
Dietary Curcumin Alleviated Acute Ileum Damage of Ducks (Anas platyrhynchos) Induced by AFB1 through Regulating Nrf2-ARE and NF-κB Signaling Pathways


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Ducks and Husbandry
2.3. Sample Collection
2.4. Assay of Antioxidant Levels in the Plasma
2.5. Assay of AFB1-DNA Adducts Levels in the Plasma
2.6. Assay of Antioxidant Ability in Ileum
2.7. RNA Isolation and Real-Time Quantitative Polymerase Chain Reaction (qRT-PCR)
2.8. Western Blotting
2.9. Statistical Analysis
3. Results
3.1. Intestinal Morphology
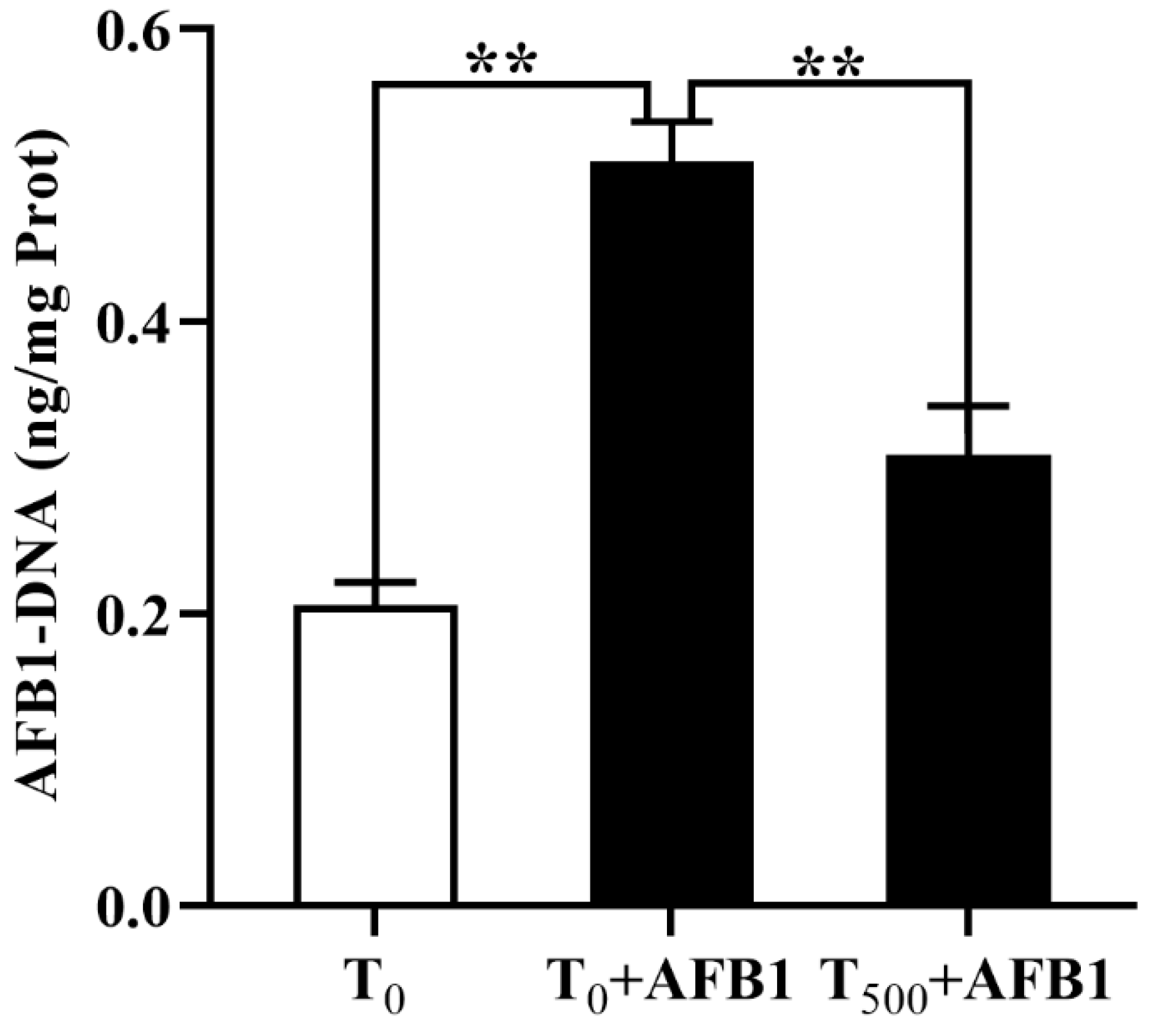
3.2. Levels of AFB1-DNA Adducts in the Plasma
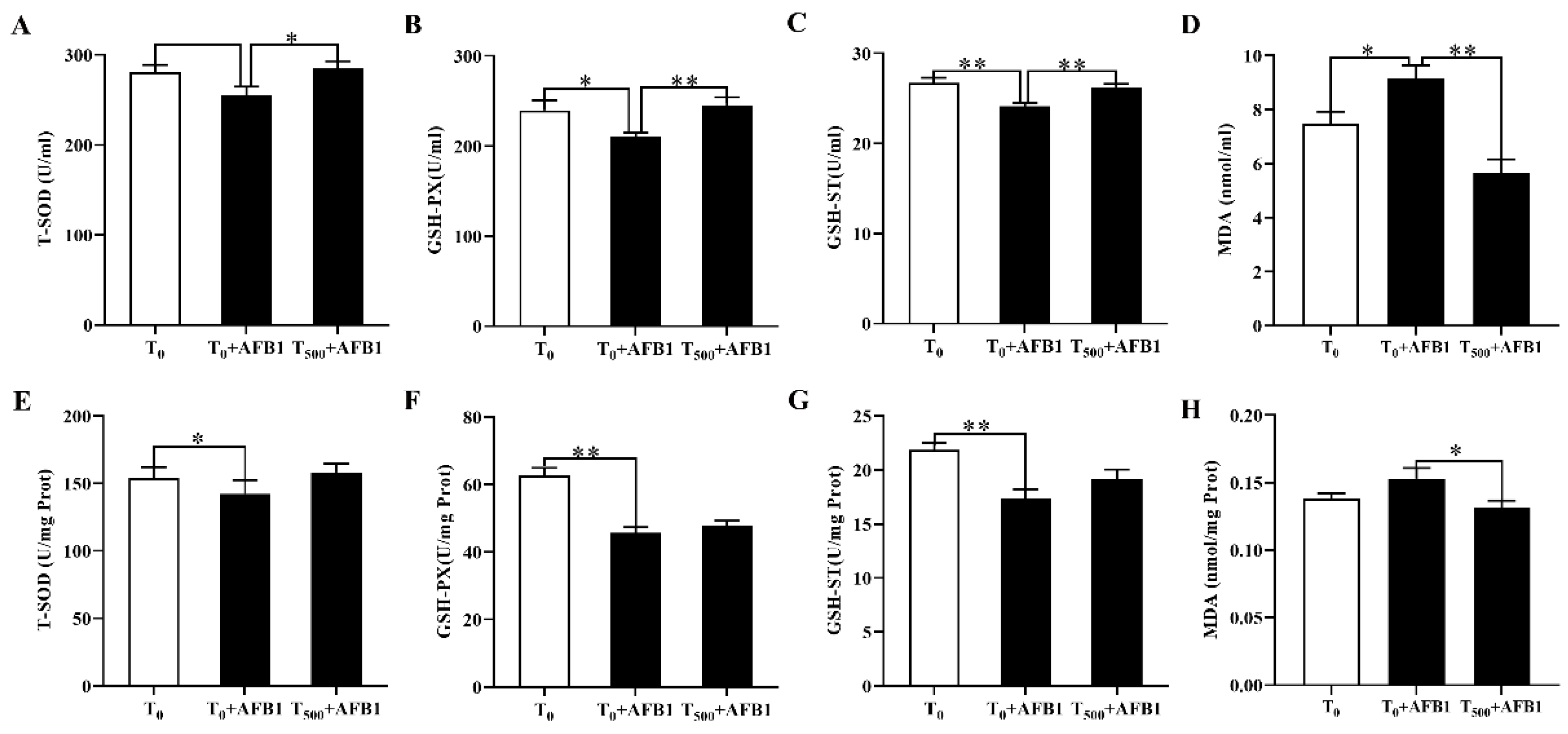
3.3. Antioxidant Capacity in the Plasma and Ileum
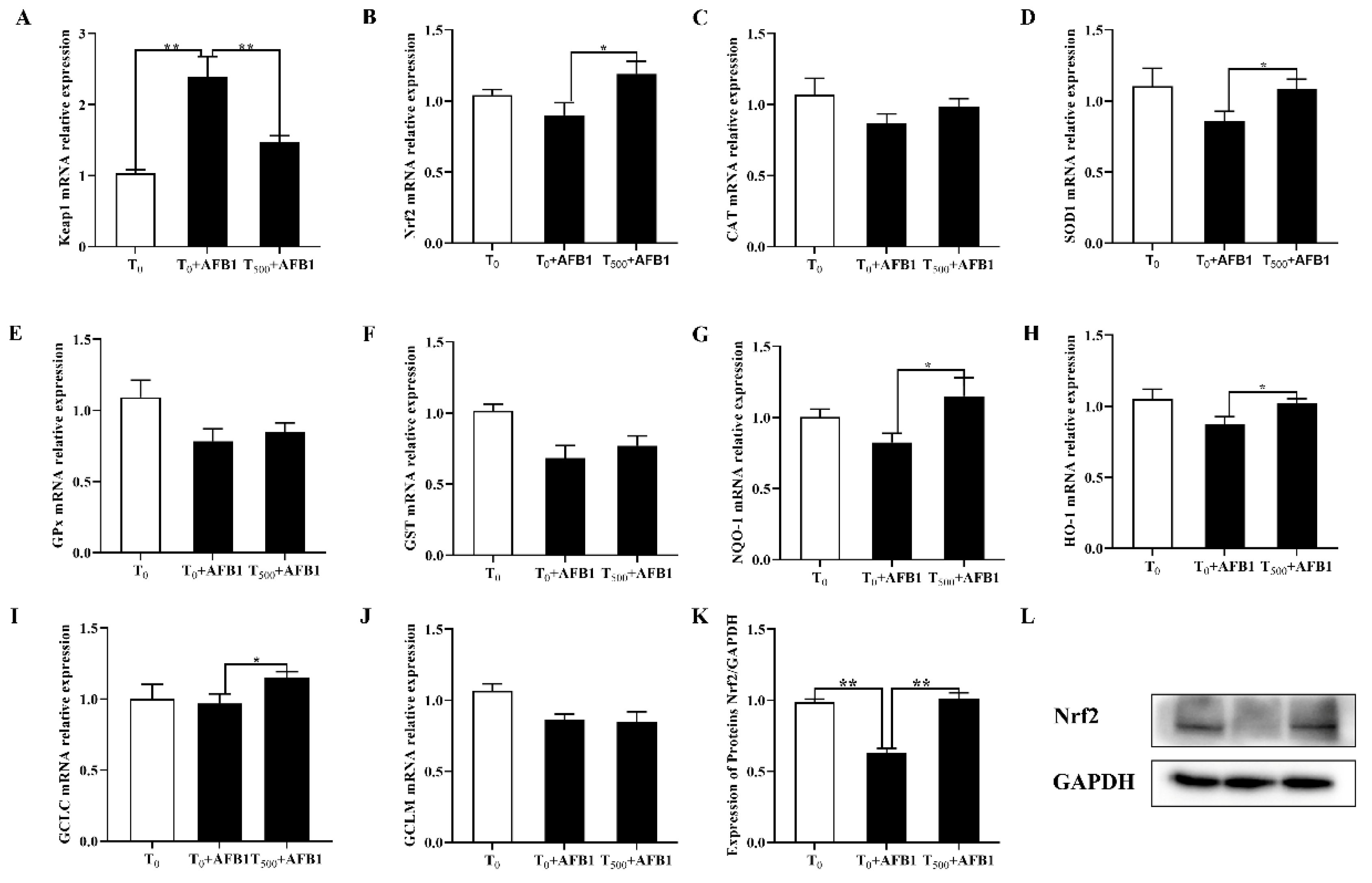
3.4. Expression of Genes Related to Nrf2-ARE Signaling Pathway in the Ileum
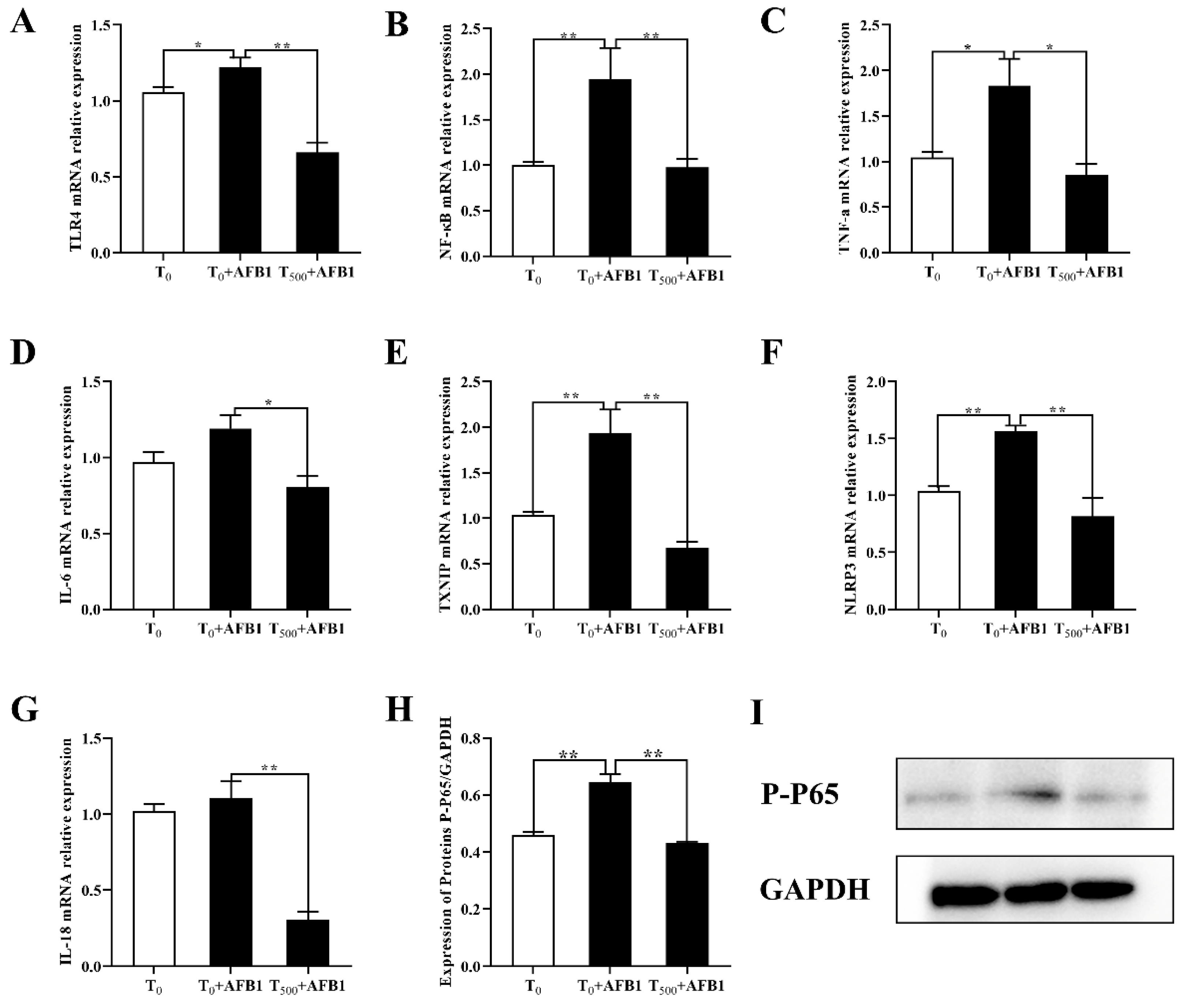
3.5. Expression of Genes Related to NF-κB Signaling Pathway in the Ileum
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khan, M.A.; Ali, S.; Yang, H.; Kamboh, A.A.; Ahmad, Z.; Tume, R.K.; Zhou, G. Improvement of color, texture and food safety of ready-to-eat high pressure-heat treated duck breast. Food Chem. 2019, 277, 646–654. [Google Scholar] [CrossRef]
- Kos, J.; Janic Hajnal, E.; Malachova, A.; Steiner, D.; Stranska, M.; Krska, R.; Poschmaier, B.; Sulyok, M. Mycotoxins in maize harvested in republic of serbia in the period 2012–2015. Part 1: Regulated mycotoxins and its derivatives. Food Chem. 2020, 312, 126034. [Google Scholar] [CrossRef]
- Wangia, R.N.; Tang, L.; Wang, J.S. Occupational exposure to aflatoxins and health outcomes: A review. J. Environ. Sci. Health Part C Environ. Carcinog. Rev. 2019, 37, 215–234. [Google Scholar] [CrossRef]
- Vineis, W.; Xun, P.V. The emerging epidemic of environmental cancers in developing countries. Ann. Oncol. 2009, 20, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Robert, H.; Payros, D.; Pinton, P.; Théodorou, V.; Mercier-Bonin, M.; Oswald, I.P. Impact of mycotoxins on the intestine: Are mucus and microbiota new targets? J. Toxicol. Environ. Health Part B Crit. Rev. 2017, 20, 249. [Google Scholar] [CrossRef] [PubMed]
- Ishfaq, M.; He, W.; Sun, X.; Wang, X.; Han, M.; Lu, Z. Dual role of dietary curcumin through attenuating afb1-induced oxidative stress and liver injury via modulating liver phase-i and phase-ii enzymes involved in afb1 bioactivation and detoxification. Front. Pharmacol. 2018, 9, 554. [Google Scholar]
