–197. [Google Scholar] [CrossRef]
- Leyva Salas, M.; Mounier, J.; Valence, F.; Coton, M.; Thierry, A.; Coton, E. Antifungal microbial agents for food biopreservation—A review. Microorganisms 2017, 5, 37. [Google Scholar] [CrossRef] [Green Version]
- Lavermicocca, P.; Valerio, F.; Evidente, A.; Lazzaroni, S.; Corsetti, A.; Gobbetti, M. Purification and characterization of novel antifungal compounds from the sourdough Lactobacillus plantarum strain 21B. Appl. Environ. Microbiol. 2000, 66, 4084–4090. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, B.; Vimont, A.; Desfossés-Foucault, É.; Daga, M.; Arora, G.; Fliss, I. Antifungal activity of lactic and propionic acid bacteria and their potential as protective culture in cottage cheese. Food Control 2017, 78, 350–356. [Google Scholar] [CrossRef]
- Nayyeri, N.; Edalatian Dovom, M.R.; Habibi Najafi, M.B.; Bahreini, M. A Preliminary study on antifungal activity of lactic acid bacteria isolated from different production stages of Lighvan cheese on Penicillium expansum and Rhodotorula mucilaginosa. J. Food Meas. Charact. 2017, 11, 1734–1744. [Google Scholar] [CrossRef]
- Russo, P.; Arena, M.P.; Fiocco, D.; Capozzi, V.; Drider, D.; Spano, G. Lactobacillus plantarum with broad antifungal activity: A promising approach to increase safety and shelf-life of cereal-based products. Int. J. Food Microbiol. 2017, 247, 48–54. [Google Scholar] [CrossRef]
- Shehata, M.G.; Badr, A.N.; El Sohaimy, S.A. Novel antifungal bacteriocin from Lactobacillus paracasei KC39 with anti-mycotoxigenic properties. Biosci. Res. 2018, 15, 4171–4183. [Google Scholar]
- Kim, J.-D. Antifungal activity of lactic acid bacteria isolated from Kimchi against Aspergillus fumigatus. Mycobiology 2005, 33, 210–214. [Google Scholar] [CrossRef] [Green Version]
- Hymery, N.; Vasseur, V.; Coton, M.; Mounier, J.; Jany, J.L.; Barbier, G.; Coton, E. Filamentous fungi and mycotoxins in cheese: A review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 437–456. [Google Scholar] [CrossRef]
- Varsha, K.K.; Nampoothiri, K.M. Appraisal of lactic acid bacteria as protective cultures. Food Control 2016, 69, 61–64. [Google Scholar] [CrossRef]
- Filtenborg, O.; Frisvad, J.C.; Thrane, U. Moulds in food spoilage. Int. J. Food Microbiol. 1996, 33, 85–102. [Google Scholar] [CrossRef]
- Guimarães, A.; Santiago, A.; Teixeira, J.A.; Venâncio, A.; Abrunhosa, L. Anti-aflatoxigenic effect of organic acids produced by Lactobacillus plantarum. Int. J. Food Microbiol. 2018, 264, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnürer, J.; Magnusson, J. Antifungal lactic acid bacteria as biopreservatives. Trends Food Sci. Technol. 2005, 16, 70–78. [Google Scholar] [CrossRef]
- Crowley, S.; Mahony, J.; van Sinderen, D. Current perspectives on antifungal lactic acid bacteria as natural bio-preservatives. Trends Food Sci. Technol. 2013, 33, 93–109. [Google Scholar] [CrossRef]
- Sadiq, F.A.; Yan, B.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. Lactic acid bacteria as antifungal and anti-mycotoxigenic agents: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1403–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, A.; Sato, Y.; Kudo, S.; Sato, S.; Nakajima, H.; Toba, T. The screening of hydrogen peroxide-producing lactic acid bacteria and their application to inactivating psychrotrophic food-borne pathogens. Curr. Microbiol. 2003, 47, 0231–0236. [Google Scholar]
- Hernandez-Valdes, J.A.; Solopova, A.; Kuipers, O.P. Development of Lactococcus lactis Biosensors for Detection of Diacetyl. Front. Microbiol. 2020, 11, 1032. [Google Scholar] [CrossRef]
- Terzic-Vidojevic, A.; Tolinacki, M.; Nikolic, M.; Veljovic, K.; Jovanovic, S.; Macej, O.; Topisirovic, L. Artisanal Vlasina raw goat’s milk cheese: Evaluation and selection of autochthonous lactic acid bacteria as starter cultures. Food Technol. Biotechnol. 2013, 51, 554–563. [Google Scholar]
- Aunsbjerg, S.; Honoré, A.; Marcussen, J.; Ebrahimi, P.; Vogensen, F.; Benfeldt, C.; Skov, T.; Knøchel, S. Contribution of volatiles to the antifungal effect of Lactobacillus paracasei in defined medium and yogurt. Int. J. Food Microbiol. 2015, 194, 46–53. [Google Scholar] [CrossRef]
- Olasupo, N.; Fitzgerald, D.; Gasson, M.; Narbad, A. Activity of natural antimicrobial compounds against Escherichia coli and Salmonella enterica serovar Typhimurium. Lett. Appl. Microbiol. 2003, 37, 448–451. [Google Scholar] [CrossRef] [Green Version]
- Liang, N.; Neužil-Bunešová, V.; Tejnecký, V.; Gänzle, M.; Schwab, C. 3-Hydroxypropionic acid contributes to the antibacterial activity of glycerol metabolism by the food microbe Limosilactobacillus reuteri. Food Microbiol. 2021, 98, 103720. [Google Scholar] [CrossRef]
