
Microalgae: Bioactive Composition, Health Benefits, Safety and Prospects as Potential High-Value Ingredients for the Functional Food Industry


1
Department of Food Science and Technology, University of California, Davis, CA 95616, USA
2
Department of Plant, Food and Environmental Sciences, Dalhousie University, 50 Pictou Road, Truro, NS B2N 5E3, Canada
*
Author to whom correspondence should be addressed.
Foods 2022, 11(12), 1744; https://doi.org/10.3390/foods11121744
Submission received: 28 April 2022
/
Revised: 7 June 2022
/
Accepted: 9 June 2022
/
Published: 14 June 2022
(This article belongs to the Special Issue Metabolomics Analysis for Food Authenticity and Safety)
Abstract
:Global population is estimated to reach about 9.22 billion by 2075. The increasing knowledge on the relationship between food biochemistry and positive health gives an indication of the urgency to exploit food resources that are not only sustainable but also impact human health beyond basic nutrition. A typical example of such novel food is microalgae, an aquatic microorganism with a plethora of diverse bioactive compounds including phenolics, carotenoids, vitamin B12 and peptides. Microalgal bioactive compounds have been shown to possess positive health effects such as antihypertensive, anti-obesity, antioxidative, anticancer and cardiovascular protection. Although, the utilization of microalgal biomass by the functional food industry has faced lots of challenges because of species diversity and variations in biomass and cultivation factors. Other documented challenges were ascribed to changes in functional structures during extraction and purification due to inefficient bio-processing techniques, inconclusive literature information on the bioavailability and safety of the microalgal bioactive compounds and the fishy odor and taste when applied in food formulations. In spite of these challenges, great opportunities exist to exploit their utilization for the development of functional foods. Microalgae are a renewable resource and have fast growth rate. Therefore, detailed research is needed to bridge these challenges to pave way for large-scale commercialization of microalgal-based healthy foods. The focus of this review is to discuss the potential of microalgae as natural ingredients for functional food development, factors limiting their acceptance and utilization in the food industry as well as their safety concerns with respect to human consumption.
1. Introduction
Functional role of foods has shifted from only the provision of energy and basic nutrients to include the supply of non-nutritive bioactive compounds capable of offering protection against the development of chronic diseases. Plant food groups (e.g., grains, legumes, nuts, fruits and vegetables, etc.) with these duo characteristics are defined as functional foods, which can be consumed as fresh foods, minimally processed or as ingredients in formulated foods [1]. However, with the increasing world population and consumer knowledge of the relationship between food intake and development of diseases, there is the need to exploit other food sources that can (i) sustain the functional food industry and (ii) meet the demanding health needs of the growing populace. In pursuit of this, an underexploited aquatic microorganism with potential functional food supply is microalgae.
Compared to terrestrial plants, microalgae are sustainable due to their rapid growth, ease of cultivation and non-competition for arable land. Microalgae are unicellular aquatic microorganisms with over 50,000 classified species. Some notable examples include Nostoc commune, Arthrospira platensis, Aphanizomenon flosaquae, Chlorella vulgaris and Chlorella pyrenoidosa [2,3]. Besides their appreciable levels of primary metabolites (e.g., protein, carbohydrates, polyunsaturated fatty acids and vitamins), the health benefits of microalgae are mainly correlated to the presence of high value secondary metabolites. Secondary metabolites are non-nutritive compounds produced in plants as defense agents against environmental stress [4]. Microalgae research has shown substantial concentrations of diverse secondary metabolites such as pigments (e.g., phenolics, carotenoids, etc.), phytosterols and mycosporine-like amino acids [5] (Figure 1).
Boussiba et al. [6] reported the total biomass of Haematococcus pluvialis to consist of 4% astaxanthin. In another study by Hifney et al. [7], phycobilin content of Arthrospira platensis was reported as 25% of its total dry biomass. According to literature, microalgal metabolites are of health value due to their capacity as antioxidant, antimicrobial, antitumor, anti-inflammatory and anticancer capacities [8,9]. Therefore, considering their renewability and plethora of high-value bioactive compounds, microalgae can be postulated as sustainable healthy food systems, which may be a reason for their increasing scientific literature over time (Figure 2).
The current review aim is to provide a comprehensive discussion on the microalgal composition of bioactive compounds, their biological activities, food applications, safety concerns and factors limiting their utilization in the functional food industry. This review further proposed innovative approaches to enhance the acceptance and utilization of microalgae as alternative plant-based novel food ingredients.
2. High-Value Bioactive Primary Metabolites
2.1. Polyunsaturated Fatty Acids (PUFAs)
Polyunsaturated fatty acids (PUFAs) cannot be produced by the human body, hence, the need to obtain them through food consumption. They are divided into two groups namely omega-3 fatty acids (including α-linolenic, ALA; eicosapentaenoic acid, EPA; and docosahexaenoic acid, DHA) and omega-6 fatty acids (including arachidonic acid ARA; linoleic acid, LA; γ-linoleic acid, GLA; and conjugated linoleic acid, CLA) [10]. The health value of microalgae can partly be directed to its composition of PUFAs, which have been shown to promote brain and eye health, as well as protect against cardiovascular diseases, obesity, diabetes and arthritis [11]. Well-known PUFA microalgal producers include Crypthecodinium, Schizochytrium and Ulkenia sp. although other genera such as Phaeodactylum, Monodus, Nannochloropsis and Porphyridium have also shown considerable levels of DHA and EPA [12]. However, it is important to highlight that a majority of research data on microalgal PUFAs are reported with reference to their biofuel applications, with very limited reports on food, health and pharmaceutical applications. Considering the health benefits of PUFAs, coupled with the low consumer acceptance of fish oil PUFAs (i.e., low oxidative stability and high off-flavors), it can be postulated that there is the demand for alternative PUFAs. Hence, scientific data on microalgal PUFA profile, their bioactive properties and stabilities under different processing conditions will be very crucial in helping promote their applications in the functional food industry. In the study of Aussant et al. [13], eight species of microalgae were cultivated under different conditions (temperature—8, 14, 20 and 26 °C; time—5, 10 and 14 days). Of the eight investigated species, Nannochloropsis oculata and Isochrysis galbana reported the highest concentration of EPA (2.52 mg/L) and DHA (1.08 mg/L) at 20 °C/day 5 and 14 °C/day 5, respectively, with their investigated in vitro nutritional indices (i.e., hypocholesterolemic, atherogenic and thrombogenic indices) falling within accepted health ranges. Similarly, an increase in bicarbonate concentration from 2 to 8 mM increased the total PUFA content by 5.6% in Pavlova lutheri [14]. However, increasing light intensity from 37.7 to 100.0 µmol/m2/s in Chlorella vulgaris reduced DHA and EPA levels by 50 and 70%, respectively [15].
2.2. Polysaccharides
From the review of Mourelle et al. [16], microalgal polysaccharides are largely exploited from the genera Porphyridium, Phaeodactylum, Chlorella, Tetraselmis, Isochrysis and Rhodella. In microalgae, polysaccharides function as protection agents, energy reservoirs and structural molecules, and are divided into pectins, glycol-protein, sulfated polysaccharides (SPS) and homo- and hetero-polysaccharides [17,18]. Among these polysaccharide groups, the most widely reported is the sulfated group with findings mainly reported on their anti-inflammatory benefits. In the study of Matsui et al. [19], extracts of sulfated polysaccharides from Porphyridium showed in vitro migratory inhibition of leukocytes to inflammation sites. These authors also observed in vivo microalgal inhibition against erythema development. Few antioxidant studies have also been reported with microalgae polysaccharides, as further discussed in Section 4 of the text. It is also important to highlight that microalgal polysaccharides are often exploited for their techno-functional applications, compared to their health benefits, thus, making it difficult to correlate their structure with health effects. Therefore, future studies focusing on the structure–activity relationship between microalgal polysaccharides and potential bioactive effects are highly recommended.
2.3. Vitamins
Although vitamins are essential elements required for proper human development, they can only be obtained through diets or supplements. Microalgae are excellent potential source of vitamins, compared to some well-known sources such as orange, carrot and soy flour [20]. Although microalgae are not natural producers of vitamin A, it is interesting to note that microalgae can accumulate vitamin A precursors such as carotenes (i.e., α- and β-carotenes) and retinol, which have been demonstrated to protect against the development of some cancer types [21]. The recent study of Ljubic et al. [22] investigated the accumulation of vitamin D3 (cholecalciferol) in Nannochloropsis oceanica, Arthrospira maxima, Rhodomonas salina and Chlorella minutissima upon exposure to different doses of ultraviolet B (0, 15, 22 and 36 kJ/m2/day) for 7 days. The authors observed the highest level of vitamin D3 (1 µ/g dry weight) with Nannochloropsis oceanica at UV-B dose of 36 kJ/m2/day, compared to the control (< 0.004 µ/g DW). Edelmann et al. [23] reported vitamin B9 contents in formulated powders of Chlorella sp. and Nannochloropsis sp. to be 25.9 and 20.8 µg/g, respectively. It can therefore be postulated that the consumption of about 5 g of Chlorella and Nannochloropsis microalgae powder can provide a quarter of the recommended daily intake (i.e., 400 µg/d) of vitamin B9. According to Tarento et al. [24], cyanobacteria has about 200 µg/g of vitamin K1, being about six times higher than levels reported for parsley (i.e., 37 µg/g), a well-known vitamin K1 food source. Hence, adult daily consumption of 1 g cyanobacteria will provide three times their daily needs for vitamin K1. Another crucial vitamin imperative for good health, especially among the aged is vitamin B12, although it is limited in plant foods [25]. Nevertheless, Edelmann et al. [23] observed Chlorella sp. to contain 2.4 µg/g of vitamin B12, concluding that 5 g Chlorella powder will provide five times the daily requirement of vitamin B12. A further important observation is that literature on bioaccessibility and bioavailability of microalgal vitamins is very limited. Thus, the need for deeper studies to enable health-regulating agencies and food industries to approve and include microalgae in the formulation of functional foods.
2.4. Peptides
Peptides are short chain amino acids (i.e., 20–50 units) linked together by peptide bonds [26]. According to Khanra et al. [27], 50% of the global protein and peptide market is currently sourced from terrestrial plants and may be replaced by proteins from microalgae and insects by 2054. Considering this trend, microalgal peptides has been exploited from Chlorella, Navicula, Tetraselmis and Nitzschia [2]. Ko et al. [26] isolated a pentapeptide with the amino acid sequence Leu-Asn-Gly-Asp-Val-Trp from Chlorella ellipsiodea, and reported appreciable peroxyl radical, 1,1-Diphenyl-2-picrylhydrazyl (DPPH) and hydroxyl radical scavenging capacities with half maximal inhibitory concentration values (IC50) of 0.02, 0.92 and 1.42 mM, respectively. Two isolated peptides from Nannochloropsis oculata with an amino acid sequence of Gly-Met-Asn-Asn-Leu-Thr-Pro and Leu-Glu-Gln were found to possess anti-hypertensive properties by inhibiting the activity of angiotensin-converting enzyme (ACE) at IC50 values of 123 and 173, respectively [28]. According to these authors, microalgal peptides can exhibit antihypertensive properties through the (i) inhibition of ACE, the main enzyme responsible for vasoconstriction of veins and arteries (ii) triggering of vasodilation effect, i.e., capacity to increase nitric oxide levels through the stimulation of the endothelial nitric oxide synthase pathway.
