
Effects of Monascus on Proteolysis, Lipolysis, and Volatile Compounds of Camembert-Type Cheese during Ripening


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cheese Manufacture and Sampling
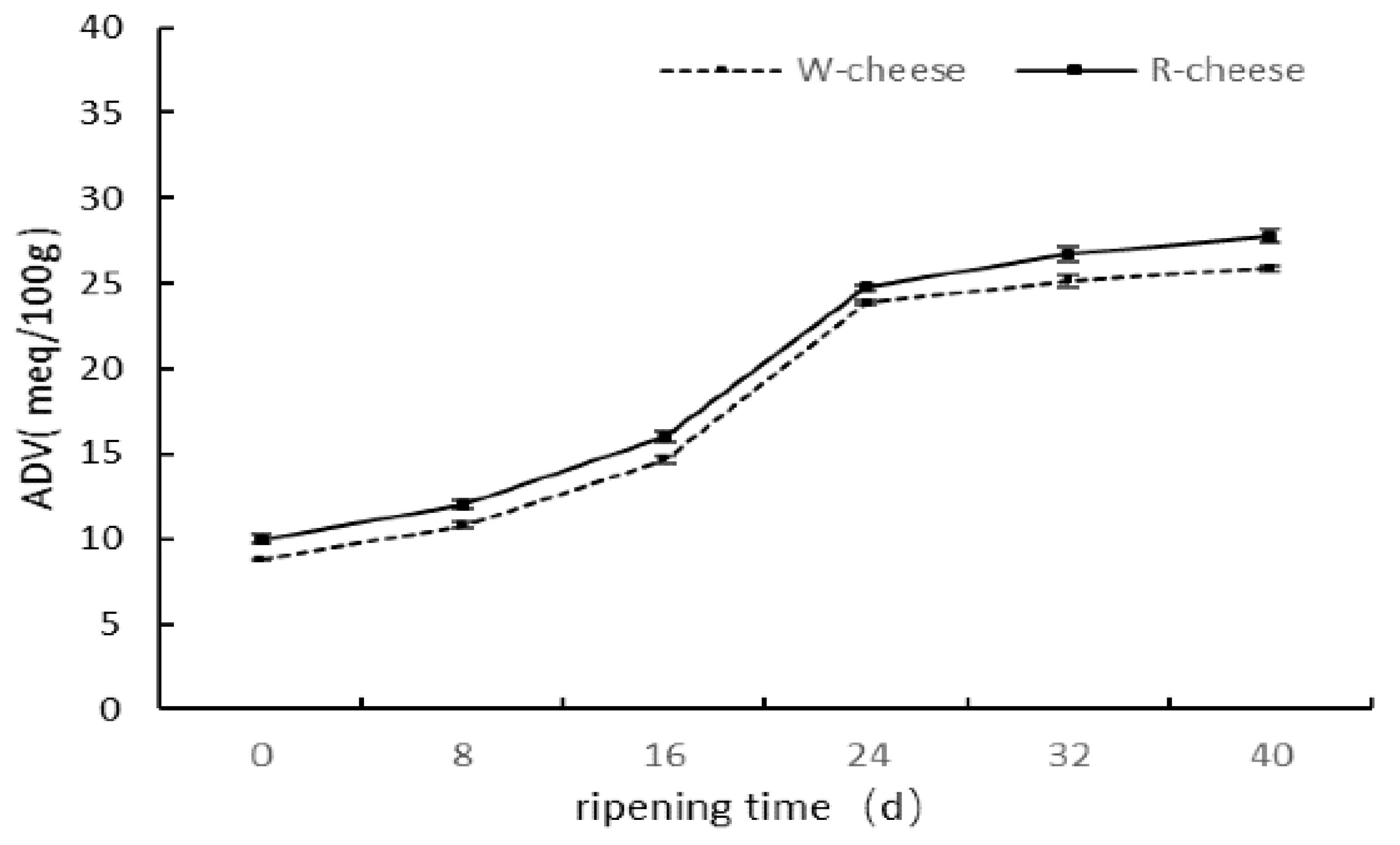
2.3. Proteolysis
2.4. Lipolysis
2.5. Volatile Compounds
2.6. Statistical Analysis
3. Results and Discussion
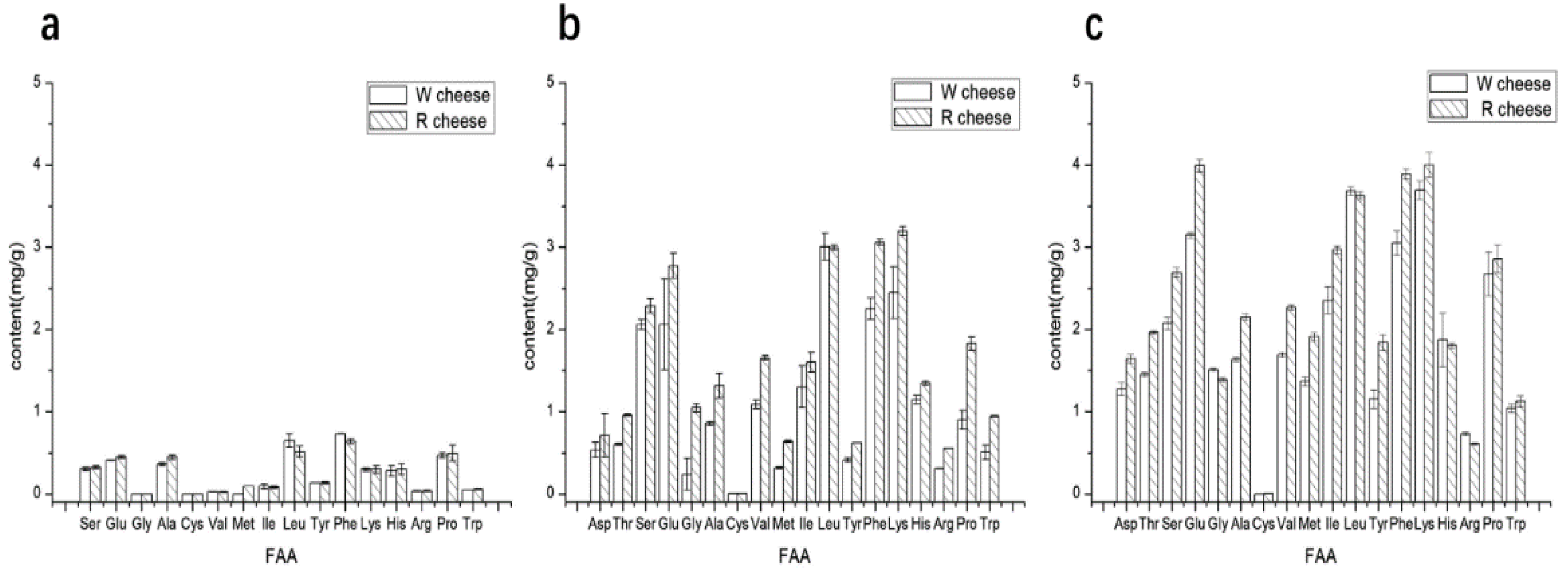
3.1. Effects of Monascus on Proteolysis and FAA of Camembert-Type Cheese
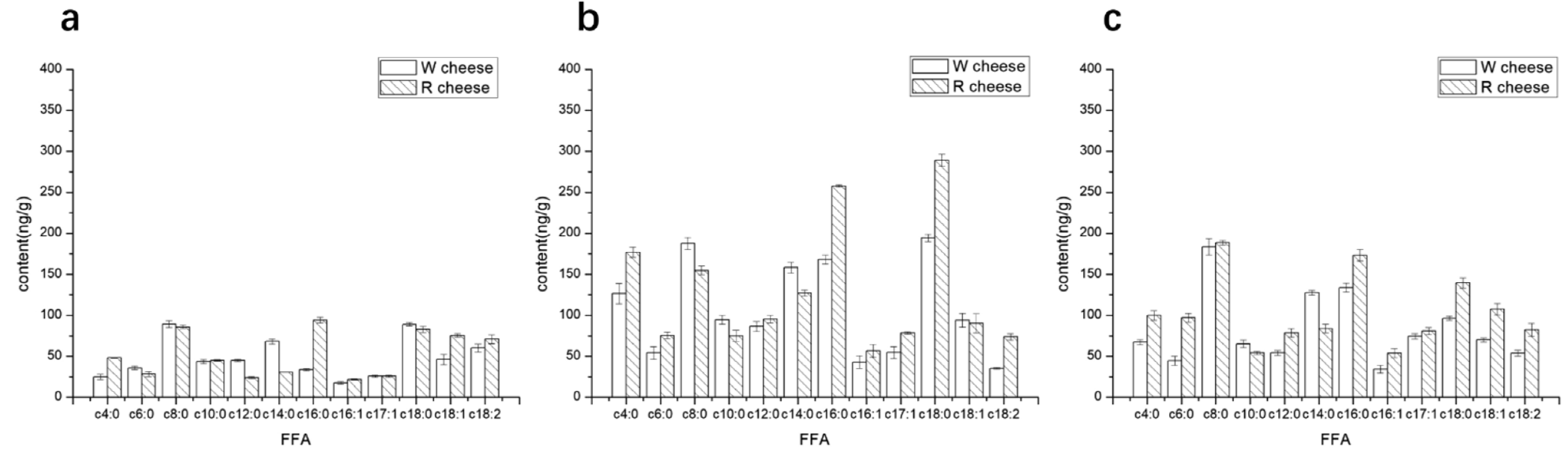
3.2. Effects of Monascus on Lipolysis and FFA of Camembert-Type Cheese
3.3. Effects of Monascus on Volatile Compounds of Camembert-Type Cheese
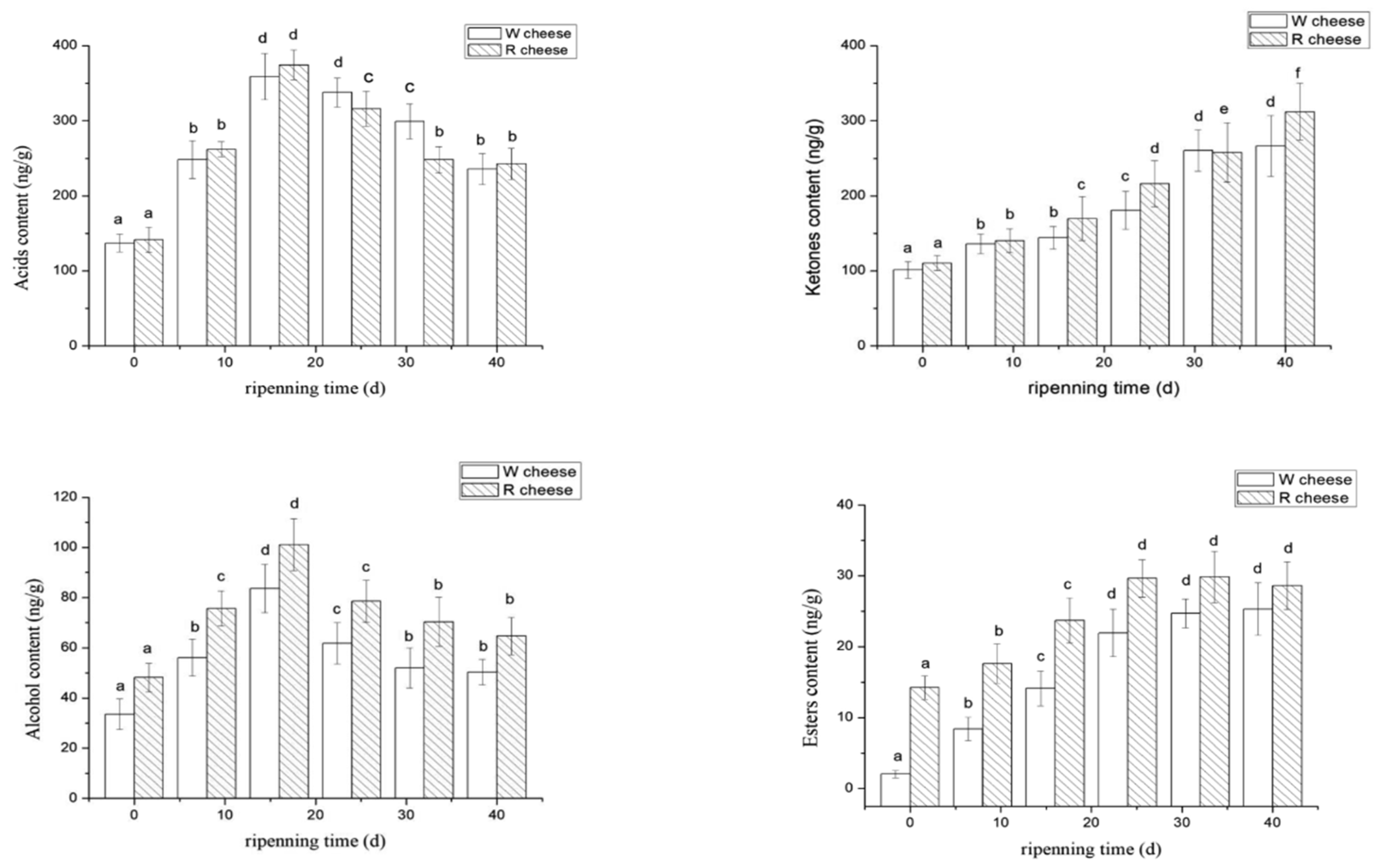
3.3.1. Acids
3.3.2. Ketones
3.3.3. Alcohols
3.3.4. Esters
3.3.5. Other Compounds
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leclercq, P.; Thiverval, G. Camembert, Brie, and related varieties. Encycl. Dairy Sci. Second Ed. 2011, 4, 773–782. [Google Scholar] [CrossRef]
- Lee, J.; Bae, I. Quality characteristics, changes in physiochemical properties and functional properties of Camembert cheese containing red ginseng powder. Korean J. Food Sci. An. 2018, 38, 64–77. [Google Scholar] [CrossRef]
- Chen, L.; Cui, J.; Ding, Q.; Ma, Y.; Chen, L.; Dong, J.Y. The effect of yeast species from raw milk in china on proteolysis and aroma compound formation in camembert-type cheese. Food Bioprocess Technol. 2012, 5, 2548–2556. [Google Scholar] [CrossRef]
- Yu, H.; Liu, Z.; Hang, F.; Mo, B. Effect of Monascus sp. as an adjunct starter on physicochemical properties and proteolysis in semi-hard cheeses during ripening. Food Sci. Biotechnol. 2016, 25, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Yuan, R.; Weng, S.; Wang, G.; Ai, L. Proteolysis, lipolysis, texture and sensory properties of cheese ripened by monascus fumeus. Food Res. Int. 2020, 137, 109657. [Google Scholar] [CrossRef] [PubMed]