- Schaefer, L.; Auchtung, T.A.; Hermans, K.E.; Whitehead, D.; Borhan, B.; Britton, R.A. The antimicrobial compound reuterin (3-hydroxypropionaldehyde) induces oxidative stress via interaction with thiol groups. Microbiology 2010, 156, 1589. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Carrillo, J.G.; Orta-Zavalza, E.; González-Rodríguez, S.; Montoya-Torres, C.; Sepúlveda-Ahumada, D.; Ortiz-Rivera, Y. Evaluation of the effectivity of reuterin in pectin edible coatings to extend the shelf-life of strawberries during cold storage. Food Packag. Shelf Life 2021, 30, 100760. [Google Scholar] [CrossRef]
- Bergsson, G.; Arnfinnsson, J.; Steingrímsson, O.l.; Thormar, H. In vitro killing of Candida albicans by fatty acids and monoglycerides. Antimicrob. Agents Chemother. 2001, 45, 3209–3212. [Google Scholar] [CrossRef] [Green Version]
- Black, B.A.; Zannini, E.; Curtis, J.M.; Gänzle, M.G. Antifungal hydroxy fatty acids produced during sourdough fermentation: Microbial and enzymatic pathways, and antifungal activity in bread. Appl. Environ. Microbiol. 2013, 79, 1866–1873. [Google Scholar] [CrossRef] [Green Version]
- Sjögren, J.r.; Magnusson, J.; Broberg, A.; Schnürer, J.; Kenne, L. Antifungal 3-hydroxy fatty acids from Lactobacillus plantarum MiLAB 14. Appl. Environ. Microbiol. 2003, 69, 7554–7557. [Google Scholar] [CrossRef] [Green Version]
- Borthwick, A.D.; Da Costa, N.C. 2, 5-diketopiperazines in food and beverages: Taste and bioactivity. Crit. Rev. Food Sci. Nutr. 2017, 57, 718–742. [Google Scholar] [CrossRef]
- Kwak, M.-K.; Liu, R.; Kang, S.-O. Antimicrobial activity of cyclic dipeptides produced by Lactobacillus plantarum LBP-K10 against multidrug-resistant bacteria, pathogenic fungi, and influenza A virus. Food Control 2018, 85, 223–234. [Google Scholar] [CrossRef]
- Dal Bello, F.; Clarke, C.; Ryan, L.; Ulmer, H.; Schober, T.; Ström, K.; Sjögren, J.; van Sinderen, D.; Schnürer, J.; Arendt, E. Improvement of the quality and shelf life of wheat bread by fermentation with the antifungal strain Lactobacillus plantarum FST 1.7. J. Cereal Sci. 2007, 45, 309–318. [Google Scholar] [CrossRef]
- O’sullivan, L.; Ross, R.; Hill, C. Potential of bacteriocin-producing lactic acid bacteria for improvements in food safety and quality. Biochimie 2002, 84, 593–604. [Google Scholar] [CrossRef]
- Soomro, A.; Masud, T.; Anwaar, K. Role of lactic acid bacteria (LAB) in food preservation and human health—A review. Pak. J. Nutr. 2002, 1, 20–24. [Google Scholar]
- Settanni, L.; Corsetti, A. Application of bacteriocins in vegetable food biopreservation. Int. J. Food Microbiol. 2008, 121, 123–138. [Google Scholar] [CrossRef]
- Mills, S.; Stanton, C.; Hill, C.; Ross, R. New developments and applications of bacteriocins and peptides in foods. Annu. Rev. Food Sci. Technol. 2011, 2, 299–329. [Google Scholar] [CrossRef]
- Ha, T.M.; Shakur, S.; Do, K.H.P. Consumer concern about food safety in Hanoi, Vietnam. Food Control 2019, 98, 238–244. [Google Scholar] [CrossRef]
- Cotter, P.D.; Hill, C.; Ross, R.P. Bacteriocins: Developing innate immunity for food. Nat. Rev. Microbiol. 2005, 3, 777–788. [Google Scholar] [CrossRef]
- Mirhosseini, M.; Nahvi, I.; Emtiazi, G.; Tavassoli, M. Characterisation of anti-Listeria monocytogenes bacteriocins from Enterococcus faecium strains isolated from dairy products. Int. J. Dairy Technol. 2010, 63, 55–61. [Google Scholar] [CrossRef]
- Kaškonienė, V.; Stankevičius, M.; Bimbiraitė-Survilienė, K.; Naujokaitytė, G.; Šernienė, L.; Mulkytė, K.; Malakauskas, M.; Maruška, A. Current state of purification, isolation and analysis of bacteriocins produced by lactic acid bacteria. Appl. Microbiol. Biotechnol. 2017, 101, 1323–1335. [Google Scholar] [CrossRef]
- Hassan, M.; Kjos, M.; Nes, I.; Diep, D.; Lotfipour, F. Natural antimicrobial peptides from bacteria: Characteristics and potential applications to fight against antibiotic resistance. J. Appl. Microbiol. 2012, 113, 723–736. [Google Scholar] [CrossRef]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef]
- Cleveland, J.; Montville, T.J.; Nes, I.F.; Chikindas, M.L. Bacteriocins: Safe, natural antimicrobials for food preservation. Int. J. Food Microbiol. 2001, 71, 1–20. [Google Scholar] [CrossRef]
- Leite, J.A.; Tulini, F.L.; dos Reis-Teixeira, F.B.; Rabinovitch, L.; Chaves, J.Q.; Rosa, N.G.; Cabral, H.; de Martinis, E.C.P. Bacteriocin-like inhibitory substances (BLIS) produced by Bacillus cereus: Preliminary characterization and application of partially purified extract containing BLIS for inhibiting Listeria monocytogenes in pineapple pulp. LWT Food Sci. Technol. 2016, 72, 261–266. [Google Scholar] [CrossRef]