- Jager, A.V.; Tonin, F.G.; Baptista, G.Z.; Souto, P.C.; Oliveira, C.A. Assessment of aflatoxin exposure using serum and urinary biomarkers in sao paulo, brazil: A pilot study. Int. J. Hyg. Environ. Health 2016, 219, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Muhammad, I.; Yu, H.; Sun, X.; Zhang, X. Detection of aflatoxin adducts as potential markers and the role of curcumin in alleviating AFB1-induced liver damage in chickens. Ecotoxicol. Environ. Saf. 2019, 176, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Wijtten, P.J.; van der Meulen, J.; Verstegen, M.W. Intestinal barrier function and absorption in pigs after weaning: A review. Br. J. Nutr. 2011, 105, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Liu, W.; Li, Q.; Li Y., F.; Yan, Y.L.; Huang, F.; Wu, X.; Zhou, Q.C.; Shu, X.G.; Ruan, Z. Dietary chlorogenic acid regulates gut microbiota, serum-free amino acids and colonic serotonin levels in growing pigs. Int. J. Food Sci. Nutr. 2017, 1–8. [Google Scholar] [CrossRef]
- Wang, F.; Zuo, Z.; Chen, K.; Gao, C.; Yang, Z.; Zhao, S.; Li, J.; Song, H.; Xi, P.; Jing, F. Histopathological Injuries, Ultrastructural Changes, and Depressed TLR Expression in the Small Intestine of Broiler Chickens with Aflatoxin B1. Toxins 2018, 10, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Peng, X.; Fang, J.; Cui, H.; Zuo, Z.; Chen, Z. Effects of aflatoxin B1 exposure and sodium selenite supplementation on the histology, cell proliferation, and cell cycle of jejunum in broilers. Biol. Trace Elem. Res. 2014, 160, 32. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Balachandran, C. Histopathological changes in broiler chickens fed aflatoxin and cyclopiazonic acid. Vet. Arh. 2009, 79, 31–40. [Google Scholar]
- Pandey, A.; Gupta, R.K.; Srivastava, R. Curcumin-the yellow magic. Asian J. Appl. Sci. 2011, 4, 343–354. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, M.; Shan, A.; Feng, X. Avian host defense cathelicidins: Structure, expression, biological functions, and potential therapeutic applications. Poult. Sci. 2020, 99, 6434–6445. [Google Scholar] [CrossRef]
- Menon, V.P.; Sudheer, A.R. Antioxidant and anti-inflammatory properties of curcumin. Oxyg. Transp. Tissue XXXIII 2007, 595, 105–125. [Google Scholar]
- He, Y.; Yue, Y.; Zheng, X.; Zhang, K.; Chen, S.; Du, Z. Curcumin, inflammation, and chronic diseases: How are they linked? Molecules 2015, 20, 26007179. [Google Scholar] [CrossRef]