- Patakova, P. Monascus, secondary metabolites: Production and biological activity. J. Ind. Microbiol. Biot. 2013, 40, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Srianta, I.; Ristiarini, S.; Nugerahani, I.; Sen, S.; Zhang, B.; Xu, G.; Blance, P. Recent research and development of monascus fermentation products. Int. Food Res. J. 2014, 21, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Katsiari, M.; Voutsinas, L. Lipolysis in reduced sodium Feta cheese made by partial substitution of NaCl by KCl. Int. Dairy J. 2000, 10, 369–373. [Google Scholar] [CrossRef]
- Chavarri, F.; Virto, M.; Martın, C.; Najera, A.; Santisteban, A.; Barron, L. Determination of free fatty acids in cheese: Comparison of two analytical methods. J. Dairy Res. 1997, 64, 445–452. [Google Scholar] [CrossRef]
- Li, L.; Wang, J. Comparative study of chemical composition and texture profile analysis between camembert cheese and Chinese Sufu. Biotechnol. Front. 2012, 1, 1–8. [Google Scholar]
- Martin, N.; Berger, C.; Le, D. Aroma compound production in cheese curd by coculturing with selected yeast and bacteria. J. Dairy Sci. 2001, 84, 2125–2135. [Google Scholar] [CrossRef]
- Gorostiza, A.; Cichoscki, A.; Valduga, A.; Valduga, E.; Bernardo, A.; Fresno, J. Changes in soluble nitrogenous compounds, caseins and free amino acids during ripening of artisanal prato cheese; a brazilian semi-hard cows variety. Food Chem. 2004, 85, 407–414. [Google Scholar] [CrossRef]
- Sablé, S.; Cottenceau, G. Current knowledge of soft cheeses flavor and related compounds. J. Agric. Food Chem. 1999, 47, 4825–4836. [Google Scholar] [CrossRef] [PubMed]
- Leclercq-Perlat, M.; Latrille, E.; Corrieu, G.; Spinnler, H. Controlled production of camembert-type cheeses. part II. Changes in the concentration of the more volatile compounds. J. Dairy Res. 2004, 71, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Buffa, M.; Guamis, B.; Pavia, M.; Trujillo, A. Lipolysis in cheese made from raw, pasteurized or high-pressure-treated goats’ milk. Int. Dairy J. 2001, 11, 175–179. [Google Scholar] [CrossRef]
- Kondyli, E.; Katsiari, M.; Masouras, T.; Voutsinas, L.P. Free fatty acids and volatile compounds of low-fat feta-type cheese made with a commercial adjunct culture. Food Chem. 2002, 79, 199–205. [Google Scholar] [CrossRef]
