
Mycotoxins Affecting Animals, Foods, Humans, and Plants: Types, Occurrence, Toxicities, Action Mechanisms, Prevention, and Detoxification Strategies—A Revisit

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Major Groups of Mycotoxins: Occurrence, Production, and Toxicities
2.1. Aflatoxins
2.2. Ochratoxins
2.3. Trichothecenes (Trichothecene Mycotoxins)
2.4. Deoxynivalenol (a Trichothecene)
2.5. Fumonisins
2.6. Emerging Fusarium Mycotoxins (Enniatins, NX-2 Toxin, Beauvericin, Moniliformin, Fusaproliferin)
2.7. Sterigmatocystin
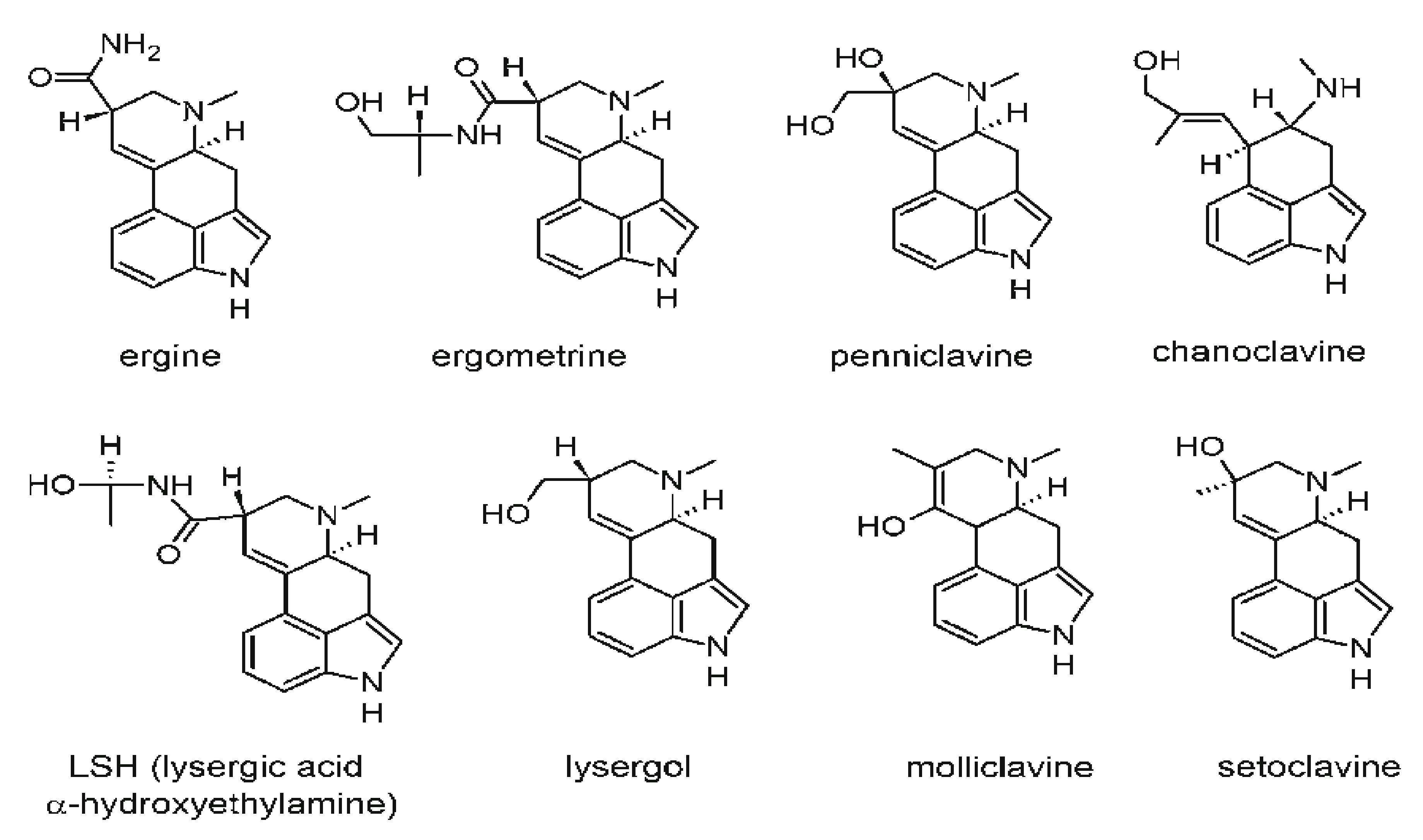
2.8. Ergot Alkaloids
2.9. Zearalenone
2.10. Alternaria Toxins (Altenuene, Tentoxin, Tenuazonic Acid, Altertoxin, Alternariol Methyl Ether, Alternariol)
2.11. Patulin (PAT)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mycotoxin | Common Fungal Species | Foods Where Commonly Found | Toxicities | Maximum Allowable Limits and Associated Remarks | Reference(s) |
|---|---|---|---|---|---|
| Aflatoxins (aflatoxins B1, B2, G1, G2, M1, M2) | Aspergillus parasiticus, Aspergillus flavus, Aspergillus bombycis, A. pseudotamarii, A. nomius, etc. | Cereals, legumes, fruits, seeds, vegetables, nuts, etc. | Liver cancer; hepatocellular carcinoma; target DNA; mutagenic and teratogenic effects | The EU set limits of 4 μg/kg and 2 μg/kg for total aflatoxins and AFB1 permitted, respectively, in nuts, dried fruits, and cereals meant for direct consumption by humans. The AFM1 maximum residue level in milk is set by the European Union and the United States at 50 ng/kg and 500 ng/kg of raw milk, respectively. The AFB1 maximum residue level in feeds of lactating cows is set at 5 μg AFB1/kg, 10 μg/kg, and 20 μg/kg of feeds in the EU, in China, and in the US, respectively | [19,26,27,34,176] |
| Ochratoxins (ochratoxins A, B, C) | Species of Aspergillus and Penicillium, including Aspergillus ochraceus, Aspergillus niger, Aspergillus carbonarius, Penicillium verrucosum | Cereals, legumes, seeds, fruits, vegetables, nuts, etc. | Immunotoxic, teratogenic, neurotoxic, hepatotoxic, and nephrotoxic activities; nephropathy in pigs; in humans, ochratoxin A was linked to urothelial tumors, chronic interstitial nephropathy, renal failure, and Balkan endemic nephropathy; etc. | In the EU, OTA limits in imported foods are set to a maximum of 10.0 μg/kg for instant coffee, 5 μg/kg for roasted coffee, 2 μg/kg for grape juice, 2 μg/kg for wine, 3 μg/kg for processed cereal food products, and 5 μg/kg for unprocessed cereal grains | [1,81] |
| Trichothecenes (trichothecene mycotoxins), examples include deoxynivalenol (vomitoxin), 3- and 15-acetyldeoxynivalenol, nivalenol, anguidine, T-2 toxin, HT-2 toxin, crotocin, diacetoxyscirpenol, macrocyclics, etc. | Species of Fusarium (Fusarium crookwellense, F culmorum, F graminearum, F poae), Myrothecium, Verticimonosporium, Trichothecium, Trichoderma, Cephalosporium, Stachybotrys, and Spicellum | Rice, oats, rye, barley, maize, wheat, vegetables, etc., and animal foods, including eggs, milk, liver, and kidneys | They can diffuse into cells and block translation by interacting with eukaryotic ribosomes; this is their primary action mechanism. Other action mechanisms for toxicity include inhibiting DNA, RNA, and protein synthesis, lipid peroxidation, apoptosis, inhibiting mitochondrial functions, neurotransmitters changes, and cytokine activation. Exposure to trichothecenes affects nearly all key systems in vertebrates … alimentary toxic aleukia (ATA) in humans, etc. | The US FDA has established a level of 1 ppm restriction for deoxynivalenol. The range of TDI of 100 ng/kg bw for the sum of T-2 and HT-2 toxins is used by the EFSA. | [16,96,98] |
| Fumonisins (fumonisins B1, B2, B3, etc.) | Fusarium species such as Fusarium verticillioides, Fusarium nyagamai, F. oxysporum, F. globosum, F. fujikuroi, F. proliferatum, Aspergillus awamori, A. niger etc. | Along with corn and corn food products, fumonisins have been reported in asparagus, sorghum, beer, rice, soybeans, beans, etc. | Fumonisins are linked to atherosclerosis in monkeys, esophageal and liver cancer in human, equine leukoencephalomalacia in horses, porcine pulmonary edema and pulmonary artery hypertrophy in swine, and kidney and liver cancer in rodents. Fumonisins inhibit sphingolipids synthesis. | The International Agency for Research on Cancer (IARC) has classified fumonisin B1 as possibly carcinogenic to humans (group 2B). The EU has put the maximum total fumonisin (fumonisins B1 and B2) limit at 1000 μg/kg for maize and maize products meant for direct consumption by humans and at 800 μg/kg for snacks and breakfast cereals produced from maize. The US FDA set a total limit of fumonisins at 2 to 4 mg/kg in corn and corn products intended for human consumption and at 3 mg/kg in corn used for popcorn. The Joint FAO/WHO Expert Committee on Food Additives put the maximum fumonisins tolerable intake per day at 2 μg/kg bw for fumonisins B1, B2, and B3, in combination or alone. | [19,114,116,117] |
| Emerging Fusarium mycotoxins (enniatins, NX-2 toxin, beauvericin, moniliformin, fusaproliferin, etc.) | Species of Fusarium, including Fusarium verticillioides, Fusarium subglutinans, Fusarium proliferatum, Fusarium acuminatum, F. avenaceum, F. arthrosporiodes, F. chlamydosporum, F. redolens, F. oxysporum, F. beomiforme, etc.; Beauveria bassiana | Corn, rice, corn products, seeds, nuts, coffee, tree nuts, dried fruits, beans, vegetable oil, etc. | As a result of their high prevalence in foods and feeds and their potential toxicity to humans and animals, the interest in emerging mycotoxins is increasing. Beauvericin has insecticidal, antifungal, and antibacterial properties and can have toxic effects, including apoptosis induction, increased cytoplasmic calcium concentration, and fragmentation of DNA in cell lines of mammals. | Not available | [126,127,133] |
| Sterigmatocystin | Aspergillus species, such as A. versicolor (major producer), A. sydowi, A. quadrilineatus, A. aureolatus, A. amstelodami, A. ruber, A. chevalieri, as well as species of Penicillium, Emiricella, Chaetomium, and Bipolaris | Peanuts, corn, barley, rice, wheat, grain products, etc. | Sterigmatocystin has teratogenic, mutagenic, and carcinogenic effects but is less potent than AFB1 and can cause hepatic toxicity in most animals; hepatocellular carcinoma and squamous cell carcinomas in rats; bloody diarrhea and death in cattle; LD50 in mice is 800 mg/kg and above | The IARC placed STC under class 2B carcinogens. The California Department of Health Services used values TD50 from the Cancer Potency Database to produce “no significant risk” intake levels for humans. The resulting level was 8 mcg/kg bw per day for a 70 kg adult. No limit has been made available in many countries | [140,144] |
| Ergot alkaloids | Comprised of a complex family of the derivatives of indole produced by the Clavicipitaceae (such as Neotyphodium and Claviceps) and Trichocomaceae (such as Penicillium and Aspergillus) families. Claviceps purpurea is the dominant producer | Rye (most common host), triticale, barley, wheat, oats, etc. | Causes ergotism; ergot alkaloids are both harmful and beneficial to humans; can cause delirious seizures, fits, St. Anthony’s Fire, etc.; can cause gangrenous and convulsive forms of toxicities | Maximum tolerable limits are in the EU commission pipeline while current ergot sclerotia content is set in unprocessed cereals at a maximum of 0.05%. In the US, rye and wheat are considered unsafe for consumption by humans if they contain above 0.3% sclerotia by weight, and barley, triticale, or oats are graded when they contain above 0.1%. The maximum ergot level set by the European Union is 0.05% in common wheat and durum, i.e., 500 mg/kg w/w sclerotia. | [147] |
| Zearalenone (formerly referred to as F-2 toxin) | Species of Fusarium, such as Fusarium crookwellense, Fusarium cerealis, Fusarium semitectum, Fusarium equiseti, Fusarium graminearum, Fusarium culmorum, etc. | Maize, soybean, rice, rye, sorghum, oats, barley, wheat, grain products, etc. | Zearalenone or its metabolic compounds are known to bind transcription factors, including pregnane X receptors involved in expressing enzymes in pathways of biosynthesis; zearalenone chronic administration can cause uterine fibroids, pituitary adenomas, hepatocellular carcinoma, and liver damage in mice, and chronic progressive hematotoxicity, testicular atrophy, cataracts, retinopathy, and nephropathy in rats; among other animals, pigs are more prone its toxicities | The tolerable daily intake (TDI) for zearalenone was set by the EFSA at 0.25 μg/kg bw/day, and is also recommended by other international bodies such as the Joint FAO/WHO Expert Committee on Food Additives (JECFA) | [155,156] |
| Alternaria toxins (altenuene, tentoxin, tenuazonic acid, altertoxin, alternariol methyl ether, alternariol) | Alternaria species such as Alternaria triticina, Alternaria tenuissima, Alternaria solani, Alternaria japonica, Alternaria dauci, Alternaria brassicae, Alternaria alternata | Fruits and vegetables, seeds, grains, plants, beer, fruit juices, vegetable juices, wine, peppers, fresh and dried tomatoes, flour, bran, wheat, dried fruit, cereal products (e.g., rice and oat flake), sunflower oil, sunflower seeds, etc. | Tenuazonic acid has phytotoxic and antibacterial properties and acute toxicities for dogs, chicken, and mice, in addition to hematological disorders in humans. Although most Alternaria toxins show low acute toxicities, alternariol methyl ether and alternariol are mostly toxic because of their genotoxic, cytotoxic, carcinogenic, and mutagenic effects, with scientific-based findings from toxicological studies in vitro involving mammalian and bacterial cells. | The toxicological concern threshold (TTC approach) was put into use by the EFSA; for genotoxic Alternaria toxins (AME and AOH), a 2.5 ng/kg body weight per day TTC value was set, while for non-genotoxic Alternaria toxins (TEN and TeA), a 1500 ng/kg body weight per day TTC value was set | [162,164] |
| Patulin | Penicillium expansum, A. clavatus, Penicillium patulum (Penicillium griseofulvum and Penicillium urticae), Penicillium crustosum, etc. | Apples, apple products, fruits, vegetables, cereals, legumes, seeds, nuts, etc. | Mutagenicity, teratogenicity, carcinogenesis, immunotoxicity, and neurotoxicity are chronic and acute effects patulin showed on cell cultures. PAT causes neurotoxic and immunotoxic effects in animals, but no reliable evidence has shown its carcinogenicity to humans. However, studies have shown human toxicities, such as hemorrhages, ulcerations, vomiting, and nausea | The US, EU, and Chinese authorities all set 50 μg/L/kg as the patulin upper limit in fruit and apple juices. The EU established a 10 μg/kg to 50 μg/kg limit depending on the type of food | [164,174,175] |
| Other common mycotoxins (tremorgenic mycotoxins, fusarins (fusarins A–F), 3-nitropropionic acid, cyclochlorotine, sporidesmin) | Tremorgenic mycotoxins are produced by Aspergillus terreus, species of the Penicillium genus, etc.; Pithomyces chartarum produces sporidesmin; cyclochlorotine is produced by Penicillium islandicum; 3-nitropropionic acid (3-NPA) is produced by the species of Arthrinium; fusarins are produced by the species of Fusarium, such as Fusarium verticillioides (formerly Fusarium moniliforme), Fusarium graminearum (Fusarium venenatum), Fusarium poae, Fusarium sporotrichioides, Fusarium oxysporum | Several foods and feeds | Tremorgenic mycotoxins cause “staggers syndrome” in livestock and are linked to neurological conditions, such as seizures, tremors, mental confusion, and even death in humans. Fusarins are mutagenic; 3-nitropropionic acid interjects mitochondrial electron transport; Cyclochlorotine interrupts myofibrils and is hepatotoxic in animals; due to the hydrophobicity of sporidesmin, it can be integrated easily into the membranes of cells, in which it changes the organization of the bilayer | The EU, the US, the WHO, etc., all have various limits for these mycotoxins | [175,176,177,178] |
2.12. Other Common Mycotoxins
3. Action Mechanisms of Mycotoxins: Key Aspects
- (a)
- Ribosomal binding: Trichothecene toxicities occur due to their capability to bind the eukaryotic ribosomes’ 60S subunit and inhibit the reaction of peptidyl transferase [80]. Ochratoxin A competes with phenylalanine–tRNA ligase and inhibits the synthesis of protein; both aspartame and phenylalanine reduce toxicity of OTA by competing with it [95,97].
