Photobiomodulation Using Different Infrared Light Sources Promotes Muscle Precursor Cells Migration and Proliferation


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
Experimental Groups
2.2. Irradiation Procedure
2.2.1. PBM Using LLL
2.2.2. PBM Using LED
2.3. May–Grunwald and Giemsa Staining for Quantifying Cell Proliferation
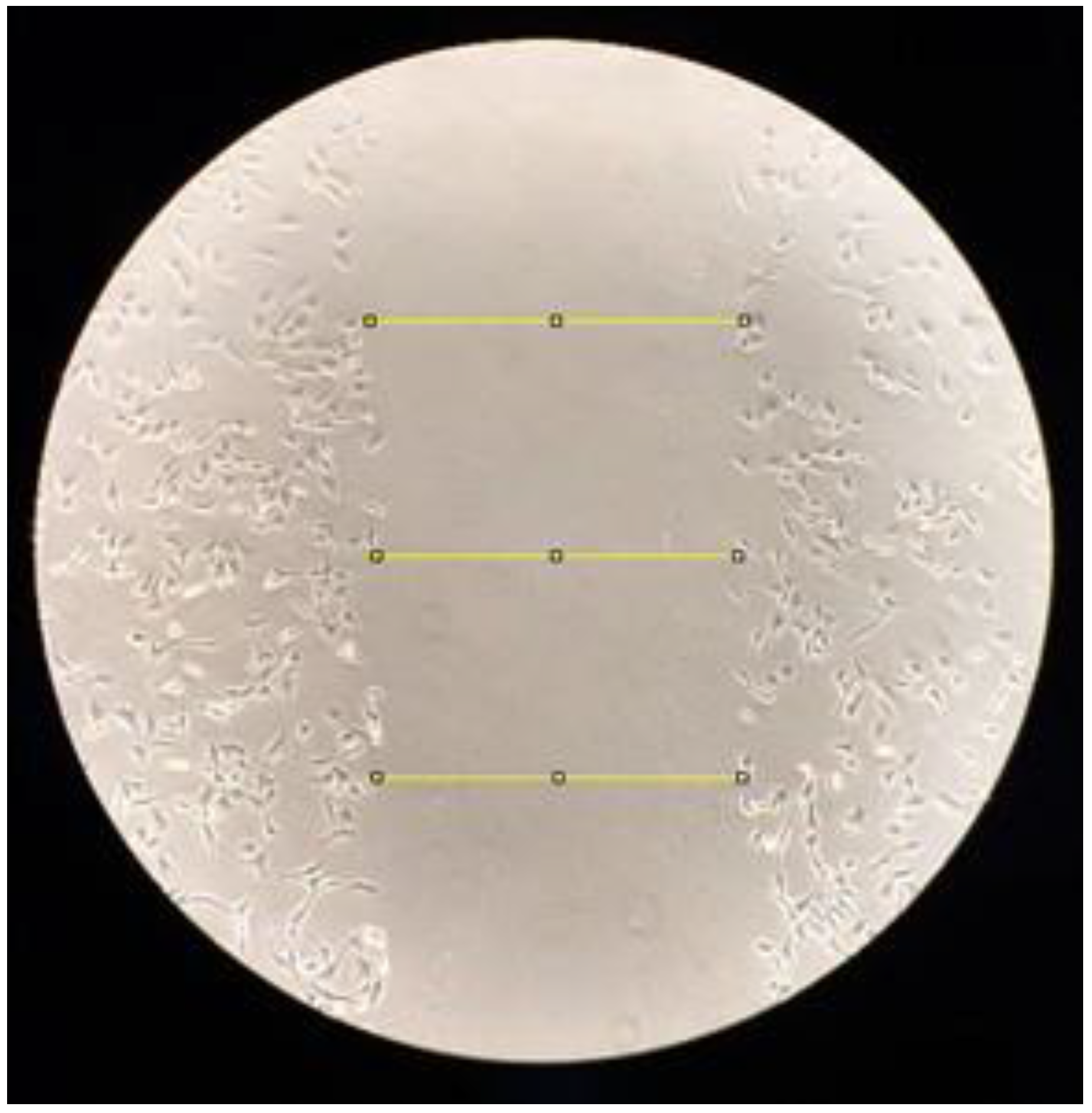
2.4. Cell Migration
2.5. Statistical Analysis
3. Results
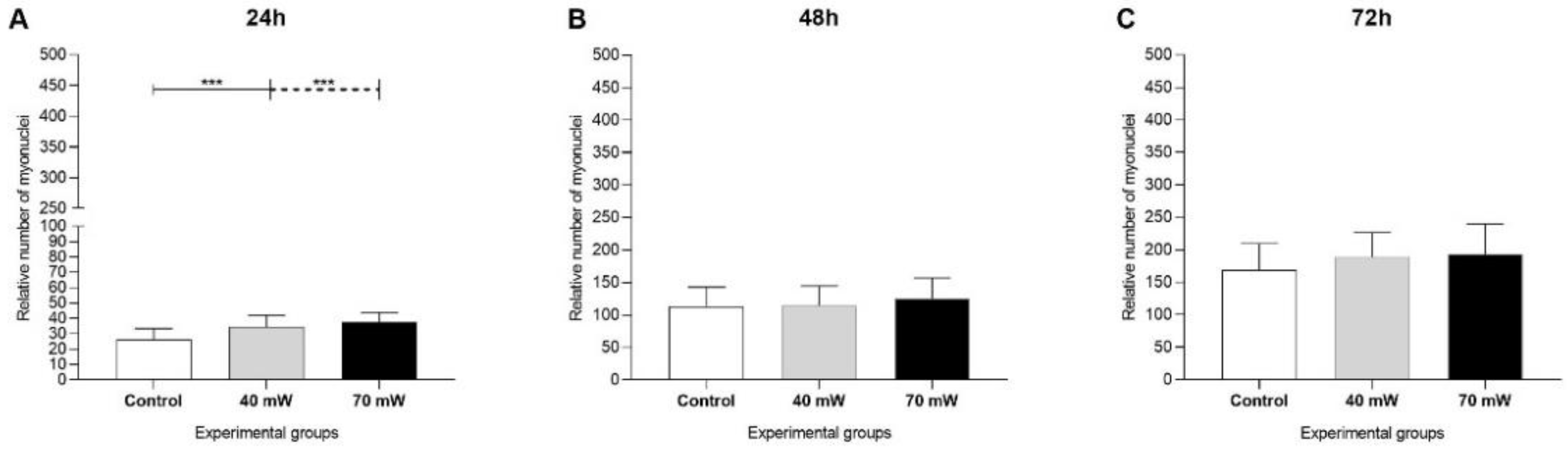
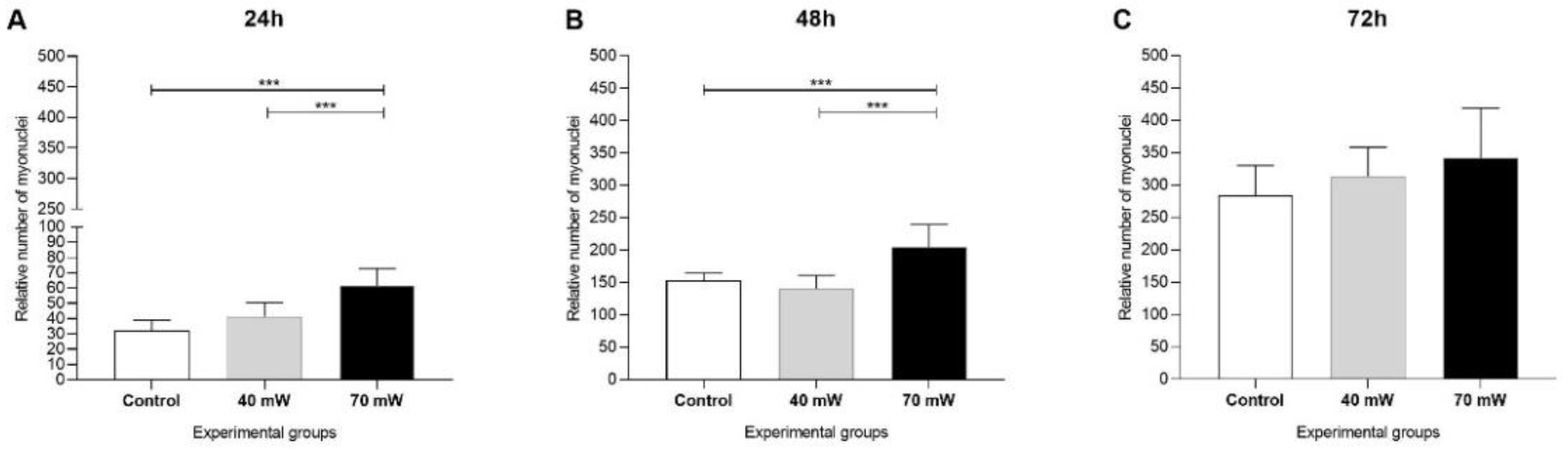
3.1. Cell Proliferation
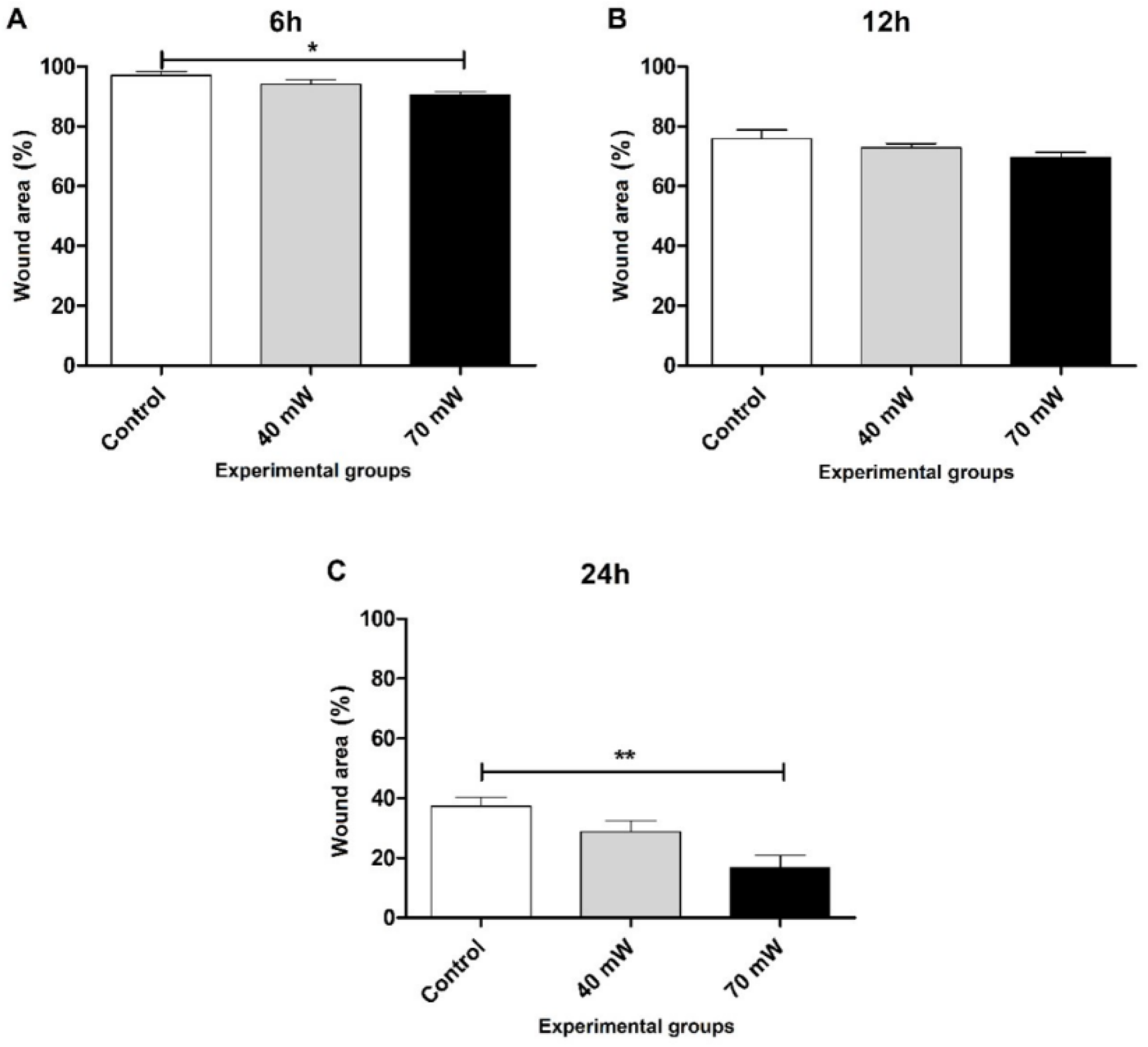
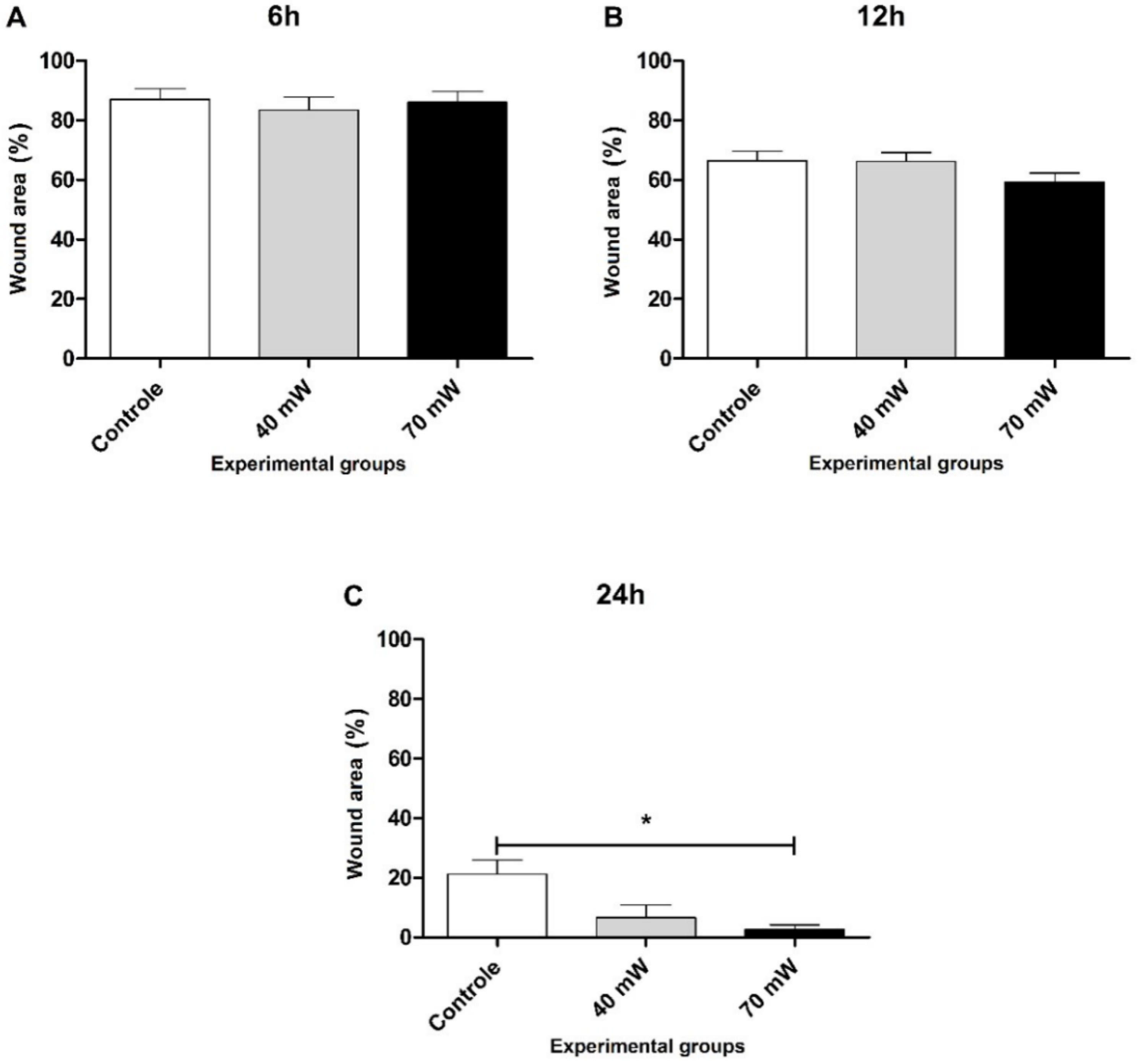
3.2. Cell Migration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, W.; Kim, J.; Park, H.S.; Jeon, J.S. Development of Microfluidic Stretch System for Studying Recovery of Damaged Skeletal Muscle Cells. Micromachines 2018, 9, 671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uezumi, A.; Ikemoto-Uezumi, M.; Tsuchida, K. Roles of Nonmyogenic Mesenchymal Progenitors in Pathogenesis and Regeneration of Skeletal Muscle. Front. Physiol. 2014, 5, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filippin, L.I.; Moreira, A.J.; Marroni, N.P.; Xavier, R.M. Nitric Oxide and Repair of Skeletal Muscle Injury. Nitric Oxide 2009, 21, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Tidball, J.G. Regulation of Muscle Growth and Regeneration by the Immune System. Nat. Rev. Immunol. 