- Wu, S.; Yu, H.; Liu, Z.; You, C. Influence of Monascus purpureus BD-M-4 on the physicochemical properties, proteolysis and volatile compounds of surface mould-ripened cheese. Food Sci. Biotechnol. 2019, 28, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Zheng, Z.; Liu, Z.; You, C. Study of the compositional, microbiological, biochemical, and volatile profile of red-veined cheese, an internal monascus-ripened variety. Front. Nutr. 2021, 8, 649611. [Google Scholar] [CrossRef]
- Leclercqperlat, M.; Corrieu, G.; Spinnler, H. Controlled production of camembert-type cheeses: Part III role of the ripening microflora on free fatty acid concentrations. J. Dairy Res. 2007, 74, 218–225. [Google Scholar] [CrossRef]
- Karahadian, C.; Josephson, D.; Lindsay, R. Volatile compounds from Penicillium sp. Contributing musty-earthy notes to Brie and Camembert cheese flavors. J. Agric. Food Chem. 1985, 33, 339–343. [Google Scholar] [CrossRef]
- Salles, C.; Sommerer, N.; Septier, C.; Issanchou, S.; Chabanet, C.; Garem, A.; Quéré, J. Goat cheese flavor: Sensory evaluation of branched-chain fatty acids and small peptide. J. Food Sci. 2002, 67, 835–841. [Google Scholar] [CrossRef]
- Mei, J.; Guo, Q.; Wu, Y.; Li, Y.; Yu, H. Study of proteolysis, lipolysis, and volatile compounds of a camembert-type cheese manufactured using a freeze-dried tibetan kefir co-culture during ripening. Food Sci. Biotechnol. 2015, 24, 393–402. [Google Scholar] [CrossRef]
- Chen, S.; Wang, J.; Van Kessel, J.; Ren, F.; Zeng, S. Effect of somatic cell count in goat milk on yield, sensory quality, and fatty acid profile of semisoft cheese. J. Dairy Sci. 2010, 93, 1345–1354. [Google Scholar] [CrossRef] [Green Version]
- Singh, T.; Drake, M.; Cadwallader, K. Flavor of cheddar cheese: A chemical and sensory perspective. Compr. Rev. Food Sci. Food Saf. 2010, 2, 166–189. [Google Scholar] [CrossRef]
- Carunchiawhetstine, M.; Karagul-Yuceer, Y.; Avsar, Y.; Avsar, Y.; Drake, M. Identification and quantification of character aroma components in fresh Chevre-style goat cheese. J. Food Sci. 2003, 68, 2441–2447. [Google Scholar] [CrossRef]
- Hong, Q.; Wang, G.; Zhang, Q.; Tian, F.; Liu, X.; Zhao, J.; Chen, Y.; Zhang, H.; Chen, W. Enhancement of ester formation in Camembert cheese by addition of ethanol. Int. J. Dairy Technol. 2017, 70, 220–227. [Google Scholar] [CrossRef]