- Soliman, M.; Nassan, M.; Ismail, T. Immunohistochemical and molecular study on the protective effect of curcumin against hepatic toxicity induced by paracetamol in Wistar rats. BMC Complement. Altern. Med. 2014, 14, 457. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, K.; Dobiasch, S.; Nguyen, L.; Schilling, D.; Combs, S.E. Modification of radiosensitivity by Curcumin in human pancreatic cancer cell lines. Sci. Rep. 2020, 10, 3815. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Harikumar, K.B. Potential Therapeutic Effects of Curcumin, the Anti-Inflammatory Agent, Against Neurodegenerative, Cardiovascular, Pulmonary, Metabolic, Autoimmune and Neoplastic Diseases. Int. J. Biochem. Cell Biol. 2009, 41, 40–59. [Google Scholar] [CrossRef] [Green Version]
- Abrahams, S.; Haylett, W.L.; Johnson, G.; Carr, J.A.; Bardien, S. Antioxidant effects of curcumin in models of neurodegeneration, aging, oxidative and nitrosative stress: A review. Neuroscience 2019, 406, 1–21. [Google Scholar] [CrossRef]
- Nakmareong, S.; Kukongviriyapan, U.; Pakdeechote, P.; Donpunha, W.; Kukongviriyapan, V.; Kongyingyoes, B.; Sompamit, K.; Phisalaphong, C. Antioxidant and vascular protective effects of curcumin and tetrahydrocurcumin in rats with L-NAME-induced hypertension. Naunyn Schmiede. Arch. Pharm. 2011, 383, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Topcu-Tarladacalisir, Y.; Akpolat, M.; Uz, Y.H.; Kizilay, G.; Sapmaz-Metin, M.; Cerkezkayabekir, A.; Omurlu, I.K. Effects of curcumin on apoptosis and oxidoinflammatory regulation in a rat model of acetic acid-induced colitis: The roles of c-Jun N-terminal kinase and p38 mitogen-activated protein kinase. J. Med. Food 2013, 16, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, A.H.; Alsahli, M.; Aly, S.M.; Khan, M.A.; Aldebasi, Y.H. Role of Curcumin in Disease Prevention and Treatment. Adv. Biomed. Res. 2018, 7, 38. [Google Scholar] [CrossRef] [PubMed]
- Limaye, A.; Yu, R.C.; Chou, C.C.; Liu, J.R.; Cheng, K.C. Protective and Detoxifying Effects Conferred by Dietary Selenium and Curcumin against AFB1-Mediated Toxicity in Livestock: A Review. Toxins 2018, 10, 25. [Google Scholar] [CrossRef] [Green Version]
- Soetikno, V.; Sari, F.R.; Lakshmanan, A.P.; Arumugam, S.; Harima, M.; Suzuki, K.; Kawachi, H.; Watanabe, K. Curcumin alleviates oxidative stress, inflammation, and renal fibrosis in remnant kidney through the Nrf2-keap1 pathway. Mol. Nutr. Food Res. 2013, 57, 1649–1659. [Google Scholar] [CrossRef]
- Wang, D.; Huang, H.; Zhou, L.; Li, W.; Zhou, H.; Hou, G.; Liu, J.; Hu, L. Effects of dietary supplementation with turmeric rhizome extract on growth performance, carcass characteristics, antioxidant capability, and meat quality of wenchang broiler chickens. Ital. J. Anim. Sci. 2016, 14, 344–349. [Google Scholar] [CrossRef]