- (b)
- Protein interaction: The plasma albumin binds to aflatoxins. After oxidation of AFB1 by cytochrome P450s, two epoxides are formed and they react with the lysine ε-amino group forming AFB1–albumin adducts [15,194,195]. Aflatoxins are immunosuppressive, and in several studies they suppressed immune response mediated by the cell and impaired phagocytosis and chemotaxis. Most immunotoxic properties of fumonisin B1 may be a result of its capability to alter the levels of mRNA and/or expression of IL-1β, IFN-γ, and TNF-α as shown in several scientific experiments [196,197]. Penitrem obstructs uptake of glutamate and GABA (γ-aminobutyric acid) into cerebellar synaptosomes, modulating the function of GABA receptors. One of the ways patulin exerts its toxicities is by causing a dose- and time-dependent phosphorylation increase of c-Jun N-terminal kinase, protein kinases 1 and 2 regulated via extracellular signal, and p38 kinase, contributing to downstream effects, including cell death and DNA damage [197,198]. A mycotoxin known as secalonic acid D, which causes “cleft palate”, phosphorylates the binding protein of the cAMP response element [198,199].
- (c)
- (d)
- Ionophore activity: Beauvericin and enniatins that are produced by species of Fusarium have ionophoric activities specific to potassium and cause an influx of potassium into the matrix of the mitochondria followed by swelling of the mitochondria [201].
- (e)
- Metabolic enzyme inhibition: OTA, citroviridin, and AFB1 affect the metabolism of carbohydrates, while rubratoxin B and trichothecenes interfere with the metabolism of lipids [117,125]. The checmical structure of fumonisins has a high similarity to those of sphinganine and sphingosine, the sphingolipid backbones. Consequently, fumonisins inhibit ceramide synthase competitively. Fumonisin B1 inhibits argininosuccinate synthetase [125].
- (f)
- (g)
- (h)
- (i)
- Necrosis and apoptosis: AFB1 cytotoxic effects in lymphocytes of humans involve necrosis, caspase activation, and apoptosis [206], which lead to programmed cell death and irreversible cell damage. The death of cells induced by necrosis does not follow the signal transduction pathway of apoptosis.
- (j)
- Mitochondrial interactions: Fumonisin B1 was found to obstruct the mitochondrial complex I in human neuroblastoma cells and rat primary astrocytes, resulting in reduced cellular and mitochondrial respiration and an increase in reactive oxygen species (ROS) generation with calcium signaling deregulation [207,208]. By binding covalently to the enzyme active site, 33-NPA permanently deactivates succinate dehydrogenase. Acrebol, from Acremonium exuviarum, inhibits mitochondrial complex III, consequently causing ATP depletion by inhibiting the chain of respiration [207,209].
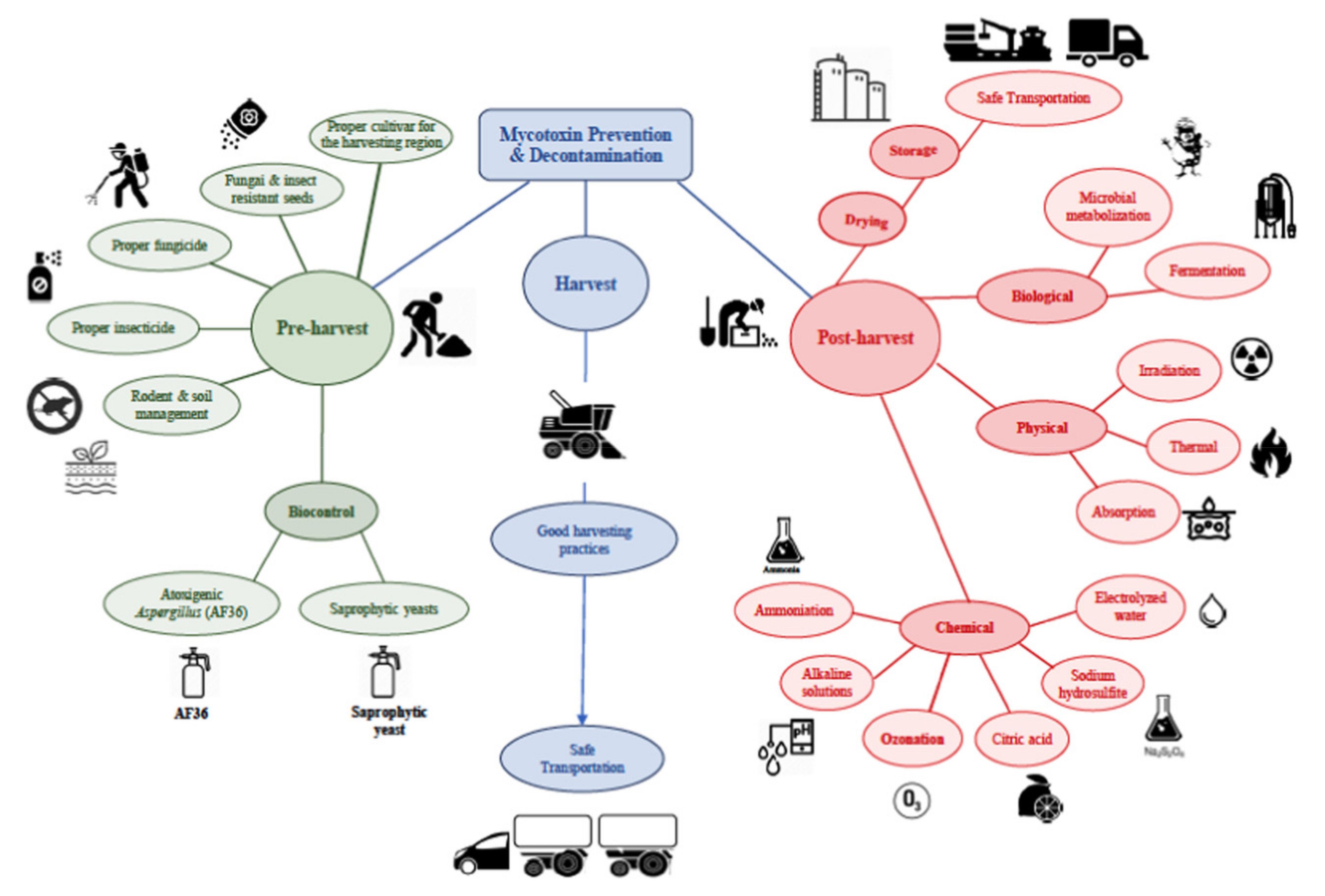
4. Mycotoxin Prevention Measures, Decontamination, and Detoxification Approaches
4.1. Pre-Harvest Preventive Measures
4.2. Post-Harvest Preventive Measures
4.2.1. Biological Strategies
Fermentation and Dietary Diversification
Fungi
Bacteria
Yeast
4.2.2. Physical Strategies
Storage Conditions
Radiation (Irradiation)
Use of Mycotoxin Binders
Sorting and Cleaning
Cold Plasma
Other Processing Methods (Frying, Baking, Peeling, Drying, etc.)
4.2.3. Detoxification with Enzymes
4.2.4. Chemical Strategies
Chitosan Usage
Ozone (O3)
Bases (Hydrated Oxide, Ammonia)
4.2.5. Other Emerging Strategies
Nanoparticles (NPs)
Extracts from Plants
Other Emerging Green Strategies
5. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Advances in Occurrence, Importance, and Mycotoxin Control Strategies: Prevention and Detoxification in Foods. Foods 2020, 9, 137. [Google Scholar] [CrossRef]
- Gnonlonfin, G.J.B.; Hell, K.; Adjovi, Y.; Fandohan, P.; Koudande, D.O.; Mensah, G.A.; Sanni, A.; Brimer, L. A Review on Aflatoxin Contamination and Its Implications in the Developing World: A Sub-Saharan African Perspective. Crit. Rev. Food Sci. Nutr. 2013, 53, 349–365. [Google Scholar] [CrossRef]
- Wu, F.; Groopman, J.D.; Pestka, J.J. Public Health Impacts of Foodborne Mycotoxins. Annu. Rev. Food Sci. Technol. 2014, 5, 351–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryden, W.L. Mycotoxins in the food chain: Human health implications. Asia Pac. J. Clin. Nutr. 2007, 16, 95–101. [Google Scholar] [PubMed]
- Paterson, R.R.M.; Lima, N. Toxicology of mycotoxins. Galanin 2010, 100, 31–63. [Google Scholar] [CrossRef]
- Egbuna, C.; Amadi, C.N.; Patrick-Iwuanyanwu, K.C.; Ezzat, S.M.; Awuchi, C.G.; Ugonwa, P.O.; Orisakwe, O.E. Emerging pollutants in Nigeria: A systematic review. Environ. Toxicol. Pharmacol. 2021, 85, 103638. [Google Scholar] [CrossRef] [PubMed]
- Awuchi, C.G.; Amagwula, I.O.; Priya, P.; Kumar, R.; Yezdani, U.; Khan, M.G. Aflatoxins In Foods And Feeds: A Review On Health Implications, Detection, And Control. Bull. Environ. Pharmacol. Life Sci. 2020, 9, 149–155. [Google Scholar]
- Urusov, A.E.; Zherdev, A.V.; Petrakova, A.V.; Sadykhov, E.G.; Koroleva, O.V.; Dzantiev, B.B. Rapid Multiple Immunoenzyme Assay of Mycotoxins. Toxins 2015, 7, 238–254. [Google Scholar] [CrossRef]
- Streit, E.; Schwab, C.; Sulyok, M.; Naehrer, K.; Krska, R.; Schatzmayr, G. Multi-Mycotoxin Screening Reveals the Occurrence of 139 Different Secondary Metabolites in Feed and Feed Ingredients. Toxins 2013, 5, 504–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anfossi, L.; Giovannoli, C.; Baggiani, C. Mycotoxin detection. Curr. Opin. Biotechnol. 2016, 37, 120–126. [Google Scholar] [CrossRef]
- Raiola, A.; Tenore, G.C.; Manyes, L.; Meca, G.; Ritieni, A. Risk analysis of main mycotoxins occurring in food for children: An overview. Food Chem. Toxicol. 2015, 84, 169–180. [Google Scholar] [CrossRef]
- Smith, L.E.; Stoltzfus, R.J.; Prendergast, A. Food Chain Mycotoxin Exposure, Gut Health, and Impaired Growth: A Conceptual Framework. Adv. Nutr. 2012, 3, 526–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avihingsanon, A.; Ramautarsing, R.A.; Suwanpimolkul, G.; Chetchotisakd, P.; Bowonwatanuwong, C.; Jirajariyavej, S.; Kantipong, P.; Tantipong, H.; Ohata, J.P.; Suankratay, C.; et al. Ergotism in Thailand Caused by Increased Access to Antiretroviral Drugs: A Global Warning. Top. Antivir. Med. 2016, 21, 165–168. [Google Scholar]
- Demir, S.; Akin, S.; Tercan, F.; Aribogan, A.; Oguzkurt, L. Ergotamine-induced lower extremity arterial vasospasm presenting as acute limb ischemia. Diagn. Interv. Radiol. 2008, 16, 165–167. [Google Scholar] [CrossRef] [Green Version]
- Haschek, W.M.; Gumprecht, L.A.; Smith, G.; Tumbleson, M.E.; Constable, P.D. Fumonisin toxicosis in swine: An overview of porcine pulmonary edema and current perspectives. Environ. Health Perspect. 2001, 109 (Suppl. S2), 251–257. [Google Scholar] [PubMed]
- Escrivá, L.; Font, G.; Manyes, L. In vivo toxicity studies of fusarium mycotoxins in the last decade: A review. Food Chem. Toxicol. 2015, 78, 185–206. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Dong, F.; Yu, M.; Xu, J.; Shi, J. Effect of preceding crop onFusariumspecies and mycotoxin contamination of wheat grains. J. Sci. Food Agric. 2016, 96, 4536–4541. [Google Scholar] [CrossRef] [PubMed]
- Klarić, M.Š.; Rašić, D.; Peraica, M. Deleterious Effects of Mycotoxin Combinations Involving Ochratoxin A. Toxins 2013, 5, 1965–1987. [Google Scholar] [CrossRef] [Green Version]
- Becker-Algeri, T.A.; Castagnaro, D.; De Bortoli, K.; De Souza, C.; Drunkler, D.A.; Badiale-Furlong, E. Mycotoxins in Bovine Milk and Dairy Products: A Review. J. Food Sci. 2016, 81, R544–R552. [Google Scholar] [CrossRef] [Green Version]
- Amaike, S.; Keller, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef]
- Iongh, D.; Beerthuis, R.K.; Vles, O.R.; Barrett, C.B.; Ord, W.O. Investigation of the factor in groundnut meal responsible for “turkey X disease”. Biochim. Biophys. Acta 1962, 65, 548–551. [Google Scholar] [CrossRef]
- Blount, W.P. Turkey “X” disease. J. Br. Turk. Fed. 1961, 9, 52–54. [Google Scholar]
- Klich, M.A. Aspergillus flavus: The major producer of aflatoxin. Mol. Plant. Pathol. 2007, 8, 713–722. [Google Scholar] [CrossRef]
- Mellon, J.E.; Cotty, P.J.; Dowd, M.K. Influence of Lipids with and without Other Cottonseed Reserve Materials on Aflatoxin B1Production byAspergillus flavus. J. Agric. Food Chem. 2000, 48, 3611–3615. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Ruyck, K.; De Boevre, M.; Huybrechts, I.; De Saeger, S. Dietary mycotoxins, co-exposure, and carcinogenesis in humans: Short review. Mutat. Res. Mutat. Res. 2015, 766, 32–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J. Current Understanding on Aflatoxin Biosynthesis and Future Perspective in Reducing Aflatoxin Contamination. Toxins 2012, 4, 1024–1057. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Jezkova, A.; Yuan, Z.; Pavlikova, L.; Dohnal, V.; Kuca, K. Biological degradation of aflatoxins. Drug Metab. Rev. 2009, 41, 1–7. [Google Scholar] [CrossRef]
- Roze, L.V.; Hong, S.-Y.; Linz, J.E. Aflatoxin Biosynthesis: Current Frontiers. Annu. Rev. Food Sci. Technol. 2013, 4, 293–311. [Google Scholar] [CrossRef]
- Kew, M.C. Aflatoxins as a cause of hepatocellular carcinoma. J. Gastrointest. Liver Dis. 2013, 22, 305–310. [Google Scholar]
- Montesano, R.; Hainaut, P.; Wild, C.P. Hepatocellular Carcinoma: From Gene to Public Health. J. Natl. Cancer Inst. 1997, 89, 1844–1851. [Google Scholar] [CrossRef] [Green Version]
- Nayak, S.; Sashidhar, R.B.; Bhat, R.V. Quantification and validation of enzyme immunoassay for urinary aflatoxin B1–N7-guanine adduct for biological monitoring of aflatoxins. Analyst 2001, 126, 179–183. [Google Scholar] [CrossRef]
- Essigmann, J.; Green, C.; Croy, R.; Fowler, K.; Buchi, G.; Wogan, G. Interactions of Aflatoxin B1 and Alkylating Agents with DNA: Structural and Functional Studies. Cold Spring Harb. Symp. Quant. Biol. 1983, 47, 327–337. [Google Scholar] [CrossRef]
- Giovati, L.; Magliani, W.; Ciociola, T.; Santinoli, C.; Conti, S.; Polonelli, L. AFM1 in Milk: Physical, Biological, and Prophylactic Methods to Mitigate Contamination. Toxins 2015, 7, 4330–4349. [Google Scholar] [CrossRef]
- Kebede, H.; Abbas, H.K.; Fisher, D.K.; Bellaloui, N. Relationship between Aflatoxin Contamination and Physiological Responses of Corn Plants under Drought and Heat Stress. Toxins 2012, 4, 1385–1403. [Google Scholar] [CrossRef] [PubMed]
- Narasaiah, K.V.; Sashidhar, R.B.; Subramanyam, C. Biochemical analysis of oxidative stress in the production of aflatoxin and its precursor intermediates. Mycopathologia 2006, 162, 179–189. [Google Scholar] [CrossRef]