3. High-Value Microalgae Secondary Metabolites
3.1. Pigments
Pigments are secondary metabolites responsible for the coloring of living organisms, harvesting of light energy for photosynthesis and protection against oxidative stress. Up until now, the main groups of microalgal pigments include carotenoids, phenolics, chlorophyll and phycobiliproteins [2]. Details of the subgroups within these pigment groups are discussed further in the text, focusing mainly on their applications in functional food development.
3.1.1. Phenolic Compounds
Basically, phenolics are benzene ringed compounds, with hydroxyl groups present in their chemical structure. Depending on their carbon skeleton, phenolic compounds are divided into phenolic acids (e.g., hydroxybenzoic acids such as gallic and vanillic acids and hydroxycinnamic acids such as caffeic and coumaric acids), flavonoids (e.g., anthocyanin, quercetin and catechin), tannins (e.g., phlorotannins) and stilbenes [4]. Although microalgae have appreciable levels of phenolic compounds, majority of literature studies on aquatic phenolics are reported with macroalgae (i.e., seaweed). Nevertheless, the few literatures available on microalgae show significant variations due to specie type, cultivation conditions and techniques used for extraction, identification and quantification. Chlorella pyrenoidosa and Arthrospira platensis showed total phenolic levels of 25.8 and 43.2 mg/g, respectively [29]. In another study by Goiris et al. [30], phenolic composition of 32 microalgae species ranged between 54 and 375 mg/g. Additionally, Widowati et al. [31] observed total phenolic levels of 16.87 and 17.80 mg/g with Tetraselmis chuii and Isochrysis galbana, respectively. Interestingly, although literature has reported red grapes (Vitis vinifera) as the leading fruit producer of phenolic compounds, a study by Manach et al. [32] showed phenolic levels of microalgae to be comparable. However, to encourage their applications in the food industry, in-depth studies on the characterization of microalgal phenolic compounds are recommended to scientifically correlate their potential health benefits with specific functional foods.
3.1.2. Carotenoids
Carotenoids are lipid-soluble pigments mainly responsible for the purple, red, orange and yellow colors of living organisms. According to Rammuni et al. [33] about 600 carotenoid structures have been identified. Structurally, carotenoids are tetraterpenes consisting of eight isoprene units with a 40-carbon skeleton, divided into two groups namely carotenes and xanthophylls [34]. The carotene groups of lycopene and β-carotene consist solely of hydrocarbons (C40H56), whereas the xanthophylls (e.g., lutein, zeaxanthin and astaxanthin) are made up of carbon, hydrogen and oxygen (C40H56O2) [35]. Among microalgal species, the dominant carotenoid producing group is Chlorophyceae, which can yield about 90% of xanthophylls and carotenes with limited levels of fucoxanthin, diatoxanthin and diadinoxanthin [36]. Previous literature such as the study of Inbaraj et al. [37] investigated the presence of different carotenoids in Chlorella pyrenoidosa. Their study observed the presence of cis lutein isomers (27975.3 µg/g), trans-α-carotene (2465.8 µg/g), zeaxanthin (2170.3 µg/g), β-carotene cis isomers (2159.3 µg/g), trans-β-carotene (2155.0 µg/g), α-carotene cis isomers (1766.7 µg/g), β-cryptoxanthin (334.9 µg/g), neoxanthin and its cis isomers (199.7 µg/g), neochrome (65.2 µg/g), auroxanthin (38.5 µg/g) and violaxanthin and its cis isomers (38.1 µg/g).
β-Carotene
From the work of Khanra et al. [27], β-carotene is prevalently produced by Scenedesmus almeriensis, Dunaliella bardawil, Dunaliella tertiolecta and Dunaliella salina. Consumption of microalgae can help meet the physiological need of vitamin A, since research has reported the body’s capacity of converting β-carotene to vitamin A. Among microalgae species, Dunaliella salina has been reported as the highest source of β-carotene, exhibiting about 98.5% β-carotene in relation to its total carotenoid content (13%, dry biomass) [33]. Compared to other sources, microalgal β-carotene has been shown to be a rich source of 9-cis β-carotene (i.e., a cyclic carotene with the structure of β-carotene, but with a cis double bond attached to positions 9 and 10) [36]. In fact, this structure is a major contributor of its free radical scavenging capacities. Secondary metabolites of some microalgae are presented in Table 1.
Lutein
Lutein has gained much research interest in recent years due to its food coloring property and nutraceutical value, especially as antioxidant and anticancer agents, with intake up to 60 mg/day safe for adults of 60 kg body weight [53]. Previous research has correlated lutein with antioxidative, antiaging, anti-cardiovascular and anticancer benefits [9]. Among microalgae, the Chlorella genus is reported as the best source of lutein, with Chlorella protothecoides being dominant (about 4.6% on dry weight basis) [54]. Other microalgal species such as Muriellopsis sp. D. salina, Scenedesmus almeriensis and Galdieria sulphuraria are also exploited for lutein [55]. Although industrial exploitation of natural lutein is sourced from marigold petals (i.e., 0.03% on dry weight basis), lutein concentrations in Chlorella protothecoides and Muriellopsis sp. are 150 and 18 times higher compared to marigold [56].
Fucoxanthin
This compound accounts for about 10% of total carotenoids found in nature. It is abundant in microalgae when compared with other aquatic plants. Fucoxanthin is a lipophilic compound characterized by a carotene structure and an oxygenated backbone, with the presence of functional groups such as hydroxyl, epoxy, carboxyl and carbonyl moieties [57]. It absorbs light within ranges of 450–540 nm, thus translating light in the blue-green to yellow-green sections of the visible spectrum. Previous studies have reported positive bioactive benefits of fucoxanthin-rich foods such as antioxidant, antimicrobial, anticancer, antihypertensive and many others [58]. These bioactive benefits have been attributed to different mechanisms of action such as induction of apoptosis, cell cycle arrest, scavenging of intracellular reactive oxygen species and inhibition of cell proliferation [59].
Astaxanthin
Among the aquatic food chain, microalgae represent the major source of astaxanthin [60]. Astaxanthin, a red colored xanthophyll, is the second most exploited microalgae carotenoid after β-carotene [21]. Among microalgae species, the main producer of astaxanthin is Haematococcus pluvialis, with about 81% in relation to its total carotenoid yield of 7% on dry weight basis [2]. Besides Haematococcus pluvialis, species such as Chlorella zofingiensis, Chlorococcum sp. and Scenedesmus sp. can also accrue astaxanthin but at limited levels. Antioxidant capacity of astaxanthin is 10 times higher compared to lutein, β-carotene, lycopene and zeaxanthin and even 100 times higher than vitamins C and E [16,41], with this effectiveness attributed to its structural composition of thirteen conjugated double bonds. Considering their high antioxidant capacity, astaxanthin-rich microalgae can be exploited as novel ingredients for functional food development. Currently, this is being applied by the aquaculture industry, where astaxanthin rich microalgae are used as feed for fish and crustaceans [61]. For instance, Kim et al. [62] observed that the consumption of astaxanthin from Haematococcus sp. decreased lipid peroxidation and oxidative damage among heavy smokers. In another study, Yoshida et al. [63] fed 61 non-obese participants with an astaxanthin extract (i.e., AstaReal®) from Haematococcus pluvalis for a period of two weeks. After the study period, participants fed with astaxanthin formulated diets (i.e., 12–18 mg/day) showed increased serum HDL and adiponectin levels compared to those that were fed on placebo.
Zeaxanthin
Zeaxanthin is predominantly produced by microalgal strains of Scenedesmus almeriensis and Nannochloropsis oculata. Research has proved a simultaneous relationship between zeaxanthin intake and health benefits such as protection against macular degeneration, cataracts, cancer and tumors [57]. Koyande et al. [21] associated visual health effects of zeaxanthin with the presence of nine conjugated double bonds and hydroxyl group in its chemical structure. This functional structure demonstrates its strong ability to absorb blue light and protect against retina damage. According to the findings of Ravi et al. [64], the daily consumption of zeaxanthin (i.e., 400 mg/kg body weight) showed no acute, sub-chronic toxicity or mutagenic effects. This is an attestation of the safety of zeaxanthin when consumed at recommended levels. It should also be added that zeaxanthin is not only beneficial for eye health but also protective against the development of certain cancer types (e.g., pancreatic and breast cancers) and type 2 diabetes [35].
3.1.3. Chlorophyll
Depending on the microalgal strain and growth conditions, the chlorophyll content of a cell ranges from 0.5 to 4% of the total dry mass [21]. Chlorophyll is a natural green pigment crucial in photosynthetic organisms for harvesting of energy from sunlight. They exist as either chlorophyll a, b or c. However, chlorophyll c is only present in brown algae and not in green algae [21]. Species of Chlorella are the main producers of chlorophyll, with other species such as Spirulina and Arthrospira producing limited concentrations [27]. Besides their photosynthetic role, chlorophyll has been shown to exhibit strong antioxidant capacity. In the recent study of Suparmi et al. [65], sodium-nitrate-treated Wistar rats were fed chlorophyll extracts (8 and 16 µg/mL) from Sanropus androgynus for 14 days, after which the authors observed strong antioxidant protection in treated Wistar rats in in vivo studies.
3.2. Phycobiliproteins
Phycobiliproteins are colored hydrophilic proteins capable of absorbing light energy between wavelength ranges of 470–660 nm [45]. According to their level of light absorption spectra, phycobiliproteins are divided into four main groups including phycoerythrins (540–570 nm), phycocyanins (610–620 nm), allophycocyanins (650–6500 nm) and phycoerythrocyanins (560–600 nm) [44]. Phycobiliproteins are predominantly found among cyanobacteria (e.g., Spirulina), red algae (e.g., Porphyridium and Galdieria), cryptophyte and glaucophyte [66]. Structurally, they are composed of two dissimilar peptide chains covalently attached to chromophores. In fact, observations by previous researchers led to the conclusion that the amino acid composition of phycobiliproteins in combination with the presence of double bonds in their chromophores are primarily responsible for their wide range of antioxidative capacities [43]. Phycocyanin is a natural, blue-colored pigment applied in formulations of confectionary, dairy, snacks and beverages. They offer health benefits such as anti-inflammatory, antiviral, antioxidative, hepatoprotective and neuroprotective effects [43,67]. Indeed, the much-advocated health benefits from spirulina consumption have been widely attributed to its appreciable concentration of phycocyanin.
3.3. Mycosporine-like Amino Acids (MAAs)
Mycosporine-like amino acids (MAAs) exhibit UV absorbance capacities between 310 and 365 nm. Structurally, MAAs are low molecular weight (<400 Da) molecules composed of either cyclohexenone or cyclohexenimine chromophores conjugated with one or two amino acids [68]. Notable examples of MAAs include shinorine, palythenic acid, mycosporine-glycine, mycosporine-taurine, palythine and porphyra-334 [69]. Majority of MAAs data on health benefits are based on their UV-absorbing capacities, with very limited literature on food applications. However, taking advantage of their reported free radical scavenging capacities, the application of MAAs as natural food preservatives can be postulated. There is the need to investigate other bioactive effects of MAAs besides their UV-absorbing and free radical scavenging capacities in-order to increase their utilization in the food industry.
3.4. Phytosterols
Phytosterols are plant-based steroids such as animal cholesterol but vary only in their carbon side chains and/or the presence of a double bond compared to animal cholesterol [70]. Phytosterols are made up of two different groups, namely sterols and stanols, with sterols possessing a double bond in its structure (i.e., making it unsaturated) and vice versa for the stanols (i.e., making it saturated) [70]. Recently, microalgal phytosterols are receiving attention from researchers due to their positive health impacts such as anticancer, antioxidative, anti-inflammatory, anti-atherogenicity, cardiovascular protection and reduction in LDL cholesterol [71].