- Piard, J.; Desmazeaud, M. Inhibiting factors produced by lactic acid bacteria. 2. Bacteriocins and other antibacterial substances. Le Lait 1992, 72, 113–142. [Google Scholar] [CrossRef] [Green Version]
- Lozo, J.; Vukasinovic, M.; Strahinic, I.; Topisirovic, L. Characterization and antimicrobial activity of bacteriocin 217 produced by natural isolate Lactobacillus paracasei subsp. paracasei BGBUK2-16. J. Food Prot. 2004, 67, 2727–2734. [Google Scholar] [CrossRef] [PubMed]
- Sobrino-López, A.; Martín-Belloso, O. Use of nisin and other bacteriocins for preservation of dairy products. Int. Dairy J. 2008, 18, 329–343. [Google Scholar] [CrossRef]
- Bali, V.; Panesar, P.S.; Bera, M.B.; Kennedy, J.F. Bacteriocins: Recent trends and potential applications. Crit. Rev. Food Sci. Nutr. 2016, 56, 817–834. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.C.; Silva, S.P.; Ribeiro, S.C. Application of bacteriocins and protective cultures in dairy food preservation. Front. Microbiol. 2018, 9, 594. [Google Scholar] [CrossRef]
- Silva, S.P.; Ribeiro, S.C.; Teixeira, J.A.; Silva, C.C. Application of an alginate-based edible coating with bacteriocin-producing Lactococcus strains in fresh cheese preservation. LWT 2022, 153, 112486. [Google Scholar] [CrossRef]
- Malini, M.; Savitha, J. Heat stable bacteriocin from Lactobacillus paracasei subsp. tolerans isolated from locally available cheese: An in vitro study. E3 J. Biotechnol. Pharm. Res. 2012, 3, 28–41. [Google Scholar]
- Lozo, J.; Topisirovic, L.; Kojic, M. Natural bacterial isolates as an inexhaustible source of new bacteriocins. Appl. Microbiol. Biotechnol. 2021, 105, 477–492. [Google Scholar] [CrossRef]
- Jena, P.K.; Trivedi, D.; Thakore, K.; Chaudhary, H.; Giri, S.S.; Seshadri, S. Isolation and characterization of probiotic properties of lactobacilli isolated from rat fecal microbiota. Microbiol. Immunol. 2013, 57, 407–416. [Google Scholar] [CrossRef]
- Colombo, M.; Castilho, N.; Todorov, S.D.; Nero, L.A. Beneficial properties of lactic acid bacteria naturally present in dairy production. BMC Microbiol. 2018, 18, 219. [Google Scholar] [CrossRef]
- Celiberto, L.S.; Pinto, R.A.; Rossi, E.A.; Vallance, B.A.; Cavallini, D.C. Isolation and characterization of potentially probiotic bacterial strains from mice: Proof of concept for personalized probiotics. Nutrients 2018, 10, 1684. [Google Scholar] [CrossRef] [Green Version]
- Żukiewicz-Sobczak, W.; Wróblewska, P.; Adamczuk, P.; Silny, W. Probiotic lactic acid bacteria and their potential in the prevention and treatment of allergic diseases. Cent.-Eur. J. Immunol. 2014, 39, 104. [Google Scholar] [CrossRef]
- Guarner, F.; Khan, A.G.; Garisch, J.; Eliakim, R.; Gangl, A.; Thomson, A.; Krabshuis, J.; Lemair, T.; Kaufmann, P.; de Paula, J.A. World gastroenterology organisation global guidelines: Probiotics and prebiotics october 2011. J. Clin. Gastroenterol. 2012, 46, 468–481. [Google Scholar] [CrossRef]
- Kook, S.-Y.; Chung, E.-C.; Lee, Y.; Lee, D.W.; Kim, S. Isolation and characterization of five novel probiotic strains from Korean infant and children faeces. PLoS ONE 2019, 14, e0223913. [Google Scholar] [CrossRef]
- Holzapfel, W.H.; Haberer, P.; Geisen, R.; Björkroth, J.; Schillinger, U. Taxonomy and important features of probiotic microorganisms in food and nutrition. Am. J. Clin. Nutr. 2001, 73, 365s–373s. [Google Scholar] [CrossRef] [Green Version]
- Azat, R.; Liu, Y.; Li, W.; Kayir, A.; Lin, D.-b.; Zhou, W.-w.; Zheng, X.-d. Probiotic properties of lactic acid bacteria isolated from traditionally fermented Xinjiang cheese. J. Zhejiang Univ. Sci. B 2016, 17, 597–609. [Google Scholar] [CrossRef] [Green Version]
- Blaabjerg, S.; Artzi, D.M.; Aabenhus, R. Probiotics for the prevention of antibiotic-associated diarrhea in outpatients—A systematic review and meta-analysis. Antibiotics 2017, 6, 21. [Google Scholar] [CrossRef] [Green Version]
- Eslami, M.; Yousefi, B.; Kokhaei, P.; Hemati, M.; Nejad, Z.R.; Arabkari, V.; Namdar, A. Importance of probiotics in the prevention and treatment of colorectal cancer. J. Cell. Physiol. 2019, 234, 17127–17143. [Google Scholar] [CrossRef]
- Sivamaruthi, B.S.; Kesika, P.; Chaiyasut, C. A mini-review of human studies on cholesterol-lowering properties of probiotics. Sci. Pharm. 2019, 87, 26. [Google Scholar] [CrossRef] [Green Version]
- Sivamaruthi, B.S.; Bharathi, M.; Kesika, P.; Suganthy, N.; Chaiyasut, C. The Administration of Probiotics against Hypercholesterolemia: A Systematic Review. Appl. Sci. 2021, 11, 6913. [Google Scholar] [CrossRef]
- Domingos-Lopes, M.; Stanton, C.; Ross, R.; Silva, C. Histamine and cholesterol lowering abilities of lactic acid bacteria isolated from artisanal Pico cheese. J. Appl. Microbiol. 2020, 129, 1428–1440. [Google Scholar] [CrossRef]