- O’Brian, G.R.; Georgianna, D.R.; Wilkinson, J.R.; Yu, J.; Abbas, H.K.; Bhatnagar, D.; Cleveland, T.E.; Nierman, W.; Payne, G.A. The effect of elevated temperature on gene transcription and aflatoxin biosynthesis. Mycologia 2007, 99, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Awuchi, C.G.; Twinomhwezi, H.; Choudghal, S.; khan, M.G.; Yezdani, U.; Akram, M.V. Nanotechnology Application in Food Science and Nutrition and Its Safety Issues; a Review. Adv. Bioresearch 2020, 11. [Google Scholar] [CrossRef]
- Awuchi, C.G.; Amagwula, I.O. The Biochemistry, Toxicology, and Uses of the Pharmacologically Active Phytochemicals: Alkaloids, Terpenes, Polyphenols, and Glycosides. Merit Res. J. Food Sci. Technol. 2020, 5, 006–021. [Google Scholar] [CrossRef]
- Awuchi, C.G.; Owuamanam, I.C.; Ogueke, C.C.; Hannington, T. The Impacts of Mycotoxins on the Proximate Composition and Functional Properties of Grains. Eur. Acad. Res. 2020, 8, 1024–1071. [Google Scholar]
- Awuchi, C.G.; Chibueze, G.A.; Ukpe, A.E.; Asoegwu, C.R.; Uyo, C.N.; Ngoka, K.E. Environmental Impacts of Food and Agricultural Production: A Systematic Review. Eur. Acad. Res. 2020, 8, 1120–1135. [Google Scholar]
- Awuchi, C.G.; Hannington, T.; Igwe, V.S.; Amagwula, I.O. Food Additives and Food Preservatives for Domestic and Industrial Food Applications. J. Anim. Health 2020, 2, 1–16. [Google Scholar]
- Awuchi, C.G.; Igwe, V.S.; Amagwula, I.O.; Echeta, C.K. Health Benefits of Micronutrients (Vitamins and Minerals) and their Associated Deficiency Diseases: A Systematic Review. Int. J. Food Sci. 2020, 3, 1–32. [Google Scholar]
- Awuchi, C.G.; Igwe, V.S.; Amagwula, I.O. Nutritional Diseases and Nutrient Toxicities: A Systematic Review of the Diets and Nutrition for Prevention and Treatment. Int. J. Adv. Acad. Res. 2020, 6, 1–46. [Google Scholar] [CrossRef]
- Twinomuhwezi, H.; Wozeyi, P.; Igwe, V.S.; Amagwula, I.O.; Awuchi, C.G. Heat of Combustion of Coffee Pulp and Husks as Alternative Sources of Renewable Energy. Eur. J. Agric. Food Sci. 2021, 3, 1–4. [Google Scholar] [CrossRef]
- Awuchi, C.G. Medicinal Plants, Bioactive Compounds, and Dietary Therapies for Treating Type 1 and Type 2 Diabetes Mellitus. IntechOpen. Available online: https://www.intechopen.com/online-first/medicinal-plants-bioactive-compounds-and-dietary-therapies-for-treating-type-1-and-type-2-diabetes-m (accessed on 20 April 2021).
- Hannington, T.; Chinaza, G.A.; Mihigo, R. Comparative Study of the Proximate Composition and Functional Properties of Composite Flours of Amaranth, Rice, Millet, and Soybean. Am. J. Food Sci. Nutr. 2020, 6, 6–19. [Google Scholar]
- Georgianna, D.R.; Payne, G.A. Genetic regulation of aflatoxin biosynthesis: From gene to genome. Fungal Genet. Biol. 2009, 46, 113–125. [Google Scholar] [CrossRef]
- Wilkinson, J.R.; Yu, J.; Bland, J.M.; Nierman, W.C.; Bhatnagar, D.; Cleveland, T.E. Amino acid supplementation reveals differential regulation of aflatoxin biosynthesis in Aspergillus flavus NRRL 3357 and Aspergillus parasiticus SRRC. Appl. Microbiol. Biotechnol. 2007, 74, 1308–1319. [Google Scholar] [CrossRef]
- Hengstler, J.G.; Van Der Burg, B.; Steinberg, P.; Oesch, F. Interspecies differences in cancer susceptibility and toxicity*. Drug Metab. Rev. 1999, 31, 917–970. [Google Scholar] [CrossRef] [PubMed]
- Ilic, Z.; Crawford, D.; Egner, P.A.; Sell, S. Glutathione-S-transferase A3 knockout mice are sensitive to acute cytotoxic and genotoxic effects of aflatoxin B. Toxicol. Appl. Pharmacol. 2010, 242, 241–246. [Google Scholar] [CrossRef] [Green Version]
- Krishnamachari, K.; Nagarajan, V.; Bhat, R.; Tilak, T. Hepatitis due to aflatoxicosis. Lancet 1975, 305, 1061–1063. [Google Scholar] [CrossRef]
- Lye, M.S.; A Ghazali, A.; Mohan, J.; Alwin, N.; Nair, R.C. An outbreak of acute hepatic encephalopathy due to severe aflatoxicosis in Malaysia. Am. J. Trop. Med. Hyg. 1995, 53, 68–72. [Google Scholar] [PubMed]
- Lewis, L.; Onsongo, M.; Njapau, H.; Schurz-Rogers, H.; Luber, G.; Kieszak, S.; Nyamongo, J.; Backer, L.; Dahiye, A.M.; Misore, A.; et al. Aflatoxin Contamination of Commercial Maize Products during an Outbreak of Acute Aflatoxicosis in Eastern and Central Kenya. Environ. Heal. Perspect. 2005, 113, 1763–1767. [Google Scholar] [CrossRef] [PubMed]
- Coppock, R.W.; Reynolds, R.D.; Buck, W.B.; Jacobsen, B.J.; Ross, S.C.; Mostrom, M.S. Acute aflatoxicosis in feeder pigs, resulting from improper storage of corn. J. Am. Veter Med. Assoc. 1989, 195, 1380–1381. [Google Scholar]
- Vesonder, R.; Haliburton, J.; Stubblefield, R.; Gilmore, W.; Peterson, S. Aspergillus flavus and aflatoxins B1, B2, and M1 in corn associated with equine death. Arch. Environ. Contam. Toxicol. 1991, 20, 151–153. [Google Scholar] [CrossRef]
- Newman, S.J.; Smith, J.R.; Stenske, K.A.; Newman, L.B.; Dunlap, J.R.; Imerman, P.M.; Kirk, C.A. Aflatoxicosis in nine dogs after exposure to contaminated commercial dog food. J. Vet. Diagn. Investig. 2007, 19, 168–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Ryu, D. Advances in Mycotoxin Research: Public Health Perspectives. J. Food Sci. 2015, 80, T2970–T2983. [Google Scholar] [CrossRef]
- Gao, S.S.; Chen, X.Y.; Zhu, R.Z.; Choi, B.-M.; Kim, B.-R. Sulforaphane induces glutathione S-transferase isozymes which detoxify aflatoxin B1-8,9-epoxide in AML 12 cells. BioFactors 2010, 36, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Eaton, D.L.; Gallagher, E.P. Mechanisms of aflatoxin carcinogenesis. Annu. Rev. Pharmacol. Toxicol. 1994, 34, 135–172. [Google Scholar] [CrossRef]
- Stern, M.C.; Umbach, D.M.; Yu, M.C.; London, S.J.; Zhang, Z.Q.; Taylor, J.A. Hepatitis B. aflatoxin B(1), and p53 codon 249 mutation in hepatocellular carcinomas from Guangxi, People’s Republic of China, and a meta-analysis of existing studies. Cancer Epidemiol. Biomark. Prev. 2001, 10, 617–625. [Google Scholar]
- Aguilar, F.; Hussain, S.P.; Cerutti, P. Aflatoxin B1 induces the transversion of G-->T in codon 249 of the p53 tumor suppressor gene in human hepatocytes. Proc. Natl. Acad. Sci. USA 1993, 90, 8586–8590. [Google Scholar] [CrossRef] [Green Version]
- Hamid, A.S.; Tesfamariam, I.G.; Zhang, Y.; Zhang, Z.G. Aflatoxin B1-induced hepatocellular carcinoma in developing countries: Geographical distribution, mechanism of action and prevention. Oncol. Lett. 2013, 5, 1087–1092. [Google Scholar] [CrossRef] [Green Version]
- Friedler, A.; DeDecker, B.S.; Freund, S.M.; Blair, C.; Rüdiger, S.; Fersht, A.R. Structural Distortion of p53 by the Mutation R249S and its Rescue by a Designed Peptide: Implications for “Mutant Conformation”. J. Mol. Biol. 2004, 336, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Hsu, I.C.; Metcalf, R.A.; Sun, T.; Welsh, J.A.; Wang, N.J.; Harris, C.C. Mutational hotspot in the p53 gene in human hepatocellular carcinomas. Nature 1991, 350, 427–428. [Google Scholar] [CrossRef] [PubMed]
- Kew, M.C. Synergistic interaction between aflatoxin B1 and hepatitis B virus in hepatocarcinogenesis. Liver Int. 2003, 23, 405–409. [Google Scholar] [CrossRef]
- Ross, R.; Yu, M.; Henderson, B.; Yuan, J.-M.; Qian, G.-S.; Tu, J.-T.; Gao, Y.-T.; Wogan, G.; Groopman, J. Urinary aflatoxin biomarkers and risk of hepatocellular carcinoma. Lancet 1992, 339, 943–946. [Google Scholar] [CrossRef]
- Wild, C.P.; Montesano, R. A model of interaction: Aflatoxins and hepatitis viruses in liver cancer aetiology and prevention. Cancer Lett. 2009, 286, 22–28. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, F. Global Burden of Aflatoxin-Induced Hepatocellular Carcinoma: A Risk Assessment. Environ. Heal. Perspect. 2010, 118, 818–824. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.-H.; Zhang, X.-H.; Li, Y.-H.; Wang, J.-L.; Yan, X.; Xing, L.-X.; Wang, F.-R. Experimental lung carcinogenic in vivo study of aflatoxin G1 in NIH mice. Zhonghua Bing Li Xue Za Zhi Chin. J. Pathol. 2004, 33, 260–263. [Google Scholar]
- Shen, H.T.; Zhang, X.H.; Huang, X.H.; Li, Y.H.; Wang, J.; Yan, X.; Wang, F. Histogenesis of lung adenocarcinoma induced by oral administration of mycotoxins in mice. Wei Sheng Yan Jiu 2005, 34, 341–344. [Google Scholar] [PubMed]
- Guindon, K.A.; Foley, J.F.; Maronpot, R.R.; Massey, T.E. Failure of catalase to protect against aflatoxin B1-induced mouse lung tumorigenicity. Toxicol. Appl. Pharmacol. 2008, 227, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Machida, M.; Asai, K.; Sano, M.; Tanaka, T.; Kumagai, T.; Terai, G.; Kusumoto, K.-I.; Arima, T.; Akita, O.; Kashiwagi, Y.; et al. Genome sequencing and analysis of Aspergillus oryzae. Nat. Cell Biol. 2005, 438, 1157–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Whitelaw, C.A.; Nierman, W.C.; Bhatnagar, D.; Cleveland, T.E. Aspergillus flavus expressed sequence tags for identification of genes with putative roles in aflatoxin contamination of crops. FEMS Microbiol. Lett. 2004, 237, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Denning, D.W.; Anderson, M.J.; Turner, G.; Latgé, J.P.; Bennett, J.W. Sequencing the Aspergillus fumigatus genome. Lancet Infect. Dis. 2002, 2, 251–253. [Google Scholar] [CrossRef]
- Faustinelli, P.C.; Wang, X.M.; Palencia, E.R.; Arias, R.S. Genome Sequences of Eight Aspergillus flavus spp. and One A. parasiticus sp.; Isolated from Peanut Seeds in Georgia: TABLE. Genome Announc. 2016, 4, 00278-16. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Bhatnagar, D.; Cleveland, T.E. Completed sequence of aflatoxin pathway gene cluster inAspergillus parasiticus. FEBS Lett. 2004, 564, 126–130. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Chang, P.-K.; Ehrlich, K.C.; Cary, J.W.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A.; Linz, J.E.; Woloshuk, C.P.; Bennett, J.W. Clustered Pathway Genes in Aflatoxin Biosynthesis. Appl. Environ. Microbiol. 2004, 70, 1253–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marín, S.; Ramos, A.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef] [PubMed]
- Bayman, P.; Baker, J.L. Ochratoxins: A global perspective. Mycopathologia 2006, 162, 215–223. [Google Scholar] [CrossRef]
- Heussner, A.H.; Bingle, L.E.H. Comparative Ochratoxin Toxicity: A Review of the Available Data. Toxins 2015, 7, 4253–4282. [Google Scholar] [CrossRef] [Green Version]
- Van Der Merwe, K.J.; Steyn, P.S.; Fourie, L.; Scott, D.B.; Theron, J.J. Ochratoxin A, a Toxic Metabolite produced by Aspergillus ochraceus Wilh. Nat. Cell Biol. 1965, 205, 1112–1113. [Google Scholar] [CrossRef]