Microalgae genera such as Dinoflagellates are key producers of 4α-methyl sterols, Pelagophyceae sp. are dense in 24-propylidenecholesterol, whereas Glaucocystophyte sp. are not only rich in sitosterols but also campesterol and stigmasterol [72]. Furthermore, Ahmed et al. [73] reported phytosterol production capacity in 10 microalgae species, with their work reporting Pavlova lutheri as the best producer of phytosterols (5186 mg/100 g) among other investigated species such as Tetraselmis sp. and Nannochloropsis sp. Thus, depicting the sustainable use of microalgae as a phytosterol resource compared to current options such as canola (Brassica napus L.) and corn (Zea mays) which produce about 820 and 850 mg phytosterol/100 g, respectively.
4. Bioavailability and Health Impacts of Microalgae as Food Ingredients
4.1. Antioxidative Properties
Because the human body is an active respiratory system, there is always the release of free radicals which can induce oxidative stress [21]. Therefore, to keep free radicals under control, the body protects itself through the actions of antioxidant enzymes (e.g., catalase and superoxide dismutase) or dietary antioxidants (e.g., pigments and peptides). Microalgae pigments such as phenolic compounds can exhibit antioxidant activities through their donation of hydrogen atoms to unstable free radicals [2]. Besides microalgae concentration of phenolic compounds, one important factor that contributes to their antioxidant capacity is the types of phenolic compounds present in its biomass.
De Jesus Raposo et al. [74] correlated antioxidant capacity of carotenoids with the presence of conjugated double bonds and specific functional groups such as epoxy, acetyl, allene and acetylene. Kent et al. [75] observed that peptides from Alexandrium minutum activated targeted cell death in human lung cancer cell lines (Wi38) at an IC50 value of 0.7 µg/mL with no detrimental effect on normal cells. Ko et al. [26] isolated and cultured a pentapeptide (amino acid sequence of Leu-Asn-Gly-Asp-Val-Trp) from Chlorella ellipsiodea with monkey kidney cells exposed to hydrochloride cytotoxicity. From their results, the pentapeptide at concentrations of 25, 50 and 100 µM enhanced cellular viability against hydrochloride-induced cytotoxicity by 56.3, 72.3 and 79.4%, respectively.
4.2. Hypertension
Currently, the drug lisinopril is applied for treatment of hypertension but has been linked with side effects such as dizziness, headache, vomiting and diarrhea [76]. Therefore, there is a need to explore natural alternatives. Ko et al. [26] observed a positive correlation between peptides from Chlorella ellipsiodea and the inhibition of the angiotensin-converting enzyme (ACE), the enzyme responsible for vasoconstriction of blood vessels. The investigated peptide (i.e., Val-Glu-Gly-Tyr amino acid sequence) showed ACE inhibition activity (IC50) of 128.4 µM. From this same study, the authors further observe that Md reduced systolic blood pressure effects when the investigated peptide fractions were fed to rats via in vivo studies.
4.3. Antimicrobial
A disc diffusion assay by Desbois et al. [77] showed that PUFAs (i.e., EPA, hexadecatrienoic and palmitoleic acids) from Phaeodactylum tricornutum exhibited high antibacterial activity against Staphylococcus aureus. Scaglioni et al. [78] also investigated antifungal and anti-mycotoxigenic potential of phenolic extracts from Nannochloropsis sp. and Spirulina sp. against cultures of Fusarium graminearum in trichothecenes mycotoxins. The authors observed that, 40 µg/mL of Nannochloropsis sp. phenolic extracts showed 98 and 100% inhibition of acetylates and nivalenol and deoxynivalenol, respectively. Additionally, phenolic extracts obtained from Spirulina sp. reduced the production of deoxynivalenol, acetylates and nivalenol by 62, 78 and 100%, respectively.
4.4. Anti-Inflammation
A 100 µg/mL carotenoid extract comprising lutein, violaxanthin, antheraxanthin, neoxanthin and carotenes from Tetraselmis suecica showed anti-inflammatory effects in human cells exposed to 30 mM H2O2 oxidative injury [79]. Saini and Keum [80] also reviewed isolated polysaccharides from Chlorella stigmatophora and Phaeodactylum tricornutum and reported higher anti-inflammatory effects (i.e., IC50 2.25 and 2.92 mg/kg, respectively) in rats exposed to carrageenan-induced paw oedema compared to the synthetic drug indomethacin (IC50 8.58 mg/kg). Chrysolaminarin, a water soluble β-1,3-D-glucan isolated from the diatom Synedra acus exhibited anti-proliferative effects against human colon cancer cell lines DLD-1 and HCT-116, at IC50 values of 47.7 and 54.5 µg/mL, respectively [81]. Additionally, due to the vigorous antioxidant and microbiota regulation capacities of microalgal fucoxanthin, it is currently being applied for the treatment of various inflammatory-related diseases [82].
4.5. Anti-Proliferative and Cardiovascular Effects
A study by Murai et al. [83] which involved 86,113 participants (i.e., 40,707 men and 45,406 women) revealed an inverse relationship between seaweed/microalgae consumption and risk of ischemic heart disease. In another study, animal diets formulated with the polysaccharide-rich red algae, Rhodella reticulata, resulted in reduced serum cholesterol levels [84]. From this same study, animals fed with polysaccharide-rich red algae showed reduced levels of glucose and insulin. From the literature, it is notable that few studies have investigated the health benefits of microalgal-based bioactive compounds either in vivo or in vitro. Indeed, if researchers believe that the world population is increasing tremendously and requires sustainable healthy food systems, then this research gap with microalgal studies should be addressed with all seriousness. Almeida et al. [85] investigated the effect of fucoxanthin on cancerous cell lines such as the human bronchopulmonary carcinoma cell line NSCLC-N6, erythromyeloblastoid leukemia cell line K562 and human lymphoblastoid cell line TK6. According to their study, all investigated cell lines showed positive effects upon treatment with fucoxanthin. Similarly, in vivo studies with mice showed reduced tumor growths of primary effusion lymphoma, sarcomas and osteosarcoma when fed with microalgal-based fucoxanthin extracts [86].
5. Microalgae Utilization for Food Formulations
Formulation of functional foods from microalgal biomass is expected to increase in the near future as the global demand for healthy foods increases. As discussed by Nova et al. [3], the application of microalgae for food formulations has opened a new phase for the functional food industry, since a continuous search for innovative raw ingredients targeted for novel food developments exists. However, as discussed in the review of Nethravathy et al. [87], essential questions to be addressed towards the efficient formulation of microalgal-based food products include:
- Interaction of microalgal biomass with other food matrix components;
- Impact of microalgal addition on food sensory attributes, especially color and taste;
- Textural and rheological changes in formulated food products upon microalgal formulation;
- Changes and stability of microalgal compounds upon exposure to processing conditions.
Development of eco-innovative techniques to meet these gaps will help maximize the utilization of microalgal-based ingredients in the current competitive functional food marketspace. Currently, the food industry is applying whole microalgal biomass or their extracted purified compounds as novel ingredients for the formulation of food products such as baked goods, pasta, noodles, plant-based milk, soups and many others, with a reflection of improved techno-functional and nutritional properties as depicted in Figure 3 and Table 2.
5.1. Snacks
Due to the increasing busy lifestyle of modern-day consumers, there is a high demand for snacks that are not only cheap but also convenient and healthy. Lucas and his colleagues [94] formulated snacks with A. platensis (at 2.6%, w/w) and observed enhanced levels of protein, minerals and lipids at 22.6, 46.4 and 28.1%, respectively, compared to the control. The authors also observed a positive coloration of the developed snacks as impacted by A. platensis with no negative effect on its texture, flavor and taste, with an overall sensory acceptability of 82%. Similarly, Bolanho et al. [105] developed cookies from A. platensis at levels of 2 and 5% (w/w). According to their study, the highest formulation level of 5%, reported maximum levels of protein, fiber and ash compared to their 2 and 0% formulated cookie counterparts. However, from their sensory studies, cookies formulated with 5% A. platensis reported the lowest score for consumer acceptability. Thus, this evidence confirms the report of Nethravathy et al. [87] that a major challenge for the microalgae-based food industry is finding the right balance between microalgae formulation level, product health quality and final consumer acceptability. Without the proper answer to these gaps, the microalgae-based food industry will suffer major market drawbacks. In another study, cookies formulated with astaxanthin extract from Haematococcus pluvialis showed improved nutritional, antioxidant and phenolic profiles [106]. An example of a microalgae-formulated snack is displayed in Figure 4.
5.2. Bread
Undoubtably, bread is one of the most widely consumed food products globally [107,108]. Based on the wide consumption and popularity of bread, developing functional bread will be a great vehicle for global food and nutrition security. In an interesting study by Uribe-Wandurraga et al. [108], breadsticks formulated with Chlorella vulgaris and A. platensis reported increased levels of iron and selenium, as well as color and texture stability over a storage period of 15 days, compared to their control forms. In another study, bread developed with 3 g of Chlorella vulgaris per 100 g of wheat flour showed improved viscoelastic and rheological properties, compared to the control and formulations above 3 g [109]. Hafsa et al. [110] formulated bread with A. platensis at 1 and 3% (w/w) substitution levels. From their study, although 3% formulation increased bread concentrations of minerals, protein and fat, this formulation level scored reduced consumer acceptability due to the green color impacted to the bread from microalgae. Thus, this evidence agrees with previous literature that the purchasing power of a food product also depends on its sensory attribute besides nutrition. Additionally, crostini formulated with different levels (0, 2, 6 and 10%, w/w) of A. platensis (Figure 5) showed increased protein, phycocyanin and total phenolic contents with increasing formulation levels, although this trend resulted in reduced in vitro digestibility [106]. Moreover, sensory analysis showed comparable acceptance for 2% microalgae formulated crostini with the control. Therefore, this suggests the strong need for a balance between microalgae food formulation, nutrition and consumer acceptability.
5.3. Others
Different food types formulated with Spirulina sp. are presented in Figure 6.
In one study, Chlorella vulgaris formulated with spreadable processed cheese at levels of 2, 4 and 6% (w/w) showed enhanced magnesium, potassium, selenium, zinc, iron and antioxidant potential, with this trend increasing with high formulation levels [112]. A functional yoghurt formulated with omega-3-rich lipid extract from Pavlova lutheri improved the concentration of n-3 PUFA for enhanced anti-inflammatory effects. However, although the study reported improved health benefits of the microalgae-formulated yoghurt, the authors also observed limited consumer acceptability upon sensory evaluation. In another detailed in vivo study, 1264.69 mg of omega-3-rich algal oil from Schizochytrium sp. was formulated with yoghurt and fed to 11 participants [113]. After 21 days of feeding, participants fed with formulated yoghurt significantly showed higher levels of bioavailable n-3-PUFA compared to those fed with unformulated yoghurt.
Consumption of soup is defined as a healthy dietary lifestyle due to its composition of different plant-based foods such as legumes, tubers and vegetables [3]. Lafarga et al. [114] developed broccoli soup formulated with 0.5 and 2% levels (w/v) of Arthrospira, Chlorella and Tetraselmis species. Upon simulated gastrointestinal digestion, the authors observed enhanced antioxidant capacity with high levels of bioaccessible polyphenols (i.e., between 32.9 and 45.6 mg/100 mL) for each microalgae/broccoli-formulated soup. Irrespective of the improved nutritional value, the authors observed reduced sensory acceptability with increasing levels of microalgae formulation. In contrast to this study, Los et al. [115] formulated a dehydrated soup with A. platensis at 15% (w/w), resulting in improved nutritional value (i.e., protein, fiber, chlorophyll, lipids and antioxidant capacity), with no compromise on acceptability when subjected to sensory evaluation. Based on these reports, there is a need for in-depth studies on formulation strategies that will enable effective microalgal food substitutions (i) for improved functionality and (ii) no/limited sacrifice of sensory quality.