- Sivamaruthi, B.S.; Prasanth, M.I.; Kesika, P.; Chaiyasut, C. Probiotics in human mental health and diseases—A minireview. Trop. J. Pharm. Res. 2019, 18, 889–895. [Google Scholar] [CrossRef]
- Zielińska, D.; Kolożyn-Krajewska, D. Food-origin lactic acid bacteria may exhibit probiotic properties. BioMed Res. Int. 2018, 2018, 1784. [Google Scholar] [CrossRef] [Green Version]
- Salminen, S.; Gueimonde, M. Human studies on probiotics: What is scientifically proven. J. Food Sci. 2004, 69, M137–M140. [Google Scholar] [CrossRef]
- Abdullah-Al-Mamun, M.; Rahman, S. Assessment of Probiotic Properties of Isolated Lactic Acid Bacteria from Human Milk Sample. Adv. Biores. 2017, 8, 140–146. [Google Scholar]
- Reuben, R.C.; Roy, P.C.; Sarkar, S.L.; Alam, R.-U.; Jahid, I.K. Isolation, characterization, and assessment of lactic acid bacteria toward their selection as poultry probiotics. BMC Microbiol. 2019, 19, 253. [Google Scholar] [CrossRef] [Green Version]
- Padmavathi, T.; Bhargavi, R.; Priyanka, P.R.; Niranjan, N.R.; Pavitra, P.V. Screening of potential probiotic lactic acid bacteria and production of amylase and its partial purification. J. Genet. Eng. Biotechnol. 2018, 16, 357–362. [Google Scholar] [CrossRef]
- Gao, J.; Li, X.; Zhang, G.; Sadiq, F.A.; Simal-Gandara, J.; Xiao, J.; Sang, Y. Probiotics in the dairy industry—Advances and opportunities. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3937–3982. [Google Scholar] [CrossRef]
- Doğan, M.; Ay, M. Evaluation of the probiotic potential of Pediococcus strains from fermented dairy product kefir. Czech J. Food Sci. 2021, 39, 376–383. [Google Scholar] [CrossRef]
- Petrova, P.; Ivanov, I.; Tsigoriyna, L.; Valcheva, N.; Vasileva, E.; Parvanova-Mancheva, T.; Arsov, A.; Petrov, K. Traditional Bulgarian dairy products: Ethnic foods with health benefits. Microorganisms 2021, 9, 480. [Google Scholar] [CrossRef] [PubMed]
- Somashekaraiah, R.; Shruthi, B.; Deepthi, B.; Sreenivasa, M. Probiotic properties of lactic acid bacteria isolated from neera: A naturally fermenting coconut palm nectar. Front. Microbiol. 2019, 10, 1382. [Google Scholar] [CrossRef] [PubMed]
- Mulaw, G.; Sisay Tessema, T.; Muleta, D.; Tesfaye, A. In vitro evaluation of probiotic properties of lactic acid bacteria isolated from some traditionally fermented Ethiopian food products. Int. J. Microbiol. 2019, 2019, 7179514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, H.S.; Grover, S.; Batish, V.K.; Gill, P. Immunological effects of probiotics and their significance to human health. In Prebiotics and Probiotics Science and Technology; Springer: New York, NY, USA, 2009; p. 901. [Google Scholar]
- Felis, G.E.; Dellaglio, F.; Torriani, S. Taxonomy of probiotic microorganisms. In Prebiotics and Probiotics Science and Technology; Springer: New York, NY, USA, 2009; pp. 591–637. [Google Scholar]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Jeevaratnam, K.; Fatima, A. Lactic Acid Bacteria: Probiotic Characteristic, Selection Criteria, and its Role in Human Health (A Review). Int. J. Emerg. Technol. Innov. Res. 2018, 5, 10. [Google Scholar]
- Siró, I. Challenges of Beneficial Health Claims. In Probiotics: Biology, Genetics and Health Aspects, Microbioloy Monographs; Liong, M.T., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 243–268. [Google Scholar]
- Harzallah, D.; Belhadj, H. Lactic acid bacteria as probiotics: Characteristics, selection criteria and role in immunomodulation of human GI muccosal barrier. In Lactic Acid Bacteria—R & D for Food, Health and Livestock Purposes; Kongo, M., Ed.; InTech: Zagreb, Croatia, 2013; pp. 197–216. [Google Scholar]
- Lawrence, G.W.; Begley, M.; Cotter, P.D.; Guinane, C.M. Potential use of biotherapeutic bacteria to target colorectal cancer-associated taxa. Int. J. Mol. Sci. 2020, 21, 924. [Google Scholar] [CrossRef] [Green Version]
- Erickson, K.L.; Hubbard, N.E. Probiotic immunomodulation in health and disease. J. Nutr. 2000, 130, 403S–409S. [Google Scholar] [CrossRef]
- Kanmani, P.; Kim, H. Immunobiotic strains modulate Toll-like receptor 3 agonist induced innate antiviral immune response in human intestinal epithelial cells by modulating IFN regulatory factor 3 and NF-κB signaling. Front. Immunol. 2019, 10, 1536. [Google Scholar] [CrossRef]
- Domingos-Lopes, M.; Nagy, A.; Stanton, C.; Ross, P.; Gelencsér, E.; Silva, C. Immunomodulatory activity of exopolysaccharide producing Leuconostoc citreum strain isolated from Pico cheese. J. Funct. Foods 2017, 33, 235–243. [Google Scholar] [CrossRef]
- Ramalho, J.B.; Spiazzi, C.C.; Bicca, D.F.; Rodrigues, J.F.; Sehn, C.P.; da Silva, W.P.; Cibin, F.W.S. Beneficial effects of Lactococcus lactis subsp. cremoris LL95 treatment in an LPS-induced depression-like model in mice. Behav. Brain Res. 2022, 426, 113847. [Google Scholar] [CrossRef]