- Bui-Klimke, T.R.; Wu, F. Ochratoxin A and Human Health Risk: A Review of the Evidence. Crit. Rev. Food Sci. Nutr. 2015, 55, 1860–1869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, L.; Bhoola, K. Ochratoxins—Food Contaminants: Impact on Human Health. Toxins 2010, 2, 771–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petzinger, E.; Ziegler, K. Ochratoxin A from a toxicological perspective. J. Veter Pharmacol. Ther. 2000, 23, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Kuruc, J.A.; Schwarz, P.; Wolf-Hall, C. Ochratoxin A in Stored U.S. Barley and Wheat. J. Food Prot. 2015, 78, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Vidal, A.; Marín, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Determination of aflatoxins, deoxynivalenol, ochratoxin A and zearalenone in wheat and oat based bran supplements sold in the Spanish market. Food Chem. Toxicol. 2013, 53, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Anselme, M.; Tangni, E.K.; Pussemier, L.; Motte, J.-C.; Van Hove, F.; Schneider, Y.-J.; Van Peteghem, C.; Larondelle, Y. Comparison of ochratoxin A and deoxynivalenol in organically and conventionally produced beers sold on the Belgian market. Food Addit. Contam. 2006, 23, 910–918. [Google Scholar] [CrossRef]
- Abouzied, M.M.; Horvath, A.D.; Podlesny, P.M.; Regina, N.P.; Metodiev, V.D.; Kamenova-Tozeva, R.M.; Niagolova, N.D.; Stein, A.D.; Petropoulos, E.A.; Ganev, V.S. Ochratoxin A concentrations in food and feed from a region with Balkan Endemic Nephropathy. Food Addit. Contam. 2002, 19, 755–764. [Google Scholar] [CrossRef]
- Malir, F.; Ostry, V.; Pfohl-Leszkowicz, A.; Novotna, E. Ochratoxin A: Developmental and Reproductive Toxicity—An Overview. Birth Defects Res. B Dev. Reprod. Toxicol. 2013, 98, 493–502. [Google Scholar] [CrossRef]
- Mayura, K.; Reddy, R.V.; Hayes, A.W.; Berndt, W.O. Embryocidal, fetotoxic and teratogenic effects of ochratoxin A in rats. Toxicology 1982, 25, 175–185. [Google Scholar] [CrossRef]
- Kanetsky, P.A.; Mitra, N.; Vardhanabhuti, S.; Vaughn, D.J.; Li, M.; Ciosek, S.L.; Letrero, R.; D’Andrea, K.; Vaddi, M.; Doody, D.R.; et al. A second independent locus within DMRT1 is associated with testicular germ cell tumor susceptibility. Hum. Mol. Genet. 2011, 20, 3109–3117. [Google Scholar] [CrossRef]
- Wang, Z.; Wu, Q.; Kuča, K.; Dohnal, V.; Tian, Z. Deoxynivalenol: Signaling pathways and human exposure risk assessment—an update. Arch. Toxicol. 2014, 88, 1915–1928. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From Simple to Complex Mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef] [PubMed]
- Foroud, N.A.; Eudes, F. Trichothecenes in Cereal Grains. Int. J. Mol. Sci. 2009, 10, 147–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostrom, M.S.; Raisbeck, M.F. Trichothecenes. Vet. Toxicol. 2012, 1239–1265. [Google Scholar] [CrossRef]
- Pestka, J. Mechanisms of deoxynivalenol-induced gene expression and apoptosis. Food Addit. Contam. Part. A 2008, 25, 1128–1140. [Google Scholar] [CrossRef]
- Wan, Q.; Wu, G.; He, Q.; Tang, H.; Wang, Y. The toxicity of acute exposure to T-2 toxin evaluated by the metabonomics technique. Mol. BioSyst. 2015, 11, 882–891. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, M.; Bhaskar, A.S.B.; Rao, P.V.L. Differential effects of route of T-2 toxin exposure on hepatic oxidative damage in mice. Environ. Toxicol. 2013, 30, 64–73. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Z.; Beier, R.C.; Shen, J.; De Smet, D.; De Saeger, S.; Zhang, S. T-2 Toxin, a Trichothecene Mycotoxin: Review of Toxicity, Metabolism, and Analytical Methods. J. Agric. Food Chem. 2011, 59, 3441–3453. [Google Scholar] [CrossRef]
- Weekley, L.B.; E O’Rear, C.; Kimbrough, T.D.; Llewellyn, G.C. Acute and chronic effects of the trichothecene mycotoxin T-2 on rat brain regional concentrations of serotonin, tryptophan, and tyrosine. Veter Hum. Toxicol. 1989, 31, 221–224. [Google Scholar]
- Königs, M.; Mulac, D.; Schwerdt, G.; Gekle, M.; Humpf, H.-U. Metabolism and cytotoxic effects of T-2 toxin and its metabolites on human cells in primary culture. Toxicol. 2009, 258, 106–115. [Google Scholar] [CrossRef]
- Joffe, A.Z. Alimentary toxic aleukia. In Microbial Toxins; Kadis, S., Ciegler, A., Ajl, S.J., Eds.; Academic Press: Cambridge, MA, USA, 1971; Volume 7, pp. 139–189. [Google Scholar]
- Wegulo, S.N. Factors Influencing Deoxynivalenol Accumulation in Small Grain Cereals. Toxins 2012, 4, 1157–1180. [Google Scholar] [CrossRef]
- Awad, W.; Ghareeb, K.; Böhm, J.; Zentek, J. The Toxicological Impacts of the Fusarium Mycotoxin, Deoxynivalenol, in Poultry Flocks with Special Reference to Immunotoxicity. Toxins 2013, 5, 912–925. [Google Scholar] [CrossRef]
- Sobrova, P.; Adam, V.; Vasatkova, A.; Beklova, M.; Zeman, L.; Kizek, R. Deoxynivalenol and its toxicity. Interdiscip. Toxicol. 2010, 3, 94–99. [Google Scholar] [CrossRef]
- Hughes, D.M.; Gahl, M.J.; Graham, C.H.; Grieb, S.L. Overt signs of toxicity to dogs and cats of dietary deoxynivalenol. J. Anim. Sci. 1999, 77, 693–700. [Google Scholar] [CrossRef]
- Audenaert, K.; Vanheule, A.; Höfte, M.; Haesaert, G. Deoxynivalenol: A Major Player in the Multifaceted Response of Fusarium to Its Environment. Toxins 2013, 6, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.-Y.; Li, Q.; Meng, F.-G.; Fu, Y.; Zhao, Z.-J.; Wang, L.-H. T-2 Toxin Contamination in Grains and Selenium Concentration in Drinking Water and Grains in Kaschin–Beck Disease Endemic Areas of Qinghai Province. Biol. Trace Element Res. 2012, 150, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.H.; Fu, Y.; Shi, Y.X.; Wang, W.G. T-2 toxin induces degenerative articular changes in rodents: Link to Kaschin-Beck disease. Toxicol. Pathol. 2011, 39, 502–507. [Google Scholar] [CrossRef]
- Kang, P.; Yao, Y.; Yang, J.; Shen, B.; Zhou, Z.; Pei, F. An animal model of Kashin–Beck disease induced by a low-nutrition diet and exposure to T-2 toxin. Osteoarthr. Cartil. 2013, 21, 1108–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Chu, Y.; Cao, J.; Wang, W.; Liu, J.; Wang, J. Effects of T-2 toxin and selenium on chondrocyte expression of matrix metalloproteinases (MMP-1, MMP-13), α2-macroglobulin (α2M) and TIMPs. Toxicol. Vitr. 2011, 25, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Marasas, W.F.; Riley, R.T.; Hendricks, K.A.; Stevens, V.L.; Sadler, T.W.; Gelineau-van Waes, J.; Missmer, S.A.; Cabrera, J.; Torres, O.; Gelderblom, W.C.; et al. Fumonisins disrupt sphingolipid metabolism, folate transport, and neural tube development in embryo culture and in vivo: A potential risk factor for human neural tube defects among populations consuming fumonisin-contaminated maize. J. Nutr. 2004, 134, 711–716. [Google Scholar] [CrossRef]
- Bryla, M.; Roszko, M.; Szymczyk, K.; Jedrzejczak, R.; Obiedzinski, M.W.; Sekul, J. Fumonisins in plant-origin food and fodder–a review. Food Addit. Contam. Part. A 2013, 30, 1626–1640. [Google Scholar] [CrossRef]
- Stockmann-Juvala, H.; Savolainen, K. A review of the toxic effects and mechanisms of action of fumonisin B. Hum. Exp. Toxicol. 2008, 27, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Scott, P.M. Recent research on fumonisins: A review. Food Addit. Contam. Part. A 2012, 29, 242–248. [Google Scholar] [CrossRef]
- Domijan, A.M. Fumonisin B(1): A neurotoxic mycotoxin. Arch. Ind. Hyg. Toxicol. 2012, 63, 531–544. [Google Scholar]
- Soriano, J.; González, L.; Catalá, A. Mechanism of action of sphingolipids and their metabolites in the toxicity of fumonisin B. Prog. Lipid Res. 2005, 44, 345–356. [Google Scholar] [CrossRef]
- Bullerman, L.B.; Bianchini, A. Stability of mycotoxins during food processing. Int. J. Food Microbiol. 2007, 119, 140–146. [Google Scholar] [CrossRef]
- Cahagnier, B.; Melcion, D.; Richard-Molard, D. Growth of Fusarium moniliforme and its biosynthesis of fumonisin B1 on maize grain as a function of different water activities. Lett. Appl. Microbiol. 1995, 20, 247–251. [Google Scholar] [CrossRef]
- Mogensen, J.M.; Nielsen, K.F.; A Samson, R.; Frisvad, J.C.; Thrane, U. Effect of temperature and water activity on the production of fumonisins by Aspergillus niger and different Fusarium species. BMC Microbiol. 2009, 9, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sydenham, E.W.; Thiel, P.G.; Marasas, W.F.O.; Shephard, G.S.; Van Schalkwyk, D.J.; Koch, K.R. Natural occurrence of some Fusarium mycotoxins in corn from low and high esophageal cancer prevalence areas of the Transkei, Southern Africa. J. Agric. Food Chem. 1990, 38, 1900–1903. [Google Scholar] [CrossRef]
- Missmer, S.A.; Suarez, L.; Felkner, M.; Wang, E.; Merrill, A.H., Jr.; Rothman, K.J.; Hendricks, K.A. Exposure to fumonisins and the occurrence of neural tube defects along the Texas-Mexico border. Environ. Health Perspect. 2006, 114, 237–241. [Google Scholar] [CrossRef]
- Magoha, H.; De Meulenaer, B.; Kimanya, M.; Hipolite, D.; Lachat, C.; Kolsteren, P. Fumonisin B1 contamination in breast milk and its exposure in infants under 6 months of age in Rombo, Northern Tanzania. Food Chem. Toxicol. 2014, 74, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, G.R.; Tolleson, W.H.; Newkirk, D.K.; Roberts, D.W.; Rowland, K.L.; Saheki, T.; Kobayashi, K.; Howard, P.C.; Melchior, W.B., Jr. Identification of fumonisin B1 as an inhibitor of argininosuccinate synthetase using fumonisin affinity chromatography and in vitro kinetic studies. J. Biochem. Mol. Toxiclogy 2000, 14, 320–328. [Google Scholar] [CrossRef]
- Sadiq, F.A.; Yan, B.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. Lactic Acid Bacteria as Antifungal and Anti-Mycotoxigenic Agents: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1403–1436. [Google Scholar] [CrossRef] [Green Version]
- Vaclavikova, M.; Malachova, A.; Veprikova, Z.; Dzuman, Z.; Zachariasova, M.; Hajslova, J. ‘Emerging’ mycotoxins in cereals processing chains: Changes of enniatins during beer and bread making. Food Chem. 2013, 136, 750–757. [Google Scholar] [CrossRef]
- European Food Safety Authority. Scientific Opinion on the risks to human and animal health related to the presence of beauvericin and enniatins in food and feed. EFSA J. 2014, 12, 3802. [Google Scholar] [CrossRef]
- Gruber-Dorninger, C.; Novak, B.; Nagl, V.; Berthiller, F. Emerging Mycotoxins: Beyond Traditionally Determined Food Contaminants. J. Agric. Food Chem. 2017, 65, 7052–7070. [Google Scholar] [CrossRef]
- Fraeyman, S.; Croubels, S.; Devreese, M.; Antonissen, G. Emerging Fusarium and Alternaria Mycotoxins: Occurrence, Toxicity and Toxicokinetics. Toxins 2017, 9, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luz, C.; Saladino, F.; Luciano, F.; Mañes, J.; Meca, G. Occurrence, toxicity, bioaccessibility and mitigation strategies of beauvericin, a minor Fusarium mycotoxin. Food Chem. Toxicol. 2017, 107, 430–439. [Google Scholar] [CrossRef]
- Stanciu, O.; Juan, C.; Miere, D.; Loghin, F.; Mañes, J. Presence of Enniatins and Beauvericin in Romanian Wheat Samples: From Raw Material to Products for Direct Human Consumption. Toxins 2017, 9, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshinari, T.; Suzuki, Y.; Sugita-Konishi, Y.; Ohnishi, T.; Terajima, J. Occurrence of beauvericin and enniatins in wheat flour and corn grits on the Japanese market, and their co-contamination with type B trichothecene mycotoxins. Food Addit. Contam. Part. A 2016, 33, 1620–1626. [Google Scholar] [CrossRef] [PubMed]