6. Safety Concerns and Acceptability of Microalgae-Based Food Products
The effective contribution of microalgae to human nutrition has great potential. However, the acceptance and application of microalgae for functional food development will be greatly defined by their safety to human health. For instance, the FDA in United States has defined microalgal-based products as “other dietary supplements” with the consumption of strains such as Spirulina, Dunaliella, Chlorella, Haematococcus, C. cohnii, P. cruentum and Schizochytrium reported as GRAS. Compared to the United States, other countries such as Canada, Australia and New Zealand approve the consumption of limited microalgal strains including Spirulina and Chlorella [93]. These limitations are due to the dearth data available with respect to the safety and efficacy of microalgae-based food products/ingredients. However, the few available literature studies on different species of Spirulina and Chlorella have shown non-toxic concentrations of heavy metals, pesticides, antibiotics and mycotoxins. Notwithstanding, there is the need for continuous screening of dietary microalgal strains due to the reported presence of polycyclic aromatic hydrocarbons (PAH) above nutritional threshold levels [116]. Another possible factor for the low application of microalgae in the functional food industry may also be caused by the limited data on human allergic reactions. To increase their utilization and consumer acceptance, there is a need for in-depth research on microalgal allergic compounds, along with the demand of regulatory bodies to ensure food industries cite these possible effects on microalgal-based food labels. Additionally, some researchers and food processors argue on the dominating presence of pigments in different microalgal strains as a possible limiting factor in food applications, with reference to its possible negative effects on taste, color, flavor and other techno-functional properties. However, it can also be discussed that this is not always the case as the effect of a pigment will depend on the type of food product, as well as the group of consumers being considered as a target market.
Another safety concern associated with food applications of microalgae is the high concentration of nucleic acid in its biomass, which has been linked to the negative effects of uric acid (i.e., responsible for gout and kidney stones) in consumers upon metabolism [87]. For example, toxins such as microcystin are elicited by Microcystis aeruginosa, anatoxin by Anabaena flos-aquae and saxitoxin by Aphanizomenon flos-aquae, with these toxic compounds associated with risks of gastrointestinal tract, liver, synapse and nerve axon functioning [117]. Thus, this provokes the conclusion that microalgae cultivation and processing when not conducted under safe conditions and continuous monitoring can lead to detrimental health. Overall, to ensure the safety and nutritional value of microalgal strains intended for human consumption, it is crucial for food industries to commercially cultivate microalgae under controlled environments compared to natural habitats such as lakes and ponds, since this approach can be less costly and also offer an efficient and consistent avenue for the industry to monitor key cultivation factors (e.g., water quality) which contribute to the possible accumulation of toxic compounds.
7. Challenges, Future Perspectives and Conclusions
With the predicted increase in global populations and climate changes, the future is faced with an unprecedented pressure for sustainable food resources capable of providing health benefits beyond basic nutrition. In this regard, microalgae are natural resources that can help meet this gap from the standpoint of sustainability, renewability and nutrition at a cheaper cost. However, some major obstacles concerning the utilization of microalgae as functional foods include accessibility, availability, sustainable techniques for extraction and purification of its metabolites, functional structure preservation during and after extraction and or in a multidimensional food matrix, bioavailability of extracted metabolites in the human gastrointestinal tract upon consumption, safety and sensory quality in formulated food products.
Although genetic engineering has been employed to enhance microalgal composition of bioactive metabolites, there still exists a large proportion of the population with less acceptance and myths for genetically modified foods [118]. In this framework, less attention has been paid on how physical techniques (e.g., pulsed electric field and ultrasonication) and abiotic environmental factors (e.g., pH, temperature, and light) can be manipulated to improve microalgal yield of bioactive metabolites via the induction of oxidative stress, a biotechnological approach known as elicitation. This approach can be proposed as an all-inclusive consumer approach to providing bioactive-rich microalgal biomass. However, investigations on metabolic pathways associated with the production of microalgal bioactive metabolites are also needed, since they will help exploit elicitation techniques targeted for optimized yield of bioactive metabolites. Thus, opening the door for an economically efficient large-scale production and utilization of microalgae as novel sustainable food systems.
With respect to the preservation of bioactive compounds from microalgal biomass during extraction and purification, there is the need to develop methods that cannot only optimize extraction yield but also protect the biochemical structures of less-stable bioactive compounds along the process, especially its functional hydroxyl groups. Another challenge concerning the functionality of microalgae-based bioactive compounds is the fact that a majority of their health benefits are reported with in vitro studies which mimic the human gastrointestinal tract and cannot be the same in reality. For instance, it has been well established that the passage of polyphenols, irrespective of the food matrix, in the stomach result in some being hydrolyzed, with about 90–95% passing into the large intestinal lumen [119]. Thus, it is very important to conduct clinical in vivo studies to not only elucidate the bioaccessibility, bioavailability and biological effects of their bioactive compounds, but to also confirm no side-effects upon consumption.
The pre-dominant sensory issues associated with microalgae-based food products are their intense color and fishy taste/aroma [120]. However, the food industry is taking advantage of the intense color, depending on the type of food product being developed and targeted consumers, as widely reported in the literature and many start-ups. Certainly, techniques such as nanotechnology and the use of safe molecules that act as inhibitors against bitter/astringent receptors are great avenues that can be exploited by the food industry for improved sensory acceptance of microalgae-based foods. Furthermore, it appears as if a majority of microalgae-based foods on the market are in the beverages, baked goods, and snack categories, with very limited application as pro/pre-biotics, although they have the potential for this purpose. A possible way to bridge this is the channeling of resources into the study of how microalgae grow and survive when being delivered alive through foods, as well as their functionalities in the human gut microbiota.
Furthermore, to increase the industrial utilization of microalgae for functional foods, there is a need to develop sustainable and eco-innovative processing techniques that are efficient for the transformation of raw microalgal biomass into value-added food products or isolated nutraceutical ingredients without compromising on their nutritional and environmental effects. With respect to the sustainable processing of microalgal biomass, novel non-thermal processing techniques such as fermentation, ultrasonication, pulsed electric field, microwave and enzyme-assisted processing can be very efficient in helping protect their nutritional value during processing while ensuring food safety concurrently. Lastly, with the increased wide choice of functional foods on the market, there is a need for effective collaboration between researchers and microalgal-based industries to better understand industrial loopholes and consumer needs. This will help guide the design of research outlines that will provide in-depth data on nutritional gaps, especially in vivo bioaccessibility, bioavailability and health benefits of functional compounds associated with microalgae consumption. Nevertheless, whatever the case, food industries should fully address rules and regulations governing the safety, quality and labelling of microalgal-based functional foods before commercialization.
Author Contributions
J.A.: Conceptualization, literature review, writing—original draft, review and editing. L.A.: Writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research did not receive any funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Barreca, D.; Trombetta, D.; Smeriglio, A.; Mandalari, G.; Romeo, O.; Felice, M.R.; Gattuso, G.; Nabavi, S.Y. Food flavonols: Nutraceuticals with Complex Health Benefits and Functionalities. Trends Food Sci. Technol. 2021, 117, 194–204. [Google Scholar] [CrossRef]
- Levasseur, W.; Perré, P.; Pozzobon, V. A Review of High Value-added Molecules Production by Microalgae in Light of the Classification. Biotechnol. Adv. 2020, 41, 1–21. [Google Scholar] [CrossRef]
- Nova, P.; Martins, A.P.; Teixeira, C.; Abreu, H.; Silva, J.G.; Silva, A.M.; Freitas, A.C.; Gomes, A.M. Foods with Microalgae and Seaweeds Fostering Consumers Health: A Review on Scientific and Market Innovations. J. Appl. Phycol. 2020, 32, 1789–1802. [Google Scholar] [CrossRef]