- Kubícková, J.; Grosch, W. Evaluation of flavour compounds of Camembert cheese. Int. Dairy J. 1998, 8, 11–16. [Google Scholar] [CrossRef]
- Malcata, F.; Reyes, H.; Garcia, H.; Hill, C.; Amundson, C. Kinetics and mechanisms of reactions catalysed by immobilized lipases. Enzyme Microb. Tech. 1992, 14, 426–446. [Google Scholar] [CrossRef]
- Varming, C.; Andersen, L.; Petersen, M.; Ardö, Y. Flavour compounds and sensory characteristics of cheese powders made from matured cheeses. Int. Dairy J. 2013, 30, 19–28. [Google Scholar] [CrossRef]
- Delgado, F.; González-Crespo, J.; Cava, R. Changes in the volatile profile of a raw goat milk cheese treated by hydrostatic high pressure at different stages of maturation. Int. Dairy J. 2011, 21, 135–141. [Google Scholar] [CrossRef]
- Sherveglieri, V.; Bhandari, M.P.; Carmona, E.N.; Betto, G.; Soprani, M.; Malla, R.; Sberveglieri, G. Spectrocolorimetry and nanowire gas sensor device S3 for the analysis of Parmigiano Reggiano cheese ripening. In Proceedings of the 2017 ISOCS/IEEE International Symposium on Olfaction and Electronic Nose (ISOEN), Montreal, QC, Canada, 28–31 May 2017; pp. 1–3. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RI | Compounds | Cheese | Ripening Time (d) | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 8 | 16 | 24 | 32 | 40 | |||
| Acids | ||||||||
| 594 | acetic acid | W | 2.82 ± 0.37 Aa | 7.42 ± 0.68 Ab | 16.27 ± 1.25 Ac | 43.15 ± 5.51 Ae | 34.18 ± 3.16 Ad | 23.68 ± 3.22 Ac |
| R | 0.73 ± 0.35 Ba | 1.49 ± 0.18 Bb | 13.96 ± 0.56 Bc | 26.92 ± 1.35 Be | 19.06 ± 0.18 Bd | 13.37 ± 1.65 Bc | ||
| 661 | propionic acid | W | 2.27 ± 0.12 a | 13.70 ± 0.96 Ac | 25.36 ± 0.82 Ab | 20.79 ± 1.48 Ab | 17.81 ± 1.35 Ab | 16.59 ± 2.29 Ab |
| R | ND | 0.66 ± 0.04 Ba | 1.29 ± 0.73 Bb | 5.74 ± 0.56 Bc | 9.06 ± 1.72 Bd | 10.57 ± 2.17 Bd | ||
| 775 | Butyric acid | W | 14.31 ± 1.23 Ab | 22.08 ± 0.26 Bc | 21.40 ± 4.26 Bc | 15.25 ± 6.35 Bb | 22.90 ± 2.53 Ac | 8.68 ± 0.20 Ba |
| R | 11.27 ± 3.47 Ba | 52.69 ± 1.16 Ad | 63.38 ± 1.99 Ae | 38.86 ± 7.89 Ac | 20.51 ± 2.23 Ab | 21.59 ± 3.19 Ab | ||
| 861 | 2-Methyl butanoic | W | 24.81 ± 2.61 Aa | 35.48 ± 1.65 Bb | 47.08 ± 17.73 Bd | 40.74 ± 1.73 Ac | 34.64 ± 0.44 Ab | 33.32 ± 0.47 Bb |