- Jin, S.; Pang, Q.; Liu, R.; Yang, H.; Liu, F.; Wang, M.; Wang, Y.; Feng, X.; Shan, A. Dietary curcumin decreased lipid oxidation and enhanced the myofibrillar protein structure of the duck (Anas platyrhynchos) breast muscle when subjected to storage. LWT-Food Sci. Technol. 2020, 133, 1–7. [Google Scholar] [CrossRef]
- Yan, H.; Ge, J.; Gao, H.; Pan, Y.; Hao, Y.; Li, J. Melatonin attenuates AFB1-induced cardiotoxicity via the NLRP3 signalling pathway. J. Med. Internet Res. 2020, 48, 300060520952656. [Google Scholar] [CrossRef]
- Luzi, A.; Cometa, M.F.; Palmery, M. Acute effects of aflatoxins on guinea pig isolated ileum. Toxicol. Vitr. Int. J. Publ. Assoc. Bibra 2002, 16, 525–529. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, J.; Zhou, M.; Li, M.; Li, M.; Tan, H. Curcumin attenuates murine lupus via inhibiting NLRP3 inflammasome. Int. Immunopharmacol. 2019, 69, 213–216. [Google Scholar] [CrossRef]
- Gong, Z.; Zhou, J.; Li, H.; Gao, Y.; Xu, C.; Zhao, S.; Chen, Y.; Cai, W.; Wu, J. Curcumin suppresses NLRP3 inflammasome activation and protects against LPS-induced septic shock. Mol. Nutr. Food Res. 2015, 59, 2132–2142. [Google Scholar] [CrossRef]
- Ruan, Z.; Mi, S.; Zhou, L.; Zhou, Y.; Li, J.; Liu, W.; Yin, Y. Chlorogenic acid enhances intestinal barrier by decreasing MLCK expression and promoting dynamic distribution of tight junction proteins in colitic rats. J. Funct. Foods 2016, 26, 698–708. [Google Scholar] [CrossRef]
- Matsui, A.; Hashiguchi, K.; Suzuki, M.; Zhang-Akiyama, Q.M. Oxidation resistance 1 functions in the maintenance of cellular survival and genome stability in response to oxidative stress-independent dna damage. Genes Environ. 2020, 42, 42–49. [Google Scholar] [CrossRef]
- Engin, A.B.; Engin, A. DNA damage checkpoint response to aflatoxin B1. Environ. Toxicol. Pharmacol. 2019, 65, 90–96. [Google Scholar] [CrossRef]
- Yilmaz, S.; Kaya, E.; Kisacam, M.A. The Effect on Oxidative Stress of Aflatoxin and Protective Effect of Lycopene on Aflatoxin Damage. In Aflatoxin-Control, Analysis, Detection and Health Risks 30; IntechOpen: London, UK, 2017; pp. 67–90. [Google Scholar]
- Topal, A.; Alak, G.; Ozkaraca, M.; Yeltekin, A.C.; Comakli, S.; Acil, G.; Kokturk, M.; Atamanalp, M. Neurotoxic responses in brain tissues of rainbow trout exposed to imidacloprid pesticide: Assessment of 8-hydroxy-2-deoxyguanosine activity 2017, oxidative stress and acetylcholinesterase activity. Chemosphere 2017, 175, 186–191. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Niu, Y.; Wang, F.; Wang, C.; Cui, T.; Bai, K.; Zhang, J.; Zhong, X.; Zhang, L.; Wang, T. Dietary curcumin supplementation attenuates inflammation, hepatic injury and oxidative damage in a rat model of intra-uterine growth retardation. Br. J. Nutr. 2018, 120, 537–548. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.Y.; Qi, M.; Zhao, L.; Zhu, M.K.; Guo, J.; Liu, J.; Gu, C.Q.; Rajput, S.A.; Krumm, C.S.; Qi, D.S.; et al. Curcumin Prevents Aflatoxin B (1) Hepatoxicity by Inhibition of Cytochrome P450 Isozymes in Chick Liver. Toxins 2016, 8, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.L.; Chu, J.G.; Jian, X.M.; Dong, J.Z.; Wang, L.P.; Li, G.X.; Yang, N.B. Curcumin attenuates lipopolysaccharide/d-galactosamine-induced acute liver injury by activating Nrf2 nuclear translocation and inhibiting NF-kB activation. Biomed. Pharmacother. 2017, 91, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Bai, K.; Su, W.; Wang, A.; Zhang, L.; Huang, K.; Wang, T. Curcumin attenuates heat-stress-induced oxidant damage by simultaneous activation of GSH-related antioxidant enzymes and Nrf2-mediated phase II detoxifying enzyme systems in broiler chickens. Poult. Sci. 2018, 97, 1209–1219. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2019, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Enayati, A.A.; Ranson, H.; Hemingway, J. Insect glutathione transferases and insecticide resistance. Insect Mol. Biol. 2005, 14, 3–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, D.; Zhu, Y.W.; Fouad, A.M.; Yan, S.J.; Chen, W.; Zhang, Y.N.; Xia, W.G.; Wang, S.; Jiang, S.Q.; Yang, L.; et al. Dietary curcumin enhances intestinal antioxidant capacity in ducklings via altering gene expression of antioxidant and key detoxification enzymes. Poult. Sci. 2019, 98, 3705–3714. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.Y.; Chen, W.F.; Zhou, B.; Yang, L.; Liu, Z.L. Inhibition of lipid peroxidation and protein oxidation in rat liver mitochondria by curcumin and its analogues. Biochim. Biophys. Acta 2006, 1760, 70–77. [Google Scholar] [CrossRef]
- El-Bahr, S.M. Effect of Curcumin on Hepatic Antioxidant Enzymes Activities and Gene Expressions in Rats Intoxicated with Aflatoxin B1. Phytother. Res. 2015, 29, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Dinkova-Kostova, A.T. The nrf2 regulatory network provides an interface between redox and intermediary metabolism. Trends Biochem. Sci. 2014, 39, 199–218. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.J.; Pang, Q.; Yang, H.; Diao, X.P.; Shan, A.S.; Feng, X.J. Effects of dietary resveratrol supplementation on the chemical composition, oxidative stability and meat quality of ducks (Anas platyrhynchos). Food Chem. 2021, 130263. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Guzmán, C.; Cortés-Reynosa, P.; Pérez-Salazar, E.; Murillo-González, F.E.; Elizondo, G. Activation of the aryl hydrocarbon receptor (ahr) induces human glutathione s transferase alpha 1 (hgsta1) expression. Chem. Biol. Interact. 2020, 331, 109284. [Google Scholar] [CrossRef] [PubMed]
- Rubiolo, J.A.; Mithieux, G.; Vega, F.V. Resveratrol protects primary rat hepatocytes against oxidative stress damage: Activation of the nrf2 transcription factor and augmented activities of antioxidant enzymes. Eur. J. Pharmacol. 2008, 591, 66–72. [Google Scholar] [CrossRef]
- Abdel-Wahhab, M.A.; Aly, S.E. Antioxidants and radical scavenging properties of vegetable extracts in rats fed aflatoxin-contaminated diet. J. Agric. Food Chem. 2003, 51, 2409–2414. [Google Scholar] [CrossRef]
- Shen, H.M.; Shi, C.Y.; Lee, H.P.; Ong, C.N. Aflatoxin b1-induced lipid peroxidation in rat liver. Toxicol. Appl. Pharmacol. 1994, 127, 145–150. [Google Scholar] [CrossRef]
- Wang, H.; Muhammad, I.; Li, W.; Sun, X.; Cheng, P.; Zhang, X. Sensitivity of Arbor Acres broilers and chemoprevention of aflatoxin B1-induced liver injury by curcumin, a natural potent inducer of phase-II enzymes and Nrf2. Environ. Toxicol. Pharmacol. 2018, 59, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, P.; Shanshan, L.; Guo, R.; Yang, Z. Liquiritin, a novel inhibitor of TRPV1 and TRPA1, protects against LPS-induced acute lung injury. Cell Calcium 2020, 88, 102198. [Google Scholar] [CrossRef]
- Shanmugam, N.; Reddy, M.A.; Guha, M.; Natarajan, R. High glucose-induced expression of proinflammatory cytokine and chemokine genes in monocytic cells. Diabetes 2003, 52, 1256–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, H.K.; Lin, A.H.; Perng, D.W.; Lee, T.S.; Kou, Y.R. Lung epithelial trpa1 mediates lipopolysaccharide-induced lung inflammation in bronchial epithelial cells and mice. Front. Physiol. 2020, 11, 596314. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Gan, F.; Zhou, X.; Zhou, Y.; Qian, G.; Liu, Z.; Huang, K. Immunotoxicity of ochratoxin A and aflatoxin B1 in combination is associated with the nuclear factor kappa B signaling pathway in 3D4/21 cells. Chemosphere 2018, 199, 718–727. [Google Scholar] [CrossRef]
- Lang, V.; Rodríguez, M.S. Innate link between nf-kappab activity and ubiquitin-like modifiers. Biochem. Soc. Trans. 2008, 36, 853. [Google Scholar] [CrossRef] [PubMed]