- Ampofo, J.O.; Ngadi, M. Ultrasonic Assisted Phenolic Elicitation and Antioxidant Potential of Common Bean (Phaseolus vulgaris) Sprouts. Ultrason. Sonochem. 2020, 64, 1–11. [Google Scholar] [CrossRef]
- Sidari, R.; Tofalo, R. A Comprehensive Overview on Microalgal Fortified/based Food and Beverages. Food Rev. Int. 2019, 35, 778–805. [Google Scholar] [CrossRef]
- Boussiba, S.; Bing, W.; Yuan, J.P.; Zarka, A.; Chen, F. Changes in Pigments Profile in the Green Alga Haeamtococcus pluvialis Exposed to Environmental Stresses. Biotechnol. Lett. 1999, 21, 601–604. [Google Scholar] [CrossRef]
- Hifney, A.F.; Issa, A.A.; Fawzy, M.A. Abiotic Stress Induced Production of ß-carotene, Allophycocyanin and Total Lipids in Spirulina sp. J. Biol. Earth Sci. 2013, 3, 54–64. [Google Scholar]
- Sandgruber, F.; Gielsdorf, A.; Baur, A.C.; Schenz, B.; Müller, S.M.; Schwerdtle, T.; Stang, G.I.; Grieh, C.; Lorkowski, S.; Dawczynski, C. Variability in Macro- and Micronutrients of 15 Commercially Available Microalgae Powders. Mar. Drugs 2021, 19, 310. [Google Scholar] [CrossRef]
- Fu, W.; Nelson, D.R.; Yi, Z.; Xu, M.; Khraiwesh, B.; Jijakli, K.; Chaiboonchoe, A.; Alzahmi, A.; Al-Khairy, D.; Brynjolfsson, S.; et al. Bioactive Compounds from Microalgae: Current Development and Prospects. In Studies in Natural Products Chemistry; Elsevier, Ltd.: Amsterdam, The Netherlands, 2017; pp. 199–225. [Google Scholar]
- Buckley, M.T.; Racimo, F.; Allentoft, M.E.; Jensen, M.K.; Jonsson, A.; Huang, H.; Hormozdiari, F.; Sikora, M.; Marnetto, D.; Eskin, E.; et al. Selection in Europeans on Fatty Acid Desaturases Associated with Dietary Changes. Mol. Biol. Evol. 2017, 34, 1307–1318. [Google Scholar] [CrossRef] [Green Version]
- Dolganyuk, V.; Andreeva, A.; Budenkova, E.; Sukhikh, S.; Babich, O.; Ivanova, S.; Prosekov, A.; Ulrikh, E. Study of Morphological Features and Determination of the Fatty Acid Composition of the Microalgae Lipid Complex. Biomolecules 2020, 10, 1571. [Google Scholar] [CrossRef]
- Sahin, D.; Tas, E.; Altindag, U.H. Enhancement of Docosahexaenoic Acid (DHA) Production from Schizochytrium sp. S31 Using Different Growth Medium Conditions. AMB Express 2018, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Aussant, J.; Guiheneuf, F.; Stengel, B.D. Impact of Temperature on Fatty Acid Composition and Nutritional Value in Eight Species of Microalgae. Appl. Microbiol. Biotechnol. 2018, 102, 5279–5297. [Google Scholar] [CrossRef] [PubMed]
- Freddy, G.; Dagmar, B.S. LC-PUFA-Enriched Oil Production by Microalgae: Accumulation of Lipid and Triacylglycerols Containing n-3 LC-PUFA Is Triggered by Nitrogen Limitation and Inorganic Carbon Availability in the Marine Haptophyte Pavlova lutheri. Mar. Drugs 2013, 11, 4246–4266. [Google Scholar]
- Khoeyi, Z.A.; Seyfabadi, J.; Ramezanpour, Z. Effect of Light Intensity and Photoperiod on Biomass and Fatty Acid Composition of the Microalgae, Chlorella vulgaris. Aquac. Int. 2012, 20, 41–49. [Google Scholar] [CrossRef]
- Mourelle, M.L.; Gomez, C.; Legido, J.L. The Potential Use of Marine Microalgae and Cyanobacteria in Cosmetics and Thalassotherapy. Cosmetics 2017, 4, 46. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, V.D. Marine Glycans in Relationship with Probiotic Microorganisms to Improve Human and Animal Health. In Marine Glycobiology: Principles and Applications; Kim, S.K., Ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 67–84. [Google Scholar]
- Chandrarathna, H.P.S.U.; Liyanage, T.D.; Edirisinghe, S.L.; Dananjaya, S.H.S.; Thulshan, E.H.T.; Nikapitiya, C.; Oh, C.; Kang, D.H.; De Zoysa, M. Marine Microalgae, Spirulina maxima-Derived Modified Pectin and Modified Pectin Nanoparticles Modulate the Gut Microbiota and Trigger Immune Responses in Mice. Mar. Drugs 2020, 18, 175. [Google Scholar] [CrossRef] [Green Version]
- Matsui, S.M.; Muizzudin, N.; Arad, S.M.; Marenus, K. Sulfated Polysaccharides from Red Microalgae Anti-inflammatory Properties In-vitro and In-vivo. Appl. Biochem. Biotechnol. 2003, 104, 13–22. [Google Scholar] [CrossRef]
- Grossman, A. Nutrient Acquisition: The generation of Bioactive Vitamin B12 by Microalgae. Curr. Biol. 2016, 26, R319–R337. [Google Scholar] [CrossRef] [Green Version]
- Koyande, A.K.; Chew, K.W.; Rambabu, K.; Tao, Y.; Chu, D.T.; Show, P.L. Microalgae: A Potential Alternative to Health Supplementation for Humans. Food Sci. Hum. Wellness 2019, 80, 16–24. [Google Scholar] [CrossRef]
- Ljubic, A.; Jacobsen, C.; Holdt, S.L.; Jakobsen, J. Microalgae Nannochloropsis oceanica as a Future New Natural Source of Vitamin D3. Food Chem. 2020, 320, 1–7. [Google Scholar] [CrossRef]
- Edelmann, M.; Aalto, S.; Chamlagain, B.; Kariluoto, S.; Piironen, V. Riboflavin, Niacin, Folate and Vitamin B12 in Commercial Microalgae Powders. J. Food Compos. Anal. 2019, 82, 1–10. [Google Scholar] [CrossRef]
- Tarento, T.D.C.; McClure, D.D.; Vasiljevski, E.; Schindeler, A.; Dehghani, F.; Kavanagh, J.M. Microalgae as a Source of Vitamin K1. Algal Res. 2018, 36, 77–87. [Google Scholar] [CrossRef]
- Amorim, M.L.; Soares, J.; dos Reis Coimbra, J.S.; de Oliveira Leite, M.; Teixeira Albino, L.F.; Arêdes, M. Microalgae Proteins: Production, Separation, Isolation, Quantification, and Application in Food and Feed. Crit. Rev. Food Sci. Nutr. 2020, 61, 1976–2002. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.C.; Kang, N.; Kim, E.A.; Kang, M.C.; Lee, S.H.; Kang, S.M.; Lee, J.B.; Jeon, B.T.; Kim, S.K.; Park, S.J.; et al. A Novel Angiotensin-I-Converting Enzyme (ACE) Inhibitory Peptide from a Marine Chlorella ellipsoidea and Its Antihypertensive Effect in Spontaneously Hypertensive Rats. Process Biochem. 2012, 470, 2005–2011. [Google Scholar] [CrossRef]
- Khanra, S.; Mondal, M.; Halder, G.; Tiwari, O.N.; Gayen, K.; Bhowmick, T.K. Downstream Processing of Microalgae for Pigments, Protein and Carbohydrate in Industrial Application: A Review. Food Bioprod. Process. 2018, 110, 60–84. [Google Scholar] [CrossRef]
- Ying, K.; Gilmour, D.J.; Zimmerman, W.B. Effects of CO2 and pH on Growth of the Microalga Dunaliella salina. J. Microb. Biochem. Technol. 2014, 6, 167–173. [Google Scholar] [CrossRef]
- Machu, L.; Misurcova, L.; Ambrozova, J.V.; Orsavova, J.; Mlcek, J.; Sochor, J.; Jurikova, T. Phenolic Content and Antioxidant Capacity in Algal Food Products. Molecules 2015, 20, 1118–1133. [Google Scholar] [CrossRef] [Green Version]
- Goiris, K.; Muylaert, K.; Fraeye, I.; Foubert, I.; De Brabanter, J.; De Cooman, L. Antioxidant Potential of Microalgae in Relation to Their Phenolic and Carotenoid Content. J. Appl. Phycol. 2012, 240, 1477–1486. [Google Scholar] [CrossRef]
- Widowati, I.; Zainuri, M.; Kusumaningrum, H.P.; Susilowati, R.; Hardivillier, Y.; Leigne, V.; Bourgougnon, N.; Mouget, J.-L. Antioxidant Activity of Three Microalgae Dunaliella salina, Tetraselmis chuii and Isochrysis galbana Clone Tahiti. IOP Conf. Ser. Earth Environ. Sci. 2017, 55, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food Sources and Bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Rammuni, M.M.; Ariyadasa, T.U.; Nimarshana, P.H.V.; Attalage, R.A. Comparative Assessment on the Extraction of Carotenoids from Microalgal Sources: Astaxanthin from H. pluvialis and β-carotene from D. salina. Food Chem. 2019, 277, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Nagarajan, D.; Zhang, Q.; Chang, J.S.; Lee, D.J. Heterotrophic Cultivation of Microalgae for Pigment Production: A review. Biotechnol. Adv. 2018, 36, 54–67. [Google Scholar] [CrossRef]
- Jaeschke, D.P.; Menegol, T.; Rech, R.; Mercali, G.D.; Marczak, L.D.F. Carotenoid and Lipid Extraction from Heterochlorella luteoviridis Using Moderate Electric Field and Ethanol. Process Biochem. 2016, 51, 1636–1643. [Google Scholar] [CrossRef]
- Sui, Y.; Mazzucchi, L.; Acharya, P.; Xu, Y.; Morgan, G.; Harvey, P.J. A Comparison of β-carotene, Phytoene and Amino Acids Production in Dunaliella salina DF 15 (CCAP 19/41) and Dunaliella salina (CCAP 19/30) Using Different Light Wavelengths. Foods 2021, 10, 2824. [Google Scholar] [CrossRef] [PubMed]
- Inbaraj, B.S.; Chien, J.T.; Chen, B.H. Improved High Performance Liquid Chromatographic Method for Determination of Carotenoids in the Microalga Chlorella pyrenoidosa. J. Chromatogr. A 2006, 1102, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Morowvat, M.H.; Ghasemi, Y. Utilization of Algae for Biofuel, Bio-Products and Bioremediation. Biocatal. Agric. Biotechnol. 2016, 7, 217–223. [Google Scholar] [CrossRef]
- Xia, S.; Wang, K.; Wan, L.; Li, A.; Hu, Q.; Zhang, C. Production, Characterization, and Antioxidant Activity of Fucoxanthin from the Marine Diatom Odontella aurita. Mar. Drugs 2013, 11, 2667–2681. [Google Scholar] [CrossRef]
- Wen, X.; Wang, Z.; Ding, Y.; Geng, Y.; Li, Y. Enhancing the Production of Astaxanthin by Mixotrophic Cultivation of Haematococcus pluvialis in Open Raceway Ponds. Aquac. Int. 2020, 28, 625–638. [Google Scholar] [CrossRef]
- Butler, T.; Golan, Y. Astaxanthin Production from Microalgae. In Microalgae Biotechnology for Food, Health and High Value Products; Alam, M., Xu, J.L., Wang, Z., Eds.; Springer: Singapore, 2020; pp. 175–242. [Google Scholar]
- Murillo, A.G.; Hu, S.; Fernandez, M.L. Zeaxanthin: Metabolism, Properties, and Antioxidant Protection of Eyes, Heart, Liver and Skin. Antioxidants 2019, 8, 390. [Google Scholar] [CrossRef] [Green Version]
- Paliwal, C.; Ghosh, T.; George, B.; Pancha, I.; Maurya, R.; Chokshi, K.; Ghosh, A.; Mishra, S. MicroAlgal Carotenoids: Potential Nutraceutical Compounds with Chemotaxonomic Importance. Algal Res. 2016, 15, 24–31. [Google Scholar] [CrossRef]
- Eltanahy, E.; Torky, A. Microalgae as cell factories: Food and feed-grade high-value metabolites. In Microalgal Biotechnology: Recent Advances, Market Potential, and Sustainability; Royal Society of Chemistry: Cambridge, UK, 2021; pp. 1–35. [Google Scholar]