| R | 23.90 ± 0.51 Aa | 58.79 ± 2.39 Ac | 56.42 ± 1.27 Ae | 42.98 ± 0.25 Ad | 34.83 ± 1.15 Ac | 38.29 ± 0.93 Ab | ||
| 877 | 3-Methyl pentanoic | W | 3.55 ± 0.45 Aa | 7.20 ± 0.97 Bb | 22.56 ± 3.07 Bd | 29.43 ± 1.89 Be | 12.66 ± 0.65 Bc | 10.28 ± 2.18 Bc |
| R | 5.46 ± 0.24 Aa | 18.68 ± 1.04 Ab | 54.22 ± 3.43 Ad | 72.45 ± 2.76 Ae | 51.36 ± 3.68 Ac | 55.46 ± 4.89 Ac | ||
| 974 | Hexanoic acid | W | 41.03 ± 5.59 Aa | 69.03 ± 6.06 Aa | 73.39 ± 4.26 Bc | 83.76 ± 17.58 Ad | 63.92 ± 5.53 Bb | 58.76 ± 11.86 Bbc |
| R | 2.76 ± 0.18 Ba | 13.23 ± 4.91 Ba | 90.30 ± 23.86 Ad | 75.86 ± 4.52 Bc | 73.01 ± 3.59 Ab | 70.74 ± 1.88 Ab | ||
| 1017 | Heptanoic acid | W | 4.89 ± 0.96 Ac | 3.42 ± 0.54 Ab | 3.02 ± 0.19 Bab | ND | 7.25 ± 1.02 Ad | 1.98 ± 0.52 Ba |
| R | 0.68 ± 0.18 Ba | 2.37 ± 0.53 Ab | 11.35 ± 2.79 Ad | 2.84 ± 0.39 b | 6.51 ± 0.79 Ac | 10.01 ± 0.05 Ad | ||
| 1272 | Nonanoic acid | W | 1.82 ± 0.03 a | 4.39 ± 0.25 c | ND | ND | 3.16 ± 0.75 Ab | 2.47 ± 0.73 Bab |
| R | ND | ND | 0.37 ± 0.16 a | 1.06 ± 0.09 c | 2.93 ± 0.26 Ac | 5.79 ± 0.68 Ad | ||
| 1372 | Decanoic acid | W | 4.62 ± 0.33 Ba | 18.59 ± 5.74 Ab | 25.13 ± 2.36 Ac | 28.04 ± 3.74 Ac | 25.40 ± 1.60 Ac | 20.93 ± 0.35 Ab |
| R | 8.88 ± 0.72 Aa | 11.78 ± 0.59 Bb | 18.28 ± 0.52 Ba | 19.26 ± 0.65 Ba | 17.30 ± 1.05 Ba | 11.83 ± 2.48 Bb | ||
| 1173 | Octanoic acid | W | 32.15 ± 1.57 Aa | 56.31 ± 1.82 Ab | 91.57 ± 4.33 Ac | 75.13 ± 15.29 Ac | 65.43 ± 3.39 Ab | 56.86 ± 3.48 Ab |
| R | 17.29 ± 1.98 Ba | 27.39 ± 3.08 Bbc | 43.76 ± 9.56 Bf | 37.24 ± 1.86 Be | 33.08 ± 15.52 Bcd | 24.72 ± 2.20 Bab | ||
| 1370 | Dodecanoic acid | W | 4.72 ± 0.24 Ad | ND | 0.60 ± 0.12 Aa | 1.43 ± 0.73 Ac | 1.91 ± 0.34 Bc | 2.11 ± 0.28 Bbc |
| R | 0.61 ± 0.06 Ba | ND | 1.19 ± 0.16 Aa | 2.84 ± 0.22 Ab | 7.62 ± 1.49 Ac | 12.56 ± 1.56 Ad | ||
| 1249 | Benzeneacetic acid | W | ND | 0.80 ± 0.19 a | 2.59 ± 0.19 b | ND | ND | ND |
| R | ND | ND | ND | ND | ND | ND | ||
| 1366 | Undecanoic acid | W | ND | ND | ND | ND | ND | ND |
| R | ND | ND | ND | ND | 2.94 ± 1.05 a | 7.62 ± 0.18 b | ||
| Ketones | ||||||||
| 594 | 2-butanone | W | 10.80 ± 1.75 Bb | 23.44 ± 2.06 B c | 6.76 ± 0.95 Ba | ND | ND | ND |