- Saliganti, V.; Kapila, R.; Kapila, S.; Bhat, M.I. Probiotics in the modulation of maternal–infant immunity: Implications for allergic diseases. Food Rev. Int. 2017, 33, 516–537. [Google Scholar] [CrossRef]
- Dinan, T.G.; Stilling, R.M.; Stanton, C.; Cryan, J.F. Collective unconscious: How gut microbes shape human behavior. J. Psychiatr. Res. 2015, 63, 1–9. [Google Scholar] [CrossRef] [Green Version]
- O’Mahony, S.M.; Clarke, G.; Borre, Y.; Dinan, T.G.; Cryan, J. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav. Brain Res. 2015, 277, 32–48. [Google Scholar] [CrossRef]
- Misra, S.; Mohanty, D. Psychobiotics: A new approach for treating mental illness? Crit. Rev. Food Sci. Nutr. 2019, 59, 1230–1236. [Google Scholar] [CrossRef]
- Sarkar, A.; Lehto, S.M.; Harty, S.; Dinan, T.G.; Cryan, J.F.; Burnet, P.W. Psychobiotics and the manipulation of bacteria–gut–brain signals. Trends Neurosci. 2016, 39, 763–781. [Google Scholar] [CrossRef] [Green Version]
- Oleskin, A.V.; Shenderov, B.A. Probiotics and psychobiotics: The role of microbial neurochemicals. Probiot. Antimicrob. Proteins 2019, 11, 1071–1085. [Google Scholar] [CrossRef]
- Tyagi, P.; Tasleem, M.; Prakash, S.; Chouhan, G. Intermingling of gut microbiota with brain: Exploring the role of probiotics in battle against depressive disorders. Food Res. Int. 2020, 137, 109489. [Google Scholar] [CrossRef]
- Barros, C.P.; Guimaraes, J.T.; Esmerino, E.A.; Duarte, M.C.K.; Silva, M.C.; Silva, R.; Ferreira, B.M.; Sant’Ana, A.S.; Freitas, M.Q.; Cruz, A.G. Paraprobiotics and postbiotics: Concepts and potential applications in dairy products. Curr. Opin. Food Sci. 2020, 32, 1–8. [Google Scholar] [CrossRef]
- Siragusa, S.; de Angelis, M.; Di Cagno, R.; Rizzello, C.G.; Coda, R.; Gobbetti, M. Synthesis of γ-aminobutyric acid by lactic acid bacteria isolated from a variety of Italian cheeses. Appl. Environ. Microbiol. 2007, 73, 7283–7290. [Google Scholar] [CrossRef] [Green Version]
- Alhaj, O.; Kanekanian, A. Milk-derived bioactive components from fermentation. In Milk and Dairy Products as Functional Foods, First Edition; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2014; pp. 237–288. [Google Scholar]
- Şanlier, N.; Gökcen, B.B.; Sezgin, A.C. Health benefits of fermented foods. Crit. Rev. Food Sci. Nutr. 2019, 59, 506–527. [Google Scholar] [CrossRef] [PubMed]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M. Functions and emerging applications of bacteriocins. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef]
- Hegarty, J.W.; Guinane, C.M.; Ross, R.P.; Hill, C.; Cotter, P.D. Bacteriocin production: A relatively unharnessed probiotic trait? F1000Research 2016, 5, 2587. [Google Scholar] [CrossRef]
- Alvarez-Sieiro, P.; Montalbán-López, M.; Mu, D.; Kuipers, O.P. Bacteriocins of lactic acid bacteria: Extending the family. Appl. Microbiol. Biotechnol. 2016, 100, 2939–2951. [Google Scholar] [CrossRef] [Green Version]
- Kommineni, S.; Bretl, D.J.; Lam, V.; Chakraborty, R.; Hayward, M.; Simpson, P.; Cao, Y.; Bousounis, P.; Kristich, C.J.; Salzman, N.H. Bacteriocin production augments niche competition by enterococci in the mammalian gastrointestinal tract. Nature 2015, 526, 719–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, S.; Kaur, S. Bacteriocins as potential anticancer agents. Front. Pharmacol. 2015, 6, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baindara, P.; Korpole, S.; Grover, V. Bacteriocins: Perspective for the development of novel anticancer drugs. Appl. Microbiol. Biotechnol. 2018, 102, 10393–10408. [Google Scholar] [CrossRef] [PubMed]
- Shuwen, H.; Miao, D.; Quan, Q.; Wei, W.; Zhongshan, Z.; Chun, Z.; Xi, Y. Protective effect of the “food-microorganism-SCFAs” axis on colorectal cancer: From basic research to practical application. J. Cancer Res. Clin. Oncol. 2019, 145, 2169–2197. [Google Scholar] [CrossRef] [PubMed]
- Brahe, L.; Astrup, A.; Larsen, L. Is butyrate the link between diet, intestinal microbiota and obesity-related metabolic diseases? Obes. Rev. 2013, 14, 950–959. [Google Scholar] [CrossRef]
- Indira, M.; Venkateswarulu, T.; Abraham Peele, K.; Bobby, N.; Krupanidhi, S. Bioactive molecules of probiotic bacteria and their mechanism of action: A review. 3 Biotech 2019, 9, 306. [Google Scholar] [CrossRef]
- Stilling, R.M.; van de Wouw, M.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. The neuropharmacology of butyrate: The bread and butter of the microbiota-gut-brain axis? Neurochem. Int. 2016, 99, 110–132. [Google Scholar] [CrossRef]