- Sonani, R.R.; Singh, N.K.; Kumar, J.; Thakar, D.; Madamwar, D. Concurrent Purification and Antioxidant Activity of Phycobiliproteins from Lyngbya sp. A09DM: An Antioxidant and Anti-Aging Potential of Phycoerythrin in Caenorhabditis elegans. Process Biochem. 2014, 49, 1757–1766. [Google Scholar] [CrossRef]
- Ardiles, P.; Cerezal-Mezquita, P.; Salinas-Fuentes, F.; Órdenes, D.; Renato, G.; Ruiz-Dominguez, M.C. Biochemical Composition and Phycoerythrin Extraction from Red Microalgae: A Comparative Study Using Green Extraction Technologies. Processes 2020, 8, 1628. [Google Scholar] [CrossRef]
- Martínez, J.M.; Luengo, E.; Saldaña, G.; Álvarez, I.; Raso, J.C. Phycocyanin Extraction Assisted by Pulsed Electric Field from Artrospira platensis. Int. Food Res. J. 2016, 99, 1042–4047. [Google Scholar] [CrossRef] [PubMed]
- Ores, J.D.C.; Amarante, M.C.A.; de Kalil, S.J. Co-Production of Carbonic Anhydrase and Phycobiliproteins by Spirulina sp. and Synechococcus nidulans. Bioresour. Technol. 2016, 219, 219–227. [Google Scholar] [CrossRef]
- Khandual, S.; Sanchez, E.O.L.; Andrews, H.E.; Padilla de la Rosa, J.D. Phycocyanin Content and Nutritional Profile of Arthrospira platensis from Mexico: Efficient Extraction Process and Stability Evaluation of Phycocyanin. BMC Chem. 2021, 15, 24. [Google Scholar] [CrossRef]
- Greque de Morais, M.; da Fontoura Prates, D.; Moreira, J.B.; Duarte, J.H.; Costa, J.A.V. Phycocyanin from Microalgae: Properties, Extraction and Purification, with Some Recent Applications. Ind. Bio-Technol. 2018, 14, 30–37. [Google Scholar] [CrossRef]
- Ajayan, K.V.; Selvaraju, M.; Thirugnanamoorthy, K. Enrichment of Chlorophyll and Phycobiliproteins in Spirulina platensis by the Use of Reflector Light and Nitrogen Sources: An In-Vitro Study. Biomass Bioenergy 2012, 47, 436–441. [Google Scholar] [CrossRef]
- López-Hernández, J.F.; García-Alamilla, P.; Palma-Ramírez, D.; Álvarez-González, C.A.; Paredes-Rojas, J.C.; Márquez-Rocha, F.J. Continuous Microalgal Cultivation for Antioxidants Production. Molecules 2020, 25, 4171. [Google Scholar] [CrossRef]
- Jia, Y.P.; Sun, L.; Yu, H.S.; Liang, L.P.; Li, W.; Ding, H.; Song, X.B.; Zhang, L.J. The Pharmacological Effects of Lutein and Zeaxanthin on Visual Disorders and Cognition Diseases. Molecules 2017, 22, 610. [Google Scholar] [CrossRef]
- Ho, S.H.; Chan, M.C.; Liu, C.C.; Chen, C.Y.; Lee, W.L.; Lee, D.J.; Chang, J.S. Enhancing Lutein Productivity of an Indigenous Microalga Scenedesmus Obliquus FSP-3 using Light-Related Strategies. Bioresour. Technol. 2014, 152, 275–282. [Google Scholar] [CrossRef]
- Sun, Z.; Li, T.; Zhou, Z.G.; Jiang, Y. Microalgae as a source of lutein: Chemistry, biosynthesis, and carotenogenesis. In Advances in Biochemical Engineering/Biotechnology; Springer: Berlin/Heidelberg, Germany, 2016; pp. 37–58. [Google Scholar]
- Lin, J.H.; Lee, D.J.; Chang, J.S. Lutein Production from Biomass: Marigold Flowers Versus Microalgae. Bioresour. Technol. 2015, 184, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.G.; Otero, P.; Echave, J.; Carreira-Casais, A.; Chamorro, F.; Collazo, N.; Jaboui, A.; Lourenço-Lopes, C.; Simal-Gandara, J.; Prieto, M.A. Xanthophylls from the Sea: Algae as Source of Bioactive Carotenoids. Mar. Drugs 2021, 19, 188. [Google Scholar] [CrossRef] [PubMed]
- Chuyen, H.V.; Eun, J.B. Marine Carotenoids: Bioactivities and Potential Benefits to Human Health. Crit. Rev. Food Sci. Nutr. 2017, 57, 2600–2610. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.; Kim, M.B.; Park, Y.K.; Lee, J.Y. Health Benefits of Fucoxanthin in the Prevention of Chronic Diseases. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2020, 1865, 158618. [Google Scholar] [CrossRef]
- Mularczyk, M.; Michalak, I.; Marycz, K. Astaxanthin and Other Nutrients from Haematococcus Pluvialis-Multifunctional Applications. Mar. Drugs 2020, 18, 459. [Google Scholar] [CrossRef]
- Lim, K.C.; Yusoff, F.M.; Shariff, M.; Kamarudin, M.S. Astaxanthin as Feed Supplement in Aquatic Animals. Rev. Aquac. 2018, 10, 738–773. [Google Scholar] [CrossRef]
- Kim, J.H.; Chang, M.J.; Choi, H.D.; Youn, Y.K.; Kim, J.T.; Oh, J.M.; Shin, W.G. Protective Effects of Haematococcus Astaxanthin on Oxidative Stress in Healthy Smokers. J. Med. Food 2011, 14, 1469–1475. [Google Scholar] [CrossRef]
- Yoshida, H.; Yanai, H.; Ito, K.; Tomono, Y.; Koikeda, T.; Tsukahara, H.; Tada, N. Administration of Natural Astaxanthin Increases Serum HDL-Cholesterol and Adiponectin in Subjects with Mild Hyperlipidemia. Atherosclerosis 2010, 209, 520–530. [Google Scholar] [CrossRef]
- Ravi, K.B.; Reddy, K.R.R.; Shankaranarayanan, J.; Deshpande, J.V.; Juturu, V.; Soni, M.G. Safety Evaluation of Zeaxanthin Concentrate (OmniXan™): Acute, Subchronic Toxicity and Mutagenicity Studies. Food Chem. Toxicol. 2014, 72, 30–3931. [Google Scholar] [CrossRef]
- Suparmi, S.; Fasitasari, M.; Martosupono, M.; Mangimbulude, J.C. Comparisons of Curative Effects of Chlorophyll from Sauropus androgynus (L) Merr Leaf Extract and Cu-Chlorophyllin on Sodium Nitrate-Induced Oxidative Stress in Rats. J. Toxicol. 2016, 2016, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Manirafasha, E.; Ndikubwimana, T.; Zeng, X.; Lu, Y.; Jing, K. Phycobiliprotein: Potential Microalgae Derived Pharmaceutical and Biological Reagent. Biochem. Eng. J. 2016, 109, 282–296. [Google Scholar] [CrossRef]
- Kannaujiya, V.K.; Singh, P.R.; Kumar, D.; Sinha, R.P. Phycobiliproteins in microalgae: Occurrence, distribution, and biosynthesis. In Pigments from Microalgae Handbook; Jacob-Lopes, E., Queiroz, M., Zepka, L., Eds.; Springer: Cham, Switzerland, 2020; pp. 43–68. [Google Scholar]
- Singh, S.P.; Kumari, S.; Rastogi, R.P.; Singh, K.L.; Sinha, R.P. Mycosporine-like Amino Acids (MAAs): Chemical Structure, Biosynthesis and Significance as UV Absorbing/Screening Compounds. Indian J. Exp. Biol. 2008, 146, 7–17. [Google Scholar]
- Koizumi, K.; Hatakeyama, M.; Boero, M.; Nobusada, K.; Hori, H.; Misonou, T.; Nakamura, S. How Seaweeds Release the Excess Energy from Sunlight to Surrounding Sea Water. Phys. Chem. Chem. Phys. 2017, 19, 15745–15753. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Lee, M.G.; Seo, H.J.; Shin, J.H.; Shin, T.S.; Yoon, Y.H.; Kim, M.Y.; Choi, J.I.; Kim, J.D. Enhancing the Feasibility of Microcystis Aeruginosa as a Feedstock for Bioethanol Production Under the Influence of Various Factors. Biomed. Res. Int. 2016, 2016, 1–9. [Google Scholar]
- Plat, J.; Baumgartner, S.; Vanmierlo, T.; Lutjohann, D.; Calkins, K.L.; Burrin, D.G.; Guthrie, G.; Thijs, C.; Te Velde, A.A.; Vreugdenhil, A.C.E.; et al. Plant-Based Sterols and Stanols in Health & Disease: “Consequences of Human Development in a Plant-Based Environment?”. Prog. Lipid Res. 2019, 74, 87–102. [Google Scholar]
- Randhir, A.; Laird, D.W.; Maker, G.; Trengove, R.; Moheimani, N.R. Microalgae: A Potential Sustainable Commercial Source of Sterols. Algal Res. 2020, 46, 101772. [Google Scholar] [CrossRef]
- Ahmed, F.; Zhou, W.; Schenk, P.M. Pavlova lutheri is a High-Level Producer of Phytosterols. Algal Res. 2015, 10, 210–217. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.F.; de Morais, A.M.M.B.; de Morais, R.M.S.C. Carotenoids from Marine MicroAlgae: A Valuable Natural Source for the Prevention of Chronic Diseases. Mar. Drugs 2015, 13, 5128–5155. [Google Scholar] [CrossRef]
- Kent, M.; Welladsen, H.M.; Mangott, A.; Li, Y. Nutritional Evaluation of Australian Microalgae as Potential Human Health Supplements. PLoS ONE 2015, 10, e0118985. [Google Scholar] [CrossRef]
- Sadat-Ebrahimi, S.R.; Parnianfard, N.; Vahed, N.; Babaei, H.; Ghojazadeh, M.; Tang, S.; Azarpazhooh, A. An Evidence-Based Systemic Review of the Off-Label uses of Lisinopril. Br. J. Clin. Pharmacol. 2018, 84, 2502–2521. [Google Scholar] [CrossRef] [Green Version]
- Desbois, A.P.; Walton, M.; Smith, V.J. Differential Antibacterial Activities of Fusiform and Oval Morphotypes of Phaeodactylum tricornutum (Bacillariophyceae). J. Mar. Biol. Assoc. 2010, 90, 769–774. [Google Scholar] [CrossRef] [Green Version]
- Scaglioni, P.T.; de Oliveira Garcia, S.; Badiale-Furlong, E. Inhibition of In-Vitro Trichothecenes Production by Microalgae Phenolic Extracts. Int. Food Res. J. 2019, 124, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Sansone, C.; Galasso, C.; Orefice, I.; Nuzzo, G.; Luongo, E.; Cutignano, A.; Romano, G.; Brunet, C.; Fontana, A.; Esposito, F.; et al. The Green Microalga Tetraselmis suecica Reduces Oxidative Stress and Induces Repairing Mechanisms in Human Cells. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.S. Omega-3 and Omega-6 Polyunsaturated Fatty Acids: Dietary Sources, Metabolism and Significance—A Review. Life Sci. 2018, 15, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Kusaikin, M.I.; Ermakova, S.P.; Shevchenko, N.M.; Isakov, V.V.; Gorshkov, A.G.; Vereshchagin, A.L.; Grachev, M.A.; Zvyagintseva, T.N. Structural Characteristics and Antitumor Activity of a New Chrysolaminaran from the Diatom Alga Synedra acus. Chem. Nat. Compd. 2010, 46, 1–4. [Google Scholar] [CrossRef]
- Liu, M.; Li, W.; Chen, Y.; Wan, X.; Wang, J. Fucoxanthin: A Promising Compound for Human Inflammation-Related Diseases. Life Sci. 2020, 255, 1178503. [Google Scholar] [CrossRef]
- Murai, U.; Yamagishi, K.; Sata, M.; Kokubo, Y.; Saito, I.; Yatsuya, H.; Ishihara, J.; Inoue, M.; Sawada, N.; Iso, H.; et al. Seaweed Intake and Risk of Cardiovascular Disease: The Japan Public Health Center-based Prospective (JPHC) Study. Am. J. Clin. Nutr. 2019, 110, 1449–1455. [Google Scholar] [CrossRef]
- Dvir, I.; Stark, A.H.; Chayoth, R.; Madar, Z.; Arad, S.M. Hypocholesterolemic Effects of Nutraceuticals Produced from the Red Microalga Porphyridium sp. in Rats. Nutrients 2009, 1, 156–167. [Google Scholar] [CrossRef] [Green Version]