| R | 32.52 ± 1.04 Ac | 34.37 ± 3.21 Ac | 20.24 ± 1.77 Ab | 5.96 ± 0.35 a | ND | ND | ||
| 694 | 2-Pentanone | W | 5.09 ± 0.35 Bb | 22.96 ± 0.55 Aa | 16.50 ± 1.23 Ac | ND | ND | ND |
| R | 9.75 ± 0.55 Ab | 20.36 ± 1.93 Ad | 14.05 ± 0.58 Ac | 4.86 ± 0.73 a | ND | ND | ||
| 853 | 2-Hexanone | W | 17.08 ± 3.79 Aa | 29.58 ± 3.29 Aa | 38.43 ± 5.69 Ba | 50.04 ± 1.10 Aa | 80.11 ± 15.67 Ab | 83.04 ± 7.54 Ab |
| R | 8.49 ± 0.29 Ba | 12.38 ± 1.92 Ba | 45.88 ± 9.56 Aa | 49.52 ± 3.55 Aa | 77.03 ± 45.82 Bb | 82.62 ± 44.27 Aa | ||
| 952 | 2-Octanone | W | ND | ND | ND | 7.13 ± 0.17 Aa | 9.81 ± 3.45 Aa | 15.16 ± 6.13 Ab |
| R | ND | ND | ND | 2.74 ± 1.81 Ba | 5.86 ± 0.67 Bb | 5.66 ± 1.04 Bb | ||
| 1052 | 2-Nonanone | W | 58.89 ± 8.14 Aa | 49.02 ± 1.73 Ba | 64.38 ± 13.59 Ab | 69.90 ± 6.70 Bb | 99.30 ± 3.35 Ac | 81.33 ± 9.44 Ac |
| R | 50.64 ± 3.39 Ba | 55.84 ± 3.17 Aa | 48.47 ± 19.62 Ba | 78.47 ± 8.59 Ab | 87.21 ± 13.78 Bbc | 75.08 ± 28.39 Ba | ||
| 1042 | 8-Nonen-2-one | W | ND | ND | ND | 29.93 ± 1.34 Ba | 37.84 ± 4.57 Ab | 41.67 ± 5.70 Bc |
| R | ND | 5.39 ± 0.89 a | 22.96 ± 1.18 d | 35.26 ± 1.74 Ab | 38.08 ± 2.46 Ac | 46.78 ± 0.62 Ac | ||
| 1151 | 2-Decanone | W | ND | ND | ND | 4.63 ± 0.66 Ab | 3.93 ± 1.06 Aa | 7.15 ± 1.32 Ac |
| R | ND | ND | ND | 1.08 ± 0.32 Ba | 1.93 ± 0.26 Ba | 2.97 ± 0.61 Bb | ||
| 1159 | 7-Decen-2-one | W | ND | ND | ND | 1.94 ± 0.70 Aa | 4.82 ± 1.19 Ab | 9.38 ± 2.25 Ac |
| R | ND | ND | ND | 2.33 ± 0.73 Ab | 1.56 ± 0.39 Ba | 3.80 ± 0.23 Bc | ||
| 1251 | 2-Undecanone | W | 9.52 ± 2.31 Ab | 10.98 ± 0.35 Aa | 18.13 ± 0.96 Ab | 17.02 ± 1.14 Bb | 24.71 ± 2.28 Bb | 28.63 ± 2.05 Bc |
| R | 9.02 ± 0.59 Aa | 11.84 ± 1.33 Ab | 17.98 ± 2.05 Ac | 29.28 ± 1.29 Ac | 32.15 ± 2.05 Ac | 56.45 ± 2.99 Ad | ||
| 1370 | 2-Tridecanone | W | ND | ND | ND | ND | ND | ND |
| R | ND | ND | ND | 5.79 ± 0.29 a | 12.26 ± 0.95 b | 35.67 ± 0.77 c | ||
| Alcohols | ||||||||
| 663 | Ethanol | W | 3.63 ± 0.42 b | 1.55 ± 0.17 a | ND | ND | ND | ND |
| R | ND | ND | ND | ND | ND | ND | ||
| 743 | 2,3-Butanediol | W | 13.4 ± 0.84 Ab | 18.26 ± 1.01 Bc | 6.76 ± 0.75 Ba | 5.35 ± 0.17 Aa | ND | ND |