- Kumara, S.S.; Gayathri, D.; Hariprasad, P.; Venkateswaran, G.; Swamy, C.T. In vivo AFB1 detoxification by lactobacillus fermentum LC5/a with chlorophyll and immunopotentiating activity in albino mice. Toxicon 2020, 187, 214–222. [Google Scholar] [CrossRef]
- Taranu, I.; Hermenean, A.; Bulgaru, C.; Pistol, G.C.; Ciceu, A.; Grosu, I.A.; Marin, D.E. Diet containing grape seed meal by-product counteracts AFB1 toxicity in liver of pig after weaning. Ecotoxicol. Environ. Saf. 2020, 203, 110899. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhang, D.; Hu, D.; Zhou, X.; Zhou, Y. The role of mitochondria in NLRP3 inflammasome activation. Mol. Immunol. 2018, 103, 115–124. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
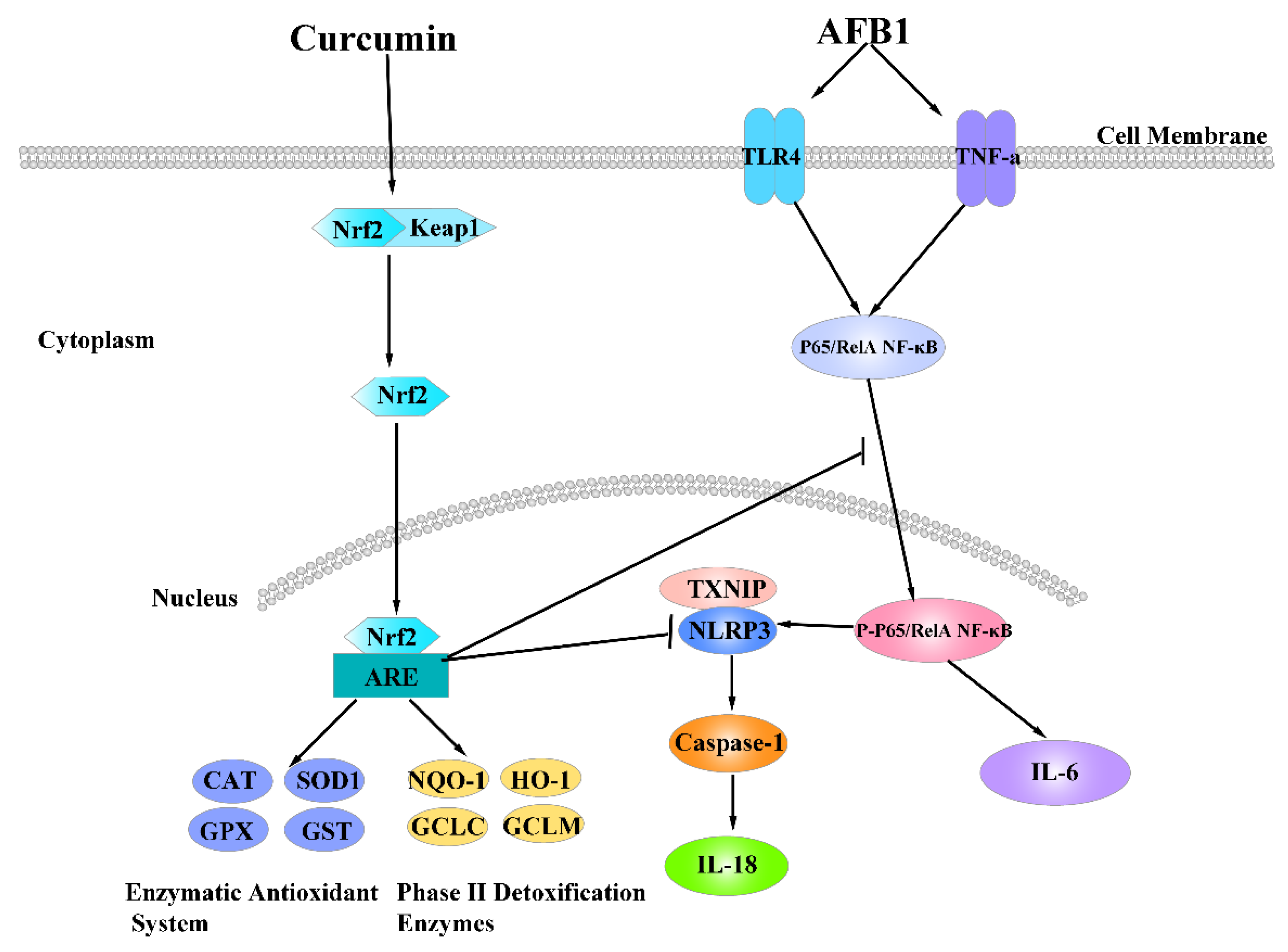{kind=link}
| Groups | Basal Diet | Curcumin (mg Curcumin/kg Basal Diet) |
|---|---|---|
| T0 | corn-soybean | 0 |
| T0 | corn-soybean | 0 + AFB1 |
| T500 | corn-soybean | 500 + AFB1 |
| Items | 1–4 Weeks 1 | 5–8 Weeks 2 | 9–10 Weeks 3 |
|---|---|---|---|
| Ingredient | |||
| Corn (7.9) | 61.70 | 68.94 | 75.80 |
| Soybean meal (45) | 26.09 | 26.80 | 20.10 |
| Corn protein flour (55) | 7.90 | - | - |
| Dicalcium phosphate | 1.40 | 1.40 | 1.40 |
| Limestone | 1.08 | 1.06 | 1.06 |
| Salt | 0.38 | 0.38 | 0.38 |
| DL-Methionine | 0.15 | 0.22 | 0.16 |
| L-Lysine | 0.20 | 0.10 | 0.00 |
| choline chloride (50%) | 0.10 | 0.10 | 0.10 |
| Premix | 1.00 | 1.00 | 1.00 |
| Total | 100 | 100 | 100 |
| Nutritional level | |||
| Calculated nutrient 4 | |||
| Metabolizable energy (MJ/kg) | 12.14 | 11.98 | 12.21 |
| CP (%) | 20.67 | 17.51 | 15.03 |
| Calcium (%) | 0.90 | 0.90 | 0.88 |
| Total phosphorus (%) | 0.68 | 0.67 | 0.65 |
| Non-phytate phosphorus (%) | 0.44 | 0.44 | 0.44 |
| Lysine (%) | 1.07 | 0.95 | 0.71 |
| Methionine (%) | 0.48 | 0.48 | 0.39 |
| Methionine + cystine (%) | 0.81 | 0.75 | 0.63 |
| Threonine (%) | 0.75 | 0.66 | 0.56 |
| Tryptophane (%) | 0.21 | 0.19 | 0.16 |
| Transcripts | Accession Number | Gene Sequence (5′–3′) | Product Length (bp) | |
|---|---|---|---|---|
| Keap1 | MF774811.1 | Forward | TCACCCTCCATAAACCCACCCAAG | 102 |
| Reverse | AGTAGCCCAAGGACTGCCGATAG | 102 | ||
| Nrf2 | NM_001310777.1 | Forward | GTTGAATCATCTGCCTGTGG | 171 |
| Reverse | TAAGCTAGGTGGTCGAGTGC | 172 | ||
| HO-1 | KU048806.1 | Forward | AAGAGCCAGGAGAACGGTCACC | 139 |
| Reverse | TGCCCACCAGGTCTGTCTGAC | 139 | ||
| SOD1 | XM_013097859.1 | Forward | CCTGTGGTGTCATCGGAATA | 116 |
| Reverse | TTGAACGAGGAAGAGCAAGTA | 127 | ||
| GCLC | XM_027455104.1 | Forward | TTCAGGTGACATTCCAGGCTTGC | 108 |