2017, 17, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Arnold, L.; Henry, A.; Poron, F.; Baba-Amer, Y.; van Rooijen, N.; Gherardi, A.P.R.K.; Chazaud, B. Inflammatory Monocytes Recruited after Skeletal Muscle Injury Switch into Anti-inflammatory Macrophages to Support Myogenesis. J. Exp. Med. 2007, 204, 1057–1069. [Google Scholar] [CrossRef] [Green Version]
- Ranzato, E.; Balbo, V.; Boccafoschi, F.; Mazzucco, L.; Burlando, B. Scratch Wound Closure of C2C12 Mouse Myoblasts Is Enhanced by Human Platelet Lysate. Cell. Biol. Int. 2009, 33, 911–917. [Google Scholar] [CrossRef]
- da Silva Neto Trajano, L.A.; Stumbo, A.C.; da Silva, C.L.; Mencalha, A.L.; Fonseca, A.S. Low-Level Infrared Laser Modulates Muscle Repair and Chromosome Stabilization Genes in Myoblasts. Lasers. Med. Sci. 2016, 31, 1161–1167. [Google Scholar] [CrossRef]
- Ferreira, J.H.; Cury, S.S.; Vechetti-júnior, I.J.; Fernandez, G.J.; Moraes, L.N.; Alves, C.A.B.; Freire, P.P.; Freitas, C.E.A.; Dal-pai-silva, M.; Carvalho, R.F. Low-Level Laser Irradiation Induces a Transcriptional Myotube-like Profile in C2C12 Myoblasts. Lasers Med. Sci. 2018, 8, 1673–1683. [Google Scholar] [CrossRef] [Green Version]
- Kitakaze, T.; Oshimo, M.; Kobayashi, Y.; Ryu, M.; Suzuki, Y.A.; Inui, H.; Harada, N.; Yamaji, R. Lactoferrin Promotes Murine C2C12 Myoblast Proliferation and Differentiation and Myotube Hypertrophy. Mol. Med. Rep. 2018, 17, 5912–5920. [Google Scholar] [CrossRef] [Green Version]