- Liu, H.; Wang, J.; He, T.; Becker, S.; Zhang, G.; Li, D.; Ma, X. Butyrate: A double-edged sword for health? Adv. Nutr. 2018, 9, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Meijer, K.; de Vos, P.; Priebe, M.G. Butyrate and other short-chain fatty acids as modulators of immunity: What relevance for health? Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 715–721. [Google Scholar] [CrossRef]
- Shokryazdan, P.; Rajion, M.A.; Meng, G.Y.; Boo, L.J.; Ebrahimi, M.; Royan, M.; Sahebi, M.; Azizi, P.; Abiri, R.; Jahromi, M.F. Conjugated linoleic acid: A potent fatty acid linked to animal and human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 2737–2748. [Google Scholar] [CrossRef]
- O’Shea, E.F.; Cotter, P.D.; Stanton, C.; Ross, R.P.; Hill, C. Production of bioactive substances by intestinal bacteria as a basis for explaining probiotic mechanisms: Bacteriocins and conjugated linoleic acid. Int. J. Food Microbiol. 2012, 152, 189–205. [Google Scholar] [CrossRef]
- Gong, M.; Hu, Y.; Wei, W.; Jin, Q.; Wang, X. Production of conjugated fatty acids: A review of recent advances. Biotechnol. Adv. 2019, 37, 107454. [Google Scholar] [CrossRef]
- Coakley, M.; Ross, R.; Nordgren, M.; Fitzgerald, G.; Devery, R.; Stanton, C. Conjugated linoleic acid biosynthesis by human-derived Bifidobacterium species. J. Appl. Microbiol. 2003, 94, 138–145. [Google Scholar] [CrossRef]
- El Roz, A.; Bard, J.; Huvelin, J.; Nazih, H. The anti-proliferative and pro-apoptotic effects of the trans9, trans11 conjugated linoleic acid isomer on MCF-7 breast cancer cells are associated with LXR activation. Prostaglandins Leukot. Essent. Fat. Acids 2013, 88, 265–272. [Google Scholar] [CrossRef]
- Kishino, S.; Takeuchi, M.; Park, S.-B.; Hirata, A.; Kitamura, N.; Kunisawa, J.; Kiyono, H.; Iwamoto, R.; Isobe, Y.; Arita, M. Polyunsaturated fatty acid saturation by gut lactic acid bacteria affecting host lipid composition. Proc. Natl. Acad. Sci. USA 2013, 110, 17808–17813. [Google Scholar] [CrossRef] [Green Version]
- Barrett, E.; Ross, R.; Fitzgerald, G.; Stanton, C. Rapid screening method for analyzing the conjugated linoleic acid production capabilities of bacterial cultures. Appl. Environ. Microbiol. 2007, 73, 2333–2337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renes, E.; Linares, D.; González, L.; Fresno, J.; Tornadijo, M.; Stanton, C. Production of conjugated linoleic acid and gamma-aminobutyric acid by autochthonous lactic acid bacteria and detection of the genes involved. J. Funct. Foods 2017, 34, 340–346. [Google Scholar] [CrossRef]
- Fuke, G.; Nornberg, J.L. Systematic evaluation on the effectiveness of conjugated linoleic acid in human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 1–7. [Google Scholar] [CrossRef]
- Kuhl, G.C.; de Dea Lindner, J. Biohydrogenation of linoleic acid by lactic acid bacteria for the production of functional cultured dairy products: A review. Foods 2016, 5, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sosa-Castañeda, J.; Hernández-Mendoza, A.; Astiazarán-García, H.; Garcia, H.; Estrada-Montoya, M.; González-Córdova, A.; Vallejo-Cordoba, B. Screening of Lactobacillus strains for their ability to produce conjugated linoleic acid in milk and to adhere to the intestinal tract. J. Dairy Sci. 2015, 98, 6651–6659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, S.C.; Stanton, C.; Yang, B.; Ross, R.P.; Silva, C.C. Conjugated linoleic acid production and probiotic assessment of Lactobacillus plantarum isolated from Pico cheese. LWT 2018, 90, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Thwe, S.M.; Kobayashi, T.; Luan, T.; Shirai, T.; Onodera, M.; Hamada-Sato, N.; Imada, C. Isolation, characterization, and utilization of γ-aminobutyric acid (GABA)-producing lactic acid bacteria from Myanmar fishery products fermented with boiled rice. Fish. Sci. 2011, 77, 279–288. [Google Scholar] [CrossRef]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-aminobutyric acid as a bioactive compound in foods: A review. J. Funct. Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Yu, H.-H.; Choi, J.H.; Kang, K.M.; Hwang, H.-J. Potential of a lactic acid bacterial starter culture with gamma-aminobutyric acid (GABA) activity for production of fermented sausage. Food Sci. Biotechnol. 2017, 26, 1333–1341. [Google Scholar] [CrossRef]
- Lee, K.W.; Shim, J.M.; Yao, Z.; Kim, J.A.; Kim, H.-J.; Kim, J.H. Characterization of a glutamate decarboxylase (GAD) from Enterococcus avium M5 isolated from Jeotgal, a Korean fermented seafood. J. Microbiol. Biotechnol. 2017, 27, 1216–1222. [Google Scholar] [CrossRef]
- Cui, Y.; Miao, K.; Niyaphorn, S.; Qu, X. Production of gamma-aminobutyric acid from lactic acid bacteria: A systematic review. Int. J. Mol. Sci. 2020, 21, 995. [Google Scholar] [CrossRef] [Green Version]