- Quiles, J.M.; Saladino, F.; Mañes, J.; Fernández-Franzón, M.; Meca, G. Occurrence of mycotoxins in refrigerated pizza dough and risk assessment of exposure for the Spanish population. Food Chem. Toxicol. 2016, 94, 19–24. [Google Scholar] [CrossRef]
- Sulyok, M.; Beed, F.; Boni, S.; Abass, A.; Mukunzi, A.; Krska, R. Quantitation of multiple mycotoxins and cyanogenic glucosides in cassava samples from Tanzania and Rwanda by an LC-MS/MS-based multi-toxin method. Food Addit. Contam. Part. A 2014, 32, 488–502. [Google Scholar] [CrossRef] [PubMed]
- Varga, E.; Wiesenberger, G.; Hametner, C.; Ward, T.J.; Dong, Y.; Schöfbeck, D.; McCormick, S.; Broz, K.; Stückler, R.; Schuhmacher, R.; et al. New tricks of an old enemy: Isolates of Fusarium graminearum produce a type A trichothecene Mycotoxin. Environ. Microbiol. 2015, 17, 2588–2600. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, N.; Keller, N.P. Mycotoxins in Conversation with Bacteria and Fungi. Front. Microbiol. 2019, 10, 403. [Google Scholar] [CrossRef] [PubMed]
- Agriopoulou, S. Enniatins: An Emerging Food Safety Issue. EC Nutr. 2016, 3, 1142–1146. [Google Scholar]
- Gunter, A.B.; Hermans, A.; Bosnich, W.; Johnson, D.A.; Harris, L.; Gleddie, S. Protein engineering of Saccharomyces cerevisiae transporter Pdr5p identifies key residues that impact Fusarium mycotoxin export and resistance to inhibition. MicrobiologyOpen 2016, 5, 979–991. [Google Scholar] [CrossRef]
- Veršilovskis, A.; De Saeger, S. Sterigmatocystin: Occurrence in foodstuffs and analytical methods—An overview. Mol. Nutr. Food Res. 2009, 54, 136–147. [Google Scholar] [CrossRef]
- Li, M.; Li, P.; Wu, H.; Zhang, Q.; Ma, F.; Zhang, Z.; Ding, X.; Wang, H. An Ultra-Sensitive Monoclonal Antibody-Based Competitive Enzyme Immunoassay for Sterigmatocystin in Cereal and Oil Products. PLoS ONE 2014, 9, e106415. [Google Scholar] [CrossRef]
- Veršilovskis, A.; Bartkevičs, V.; Miķelsone, V. Sterigmatocystin presence in typical Latvian grains. Food Chem. 2008, 109, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Reiss, J. Mycotoxins in foodstuffs. VI. Formation of sterigmatocystin in bread by Aspergillus versicolor. Z. Lebensm. Unters. Und-Forsch. 1976, 160, 313–319. [Google Scholar]
- Gao, W.; Jiang, L.; Ge, L.; Chen, M.; Geng, C.; Yang, G.; Li, Q.; Ji, F.; Yan, Q.; Zou, Y.; et al. Sterigmatocystin-induced oxidative DNA damage in human liver-derived cell line through lysosomal damage. Toxicol. Vitr. 2015, 29, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xing, X.; Wang, J.; Xing, L.; Su, Y.; Yao, Z.; Yan, X.; Wang, J.; Zhang, X. Sterigmatocystin alters the number of FoxP3+ regulatory T cells and plasmacytoid dendritic cells in BALB/c mice. Food Chem. Toxicol. 2012, 50, 1920–1926. [Google Scholar] [CrossRef]
- Wallwey, C.; Li, S.-M. Ergot alkaloids: Structure diversity, biosynthetic gene clusters and functional proof of biosynthetic genes. Nat. Prod. Rep. 2010, 28, 496–510. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.L.; Panaccione, D.G. Diversification of Ergot Alkaloids in Natural and Modified Fungi. Toxins 2015, 7, 201–218. [Google Scholar] [CrossRef] [Green Version]
- Rimar, Y.; Rimar, D. Witches saints and other diseases. Harefuah 2003, 142, 383–386396. [Google Scholar] [PubMed]
- Alm, T. The Witch Trials of Finnmark, Northern Norway, during the 17th Century: Evidence for Ergotism as a Contributing Factor. Econ. Bot. 2003, 57, 403–416. [Google Scholar] [CrossRef]
- Schiff, P.L. Ergot and Its Alkaloids. Am. J. Pharm. Educ. 2006, 70, 98. [Google Scholar] [CrossRef]
- Strickland, J.R.; Looper, M.L.; Matthews, J.C.; Rosenkrans, C.F., Jr.; Flythe, M.D.; Brown, K.R. Board-invited review: St. Anthony’s Fire in livestock: Causes, mechanisms, and potential solutions. J. Anim. Sci. 2011, 89, 1603–1626. [Google Scholar] [CrossRef]
- Klotz, J.L. Activities and Effects of Ergot Alkaloids on Livestock Physiology and Production. Toxins 2015, 7, 2801–2821. [Google Scholar] [CrossRef]
- Craig, A.M.; Klotz, J.L.; Duringer, J.M. Cases of ergotism in livestock and associated ergot alkaloid concentrations in feed. Front. Chem. 2015, 3, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fink-Gremmels, J.; Malekinejad, H. Clinical effects and biochemical mechanisms associated with exposure to the mycoestrogen zearalenone. Anim. Feed. Sci. Technol. 2007, 137, 326–341. [Google Scholar] [CrossRef]
- Dänicke, S.; Winkler, J. Invited review: Diagnosis of zearalenone (ZEN) exposure of farm animals and transfer of its residues into edible tissues (carry over). Food Chem. Toxicol. 2015, 84, 225–249. [Google Scholar] [CrossRef]
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef]
- Tomaszewski, J.; Miturski, R.; Semczuk, A.; Kotarski, J.; Jakowicki, J. Tissue zearalenone concentration in normal, hyperplastic and neoplastic human endometrium. Ginekol. Polska 1998, 69, 363–366. [Google Scholar]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; De Saeger, S.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2012, 57, 165–186. [Google Scholar] [CrossRef] [PubMed]
- De Boevre, M.; Jacxsens, L.; Lachat, C.; Eeckhout, M.; Di Mavungu, J.D.; Audenaert, K.; Maene, P.; Haesaert, G.; Kolsteren, P.; De Meulenaer, B.; et al. Human exposure to mycotoxins and their masked forms through cereal-based foods in Belgium. Toxicol. Lett. 2013, 218, 281–292. [Google Scholar] [CrossRef] [Green Version]
- Logrieco, A.; Moretti, A.; Solfrizzo, M. Alternaria toxins and plant diseases: An overview of origin, occurrence and risks. World Mycotoxin J. 2009, 2, 129–140. [Google Scholar] [CrossRef]
- Sivagnanam, K.; Komatsu, E.; Rampitsch, C.; Perreault, H.; Gräfenhan, T. Rapid screening of Alternaria mycotoxins using MALDI-TOF mass spectrometry. J. Sci. Food Agric. 2017, 97, 357–361. [Google Scholar] [CrossRef]
- European Food Safety Authority. Scientific Opinion on the risks for animal and public health related to the presence of Alternaria toxins in feed and food. EFSA J. 2011, 10, 2407. [Google Scholar]
- Zwickel, T.; Klaffke, H.; Richards, K.; Rychlik, M. Development of a high performance liquid chromatography tandem mass spectrometry based analysis for the simultaneous quantification of various Alternaria toxins in wine, vegetable juices and fruit juices. J. Chromatogr. A 2016, 1455, 74–85. [Google Scholar] [CrossRef]
- Bernal, Á.R.R.; Reynoso, C.M.; Londoño, V.A.G.; Broggi, L.E.; Resnik, S.L. Alternaria toxins in Argentinean wheat, bran, and flour. Food Addit. Contam. Part. B 2019, 12, 24–30. [Google Scholar] [CrossRef]
- Sanzani, S.M.; Gallone, T.; Garganese, F.; Caruso, A.G.; Amenduni, M.; Ippolito, A. Contamination of fresh and dried tomato by Alternaria toxins in southern Italy. Food Addit. Contam. Part. A 2019, 36, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Walravens, J.; Mikula, H.; Rychlik, M.; Asamd, S.; Ediagea, E.N.; Di Mavungua, J.D.; Landschoote, A.V.; Vanhaeckef, L.; De Saeger, S. Development and validation of an ultra-high-performance liquid chromatography tandem mass spectrometric method for the simultaneous determination of free and conjugated Alternaria toxins in cereal-based foodstuffs. J. Chromatogr. A 2014, 1372, 91–101. [Google Scholar] [CrossRef]
- Brzonkalik, K.; Herrling, T.; Syldatk, C.; Neumann, A. Process development for the elucidation of mycotoxin formation in Alternaria alternata. AMB Express 2011, 1, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleck, S.C.; Burkhardt, B.; Pfeiffer, E.; Metzler, M. Alternaria toxins: Altertoxin II is a much stronger mutagen and DNA strand breaking mycotoxin than alternariol and its methyl ether in cultured mammalian cells. Toxicol. Lett. 2012, 214, 27–32. [Google Scholar] [CrossRef]
- Müller, M.E.; Korn, U. Alternaria mycotoxins in wheat—A 10 years survey in the Northeast of Germany. Food Control. 2013, 34, 191–197. [Google Scholar] [CrossRef]
- Lee, H.B.; Patriarca, A.; Magan, N. Alternaria in Food: Ecophysiology, Mycotoxin Production and Toxicology. Mycobiology 2015, 43, 93–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, M.Á.P.; Alonso, I.G.; De Santos, R.M.; Lacarra, T.G. The importance of genus Alternaria in mycotoxins production and human diseases. Nutr. Hosp. 2013, 27, 1772–1781. [Google Scholar]
- Vidal, A.; Ouhibi, S.; Ghali, R.; Hedhili, A.; De Saeger, S.; De Boevre, M. The mycotoxin patulin: An updated short review on occurrence, toxicity and analytical challenges. Food Chem. Toxicol. 2019, 129, 249–256. [Google Scholar] [CrossRef]
- Zhong, L.; Carere, J.; Lu, Z.; Lu, F.; Zhou, T. Patulin in Apples and Apple-Based Food Products: The Burdens and the Mitigation Strategies. Toxins 2018, 10, 475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mardani, M.; Rezapour, S.; Rezapour, P. Survey of aflatoxins in Kashkineh: A traditional Iranian food. Iran. J. Microbiol. 2011, 3, 147–151. [Google Scholar] [PubMed]
- Eriksen, G.S.; Jaderlund, K.H.; Moldes-Anaya, A.; Schonheit, J.; Bernhoft, A.; Jaeger, G.; Rundberget, T.; Skaar, I. Poisoning of dogs with tremorgenic Penicillium toxins. Medical Mycol. 2010, 48, 188–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vettorazzi, A.; López de Cerain, A. Chapter 17-Mycotoxins as Food Carcinogens. In Environmental Mycology in Public Health—Fungi and Mycotoxins Risk Assessment and Management; Viegas, C., Pinheiro, A.C., Sabino, R., Viegas, S., Brandão, J., Veríssimo, C., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2016; pp. 261–298. [Google Scholar]
- Mizutani, K.; Kumagai, S.; Mochizuki, N.; Kitagawa, Y.; Sugita-Konishi, Y. Determination of a Yellow Rice Toxin, Luteoskyrin, in Rice by Using Liquid Chromatography–Tandem Mass Spectrometry with Electrospray Ionization. J. Food Prot. 2009, 72, 1321–1326. [Google Scholar] [CrossRef]
- Srinivasan, U.; Bala, A.; Jao, S.-C.; Starke, D.W.; Jordan, T.W.; Mieyal, J.J. Selective Inactivation of Glutaredoxin by Sporidesmin and Other Epidithiopiperazinediones. Biochemistry 2006, 45, 8978–8987. [Google Scholar] [CrossRef] [Green Version]
- Bradford, H.F.; Norris, P.J.; Smith, C.C. Changes in transmitter release patterns in vitro induced by tremorgenic mycotoxins. J. Environ. Pathol. Toxicol. Oncol. 1990, 10, 17–30. [Google Scholar]
- Boysen, S.R.; Rozanski, E.A.; Chan, D.L.; Grobe, T.L.; Fallon, M.J.; Rush, J.E. Tremorgenic mycotoxicosis in four dogs from a single household. J. Am. Veter Med. Assoc. 2002, 221, 1441–1444. [Google Scholar] [CrossRef] [PubMed]
- Maragos, C.M.; Busman, M.; Plattner, R.D. Development of monoclonal antibodies for the fusarin mycotoxins. Food Addit. Contam. Part. A 2007, 25, 105–114. [Google Scholar] [CrossRef]
- Kleigrewe, K.; Aydin, F.; Hogrefe, K.; Piecuch, P.; Bergander, K.; Würthwein, E.U.; Humpf, H.U. Structure Elucidation of New Fusarins Revealing Insights in the Rearrangement Mechanisms of theFusariumMycotoxin Fusarin C. J. Agric. Food Chem. 2012, 60, 5497–5505. [Google Scholar] [CrossRef]