- da Silva Neto Trajano, L.A.; da Silva, C.L.; de Carvalho, S.N.; Cortez, E.; Mencalha, A.L.; de Souza da Fonseca, A.; Stumbo, A.C. Cell Viability, Reactive Oxygen Species, Apoptosis, and Necrosis in Myoblast Cultures Exposed to Low-Level Infrared Laser. Lasers Med. Sci. 2016, 31, 841–848. [Google Scholar] [CrossRef]
- Santos, T.C.; de Brito Sousa, K.; Andreo, L.; Martinelli, A.; Rodrigues, M.F.S.D.; Bussadori, S.K.; Fernandes, K.P.S.; Mesquita-Ferrari, R.A. Effect of Photobiomodulation on C2C12 Myoblasts Cultivated in M1 Macrophage-Conditioned Media. Photoch. Photob. 2020, 4, 906–916. [Google Scholar] [CrossRef] [PubMed]
- Bolus, D.J.; Shanmugam, G.; Narasimhan, M.; Rajasekaran, N.S. Recurrent Heat Shock Impairs the Proliferation and Differentiation of C2C12 Myoblasts. Cell Stress Chaper. 2018, 23, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Andreo, L.; Mesquita-ferrari, R.A.; Ribeiro, Ã.B.G.; Benitte, A. Effects of Myogenic Precursor Cells (C2C12) Transplantation and Low-Level Laser Therapy on Muscle Repair. Lasers Surg. Med. 2018, 50, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.M.D.; Malamo, A.G.; Larkin-Kaiser, K.A.; Borsa, P.A.; Adhihetty, P.J. Effect of Near-Infrared Light Exposure on Mitochondrial Signaling in C2C12 Muscle Cells. Mitoch. Resear. Soc. 2014, 14, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Serrage, H.J.; Joanisse, S.; Cooper, P.R.; Palin, W.; Hadis, M.; Darch, O.; Philp, A.; Milward, M.R. Differential Responses of Myoblasts and Myotubes to Photobiomodulation Are Associated with Mitochondrial Number. J. Biophot. 2019, 12, e201800411. [Google Scholar] [CrossRef] [Green Version]
- Alves, A.N.; Ribeiro, B.G.; Fernandes, K.P.S.; Souza, N.H.C.; Rocha, L.A.; Nunes, F.D.; Bussadori, S.K.; Mesquita-Ferrari, R.A. Comparative Effects of Low-Level Laser Therapy Pre- and Post-Injury on MRNA Expression of MyoD, Myogenin, and IL-6 during the Skeletal Muscle Repair. Lasers Med. Sci. 2016, 31, 679–685. [Google Scholar] [CrossRef]
- Souza, N.H.C.; Mesquita-Ferrari, R.A.; Rodrigues, M.F.S.D.; da Silva, D.F.T.; Ribeiro, B.G.; Alves, A.N.; Garcia, M.P.; Nunes, F.D.; da Silva Junior, E.M.; França, C.M.; et al. Photobiomodulation and Different Macrophages Phenotypes during Muscle Tissue Repair. J. Cell. Mol. Med. 2018, 22, 4922–4934. [Google Scholar] [CrossRef]
- Ferraresi, C.; Kaippert, B.; Avci, P.; Huang, Y.; Victor, M.; De Sousa, P.; Bagnato, V.S.; Parizotto, N.A. Low-Level Laser (Light) Therapy Increases Mitochondrial Membrane Potential and ATP Synthesis in C2C12 Myotubes with a Peak Response at 3–6 Hours. Photochem. Photobiol. 2016, 91, 411–416. [Google Scholar] [CrossRef]