- Mazzoli, R.; Pessione, E. The neuro-endocrinological role of microbial glutamate and GABA signaling. Front. Microbiol. 2016, 7, 1934. [Google Scholar] [CrossRef] [Green Version]
- Barrett, E.; Ross, R.; O’Toole, P.W.; Fitzgerald, G.F.; Stanton, C. γ-Aminobutyric acid production by culturable bacteria from the human intestine. J. Appl. Microbiol. 2012, 113, 411–417. [Google Scholar] [CrossRef]
- Franciosi, E.; Carafa, I.; Nardin, T.; Schiavon, S.; Poznanski, E.; Cavazza, A.; Larcher, R.; Tuohy, K.M. Biodiversity and γ-aminobutyric acid production by lactic acid bacteria isolated from traditional alpine raw cow’s milk cheeses. BioMed Res. Int. 2015, 2015, 625740. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Han, X.; Cen, S.; Duan, H.; Feng, S.; Xue, Y.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q. Beneficial effect of GABA-rich fermented milk on insomnia involving regulation of gut microbiota. Microbiol. Res. 2020, 233, 126409. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, R.; Zhang, S.; Zhang, X.; Zhou, H.; Wen, T.; Wang, J. Depression-like symptoms due to Dcf1 deficiency are alleviated by intestinal transplantation of Lactobacillus murine and Lactobacillus reuteri. Biochem. Biophys. Res. Commun. 2022, 593, 137–143. [Google Scholar] [CrossRef]
- Ko, C.Y.; Lin, H.-T.V.; Tsai, G.J. Gamma-aminobutyric acid production in black soybean milk by Lactobacillus brevis FPA 3709 and the antidepressant effect of the fermented product on a forced swimming rat model. Process Biochem. 2013, 48, 559–568. [Google Scholar] [CrossRef]
- He, Y.; Ouyang, J.; Hu, Z.; Yang, J.; Chu, Y.; Huang, S.; Yang, Y.; Liu, C. Intervention mechanism of repeated oral GABA administration on anxiety-like behaviors induced by emotional stress in rats. Psychiatry Res. 2019, 271, 649–657. [Google Scholar] [CrossRef]
- Boonstra, E.; de Kleijn, R.; Colzato, L.S.; Alkemade, A.; Forstmann, B.U.; Nieuwenhuis, S. Neurotransmitters as food supplements: The effects of GABA on brain and behavior. Front. Psychol. 2015, 6, 1520. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Gong, T.; Xiang, Y.; Mikkelsen, M.; Wang, G.; Edden, R.A. Single-dose L-dopa increases upper brainstem GABA in Parkinson’s disease: A preliminary study. J. Neurol. Sci. 2021, 422, 117309. [Google Scholar] [CrossRef]
- Hinton, T.; Jelinek, H.F.; Viengkhou, V.; Johnston, G.A.; Matthews, S. Effect of GABA-fortified oolong tea on reducing stress in a university student cohort. Front. Nutr. 2019, 6, 27. [Google Scholar] [CrossRef]
- Mandal, P.K.; Kansara, K.; Dabas, A. The GABA–working memory relationship in Alzheimer’s disease. J. Alzheimer’s Dis. Rep. 2017, 1, 43–45. [Google Scholar] [CrossRef] [Green Version]
- Whissell, P.D.; Bang, J.Y.; Khan, I.; Xie, Y.-F.; Parfitt, G.M.; Grenon, M.; Plummer, N.W.; Jensen, P.; Bonin, R.P.; Kim, J.C. Selective activation of cholecystokinin-expressing GABA (CCK-GABA) neurons enhances memory and cognition. Eneuro 2019, 6. [Google Scholar] [CrossRef]
- Nakamura, H.; Takishima, T.; Kometani, T.; Yokogoshi, H. Psychological stress-reducing effect of chocolate enriched with γ-aminobutyric acid (GABA) in humans: Assessment of stress using heart rate variability and salivary chromogranin A. Int. J. Food Sci. Nutr. 2009, 60, 106–113. [Google Scholar] [CrossRef]
- Xie, Z.; Xia, S.; Le, G.-W. Gamma-aminobutyric acid improves oxidative stress and function of the thyroid in high-fat diet fed mice. J. Funct. Foods 2014, 8, 76–86. [Google Scholar] [CrossRef]
- Varghese, T.; Rejish Kumar, V.; Anand, G.; Dasgupta, S.; Pal, A.K. Dietary GABA enhances hypoxia tolerance of a bottom-dwelling carp, Cirrhinus mrigala by modulating HIF-1α, thyroid hormones and metabolic responses. Fish Physiol. Biochem. 2020, 46, 199–212. [Google Scholar] [CrossRef]
- Hwang, I.; Jo, K.; Shin, K.C.; Kim, J.I.; Ji, Y.; Park, Y.J.; Park, J.; Jeon, Y.G.; Ka, S.; Suk, S. GABA-stimulated adipose-derived stem cells suppress subcutaneous adipose inflammation in obesity. Proc. Natl. Acad. Sci. USA 2019, 116, 11936–11945. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-Y.; Lee, G.-H.; Hoang, T.-H.; Kim, Y.-M.; Jang, G.-H.; Seok, C.-H.; Gwak, Y.-G.-S.; Lim, J.; Kim, J.; Chae, H.-J. GABA and Fermented Curcuma longa L. Extract Enriched with GABA Ameliorate Obesity through Nox4-IRE1α Sulfonation-RIDD-SIRT1 Decay Axis in High-Fat Diet-Induced Obese Mice. Nutrients 2022, 14, 1680. [Google Scholar] [CrossRef]
- Sato, K.; Komaru, T.; Arima, T.; Jardson, C.; Yanaka, N.; Kumrungsee, T. Dietary GABA and its combination with vigabatrin mimic calorie restriction and induce antiobesity-like effects in lean mice. J. Funct. Foods 2021, 78, 104367. [Google Scholar] [CrossRef]
- Oketch-Rabah, H.A.; Madden, E.F.; Roe, A.L.; Betz, J.M. United States Pharmacopeia (USP) safety review of gamma-aminobutyric acid (GABA). Nutrients 2021, 13, 2742. [Google Scholar] [CrossRef] [PubMed]