- Behrens, M.I.; Koh, J.; Canzoniero, L.M.T.; Sensi, S.L.; Csernansky, C.A.; Choi, D.W. 3-Nitropropionic acid induces apoptosis in cultured striatal and cortical neurons. NeuroReport 1995, 6, 545–548. [Google Scholar] [CrossRef] [PubMed]
- Patočka, J.; Bielavský, J.; Cabal, J.; Fusek, J. 3-Nitropropionic Acid and Similar Nitrotoxins. Acta Medica 2000, 43, 9–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peraica, M.; Radić, B.; Lucić, A.; Pavlović, M. Toxic effects of mycotoxins in humans. Bull. World Heal. Organ. 1999, 77, 754–766. [Google Scholar]
- Ludolph, A.; He, F.; Spencer, P.; Hammerstad, J.; Sabri, M. 3-Nitropropionic Acid—Exogenous Animal Neurotoxin and Possible Human Striatal Toxin. Can. J. Neurol. Sci. J. Can. Sci. Neurol. 1991, 18, 492–498. [Google Scholar] [CrossRef]
- Ueno, Y. Hepatotoxicity of cyclochlorotine—A cyclic peptide produced by Penicillium islandicum. Прикладная Биoхимия И Микрoбиoлoгия 1992, 28, 899–906. [Google Scholar]
- Zhou, Z.-H.; Komiyama, M.; Terao, K.; Shimada, Y. Effects of cyclochlorotine on myofibrils in cardiomyocytes and on actin filament bundles in fibroblasts in vitro. Nat. Toxins 1994, 2, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Masuda, T.; Ito, J.; Akuzawa, S.; Ishii, K.; Takagi, H.; Ueno, Y. Hepatic accumulation and hepatotoxicity of luteoskyrin in mice. Toxicol. Lett. 1992, 61, 9–20. [Google Scholar]
- Mori, S.; Sugihara, Y.; Kitagawa, A.; Kawai, K.; Nozawa, Y.; Ogihara, Y. The respiration-impairing effect of rubroskyrin, a toxic metabolite. Mycotoxin Res. 1996, 12, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Upreti, G.; Jain, M.K. Interaction of sporidesmin, a mycotoxin from Pithomyces chartarum, with lipid bilayers. Biosci. Rep. 1993, 13, 233–243. [Google Scholar] [CrossRef]
- Smith, B. Effects of low dose rates of sporidesmin given orally to sheep. New Zealand Veter J. 2000, 48, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Munday, R. Studies on the mechanism of toxicity of the mycotoxin, sporidesmin. V. Generation of hydroxyl radical by sporidesmin. J. Appl. Toxicol. 1987, 7, 17–22. [Google Scholar] [CrossRef]
- Scholl, P.F.; Groopman, J.D. Long-term Stability of Human Aflatoxin B1 Albumin Adducts Assessed by Isotope Dilution Mass Spectrometry and High-Performance Liquid Chromatography-Fluorescence. Cancer Epidemiol. Biomark. Prev. 2008, 17, 1436–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.-S.; Yu, F.-Y.; Su, C.-C.; Kan, J.-C.; Chung, C.-P.; Liu, B.-H. Activation of ERK mitogen-activated protein kinase in human cells by the mycotoxin patulin. Toxicol. Appl. Pharmacol. 2005, 207, 103–111. [Google Scholar] [CrossRef]
- Liu, B.-H.; Wu, T.-S.; Yu, F.-Y.; Wang, C.-H. Mycotoxin Patulin Activates the p38 Kinase and JNK Signaling Pathways in Human Embryonic Kidney Cells. Toxicol. Sci. 2005, 89, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Hanumegowda, U.M.; Dhulipala, V.C.; Reddy, C.S. Mechanism of secalonic acid D-induced inhibition of transcription factor binding to cyclic AMP response element in the developing murine palate. Toxicol. Sci. 2002, 70, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Moldes-Anaya, A.S.; Fonnum, F.; Eriksen, G.S.; Rundberget, T.; Walaas, S.I.; Wigestrand, M.B. In vitro neuropharmacological evaluation of penitrem-induced tremorgenic syndromes: Importance of the GABAergic system. Neurochem. Int. 2011, 59, 1074–1081. [Google Scholar] [CrossRef]
- Bruneau, J.C.; Stack, E.; O’Kennedy, R.; Loscher, C.E. Aflatoxins B1, B2 and G1 modulate cytokine secretion and cell surface marker expression in J774A.1 murine macrophages. Toxicol. Vitr. 2012, 26, 686–693. [Google Scholar] [CrossRef]
- Kiessling, K.-H. Biochemical mechanism of action of mycotoxins. Pure Appl. Chem. 1986, 58, 327–338. [Google Scholar] [CrossRef]
- Tonshin, A.A.; Teplova, V.V.; Andersson, M.A.; Salkinoja-Salonen, M.S. The Fusarium mycotoxins enniatins and beauvericin cause mitochondrial dysfunction by affecting the mitochondrial volume regulation, oxidative phosphorylation and ion homeostasis. Toxicology 2010, 276, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Zbib, N.; Repussard, C.; Tardieu, D.; Priymenko, N.; Domange, C.; Guerre, P. Toxicity of endophyte-infected ryegrass hay containing high ergovaline level in lactating ewes. J. Anim. Sci. 2015, 93, 4098–4109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.-C.; Hou, Y.-J.; Han, J.; Liu, H.-L.; Cui, X.-S.; Kim, N.-H.; Sun, S.-C. Effect of Mycotoxin-Containing Diets on Epigenetic Modifications of Mouse Oocytes by Fluorescence Microscopy Analysis. Microsc. Microanal. 2014, 20, 1158–1166. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, R.K.; Misra, R.S. Effect of aflatoxin B1 on chromatin-bound ribonucleic acid polymerase and nucleic acid and protein synthesis in germinating maize seeds. Appl. Environ. Microbiol. 1981, 42, 389–393. [Google Scholar] [CrossRef] [Green Version]
- Tashiro, F.; Hiral, K.; Ueno, Y. Inhibitory effects of carcinogenic mycotoxins on deoxyribonucleic acid-dependent ribonucleic acid polymerase and ribonuclease H. Appl. Environ. Microbiol. 1979, 38, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Al-Hammadi, S.; Marzouqi, F.; Al-Mansouri, A.; Shahin, A.; Al-Shamsi, M.; Brown, E.M.; Souid, A.-K. The Cytotoxicity of Aflatoxin B1 in Human Lymphocytes. Sultan Qaboos Univ. Med. J. 2014, 14, 65–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scallet, A.C.; Haley, R.L.; Scallet, D.M.; Duhart, H.M.; Binienda, Z.K. 3-nitropropionic acid inhibition of succinate dehydrogenase (complex II) activity in cultured Chinese hamster ovary cells: Antagonism by L-carnitine. Ann. N. Y. Acad. Sci. 2003, 993, 305–312, discussion 345–309. [Google Scholar] [CrossRef]
- Kruglov, A.G.; Andersson, M.A.; Mikkola, R.; Roivainen, M.; Kredics, L.; Saris, N.-E.L.; Salkinoja-Salonen, M.S. Novel Mycotoxin fromAcremonium exuviarumIs a Powerful Inhibitor of the Mitochondrial Respiratory Chain Complex III. Chem. Res. Toxicol. 2009, 22, 565–573. [Google Scholar] [CrossRef]
- Domijan, A.-M.; Abramov, A. Fumonisin B1 inhibits mitochondrial respiration and deregulates calcium homeostasis—Implication to mechanism of cell toxicity. Int. J. Biochem. Cell Biol. 2011, 43, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Adeyeye, S.A.O. Aflatoxigenic fungi and mycotoxins in food: A review. Crit. Rev. Food Sci. Nutr. 2019, 60, 709–721. [Google Scholar] [CrossRef]
- Afsah-Hejri, L.; Hajeb, P.; Ehsani, R.J. Application of ozone for degradation of mycotoxins in food: A review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1777–1808. [Google Scholar] [CrossRef] [PubMed]
- Asoegwu, C.R.; Chibueze, G.A.; Nelson, K.C.T.; Orji, C.G.; Nwosu, O.U.; Egbufor, U.C.; Chinaza, G.A. A Review on the Role of Biofertilizers In Reducing Soil Pollution and Increasing Soil Nutrients. Himal. J. Agric. 2020, 1, 34–38. [Google Scholar]
- Ahaotu, N.N.; Ibeabuchi, C.J.; Agunwa, I.; Echeta, C.K.; Awuchi, C.G.; Ohia, P. Antinutritional and phytochemical composition of fermented condiment (Ogiri) made from Sandbox (Hura crepitan) Seed. Eur. Acad. Res. 2020, 8, 1871–1883. [Google Scholar]
- Nnennaya, A.N.; Kate, E.C.; Evelyn, B.N.; Godswill, A.C.; Linda, A.C.; Julian, I.C.; Moses, O. Study on the nutritional and chemical composition of “Ogiri” condiment made from sandbox seed (Hura crepitans) as affected by fermentation time. GSC Biol. Pharm. Sci. 2020, 11, 105–113. [Google Scholar] [CrossRef]
- Natumanya, P.; Twinomuhwezi, H.; Igwe, V.S.; Maryam, S.; Awuchi, C.G. Effects of Drying Techniques on Nutrient Retention and Phytochemicals in Selected Vegetables. Eur. J. Agric. Food Sci. 2021, 3, 5–14. [Google Scholar] [CrossRef]
- Luo, Y.; Liu, X.; Li, J. Updating techniques on controlling mycotoxins—A review. Food Control. 2018, 89, 123–132. [Google Scholar] [CrossRef]
- Mishra, H.N.; Das, C. A Review on Biological Control and Metabolism of Aflatoxin. Crit. Rev. Food Sci. Nutr. 2003, 43, 245–264. [Google Scholar] [CrossRef]
- Wagacha, J.; Muthomi, J. Mycotoxin problem in Africa: Current status, implications to food safety and health and possible management strategies. Int. J. Food Microbiol. 2008, 124, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Munkvold, G.P. Cultural and genetic approaches to managing mycotoxins in maize. Annu. Rev. Phytopathol. 2003, 41, 99–116. [Google Scholar] [CrossRef] [PubMed]
- Adebiyi, J.A.; Kayitesi, E.; Adebo, O.A.; Changwa, R.; Njobeh, P.B. Food fermentation and mycotoxin detoxification: An African perspective. Food Control. 2019, 106, 106731. [Google Scholar] [CrossRef]
- Alberts, J.; Lilly, M.; Rheeder, J.; Burger, H.-M.; Shephard, G.S.; Gelderblom, W. Technological and community-based methods to reduce mycotoxin exposure. Food Control. 2017, 73, 101–109. [Google Scholar] [CrossRef]
- Polonelli, L.; Giovati, L.; Magliani, W.; Conti, S.; Sforza, S.; Calabretta, A.; Casoli, C.; Ronzi, P.; Grilli, E.; Gallo, A.; et al. Vaccination of Lactating Dairy Cows for the Prevention of Aflatoxin B1 Carry Over in Milk. PLoS ONE 2011, 6, e26777. [Google Scholar] [CrossRef]
- Brown, R.L.; Menkir, A.; Chen, Z.-Y.; Bhatnagar, D.; Yu, J.; Yao, H.; Cleveland, T.E. Breeding aflatoxin-resistant maize lines using recent advances in technologies—A review. Food Addit. Contam. Part. A 2013, 30, 1382–1391. [Google Scholar] [CrossRef]
- Yang, J.; Li, J.; Jiang, Y.; Duan, X.; Qu, H.; Yang, B.; Chen, F.; Sivakumar, D. Natural Occurrence, Analysis, and Prevention of Mycotoxins in Fruits and their Processed Products. Crit. Rev. Food Sci. Nutr. 2014, 54, 64–83. [Google Scholar] [CrossRef] [PubMed]
- Giovati, L.; Gallo, A.; Masoero, F.; Cerioli, C.; Ciociola, T.; Conti, S.; Magliani, W.; Polonelli, L. Vaccination of Heifers with Anaflatoxin Improves the Reduction of Aflatoxin B1 Carry Over in Milk of Lactating Dairy Cows. PLoS ONE 2014, 9, e94440. [Google Scholar] [CrossRef] [Green Version]
- Magan, N.; Aldred, D.; Mylona, K.; Lambert, R.J. Limiting mycotoxins in stored wheat. Food Addit. Contam. Part. A 2010, 27, 644–650. [Google Scholar] [CrossRef] [Green Version]
- Chulze, S. Strategies to reduce mycotoxin levels in maize during storage: A review. Food Addit. Contam. Part. A 2010, 27, 651–657. [Google Scholar] [CrossRef]
- Sarrocco, S.; Mauro, A.; Battilani, P. Use of Competitive Filamentous Fungi as anAlternative Approach for Mycotoxin Risk Reductionin Staple Cereals: State of Art and Future Perspectives. Toxins 2019, 11, 701. [Google Scholar] [CrossRef] [Green Version]