- Mesquita-Ferrari, R.A.; Alves, A.N.; de Oliveira Cardoso, V.; Artilheiro, P.P.; Bussadori, S.K.; Rocha, L.A.; Nunes, F.D.; Fernandes, K.P.S. Low-Level Laser Irradiation Modulates Cell Viability and Creatine Kinase Activity in C2C12 Muscle Cells during the Differentiation Process. Lasers Med. Sci. 2015, 30, 2209–2213. [Google Scholar] [CrossRef]
- Silveira, P.C.L.; Ferreira, G.K.; Zaccaron, R.P.; Glaser, V.; Remor, A.P.; Mendes, C.; Pinho, R.A.; Latini, A. Effects of Photobiomodulation on Mitochondria of Brain, Muscle, and C6 Astroglioma Cells. Med. Eng. Phys. 2019, 71, 108–113. [Google Scholar] [CrossRef]
- Silva, D.F.T.; Mesquita-Ferrari, R.A.; Fernandes, K.P.S.; Raele, M.P.; Wetter, N.U.; Deana, A.M. Effective Transmission of Light for Media Culture, Plates and Tubes. Photoch. Photob. 2012, 88, 1211–1216. [Google Scholar] [CrossRef] [PubMed]
- Ono, Y.; Sakamoto, K. Lipopolysaccharide Inhibits Myogenic Differentiation of C2C12 Myoblasts through the Toll-like Receptor 4-Nuclear Factor-ΚB Signaling Pathway and Myoblast-Derived Tumor Necrosis Factor-α. PLoS ONE 2017, 12, e0182040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veliça, P.; Bunce, C.M. A Quick, Simple and Unbiased Method to Quantify C2C12 Myogenic Differentiation. Muscle Nerve 2011, 44, 366–370. [Google Scholar] [CrossRef] [PubMed]