| R | 11.69 ± 1.89 Ab | 21.97 ± 1.28 Ac | 24.32 ± 2.97 Ac | 4.69 ± 0.75 Aa | 6.12 ± 0.64 a | ND | ||
| 697 | 3-methyl-1-Butanol | W | 3.19 ± 0.92 ab | 14.09 ± 3.38 c | 31.59 ± 3.07 d | 14.58 ± 1.58 Ac | 4.61 ± 1.06 Ab | ND |
| R | ND | ND | ND | 5.86 ± 0.27 Ba | 6.34 ± 0.04 Ab | 7.25 ± 1.28 b | ||
| 681 | Pentanol | W | ND | ND | ND | ND | ND | ND |
| R | 1.80 ± 0.25 a | 4.71 ± 0.83 b | 19.49 ± 1.46 d | 10.42 ± 2.37 c | 6.72 ± 1.33 b | 3.93 ± 0.56 b | ||
| 815 | 5-methyl- 2-Hexanol | W | ND | 4.60 ± 1.51 Aa | 14.98 ± 0.12 b | ND | ND | ND |
| R | ND | 1.97 ± 0.56 Ba | ND | ND | ND | ND | ||
| 960 | 2-Heptanol | W | ND | ND | ND | ND | 8.41 ± 0.98 Ba | 11.61 ± 1.43 Ab |
| R | 2.32 ± 0.75 a | 3.89 ± 0.69 a | 11.14 ± 1.52 b | 16.23 ± 2.93 c | 26.55 ± 3.02 Ad | 8.46 ± 1.19 Bb | ||
| 979 | 2-Octanol | W | 1.36 ± 0.59 a | 4.31 ± 0.32 b | 3.85 ± 0.62 b | 1.78 ± 0.18 a | ND | ND |
| R | ND | ND | ND | ND | 4.71 ± 0.29 b | 3.26 ± 1.06 a | ||
| 969 | 1-Octen-3-ol | W | 5.77 ± 0.25 a | 4.19 ± 1.10 Ba | 17.79 ± 2.75 Ad | 19.55 ± 1.87 Ae | 25.67 ± 1.56 Aa | 20.62 ± 0.87 Aa |
| R | ND | 7.57 ± 0.48 Aa | 10.06 ± 1.42 Bb | 15.29 ± 6.41 Bc | 13.62 ± 2.42 Bd | 15.73 ± 1.73 Bc | ||
| 1078 | 2-Nonanol | W | ND | ND | 3.24 ± 0.35 Ba | 6.85 ± 0.25 Ab | 10.44 ± 0.65 Ac | 17.02 ± 1.87 Ad |
| R | 16.77 ± 3.15 b | 9.34 ± 0.34 a | 14.75 ± 1.02 Ab | 7.64 ± 0.26 Aa | 5.36 ± 0.74 Aa | 7.16 ± 1.21 Ba | ||
| 1136 | Phenylethyl Alcohol | W | 3.76 ± 0.59 Bc | 7.96 ± 0.87 Be | 5.44 ± 0.17 Bd | 3.66 ± 0.44 Bc | 2.87 ± 0.06 Bb | 1.08 ± 0.01 Ba |
| R | 11.02 ± 1.48 Aa | 24.34 ± 2.57 Ac | 21.32 ± 1.58 Ac | 18.43 ± 6.31 Abc | 17.86 ± 1.06 Ab | 18.89 ± 2.33 Ab | ||
| 1178 | Decanol | W | 2.43 ± 0.21 Bb | 1.16 ± 0.63 Aa | ND | ND | ND | ND |
| R | 4.55 ± 0.85 Ab | 1.91 ± 0.09 Aa | ND | ND | ND | ND | ||
| Esters | ||||||||
| 1386 | delta-Decanolide | W | 2.05 ± 0.10 Aa | 2.17 ± 0.04 Ab | 1.96 ± 0.56 Aa | 1.79 ± 0.15 Bab | 2.48 ± 0.49 Bb | 1.87 ± 0.23 Ba |
| R | 2.42 ± 0.39 Aa | 2.78 ± 0.74 Abc | 2.46 ± 0.28 Ab | 3.62 ± 1.02 Ac | 4.63 ± 0.74 Ad | 4.16 ± 0.61 Ad | ||
| 1602 | delta-Dodecalactone | W | ND | 1.68 ± 0.27 Ab | 2.39 ± 0.75 Ac | 2.44 ± 0.18 Ac | 1.82 ± 0.11 Bb | 1.18 ± 0.03 Ba |