| Reverse | AGAACGGAGATGCAGCACTCAATG | 108 | ||
| GCLM | XM_027462629.1 | Forward | TGTTGTGTGATGCCACCTGATCTC | 150 |
| Reverse | CCATTCGTGTGCTTTGACGTTCTG | 150 | ||
| CAT | KU048802.1 | Forward | TGTGCGTGACTGACAACCAAGG | 96 |
| Reverse | ACATGCGGCTCTCCTTCACAAC | 96 | ||
| NQO-1 | XM_027466610.1 | Forward | CGTCGCCGAGCAGAAGAAGATC | 195 |
| Reverse | CTGGTGGTGAACGACAGCATGG | 195 | ||
| GST | LOC101797566 | Forward | ACAAGGCTGCAACCAGATACTTCC | 178 |
| Reverse | ACTGCACATCTGCTCTGCTAAGC | 178 | ||
| GPX | XM_027459004.1 | Forward | GAACGGCACCAACGAGGAGATC | 99 |
| Reverse | TTCACCTGGCACTTCTGGAACAG | 99 | ||
| NLRP3 | MH373356.1 | Forward | CGCTGAACGAGGACGCACTG | 124 |
| Reverse | TGGAAGGGTAGTCGGGACATAGC | 124 | ||
| TXNIP | XM_032204531.1 | Forward | GCTGCCAAGAAGGAGAAGAAGGTG | 130 |
| Reverse | TGTTCTCGAAGTCGGCGTTGATG | 130 | ||
| Caspase-1 | XM_027446016.1 | Forward | GCGGAACCAAGAGCAGAGATGAG | 130 |
| Reverse | CCACGGCAGGACTGGATAATAACC | 130 | ||
| IL-18 | NM_001310420.1 | Forward | GGCTCTGTCCCAAGGCAGGAG | 124 |
| Reverse | GCCACTCTGCGTCAGCTTCAC | 124 | ||
| IL-6 | JQ728554.1 | Forward | ATGTGCGAGAAGTTCACCGTCTG | 113 |
| Reverse | TCGTCGAAGCCAGCCAGGAG | 113 | ||
| TNF-a | XM_013105371.3 | Forward | AGCTGGCTAAGACCGTGGTCAG | 151 |
| Reverse | ACGTTGTTGACTCGTCCATGTGAC | 151 | ||
| TLR4 | NM_001310413.1 | Forward | GACCTCCAGCACACGAAGTTAGAC | 165 |
| Reverse | GGAGTTGCCTGCCATCTTGAGC | 165 | ||
| NF-κB | XM_027453277.2 | Forward | GGAGCAGTGGCGGTGTCAAC | 126 |
| Reverse | AGTGCAGTTCATGTCATCGGTCTC | 126 | ||
| β-actin | EF667345.1 | Forward | ATGTCGCCCTGGATTTCG | 62 |
| Reverse | CACAGGACTCCATACCCAAGAA | 62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, S.; Yang, H.; Jiao, Y.; Pang, Q.; Wang, Y.; Wang, M.; Shan, A.; Feng, X. Dietary Curcumin Alleviated Acute Ileum Damage of Ducks (Anas platyrhynchos) Induced by AFB1 through Regulating Nrf2-ARE and NF-κB Signaling Pathways. Foods 2021, 10, 1370. https://doi.org/10.3390/foods10061370
Jin S, Yang H, Jiao Y, Pang Q, Wang Y, Wang M, Shan A, Feng X. Dietary Curcumin Alleviated Acute Ileum Damage of Ducks (Anas platyrhynchos) Induced by AFB1 through Regulating Nrf2-ARE and NF-κB Signaling Pathways. Foods. 2021; 10(6):1370. https://doi.org/10.3390/foods10061370
Chicago/Turabian StyleJin, Sanjun, Hao Yang, Yihan Jiao, Qian Pang, Yingjie Wang, Min Wang, Anshan Shan, and Xingjun Feng. 2021. "Dietary Curcumin Alleviated Acute Ileum Damage of Ducks (Anas platyrhynchos) Induced by AFB1 through Regulating Nrf2-ARE and NF-κB Signaling Pathways" Foods 10, no. 6: 1370. https://doi.org/10.3390/foods10061370