- Pansters, N.A.M.; van der Velden, J.L.J.; Kelders, M.C.J.M.; Laeremans, H.; Schols, A.M.W.J.; Langen, R.C.J. Segregation of Myoblast Fusion and Muscle-Specific Gene Expression by Distinct Ligand-Dependent Inactivation of GSK-3b. Cell. Mol. Life Sci. 2011, 68, 523–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizunoya, W.; Tashima, A.; Sato, Y.; Tatsumi, R.; Ikeuchi, Y. The Growth-Promoting Activity of Egg White Proteins in the C2C12 Myoblast Cell Line. Anim. Sci. J. 2015, 86, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Hamblin, M.R. Mechanisms and Applications of the Anti-Inflammatory Effects of Photobiomodulation. AIMS Bioph. 2017, 4, 337–361. [Google Scholar] [CrossRef] [PubMed]
- Baskov, A.; Borshchenko, I.A.; Baskov, V.; Shekhter, A.; Sobol, E. Laser Reconstruction of Spinal Discs Experiments and Clinic. Appl. Sci. 2022, 12, 675. [Google Scholar] [CrossRef]
- Zein, R.; Selting, W.; Hamblin, M.R. Review of Light Parameters and Photobiomodulation Efficacy: Dive into Complexity. J. Biomed. Opt. 2018, 23, 120901. [Google Scholar] [CrossRef] [Green Version]
- Bajaj, P.; Reddy, B.; Millet, L.; Wei, C.; Zorlutuna, P.; Bao, G.; Bashir, R. Patterning the Differentiation of C2C12 Skeletal Myoblasts. Integr. Biol. 2011, 3, 897–909. [Google Scholar] [CrossRef]
- Ferreira, M.P.P.; Ferrari, R.A.M.; Gravalos, E.D.; Martins, M.D.; Bussadori, S.K.; Gonzalez, D.A.B.; Fernandes, K.P.S. Effect of Low-Energy Gallium-Aluminum-Arsenide and Aluminium Gallium Indium Phosphide Laser Irradiation on the Viability of C2C12 Myoblasts in a Muscle Injury Model. Photomed. Laser Surg. 2009, 27, 901–906. [Google Scholar] [CrossRef]