- Almeida, T.P.; Ferreira, J.; Vettorazzi, A.; Azqueta, A.; Rocha, E.; Ramos, A.A. Cytotoxic Activity of Fucoxanthin, Alone and in Combination with the Cancer Drugs Imatinib and Doxorubicin, in CML Cell Lines. Environ. Toxicol. Pharmacol. 2018, 59, 24–33. [Google Scholar] [CrossRef]
- Cianciosi, D.; Varela-Lopez, A.; Forbes-Hernandez, T.Y.; Gasparrini, M.; Afrin, S.; Reboredo-Rodriguez, P.; Zhang, J.J.; Quiles, J.L.; Nabavi, S.F.; Battino, M.; et al. Targeting Molecular Pathways in Cancer Stem Cells by Natural Bioactive Compounds. Pharmacol. Res. 2018, 135, 150–165. [Google Scholar] [CrossRef]
- Nethravathy, M.U.; Jitendra, G.M.; Sandeep, N.M.; Ajam, Y.S. Recent Advances in Microalgal Bioactives for Food, Feed, and Healthcare Products: Commercial Potential, Market Space, and Sustainability. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1882–1897. [Google Scholar]
- Abd El Baky, H.H.; El Baroty, G.S.; Ibrahem, E.A. Functional Characters Evaluation of Biscuits Sublimated with Pure Phycocyanin Isolated from Spirulina and Spirulina biomass. Nutr. Hosp. 2015, 32, 231–241. [Google Scholar] [PubMed]
- Singh, P.; Singh, R.; Jha, A.; Rasane, P.; Gautam, A.K. Optimization of a Process for High Fibre and High Protein Biscuit. J. Food Sci. Technol. 2015, 52, 1394–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista, A.P.; Niccolai, A.; Fradinho, P.; Fragoso, S.; Bursic, I.; Rodolfi, L. Microalgae Biomass as an Alternative Ingredient in Cookies: Sensory, Physical and Chemical Properties, Antioxidant Activity and In-Vitro Digestibility. Algal Res. 2017, 26, 161–171. [Google Scholar] [CrossRef]
- Mofasser-Hossain, A.K.M.; Brennan, M.A.; Mason, S.L.; Guo, X.; Zeng, X.A.; Brennan, C.S. The Effect of Astaxanthin-Rich Microalgae “Haematococcus pluvialis” and Wholemeal Flours Incorporation in Improving the Physical and Functional Properties of Cookies. Foods 2017, 6, 1–10. [Google Scholar]
- Ak, B.; Avşaroglu, E.; Işik, O.; Özyurt, G.; Kafkas, E.; Etyemez, M. Nutritional and Physicochemical Characteristics of Bread Enriched with Microalgae Spirulina platensis. Int. J. Eng. Res. Appl. 2016, 6, 30–38. [Google Scholar]
- Garcıa, J.L.; Vicente, D.M.; Galan, B. Microalgae, Old Sustainable Food and Fashion Nutraceuticals. Microb. Biotechnol. 2017, 10, 1017–1024. [Google Scholar] [CrossRef] [Green Version]
- Lucas, B.F.; de Morais, M.G.; Santos, T.D.; Costa, J.A.V. Spirulina for Snack Enrichment: Nutritional, Physical and Sensory Evaluations. LWT Food Sci. Technol. 2018, 90, 270–276. [Google Scholar] [CrossRef]
- Gouveia, L.; Batista, A.P.; Miranda, A.; Empis, J.; Raymundo, A. Chlorella vulgaris Biomass Used as Colouring Source in Traditional Butter Cookies. Innov. Food Sci. Emerg. Technol. 2007, 8, 433–436. [Google Scholar] [CrossRef]
- Da Silva, S.P.; Ferreira do Valle, A.; Perrone, D. Microencapsulated Spirulina maxima Biomass as an Ingredient for the Production of Nutritionally Enriched and Sensorially Well-Accepted Vegan Biscuits. LWT Food Sci. Technol. 2021, 142, 1–7. [Google Scholar] [CrossRef]
- Sahni, P.; Sharma, S.; Singh, B. Evaluation and Quality Assessment of Defatted Microalgae Meal of Chlorella as an Alternative Food Ingredient in Cookies. Nutr. Food Sci. 2019, 49, 221–231. [Google Scholar] [CrossRef]
- Onacik-Gür, S.; Zbikowska, A.; Majewska, B. Effect of Spirulina (Spirulina platensis) Addition on Textural and Quality Properties of Cookies. Ital. J. Food Sci. 2017, 30, 1–12. [Google Scholar]
- Kumoro, A.C.; Johnny, D.; Alfilovita, D. Incorporation of Microalgae and Seaweed in Instant Fried Wheat Noodles Manufacturing: Nutrition and Culinary Properties Study. Int. Food Res. J. 2016, 23, 715–722. [Google Scholar]
- De Marco, E.R.; Steffolani, M.E.; Martínez, C.S.; León, C.S. Effects of Spirulina Biomass on the Technological and Nutritional Quality of Bread Wheat Pasta. LWT Food Sci. Technol. 2014, 58, 102–108. [Google Scholar] [CrossRef]
- El-Baz, F.K.; Abdo, S.M.; Hussein, A.M.S. Microalgae Dunaliella salina for Use as Food Supplement to Improve Pasta Quality. Int. J. Pharm. Sci. Rev. Res. 2017, 46, 45–51. [Google Scholar]
- Fradique, M.; Batista, A.P.; Nunes, M.C.; Gouveia, L.; Bandarra, N.M.; Raymundo, A. Incorporation of Chlorella vulgaris and Spirulina maxima Biomass in Pasta Products. Part 1: Preparation and Evaluation. J. Sci. Food Agric. 2010, 90, 1656–1664. [Google Scholar] [CrossRef] [PubMed]
- Raja, R.; Coelho, A.; Hemaiswarya, S.; Kumar, P.; Carvalho, I.S.; Alagarsamy, A. Applications of Microalgal Paste and Powder as Food and Feed: An Update Using Text Mining Tool. Beni-Suef Univ. J. Basic Appl. Sci. 2018, 7, 740–747. [Google Scholar] [CrossRef]
- Henchion, M.; Hayes, M.; Mullen, A.; Fenelon, M.; Tiwari, B. Future Protein Supply and Demand: Strategies and Factors Influencing a Sustainable Equilibrium. Foods 2017, 6, 53. [Google Scholar] [CrossRef] [Green Version]
- Bolanho, B.C.; Egea, M.B.; Jácome, A.L.M.; Campos, I.; De Carvalho, J.C.M.; Danesi, E.D.G. Antioxidant and Nutritional Potential of Cookies Enriched with Spirulina platensis and Sources of Fibre. J. Food Nutr. Res. 2014, 53, 171–179. [Google Scholar]
- Niccolai, A.; Venturi, M.; Galli, V.; Pini, N.; Rodolfi, L.; Biondi, N.; D’Ottavio, M.; Batista, P.A.; Raymundo, A.; Granchi, L.; et al. Development of New Microalgae-Based Sourdough “Crostini”: Functional Effects of Arthrospira platensis (Spirulina) Addition. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Angelino, D.; Cossu, M.; Marti, A.; Zanoletti, M.; Chiavaroli, L.; Brighenti, F.; Del Rio, D.; Martini, D. Bioaccessibility and Bioavailability of Phenolic Compounds in Bread: A Review. Food Funct. J. 2017, 8, 2368–2393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uribe-Wandurraga, Z.N.; Igual, M.; García-Segovia, P.; Martínez-Monzó, J. Effect of Microalgae Addition on Mineral Content, Colour and Mechanical Properties of Breadsticks. Food Funct. J. 2019, 10, 4685–4692. [Google Scholar] [CrossRef] [PubMed]
- Graça, C.; Fradinho, P.; Sousa, I.; Raymundo, A. Impact of Chlorella vulgaris on the Rheology of Wheat Flour Dough and Bread Texture. LWT Food Sci. Technol. 2018, 89, 466–474. [Google Scholar] [CrossRef]
- Hafsa, Y.A.; Amel, D.; Samia, S.; Sidahmed, S. Evaluation of Nutritional and Sensory Properties of Bread Enriched with Spirulina. Ann. Rev. Food Sci. Technol. 2014, 15, 270–275. [Google Scholar]
- Da Silva Vaz, B.; Moreira, J.B.; de Morais, M.G.; Costa, J.A.V. Microalgae as a New Source of Bioactive Compounds in Food Supplements. Curr. Opin. Food Sci. 2016, 7, 73–77. [Google Scholar]
- Tohamy, M.M.; Ali, M.A.; Shaaban, H.A.; Mohammas, A.G.; Hasanain, A.M. Production of Functional Spreadable Processed Cheese using Chlorella vulgaris. Acta Sci. Pol. Technol. Aliment. 2018, 17, 347–358. [Google Scholar]
- Lane, K.E.; Li, W.; Smith, C.; Derbyshire, E. The Bioavailability of an Omega-3-rich Algal Oil is Improved by Nanoemulsion Technology Using Yogurt as a Food Vehicle. Int. J. Food Sci. Technol. 2014, 49, 1264–1271. [Google Scholar] [CrossRef] [Green Version]
- Lafarga, T.; Acién-Fernández, F.G.; Castellari, M.; Villaró, S.; Bobo, G.; Aguiló-Aguayo, I. Effect of Microalgae Incorporation on the Physicochemical, Nutritional, and Sensorial Properties of an Innovative Broccoli Soup. LWT Food Sci. Technol. 2019, 111, 167–174. [Google Scholar] [CrossRef]
- Los, P.R.; Simões, D.R.S.; de Leone, R.S.; Bolanho, B.C.; Crdoso, T.; Danesi, E.D.G. Viability of Peach Palm By-Product, Spirulina platensis, and Spinach for the Enrichment of Dehydrated Soup. Pesqui. Agropecu. Bras. 2018, 53, 1259–1267. [Google Scholar] [CrossRef]
- Muys, M.; Sui, Y.; Schwaiger, B.; Lesueur, C.; Vandenheuve, D.; Vermeir, P.; Vlaeminck, S.E. High Variability in Nutritional Value and Safety of Commercially Available Chlorella and Spirulina Biomass Indicates the Need for Smart Production Strategies. Bioresour. Technol. 2019, 275, 247–257. [Google Scholar] [CrossRef]
- Kratzer, R.; Murkovic, M. Food Ingredients and Nutraceuticals from Microalgae: Main Product Classes and Biotechnological Production. Foods 2021, 10, 1626. [Google Scholar] [CrossRef] [PubMed]
- Paull, J. The Threat of Genetically Modified Organisms (GMOs) to Organic Agriculture: A Case Study Update. Agric. Food 2013, 3, 56–63. [Google Scholar]
- Faria, A.; Fernandes, I.; Norberto, S.; Mateus, N.; Calhau, C. Interplay Between Anthocyanins and Gut Microbiota. J. Agric. Food Chem. 2014, 62, 6898–6902. [Google Scholar] [CrossRef] [PubMed]
- Camacho, F.; Macedo, A.; Malcata, F. Potential Industrial Applications and Commercialization of Microalgae in the Functional Food and Feed Industries: A Short Review. Mar. Drugs 2019, 17, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Bioactive composition of microalgal biomass. PUFA—polyunsaturated fatty acids; MAAs—mycosporine-like amino acids.
Figure 1.
Bioactive composition of microalgal biomass. PUFA—polyunsaturated fatty acids; MAAs—mycosporine-like amino acids.

Figure 2.
Bibliometric analysis on “microalgae and bioactive compounds” from the Web of Science database. The chart displays published research papers from January 1991 to December 2021.
Figure 2.
Bibliometric analysis on “microalgae and bioactive compounds” from the Web of Science database. The chart displays published research papers from January 1991 to December 2021.

Figure 3.
Schematic description of microalgae health benefits and potential food applications.

Figure 4.
Snacks enriched with different substitution levels (2 and 6%) of microalgae biomass. Ap—A. platensis; Cv—C. vulgaris; Ts—T. suecica; Pt—P. tricornutum. Figure adapted from Batista et al. [90].
Figure 4.
Snacks enriched with different substitution levels (2 and 6%) of microalgae biomass. Ap—A. platensis; Cv—C. vulgaris; Ts—T. suecica; Pt—P. tricornutum. Figure adapted from Batista et al. [90].

Figure 5.