| R | ND | 1.37 ± 0.79 Aa | 2.01 ± 0.54 Ab | 2.77 ± 0.07 Ab | 3.94 ± 0.76 Ac | 3.76 ± 0.34 Ac | ||
| 984 | Ethyl hexanoate | W | ND | 1.75 ± 0.02 Bb | 1.95 ± 0.54 Bb | 2.67 ± 0.67 Bc | 1.34 ± 0.23 Ab | 0.98 ± 0.47 Ba |
| R | 5.69 ± 1.08 b | 4.91 ± 0.39 Aab | 8.66 ± 0.57 Ac | 10.66 ± 3.15 Ad | 7.30 ± 1.75 Bc | 7.67 ± 0.98 Ac | ||
| 796 | Ethyl butyrate | W | ND | 0.78 ± 0.06 ab | 2.07 ± 0.74 bc | 7.68 ± 2.35 d | 9.04 ± 0.32 d | 6.82 ± 0.94 c |
| R | ND | ND | ND | ND | ND | ND | ||
| 1183 | Ethyl caprylate | W | ND | 1.36 ± 0.43 Bb | 1.87 ± 0.18 Bc | 2.14 ± 0.49 Bc | 0.75 ± 0.27 Ba | ND |
| R | 2.04 ± 0.34 a | 3.62 ± 0.45 Ab | 4.85 ± 0.70 Ab | 3.88 ± 0.66 Ab | 6.72 ± 0.15 Ad | 5.41 ± 0.88 c | ||
| 963 | Methyl pentanoic | W | ND | 0.67 ± 0.16 a | 2.42 ± 1.33 b | 2.86 ± 0.14 b | 6.61 ± 0.17 c | 8.59 ± 0.93 d |
| R | 1.63 ± 0.46 b | 1.06 ± 0.03 a | ND | ND | ND | ND | ||
| 1381 | Ethyl decanoate | W | ND | ND | 1.46 ± 0.11 Bb | 0.84 ± 0.27 Ba | ND | ND |
| R | 0.61 ± 0.06 a | 3.18 ± 0.79 b | 4.64 ± 0.15 Ac | 7.83 ± 0.31 Ad | 7.25 ± 0.85 d | 7.62 ± 1.09 d | ||
| 1083 | Heptyl acetate | W | ND | ND | ND | 1.52 ± 2.24 a | 2.65 ± 0.48 b | 5.88 ± 0.36 c |
| R | ND | ND | ND | ND | ND | ND | ||
| 981 | Hexyl formate | W | ND | ND | ND | ND | ND | ND |
| R | ND | ND | 1.09 ± 0.16 b | 0.89 ± 0.07 a | ND | ND | ||
| 1081 | Methyl pentanoic | W | ND | ND | ND | ND | ND | ND |
| R | 1.83 ± 0.28 b | 0.69 ± 0.14 a | ND | ND | ND | ND | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Wang, T.; Zhang, Y.; Song, B.; Pang, X.; Lv, J. Effects of Monascus on Proteolysis, Lipolysis, and Volatile Compounds of Camembert-Type Cheese during Ripening. Foods 2022, 11, 1662. https://doi.org/10.3390/foods11111662
Zhang S, Wang T, Zhang Y, Song B, Pang X, Lv J. Effects of Monascus on Proteolysis, Lipolysis, and Volatile Compounds of Camembert-Type Cheese during Ripening. Foods. 2022; 11(11):1662. https://doi.org/10.3390/foods11111662
Chicago/Turabian StyleZhang, Shuwen, Tong Wang, Yumeng Zhang, Bo Song, Xiaoyang Pang, and Jiaping Lv. 2022. "Effects of Monascus on Proteolysis, Lipolysis, and Volatile Compounds of Camembert-Type Cheese during Ripening" Foods 11, no. 11: 1662. https://doi.org/10.3390/foods11111662