- Mesquita-Ferrari, R.A.; Ribeiro, R.; Souza, N.H.C.; Silva, C.A.A.; Martins, M.D.; Bussadori, S.K.; Fernandes, K.P.S. No Effect of Low-Level Lasers on in Vitro Myoblast Culture. Indian J. Exp. Biol. 2011, 49, 423–428. [Google Scholar] [PubMed]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Freitas, L.F.; Hamblin, M.R. Proposed Mechanisms of Photobiomodulation or Low-Level Light Therapy. IEEE J. Sel. Top. Quantum Electron. 2016, 22, 348–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naderi, M.S.; Razzaghi, M.; Djavid, G.E.; Hajebrahimi, Z. A Comparative Study of 660 nm Low-Level Laser and Light Emitted Diode in Proliferative Effects of Fibroblast Cells. Lasers Med. Sci. 2017, 8, S46–S50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heiskanen, V.; Hamblin, M.R. Photobiomodulation: Lasers vs. light emitting diodes? Photoch. Photob. 2018, 17, 1003–1017. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | LLL | LLL |
|---|---|---|
| Wavelenght (nm) | 780 | 780 |
| Operating mode | CW | CW |
| Area irradiated (cm2) | 0.04 | 0.04 |
| Output power (mW) | 40 | 70 |
| Irradiance (W/cm2) | 1 | 1.75 |
| Radiant exposure (J/cm2) | 10 | 10 |
| Number of points irradiated | 1 | 1 |
| Total exposure time (s) | 10 | 6 |
| Total energy (J) | 0.4 | 0.4 |
| Parameter | LED | LED |