Sourdough crostini developed from A. platensis F&M-C256 at formulation levels 2, 6 and 10%, w/w. C—control; Ap—A. platensis. Figure adapted from Niccolai et al. [106]; Source—Springer Nature; https://creativecommons.org/licenses/by/4.0/legalcode, accessed on 27 April 2022; no changes were made to the figure in re-printing.
Figure 5.
Sourdough crostini developed from A. platensis F&M-C256 at formulation levels 2, 6 and 10%, w/w. C—control; Ap—A. platensis. Figure adapted from Niccolai et al. [106]; Source—Springer Nature; https://creativecommons.org/licenses/by/4.0/legalcode, accessed on 27 April 2022; no changes were made to the figure in re-printing.

Figure 6.
Different food formulations from Spirulina sp. ((a) isotonic beverages; (b) cereal bars; (c) instant soup; (d) pudding; (e) cake powder mix; (f) biscuit). Adapted with permission from [111], 2016, Elsevier Ltd.
Figure 6.
Different food formulations from Spirulina sp. ((a) isotonic beverages; (b) cereal bars; (c) instant soup; (d) pudding; (e) cake powder mix; (f) biscuit). Adapted with permission from [111], 2016, Elsevier Ltd.


{kind=link}
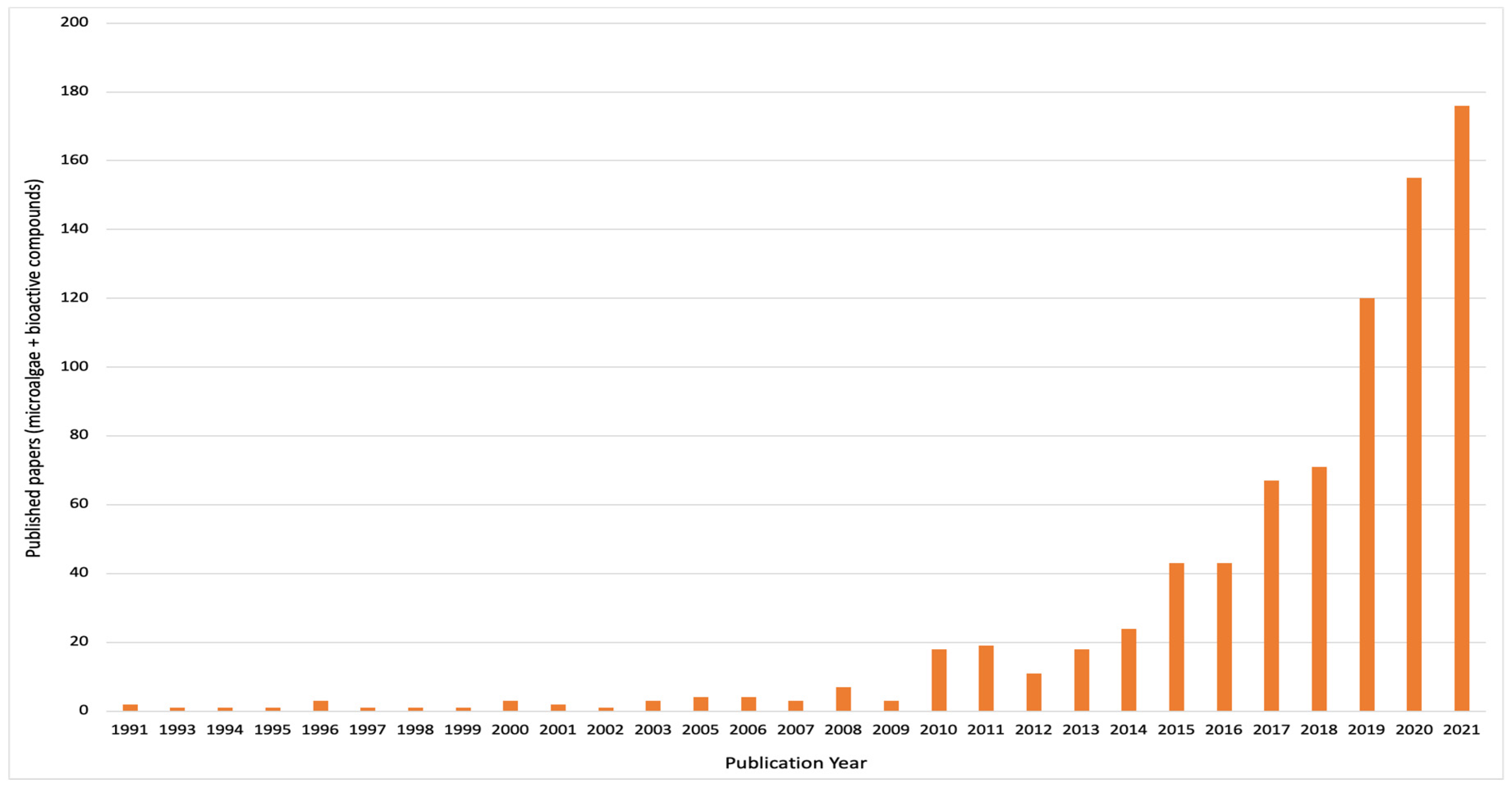{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Health benefits of secondary metabolites from selected microalgal species.
| Genus/Species | Metabolite | Concentration | Bioactive Effect | References |
|---|---|---|---|---|
| Dunaliella salina | Lutein | 0.4–0.8% | Antioxidative, anticancer, cataract and macular degeneration protection | [9,35] |
| Chlorella sorokiniana | 5.21 mg/g | |||
| Heterochlorella luteoviridis | β-carotene | 0.19 mg/g | Anticancer, antioxidative, antihypertensive, neuroprotective, protection against macular degeneration, anticholesterol | [36,38] |
| Dunaliella salina | 0.01–15.0 g/L | |||
| Chaetoceros sp. | Fucoxanthin | 1.5–2.0% 5.13 mg/g | Antioxidative, anticancer, anti-cholesterol, antidiabetic, antitumor | [39] |
| Odontella sp. | ||||
| Chaetoceros sp. | ||||
| Haematococcus pluvialis | Astaxanthin | 1.95–2.75% | Antibacterial, anticancer, anti-inflammatory, antioxidative, neuroprotective, antimicrobial | [40] |
| Chlorella zofingiensis | Astaxanthin | 1.5% | Antibacterial, anticancer, anti-inflammatory, antioxidative, neuroprotective, antimicrobial | [41] |
| Heterochlorella luteoviridis | Zeaxanthin | 0.244 mg/g | Improve eye health, antidiabetic | [35,42,43,44,45,46] |
| Dunaliella salina | 6 mg/g | |||
| Lyngbya sp. | 0.13 mg/g | |||
| Lyngbya sp. | 27.47 mg/g | |||
| Anabaena sp. Cyanidium caldarium | Phycocyanin | 8.3% 4–53 mg/g | Neuroprotective, anti-inflammatory, antioxidative, hepatoprotective, antiaging, antitumor | [47,48,49,50] |
| Spirulina platensis Arthrospira platensis | 6.5–152 mg/g 54.65 mg/g | |||
| Oscillatoria quadripunctulata | 27.43 mg/g | |||
| Spirulina platensis | Allophycocy-anins | 26.8–45.2 mg/g | Hepatoprotective, antioxidative, anti-inflammatory | [47,51,52] |
| Spirulina sp. Porphyridium cruentum | 20–70 mg/g 13.20 mg/g | |||
| Spirulina platensis | 92 mg/g |
Table 2.
Nutritional and techno-functional properties of microalgae-based formulated food products.
| Food Product | Genus/Species | Formulation Levels | Measured Effects | References |
|---|---|---|---|---|
| Biscuits | Arthrospira platensis | Whole A. platensis (0.3, 0.6 and 0.9%); phycocyanin extract: 0.3% w/w to wheat flour | Increased nutritional properties | [88] |
| Biscuits | Arthrospira platensis | 1.63, 3, 5, 7 and 8.36% w/w to wheat flour | Improved techno-functional attributes and enhanced protein, fiber and antioxidant levels | [89] |
| Biscuits | Arthrospira platensis, Chlorella vulgaris, Tetraselmis suecica and P. tricornutum | 2 and 6% w/w to wheat flour | Improved techno-functional and antioxidant properties | [90] |
| Cookies | Haematococcus pluvialis | Extracted astaxanthin powder at 5, 10 and 15% w/w to wheat flour | Enhanced techno-functional and antioxidant properties | [91] |
| Bread | Arthrospira platensis | 11% w/w to flour | Improved techno-functional attributes, protein and mineral levels | [92] |
| Bread | Isochrysis galbana, Tetraselmis suecica, S. almeriensis, and N. gaditana | 0.47% w/w to wheat flour | Increased techno-functional attributes | [93] |
| Extruded snacks | Arthrospira sp. | 0.4, 1.0, 1.8, 2.6, and 3.2%, w/w to flour | Increased techno- functional attributes and protein levels | [94] |
| Butter cookies | Chlorella vulgaris | 0.5, 1.0, 2.0 and 3.0%, w/w to wheat flour | Improved textural and color attributes | [95] |
| Biscuits | Spirulina maxima | 5, 10 and 15%, w/w of wheat flour | Improved concentrations of protein, iron and PUFAs | [96] |
| Cookies | Chlorella sp. | 3, 6, 9 and 12%, w/w of wheat flour | Improved techno-functional and sensory attributes | [97] |
| Spirulina platensis | 0, 1, 2 and 3%, w/w of wheat flour | Increased textural and colour properties; low overall sensory quality | [98] | |
| Noodles | Chlorella vulgaris and Spirulina platensis | 2.5, 5, 7.5, 10, 12.5, 15, 20 and 25 g, w/w of wheat flour | Enhanced techno-functionalities, protein, ash and dietary fiber; reduced carbohydrate content | [99] |
| Pasta | Spirulina platensis | 5, 10 and 20 g, w/w to wheat flour | Enhanced techno-functionalities, protein, phenolics and antioxidant properties; limited protein digestibility | [100] |
| Dunaliella salina | 0, 1, 2 and 3%, w/w to wheat flour | Improved protein, minerals, antioxidants, techno-functional and sensory properties | [101] | |
| Chlorella vulgaris and Spirulina maxima | 0.5, 1 and 2%, w/w of wheat flour | Elevated techno-functional and sensory qualities | [102] | |
| Noodles | Spirulina sp. | 3–5%, w/w of wheat flour | Improved nutritional quality | [103] |
| Protein drink | Spirulina sp. | 10–12% | Improved protein content | [103] |
| Noodles | Chlorella sp. | - | Enhanced nutritional value | [103] |
| Energy bar | Chlorella sp. | 5–10% | Enhanced protein content | [103] |
| Burger | Chlorella sp. | 50% | Improved textural and sensory qualities | [104] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ampofo, J.; Abbey, L. Microalgae: Bioactive Composition, Health Benefits, Safety and Prospects as Potential High-Value Ingredients for the Functional Food Industry. Foods 2022, 11, 1744. https://doi.org/10.3390/foods11121744
AMA Style
Ampofo J, Abbey L. Microalgae: Bioactive Composition, Health Benefits, Safety and Prospects as Potential High-Value Ingredients for the Functional Food Industry. Foods. 2022; 11(12):1744. https://doi.org/10.3390/foods11121744
Chicago/Turabian StyleAmpofo, Josephine, and Lord Abbey. 2022. "Microalgae: Bioactive Composition, Health Benefits, Safety and Prospects as Potential High-Value Ingredients for the Functional Food Industry" Foods 11, no. 12: 1744. https://doi.org/10.3390/foods11121744
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.