- Tanamool, V.; Hongsachart, P.; Soemphol, W. Screening and characterisation of gamma-aminobutyric acid (GABA) producing lactic acid bacteria isolated from Thai fermented fish (Plaa-som) in Nong Khai and its application in Thai fermented vegetables (Som-pak). Food Sci. Technol. 2019, 40, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Barla, F.; Koyanagi, T.; Tokuda, N.; Matsui, H.; Katayama, T.; Kumagai, H.; Michihata, T.; Sasaki, T.; Tsuji, A.; Enomoto, T. The γ-aminobutyric acid-producing ability under low pH conditions of lactic acid bacteria isolated from traditional fermented foods of Ishikawa Prefecture, Japan, with a strong ability to produce ACE-inhibitory peptides. Biotechnol. Rep. 2016, 10, 105–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, S.C.; Domingos-Lopes, M.F.; Stanton, C.; Ross, R.P.; Silva, C.C. Production of-aminobutyric acid (GABA) by Lactobacillus otakiensis and other Lactobacillus sp. isolated from traditional Pico cheese. Int. J. Dairy Technol. 2018, 71, 1012–1017. [Google Scholar] [CrossRef]
- Li, H.; Cao, Y. Lactic acid bacterial cell factories for gamma-aminobutyric acid. Amino Acids 2010, 39, 1107–1116. [Google Scholar] [CrossRef]
- Dhakal, R.; Bajpai, V.K.; Baek, K.-H. Production of GABA (γ-aminobutyric acid) by microorganisms: A review. Braz. J. Microbiol. 2012, 43, 1230–1241. [Google Scholar] [CrossRef] [Green Version]
- Cunha, D.S.; Coelho, M.C.; Ribeiro, S.C.; Silva, C.C. Application of Enterococcus malodoratus SJC25 for the Manufacture of Whey-Based Beverage Naturally Enriched with GABA. Foods 2022, 11, 447. [Google Scholar] [CrossRef]
- Angelin, J.; Kavitha, M. Exopolysaccharides from probiotic bacteria and their health potential. Int. J. Biol. Macromol. 2020, 162, 853–865. [Google Scholar] [CrossRef]
- Saadat, Y.R.; Khosroushahi, A.Y.; Gargari, B.P. A comprehensive review of anticancer, immunomodulatory and health beneficial effects of the lactic acid bacteria exopolysaccharides. Carbohydr. Polym. 2019, 217, 79–89. [Google Scholar] [CrossRef]
- Ruas-Madiedo, P.; de los Reyes-Gavilan, C.G. Invited review: Methods for the screening, isolation, and characterization of exopolysaccharides produced by lactic acid bacteria. J. Dairy Sci. 2005, 88, 843–856. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.A.; Lopes Neto, J.H.P.; Cardarelli, H.R. Exopolysaccharides produced by Lactobacillus plantarum: Technological properties, biological activity, and potential application in the food industry. Ann. Microbiol. 2019, 69, 321–328. [Google Scholar] [CrossRef]
- Torino, M.I.; Font de Valdez, G.; Mozzi, F. Biopolymers from lactic acid bacteria. Novel applications in foods and beverages. Front. Microbiol. 2015, 6, 834. [Google Scholar] [CrossRef] [Green Version]
- Paulo, E.M.; Vasconcelos, M.P.; Oliveira, I.S.; Affe, H.M.d.J.; Nascimento, R.; Melo, I.S.d.; Roque, M.R.d.A.; Assis, S.A.d. An alternative method for screening lactic acid bacteria for the production of exopolysaccharides with rapid confirmation. Food Sci. Technol. 2012, 32, 710–714. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Yang, Z.; Jing, X.; Yu, P.; Zhang, Y.; Yi, H.; Zhang, L. Improvement of the texture of yogurt by use of exopolysaccharide producing lactic acid bacteria. BioMed Res. Int. 2016, 2016, 7945675. [Google Scholar] [CrossRef]
- Patten, D.; Laws, A.P. Lactobacillus-produced exopolysaccharides and their potential health benefits: A review. Benef. Microbes 2015, 6, 457–471. [Google Scholar] [CrossRef]
- Korcz, E.; Kerényi, Z.; Varga, L. Dietary fibers, prebiotics, and exopolysaccharides produced by lactic acid bacteria: Potential health benefits with special regard to cholesterol-lowering effects. Food Funct. 2018, 9, 3057–3068. [Google Scholar] [CrossRef]
- Caggianiello, G.; Kleerebezem, M.; Spano, G. Exopolysaccharides produced by lactic acid bacteria: From health-promoting benefits to stress tolerance mechanisms. Appl. Microbiol. Biotechnol. 2016, 100, 3877–3886. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coelho, M.C.; Malcata, F.X.; Silva, C.C.G. Lactic Acid Bacteria in Raw-Milk Cheeses: From Starter Cultures to Probiotic Functions. Foods 2022, 11, 2276. https://doi.org/10.3390/foods11152276
Coelho MC, Malcata FX, Silva CCG. Lactic Acid Bacteria in Raw-Milk Cheeses: From Starter Cultures to Probiotic Functions. Foods. 2022; 11(15):2276. https://doi.org/10.3390/foods11152276
Chicago/Turabian StyleCoelho, Márcia C., Francisco Xavier Malcata, and Célia C. G. Silva. 2022. "Lactic Acid Bacteria in Raw-Milk Cheeses: From Starter Cultures to Probiotic Functions" Foods 11, no. 15: 2276. https://doi.org/10.3390/foods11152276