- Magan, N.; Aldred, D. Post-harvest control strategies: Minimizing mycotoxins in the food chain. Int. J. Food Microbiol. 2007, 119, 131–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexa, E.; Dehelean, C.A.; Poiana, M.-A.; Radulov, I.; Cimpean, A.-M.; Bordean, D.-M.; Tulcan, C.; Pop, G. The occurrence of mycotoxins in wheat from western Romania and histopathological impact as effect of feed intake. Chem. Central J. 2013, 7, 99. [Google Scholar] [CrossRef] [Green Version]
- Lyagin, I.; Efremenko, E. Enzymes for Detoxification of Various Mycotoxins: Origins and Mechanisms of Catalytic Action. Molecules 2019, 24, 2362. [Google Scholar] [CrossRef] [Green Version]
- Chinaza, G.A.; Chibueze, G.A. Physiological Effects of Plastic Wastes on the Endocrine System (Bisphenol A, Phthalates, Bisphenol S, PBDEs, TBBPA). Int. J. Bioinform. Comput. Biol. 2019, 4, 11–29. [Google Scholar]
- Chinaza, G.A.; Igwe, V.S.; Amagwula, I.O. Ready-to-Use Therapeutic Foods (RUTFs) for Remedying Malnutrition and Preventable Nutritional Diseases. Int. J. Adv. Acad. Res. 2020, 6, 47–81. [Google Scholar]
- Wang, L.; Wu, J.; Liu, Z.; Shi, Y.; Liu, J.; Xu, X.; Hao, S.; Mu, P.; Deng, F.; Deng, Y. Aflatoxin B1 Degradation and Detoxification by Escherichia coli CG1061 Isolated From Chicken Cecum. Front. Pharmacol. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Chinaza, G.A. Proximate Composition and Functional Properties of Different Grain Flour Composites for Industrial Applications. Int. J. Food Sci. 2019, 2, 43–64. [Google Scholar]
- Hassan, Y.; Zhou, T. Addressing the mycotoxin deoxynivalenol contamination with soil-derived bacterial and enzymatic transformations targeting the C3 carbon. World Mycotoxin J. 2018, 11, 101–112. [Google Scholar] [CrossRef]
- Xia, X.; Zhang, Y.; Li, M.; Garba, B.; Zhang, Q.; Wang, Y.; Zhang, H.; Li, P. Isolation and characterization of a Bacillus subtilis strain with aflatoxin B 1 biodegradation capability. Food Control. 2017, 75, 92–98. [Google Scholar] [CrossRef]
- Kabak, B.; Var, I. Factors affecting the removal of aflatoxin M1 from food model by Lactobacillus and Bifidobacterium strains. J. Environ. Sci. Heal. Part. B 2008, 43, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Gerbaldo, G.A.; Barberis, C.; Pascual, L.; Dalcero, A.; Barberis, L. Antifungal activity of two Lactobacillus strains with potential probiotic properties. FEMS Microbiol. Lett. 2012, 332, 27–33. [Google Scholar] [CrossRef]
- Prado, G.; Madeira, J.E.G.C.; Morais, V.A.D.; Oliveira, M.S.; Souza, R.A.; Peluzio, J.M.; Godoy, I.J.; Silva, J.F.M.; Pimenta, R.S. Reduction of Aflatoxin B1 in Stored Peanuts (Arachis hypogaea L.) Using Saccharomyces cerevisiae. J. Food Prot. 2011, 74, 1003–1006. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, M.; Wu, C.; Peng, B. Physical adsorption of patulin by Saccharomyces cerevisiae during fermentation. J. Food Sci. Technol. 2019, 56, 2326–2331. [Google Scholar] [CrossRef]
- Wu, F.; Mitchell, N.J.; Male, D.; Kensler, T.W. Reduced Foodborne Toxin Exposure Is a Benefit of Improving Dietary Diversity. Toxicol. Sci. 2014, 141, 329–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magoha, H.S.; Kimanya, M.; De Meulenaer, B.; Roberfroid, D.; Lachat, C.; Kolsteren, P. Risk of dietary exposure to aflatoxins and fumonisins in infants less than 6 months of age in Rombo, Northern Tanzania. Matern. Child. Nutr. 2016, 12, 516–527. [Google Scholar] [CrossRef]
- Cvetnić, Z.; Pepeljnjak, S. Interaction Between Certain Moulds and Aflatoxin B1 Producer Aspergillus Flavus NRRL. Arch. Ind. Hyg. Toxicol. 2007, 58, 429–434. [Google Scholar] [CrossRef] [Green Version]
- Umesha, S.; Manukumar, H.M.G.; Chandrasekhar, B.; Shivakumara, P.; Kumar, J.S.; Raghava, S.; Avinash, P.; Shirin, M.; Bharathi, T.R.; Rajini, S.B.; et al. Aflatoxins and food pathogens: Impact of biologically active aflatoxins and their control strategies. J. Sci. Food Agric. 2017, 97, 1698–1707. [Google Scholar] [CrossRef] [PubMed]
- Adebo, O.A.; Kayitesi, E.; Njobeh, P.B. Reduction of Mycotoxins during Fermentation of Whole Grain Sorghum to Whole Grain Ting (a Southern African Food). Toxins 2019, 11, 180. [Google Scholar] [CrossRef] [Green Version]
- Tilocca, B.; Balmas, V.; Hassan, Z.U.; Jaoua, S.; Migheli, Q. A proteomic investigation of Aspergillus carbonarius exposed to yeast volatilome or to its major component 2-phenylethanol reveals major shifts in fungal metabolism. Int. J. Food Microbiol. 2019, 306, 108265. [Google Scholar] [CrossRef]
- Liu, Y.; Chang, J.; Wang, P.; Yin, Q.-Q.; Huang, W.-W.; Liu, C.-Q.; Bai, X.-X.; Zhu, Q.; Gao, T.-Z.; Zhou, P. Effects of Saccharomyces cerevisiae on alleviating cytotoxicity of porcine jejunal epithelia cells induced by deoxynivalenol. AMB Express 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Mendieta, C.R.; Gómez, G.V.; Del Río, J.C.G.; Cuevas, A.C.; Arce, J.M.; Ávila, E.G. Effect of the Addition of Saccharomyces Cerevisiae Yeast Cell Walls to Diets with Mycotoxins on the Performance and Immune Responses of Broilers. J. Poult. Sci. 2018, 55, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Wang, J.; Zhang, H.; Li, C.; Zhang, X. Ochratoxin A is degraded by Yarrowia lipolytica and generates non-toxic degradation products. World Mycotoxin J. 2016, 9, 269–278. [Google Scholar] [CrossRef]
- Awuchi, C.G.; Igwe, V.S.; Echeta, C.K. The Functional Properties of Foods and Flours. Int. J. Adv. Acad. Res. 2019, 5, 139–160. [Google Scholar]
- Chilaka, C.A.; De Boevre, M.; Atanda, O.O.; De Saeger, S. The Status of Fusarium Mycotoxins in Sub-Saharan Africa: A Review of Emerging Trends and Post-Harvest Mitigation Strategies towards Food Control. Toxins 2017, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Chinaza, G.A.; Clifford, I.O.; Chika, C.O.; Victory, S.I. Evaluation of Patulin Levels and impacts on the Physical Characteristics of Grains. Int. J. Adv. Acad. Res. 2019, 5, 10–25. [Google Scholar]
- Gonçalves, A.; Gkrillas, A.; Dorne, J.L.; Dall’Asta, C.; Palumbo, R.; Lima, N.; Battilani, P.; Venâncio, A.; Giorni, P. Pre- and Postharvest Strategies to Minimize Mycotoxin Contamination in the Rice Food Chain. Compr. Rev. Food Sci. Food Saf. 2019, 18, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Neme, K.; Mohammed, A. Mycotoxin occurrence in grains and the role of postharvest management as a mitigation strategies. A review. Food Control. 2017, 78, 412–425. [Google Scholar] [CrossRef]
- Shanakhat, H.; Sorrentino, A.; Raiola, A.; Romano, A.; Masi, P.; Cavella, S. Current methods for mycotoxins analysis and innovative strategies for their reduction in cereals: An overview. J. Sci. Food Agric. 2018, 98, 4003–4013. [Google Scholar] [CrossRef]
- Karlovsky, P.; Suman, M.; Berthiller, F.; De Meester, J.; Eisenbrand, G.; Perrin, I.; Oswald, I.P.; Speijers, G.; Chiodini, A.; Recker, T.; et al. Impact of food processing and detoxification treatments on mycotoxin contamination. Mycotoxin Res. 2016, 32, 179–205. [Google Scholar] [CrossRef]
- Kalagatur, N.K.; Kamasani, J.R.; Mudili, V. Assessment of Detoxification Efficacy of Irradiation on Zearalenone Mycotoxin in Various Fruit Juices by Response Surface Methodology and Elucidation of Its in-vitro Toxicity. Front. Microbiol. 2018, 9, 2937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamle, M.; Mahato, D.K.; Devi, S.; Lee, K.E.; Kang, S.G.; Kumar, P. Fumonisins: Impact on Agriculture, Food, and Human Health and their Management Strategies. Toxins 2019, 11, 328. [Google Scholar] [CrossRef] [Green Version]
- Pereyra, M.G.; Martínez, M.; Cavaglieri, L. Presence of aiiA homologue genes encoding for N-Acyl homoserine lactone-degrading enzyme in aflatoxin B1-decontaminating Bacillus strains with potential use as feed additives. Food Chem. Toxicol. 2019, 124, 316–323. [Google Scholar] [CrossRef]
- Alizadeh, A.M.; Hashempour-Baltork, F.; Khaneghah, A.M.; Hosseini, H. New perspective approaches in controlling fungi and mycotoxins in food using emerging and green technologies. Curr. Opin. Food Sci. 2021, 39, 7–15. [Google Scholar] [CrossRef]
- Wielogorska, E.; Ahmed, Y.; Meneely, J.; Graham, W.G.; Elliott, C.T.; Gilmore, B.F. A holistic study to understand the detoxification of mycotoxins in maize and impact on its molecular integrity using cold atmospheric plasma treatment. Food Chem. 2019, 301, 125281. [Google Scholar] [CrossRef] [PubMed]
- Basaran, P.; Basaran-Akgul, N.; Oksuz, L. Elimination of Aspergillus parasiticus from nut surface with low pressure cold plasma (LPCP) treatment. Food Microbiol. 2008, 25, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Hojnik, N.; Modic, M.; Tavčar-Kalcher, G.; Babič, J.; Walsh, J.L.; Cvelbar, U. Mycotoxin Decontamination Efficacy of Atmospheric Pressure Air Plasma. Toxins 2019, 11, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awuchi, C.G.; Echeta, K.C. Current Developments in Sugar Alcohols: Chemistry, Nutrition, and Health Concerns of Sorbitol, Xylitol, Glycerol, Arabitol, Inositol, Maltitol, and Lactitol. Int. J. Adv. Acad. Res. 2019, 5, 1–33. [Google Scholar]
- Somtochukwu, I.V.; Chinenyenwa, O.G.; Godswill, A.C.; Ibrahim, K.M.; Uchenna, O.N.; Otuosorochi, A.I. Ethyl Carbamate in Burukutu Produced from Different Sorghum Varieties Under Varying Storage Conditions Using Response Surface Methodology. Am. J. Food Sci. Nutr. 2018, 5, 82–88. [Google Scholar]
- Somtochukwu, I.V.; Chinenyenwa, O.G.; Godswill, A.C.; Ibrahim, K.M.; Uchenna, O.N.; Otuosorochi, A.I. Effect of Storage Conditions on the Methanol Content of Burukutu Produced from Different Sorghum Varieties; a Response Surface Methodology Approach. Am. J. Food Nutr. Health 2018, 3, 42–47. [Google Scholar]
- Chinaza, G.A.; Ikechukwu, O.A.; Victory, S.I.; Allan, I.G. Effects of Repeated Deep Frying on Refractive Index and Peroxide Value of Selected Vegetable Oils. Int. J. Adv. Acad. Res. 2018, 4, 106–119. [Google Scholar]
- Awuchi, C.G.; Nwankwere, E.T. Residual Calcium Content of Sweet Potato Slices after Osmotic Pre-treatment with Salt (NaCl) Solution. Am. J. Food Nutr. Health 2018, 3, 8–15. [Google Scholar]
- Awuchi, C.G. Sugar Alcohols: Chemistry, Production, Health Concerns and Nutritional Importance of Mannitol, Sorbitol, Xylitol, and Erythritol. Int. J. Adv. Acad. Res. 2017, 3, 31–66. [Google Scholar]
- Udeogu, E.; Awuchi, C.G. Effect of Processing on Hemagglutinin Activity of Lectin Extracts from Selected Cereals and Legumes. Int. J. Adv. Acad. Res. 2016, 2, 24–59. [Google Scholar]
- Rushing, B.R.; Selim, M.I. Aflatoxin B1: A review on metabolism, toxicity, occurrence in food, occupational exposure, and detoxification methods. Food Chem. Toxicol. 2019, 124, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Kamala, A.; Kimanya, M.; Haesaert, G.; Tiisekwa, B.; Madege, R.; Degraeve, S.; Cyprian, C.; De Meulenaer, B. Local post-harvest practices associated with aflatoxin and fumonisin contamination of maize in three agro ecological zones of Tanzania. Food Addit. Contam. Part. A 2016, 33, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Francesconi, S.; Steiner, B.; Buerstmayr, H.; Lemmens, M.; Sulyok, M.; Balestra, G.M. Chitosan Hydrochloride Decreases Fusarium graminearum Growth and Virulence and Boosts Growth, Development and Systemic Acquired Resistance in Two Durum Wheat Genotypes. Molecules 2020, 25, 4752. [Google Scholar] [CrossRef]