|---|---|---|
| Wavelenght (nm) | 850 | 850 |
| Operating mode | CW | CW |
| Area irradiated (cm2) | 3 | 3 |
| Output power (mW) | 40 | 70 |
| Irradiance (W/cm2) | 0.013 | 0.023 |
| Radiant exposure (J/cm2) | 0.13 | 0.13 |
| Number of points irradiated | 1 | 1 |
| Total exposure time (s) | 10 | 6 |
| Total energy (J) | 0.4 | 0.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lovisetto, R.; Malavazzi, T.C.d.S.; Andreo, L.; Rodrigues, M.F.S.D.; Bussadori, S.K.; Fernandes, K.P.S.; Mesquita-Ferrari, R.A. Photobiomodulation Using Different Infrared Light Sources Promotes Muscle Precursor Cells Migration and Proliferation. Photonics 2022, 9, 469. https://doi.org/10.3390/photonics9070469
Lovisetto R, Malavazzi TCdS, Andreo L, Rodrigues MFSD, Bussadori SK, Fernandes KPS, Mesquita-Ferrari RA. Photobiomodulation Using Different Infrared Light Sources Promotes Muscle Precursor Cells Migration and Proliferation. Photonics. 2022; 9(7):469. https://doi.org/10.3390/photonics9070469
Chicago/Turabian StyleLovisetto, Renan, Tainá Caroline dos Santos Malavazzi, Lucas Andreo, Maria Fernanda Setubal Destro Rodrigues, Sandra Kalil Bussadori, Kristianne Porta Santos Fernandes, and Raquel Agnelli Mesquita-Ferrari. 2022. "Photobiomodulation Using Different Infrared Light Sources Promotes Muscle Precursor Cells Migration and Proliferation" Photonics 9, no. 7: 469. https://doi.org/10.3390/photonics9070469