- Gunupuru, L.R.; Patel, J.S.; Sumarah, M.W.; Renaud, J.B.; Mantin, E.G.; Prithiviraj, B. A plant biostimulant made from the marine brown algae Ascophyllum nodosum and chitosan reduceFusarium head blight and mycotoxin contamination in wheat. PLoS ONE 2019, 14, e0220562. [Google Scholar] [CrossRef] [PubMed]
- Solís-Cruz, B.; Hernández-Patlán, D.; Beyssac, E.; Latorre, J.D.; Hernandez-Velasco, X.; Merino-Guzman, R.; Tellez, G.; López-Arellano, R. Evaluation of Chitosan and Cellulosic Polymers as Binding Adsorbent Materials to Prevent Aflatoxin B1, Fumonisin B1, Ochratoxin, Trichothecene, Deoxynivalenol, and Zearalenone Mycotoxicoses Through an In Vitro Gastrointestinal Model for Poultry. Polymers 2017, 9, 529. [Google Scholar] [CrossRef] [PubMed]
- Pirouz, A.A.; Selamat, J.; Iqbal, S.Z.; Samsudin, N.I.P. Efficient and Simultaneous Chitosan-Mediated Removal of 11 Mycotoxins from Palm Kernel Cake. Toxins 2020, 12, 115. [Google Scholar] [CrossRef] [Green Version]
- Zachetti, V.G.L.; Cendoya, E.; Nichea, M.J.; Chulze, S.N.; Ramirez, M.L. Preliminary Study on the Use of Chitosan as an Eco-Friendly Alternative to Control Fusarium Growth and Mycotoxin Production on Maize and Wheat. Pathogens 2019, 8, 29. [Google Scholar] [CrossRef] [Green Version]
- Porto, Y.D.; Trombete, F.M.; Freitas-Silva, O.; De Castro, I.M.; Direito, G.M.; Ascheri, J.L.R. Gaseous Ozonation to Reduce Aflatoxins Levels and Microbial Contamination in Corn Grits. Microorganisms 2019, 7, 220. [Google Scholar] [CrossRef] [Green Version]
- Piemontese, L.; Messia, M.C.; Marconi, E.; Falasca, L.; Zivoli, R.; Gambacorta, L.; Perrone, G.; Solfrizzo, M. Effect of gaseous ozone treatments on DON, microbial contaminants and technological parameters of wheat and semolina. Food Addit. Contam. Part. A 2018, 35, 761–772. [Google Scholar] [CrossRef]
- Li, M.; Guan, E.; Bian, K. Effect of ozone treatment on deoxynivalenol and quality evaluation of ozonised wheat. Food Addit. Contam. Part. A 2014, 32, 544–553. [Google Scholar] [CrossRef]
- Okpala, C.O.R. Ozone delivery on food materials incorporating some bio-based processes: A succinct synopsis. Adv. Mater. Proc. 2017, 2, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Okpala, C.O.R. Changes in some biochemical and microbiological properties of ozone-processed shrimp: Effects of increased ozone discharge combined with iced storage. J. Food Nutr. Res. 2018, 57, 48–56. [Google Scholar]
- Pandiselvam, R.; Sunoj, S.; Manikantan, M.R.; Kothakota, A.; Hebbar, K.B. Application and Kinetics of Ozone in Food Preservation. Ozone Sci. Eng. 2016, 39, 115–126. [Google Scholar] [CrossRef]
- Li, M.; Guan, E.; Bian, K. Structure Elucidation and Toxicity Analysis of the Degradation Products of Deoxynivalenol by Gaseous Ozone. Toxins 2019, 11, 474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agriopoulou, S.; Koliadima, A.; Karaiskakis, G.; Kapolos, J. Kinetic study of aflatoxins’ degradation in the presence of ozone. Food Control. 2016, 61, 221–226. [Google Scholar] [CrossRef]
- Luo, X.; Wang, R.; Wang, L.; Li, Y.; Wang, Y.; Chen, Z. Detoxification of aflatoxin in corn flour by ozone. J. Sci. Food Agric. 2014, 94, 2253–2258. [Google Scholar] [CrossRef]
- Jalili, M. A review on aflatoxins reduction in food. Iran. J. Health Saf. Environ. 2016, 3, 445–459. [Google Scholar]
- Wang, L.; Luo, Y.; Luo, X.; Wang, R.; Li, Y.; Li, Y.; Shao, H.; Chen, Z. Effect of deoxynivalenol detoxification by ozone treatment in wheat grains. Food Control. 2016, 66, 137–144. [Google Scholar] [CrossRef]
- Brodowska, A.J.; Nowak, A.; Śmigielski, K. Ozone in the food industry: Principles of ozone treatment, mechanisms of action, and applications: An overview. Crit. Rev. Food Sci. Nutr. 2018, 58, 2176–2201. [Google Scholar] [CrossRef]
- Öztekin, S.; Zorlugenç, B.; Zorlugenç, F.K. Effects of ozone treatment on microflora of dried figs. J. Food Eng. 2006, 75, 396–399. [Google Scholar] [CrossRef]
- Palou, L.; Smilanick, J.L.; Crisosto, C.H.; Mansour, M. Effect of Gaseous Ozone Exposure on the Development of Green and Blue Molds on Cold Stored Citrus Fruit. Plant. Dis. 2001, 85, 632–638. [Google Scholar] [CrossRef]
- Wu, J.; Doan, H.; Cuenca, M.A. Investigation of gaseous ozone as an anti-fungal fumigant for stored wheat. J. Chem. Technol. Biotechnol. 2006, 81, 1288–1293. [Google Scholar] [CrossRef]
- Young, J.C.; Zhu, H.; Zhou, T. Degradation of trichothecene mycotoxins by aqueous ozone. Food Chem. Toxicol. 2006, 44, 417–424. [Google Scholar] [CrossRef]
- Inan, F.; Pala, M.; Doymaz, I. Use of ozone in detoxification of aflatoxin B1 in red pepper. J. Stored Prod. Res. 2007, 43, 425–429. [Google Scholar] [CrossRef]
- Zorlugenç, B.; Zorlugenç, F.K.; Öztekin, S.; Evliya, I.B. The influence of gaseous ozone and ozonated water on microbial flora and degradation of aflatoxin B1 in dried figs. Food Chem. Toxicol. 2008, 46, 3593–3597. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Xie, W. Detoxification of Aflatoxin B1 by magnetic graphene composite adsorbents from contaminated oils. J. Hazard. Mater. 2020, 381, 120915. [Google Scholar] [CrossRef]
- Bankole, S.A. Effect of ethylene oxide and methyl formate fumigation on seeds mycoflora and germination of some stored oil seeds in Nigeria. Crop. Res. 1996, 11, 224–227. [Google Scholar]
- Kavita, W.; Reddy, M.U. Effect of chemicals on aflatoxin B1production, germination and viability in maize and groundnuts. J. Res. Angrau 2000, 28, 57–64. [Google Scholar]
- Tarazona, A.; Gómez, J.V.; Mateo, E.M.; Jiménez, M.; Mateo, F. Antifungal effect of engineered silver nanoparticles on phytopathogenic and toxigenic Fusarium spp. and their impact on mycotoxin accumulation. Int. J. Food Microbiol. 2019, 306, 108259. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wu, S.; Wang, F.; Li, Q.; He, C.; Duan, N.; Wang, Z. Assessing the toxicity in vitro of degradation products from deoxynivalenol photocatalytic degradation by using upconversion nanoparticles@ TiO2 composite. Chemosphere 2020, 238, 124648. [Google Scholar] [CrossRef] [PubMed]
- González-Jartín, J.M.; Alves, L.D.C.; Alfonso, A.; Piñeiro, Y.; Vilar, S.Y.; Gomez, M.G.; Osorio, Z.V.; Sainz, M.J.; Vieytes, M.R.; Rivas, J.; et al. Detoxification agents based on magnetic nanostructured particles as a novel strategy for mycotoxin mitigation in food. Food Chem. 2019, 294, 60–66. [Google Scholar] [CrossRef]
- Chaudhari, A.K.; Dwivedy, A.K.; Singh, V.K.; Das, S.; Singh, A.; Dubey, N.K. Essential oils and their bioactive compounds as green preservatives against fungal and mycotoxin contamination of food commodities with special reference to their nanoencapsulation. Environ. Sci. Pollut. Res. 2019, 26, 25414–25431. [Google Scholar] [CrossRef] [PubMed]
- Perczak, A.; Juś, K.; Gwiazdowska, D.; Marchwińska, K.; Waśkiewicz, A. The Efficiency of Deoxynivalenol Degradation by Essential Oils under In Vitro Conditions. Foods 2019, 8, 403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aiko, V.; Mehta, A. Occurrence, detection and detoxification of mycotoxins. J. Biosci. 2015, 40, 943–954. [Google Scholar] [CrossRef]
- Sánchez-Montero, L.; Córdoba, J.J.; Alía, A.; Peromingo, B.; Núñez, F. Effect of Spanish smoked paprika “Pimentón de La Vera” on control of ochratoxin A and aflatoxins production on a dry-cured meat model system. Int. J. Food Microbiol. 2019, 308, 108303. [Google Scholar] [CrossRef]
- Kollia, E.; Proestos, C.; Zoumpoulakis, P.; Markaki, P. Capsaicin, an inhibitor of Ochratoxin A production by Aspergillus section Nigri strains in grapes (Vitis vinifera L.). Food Addit. Contam. Part. A 2019, 36, 1709–1721. [Google Scholar] [CrossRef] [PubMed]
- Kalagatur, N.K.; Kamasani, J.R.; Mudili, V.; Krishna, K.; Chauhan, O.P.; Sreepathi, M.H. Effect of high pressure processing on growth and mycotoxin production of Fusarium graminearum in maize. Food Biosci. 2018, 21, 53–59. [Google Scholar] [CrossRef]
- Timmermans, R.; Hayrapetyan, H.; Vollebregt, M.; Dijksterhuis, J. Comparing thermal inactivation to a combined process of moderate heat and high pressure: Effect on ascospores in strawberry puree. Int. J. Food Microbiol. 2020, 325, 108629. [Google Scholar] [CrossRef] [PubMed]
- Suchanek, M.; Olejniczak, Z. Low field MRI study of the potato cell membrane electroporation by pulsed electric field. J. Food Eng. 2018, 231, 54–60. [Google Scholar] [CrossRef]
- Vijayalakshmi, S.; Nadanasabhapathi, S.; Kumar, R.; Kumar, S.S. Effect of pH and pulsed electric field process parameters on the aflatoxin reduction in model system using response surface methodology. J. Food Sci. Technol. 2018, 55, 868–878. [Google Scholar] [CrossRef]
- Zhong, C.; Guan, X.; Fan, Z.; Song, W.; Chen, R.; Wang, Y.; Sun, X.; He, S. Pulsed electric field disinfection treatment of Fusarium oxysporum in nutrient solution. Water Supply 2019, 19, 2116–2122. [Google Scholar] [CrossRef]
- Rudik, F.Y.; Morgunova, N.L.; Krasnikova, E.S. Decontamination of grain by ultrasound. IOP Conf. Ser. Earth Environ. Sci. IOP Conf. Ser. Earth Environ. Sci. 2020, 421, 022022. [Google Scholar] [CrossRef]
- Del Villalobos, M.; Serradilla, M.J.; Martín, A.; Ruíz-Moyano, S.; Casquete, R.; Hernández, A.; Córdoba, M.G. Use of efficient drying methods to improve the safety and quality of dried fig. J. Food Process. Preserv. 2018, 43, e13853. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Awuchi, C.G.; Ondari, E.N.; Ogbonna, C.U.; Upadhyay, A.K.; Baran, K.; Okpala, C.O.R.; Korzeniowska, M.; Guiné, R.P.F. Mycotoxins Affecting Animals, Foods, Humans, and Plants: Types, Occurrence, Toxicities, Action Mechanisms, Prevention, and Detoxification Strategies—A Revisit. Foods 2021, 10, 1279. https://doi.org/10.3390/foods10061279
Awuchi CG, Ondari EN, Ogbonna CU, Upadhyay AK, Baran K, Okpala COR, Korzeniowska M, Guiné RPF. Mycotoxins Affecting Animals, Foods, Humans, and Plants: Types, Occurrence, Toxicities, Action Mechanisms, Prevention, and Detoxification Strategies—A Revisit. Foods. 2021; 10(6):1279. https://doi.org/10.3390/foods10061279
Chicago/Turabian StyleAwuchi, Chinaza Godswill, Erick Nyakundi Ondari, Chukwuka U. Ogbonna, Anjani K. Upadhyay, Katarzyna Baran, Charles Odilichukwu R. Okpala, Małgorzata Korzeniowska, and Raquel P. F. Guiné. 2021. "Mycotoxins Affecting Animals, Foods, Humans, and Plants: Types, Occurrence, Toxicities, Action Mechanisms, Prevention, and Detoxification Strategies—A Revisit" Foods 10, no. 6: 1279. https://doi.org/10.3390/foods10061279